
Management of Rhizosphere Microbiota and Plant Production under Drought Stress: A Comprehensive Review

 , ,
, ,  ,
,  , and
, and 
Abstract
1. Introduction
2. The Drought as Main Constraint for Plant Production in a Global Climate Change Scenario
3. Rhizosphere Microbial Groups and Ecological Roles for Plant Production under Drought Conditions

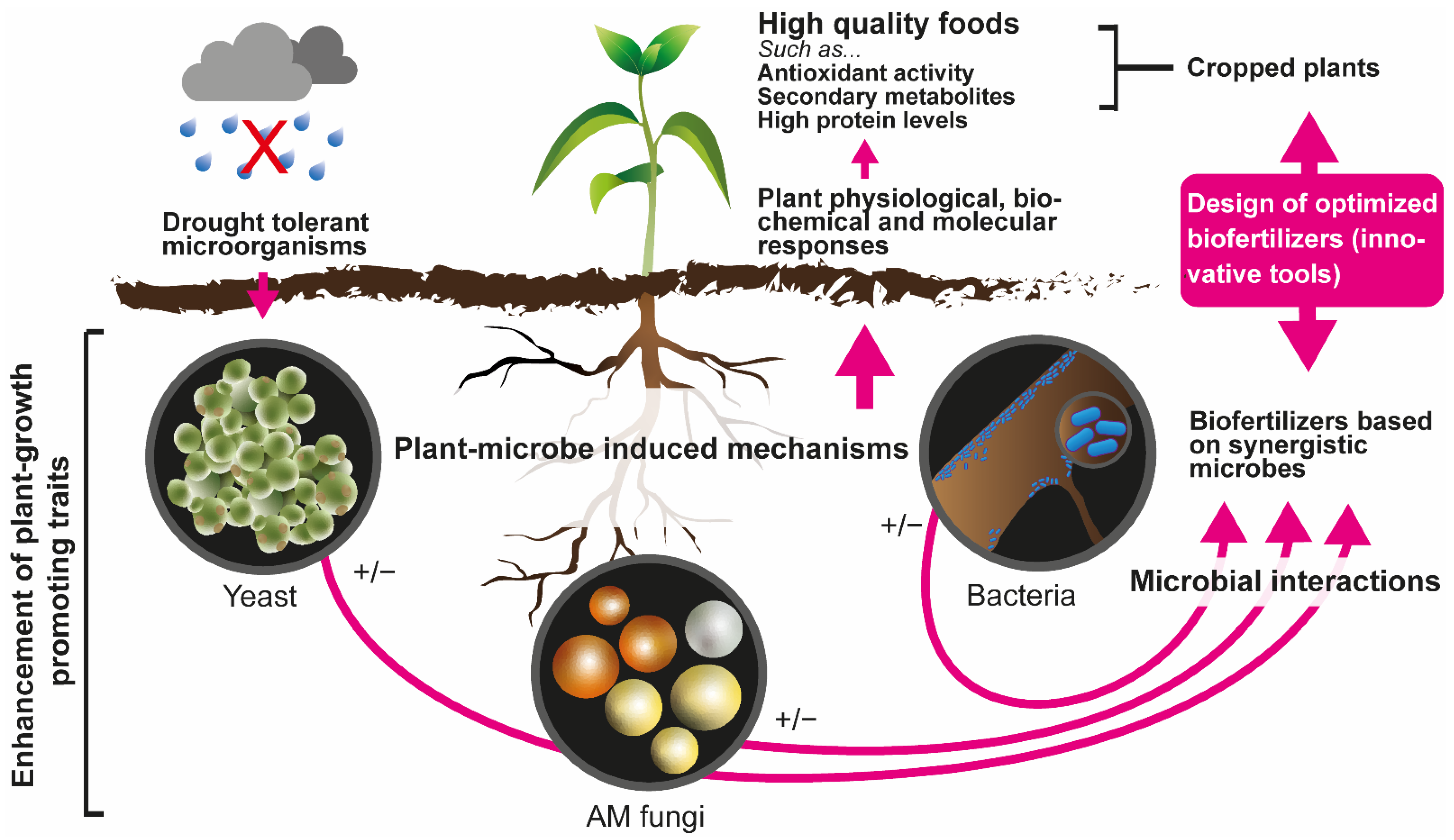{kind=link}
| Crop | Microorganism | PGP Traits Evidenced | Specific Effects | References |
|---|---|---|---|---|
| Sorghum bicolor L. | Streptomyces laurentii and Penicillium sp. | P and Zn-solubilization. Siderophores, hydrogen cyanide, NH3 and IAA production | + Plant growth + Chlorophyll content + Production of osmolytes − Lipid peroxidation | [89] |
| Poncirus trifoliata | Ochrobacetrum sp., Microbacterium sp., Enterobacter sp., and Enterobacter cloacae | N-fixation, P-solubilization, ACC deaminase activity, siderophore and IAA production | + Proline accumulation in leaves + Relative water content + Cell membrane stability index + Genes like sbP5CS2 and sbP5CS1 | [59] |
| Zea mays L. | Arthrobacter arilaitensis and Streptomyces pseudovenezuelae | P-solubilization, ACC deaminase activity, IAA, siderophore, and ammonia production | + Shoot and root lengths + Dry shoot and root weights + Chlorophyll content + Numbers of leaves | [90] |
| Triticum aestivum | Pseudomonas azotoformans | P-solubilization, ACC deaminase activity, EPS and IAA production. Expression of biofilm genes AdnA and FliC | + Plant growth + Dry weight of root and shoot + Photosynthetic pigments content − CAT, SOD, and GR activity | [91] |
| Glycine Max L. | Bacillus cereus, Pseudomonas otitidis, and Pseudomonas sp. | P-solubilization, ACC deaminase activity, IAA and ammonia production | + Plant growth + Stomatal density + Relative water content + Chlorophyll pigments + Sugar, protein, and proline content − MDA and H2O2 | [92] |
| Oryza sativa L. | Bacillus megaterium, Bacillus altitudinis, and Bacillus endophyticus | ACC deaminase activity, IAA, EPS, and GA production under stress conditions | + Plant growth + Carotenoids + Total proteins + Sugar content | [93] |
| Zea mayz L. | Bacillus subtilis strains (DHK and B1N1) | P-solubilization, ACC deaminase activity, IAA and siderophore production. Antagonism with Fusarium oxysporum and Rhizoctonia solani | + Plant growth + SOD, POD, and CAT activity + Chlorophyll content + Amino acid content − ROS species | [94] |
| Solanum lycopersicum | Streptomyces strains (IT25 and C-2012) | P-solubilization, siderophore production, ACC deaminase activity. Salinity tolerance (NaCl 13%) | + Plant growth + Leaf relative water content + Proline, MDA, H2O2, and total sugar content + Gene expression of ERF1 and WRKY70 + APX activity − CAT and GPX activity | [95] |
| Triticum aestivum L. | Pseudomonas sp. and Serratia marcescens | P-solubilization, Zn-solubilization, ACC deaminase activity, IAA, EPS, siderophore, and ammonia production | + Osmolyte accumulation + Chlorophyll and carotenoids + Zn and Fe content in grains | [96] |
| Triticum aestivum L. | Pseudomona helmanticensis and Pseudomona baetica | P-solubilization and IAA production in presence of salinity (NaCl 4%) in different drought stress | + Soil p-available + Shoot and root dry weight + Grain yield + P uptake by shoot | [97] |
| Oryza Sativa L. | Diverse PGP microorganism | P-solubilization, siderophore, EPS, N-fixation, expression of nifH and polR genes. Drought tolerance | + Rice seedling + Shoot length + Shoot and root fresh weight + Antioxidant capability + Proline and soluble sugar content | [98] |
| Zea mays L. | Bacillus subtilis and Bacillus safensis | P-solubilization, ACC deaminase activity, IAA, EPS, biofilm, and alginate production | + Total chlorophyll, carotenoid, and soluble sugar − Proline accumulation − Antioxidant enzymes − ACC accumulation, ACC oxidase and ACC synthase under salt stress | [14] |
| Eleusine coracana L. | Variovorax paradoxus, Ochrobactrum anthropi, Pseudomonas palleroniana, Pseudomonas fluorescens, and Pseudomonas palleroniana | P-solubilization, ACC deaminase activity, IAA and siderophore production, N-fixation | + Overall growth parameters and nutrient concentration + SOD, GPX, CAT, and APX activity + Proline, phenol, and chlorophyll − H2O2 and MDA | [99] |
| Sorghum bicolor | Streptomyces sp. and Nocardiopsis sp. | P-solubilization, ACC deaminase activity, IAA and siderophore production under drought, heat, and Cd stress | + Plant growth and photosynthetic pigments + Translocation of Cd from root to shoot + SOD, APX, and CAT activity − MDA concentration | [100] |
| TCrop | AMF + Complement | Specific Effects | Reference |
|---|---|---|---|
| Glycine max L. | Glomus clarum, Glomus mosseae, and Gigaspora margarita + Bradyrhizobium japonicum | + Number of nodules + Grain yield and growth + CAT and POD in seeds + Proline content + Gene expression of CAT and POD − Gene expression of P5CS, P5CR, PDH, and P5CDH | [105] |
| Poncirus trifoliata | Funneliformis mosseae | + Leaf gas exchange + Soil pH, and ammonium content + H+-ATPase activity on shoot and roots + Regulation of H+-ATPase gene PtAHA2 | [106] |
| Poncirus trifoliata | Funneliformis mosseae | + Growth traits and leaf water potential + Gene expression of two aquaporin protein + Chlorophyll concentration | [107] |
| Glycine max L. | Acaulospora laevis, Septoglomus deserticola, and Rhizophagus irregularis + Bacillus amyloliquefaciens | + Plant biomass + Phenol, flavonoid, glycine betaine content and GTS + GA, trans-zeatin-riboside, and IAA in seeds + ATP content and hydrolytic activities of plasma membrane − ABA | [108] |
| Ephedra foliate | Claroideoglomus etunicatum, Rhizophagus intraradices, and Funneliformis mosseae | + Plant growth, chlorophyll content, nitrate and nitrite reductase activity, antioxidant activity, and ascorbic acid content + Content of proline, glucose, and total soluble protein + Sucrose phosphate synthetase activity, IAA, IBA, GA, and ABA − Glutathione level | [109] |
| Nicotiana tabacum | Glomus versiforme + Phosphorus supplementation | + Osmolytes content, proline, sugars, and free amino acids + Antioxidant activities of SOD, CAT, APX, POD, and GR, and AsA and GSH content. + IAA, ABA concentrations in roots and leaves. − ROS accumulation and lipid peroxidation | [110] |
| Solanum lycopersicum | Glomus sp., Sclerocystis sp. and Acaulospora sp. + Acinetobacter sp., and Rahnella aquatilis + compost | + Biomass, fruit number per plant, and fruit yield + Sugar content on shoot − PPO activity and increase of POD activity | [111] |
| Camellia sinensis | Claroideoglomus etunicatum | + Plant growth and leaf water content + Antioxidant activity as SOD, CAT, GPX, and APX + Regulation of CsSODin and CsCAT genes − O2− and MDA content | [112] |
| Vaccinium corymbosum | Funneliformis mosseae | + Proteins involved in amino acid metabolism, antioxidant system, signal transduction, and photosynthesis + Carotenoid biosynthesis + Photosynthetic capacity | [113] |
| Malus hupehensis | Rhizophagus irregularis | + Plant growth + Total chlorophyll content, net photosynthetic rate, stomatal conductance, and transpiration + SOD, POD, and CAT + Proline and total sugar content − Accumulation of MDA, H2O2 and O2− | [114] |
| Populus cathayana | Rhizophagus intraradices | + Plant biomass, root-to-root radio, photosynthetic rate, stomatal conductance + Intercellular CO2 concentration and transpiration rate. + SOD, POD, soluble sugar content especially on shoot + Gene expression of PcGRF10 and PcGRF11 genes induced by AMF | [115] |
| Pheonix dactylifera | AMF consortium + plant growth-promoting rhizobacteria (PGPR) consortium | + Plant biomass, rise of phosphorus uptake, and boosted plant-water relationship + Total soluble sugar and protein content. + Soil organic matter, phosphorus, and glomalin content − H2O2 and MDA accumulation | [116] |
| Solanum lycopersicum | Funneliformis mosseae, Rhizophagus irregularis and Funneliformis coronatum | − H2O2 and MDA content (especially F. mosseae) | [117] |
| Thymus daenensis and Thymus bulgaris | Funneliformis moseae and Rhizophagus intraradices | + Root and shoot dry weight, relative water content, photosynthetic pigments, gas change, and nutritional parameters + Essential oil production + Root colonization and soil spore density − Proline, MDA, electrolyte leakage, and stomatal resistance | [118] |
| Trifolium repens L. | Funneliformis mosseae and Paraglomus occultum | + Root total length, surface area, and volume + Leaf relative water content + SOD, CAT, POD, and ABA levels in root − MDA content | [119] |
4. The “Omics” Approaches and the Development of Optimized Bioinoculants
5. Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010–2018): A Climate Dynamics Perspective. Int. J. Climatol. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of Climate Change on Agriculture and Its Mitigation Strategies: A Review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Gerten, D.; Heck, V.; Jägermeyr, J.; Bodirsky, B.L.; Fetzer, I.; Jalava, M.; Kummu, M.; Lucht, W.; Rockström, J.; Schaphoff, S.; et al. Feeding Ten Billion People Is Possible within Four Terrestrial Planetary Boundaries. Nat. Sustain. 2020, 3, 200–208. [Google Scholar] [CrossRef]
- ODEPA. Reflexiones y Desafíos Al 2030; Apey, A., Barrera, D., Rivas, T., Eds.; ODEPA: Santiago, Chile, 2018; ISBN 9789567244300. [Google Scholar]
- Santander, C.; Aroca, R.; Ruiz-Lozano, J.M.; Olave, J.; Cartes, P.; Borie, F.; Cornejo, P. Arbuscular Mycorrhiza Effects on Plant Performance under Osmotic Stress. Mycorrhiza 2017, 27, 639–657. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Cornejo, P.; Abraham, J.; Valentine, A. Simultaneous Mitigation of Aluminum, Salinity and Drought Stress in Lactuca Sativa Growth via Formulated Plant Growth Promoting Rhodotorula mucilaginosa CAM4. Ecotoxicol. Environ. Saf. 2019, 180, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Silambarasan, S.; Logeswari, P.; Cornejo, P.; Kannan, V.R. Role of Plant Growth–promoting Rhizobacterial Consortium in Improving the Vigna radiata Growth and Alleviation of Aluminum and Drought Stresses. Environ. Sci. Pollut. Res. 2019, 26, 27647–27659. [Google Scholar] [CrossRef] [PubMed]
- Silambarasan, S.; Logeswari, P.; Valentine, A.; Cornejo, P. Role of Curtobacterium herbarum Strain CAH5 on Aluminum Bioaccumulation and Enhancement of Lactuca sativa Growth under Aluminum and Drought Stresses. Ecotoxicol. Environ. Saf. 2019, 183, 109573. [Google Scholar] [CrossRef]
- Rojas, M.; Lambert, F.; Ramirez-Villegas, J.; Challinor, A.J. Emergence of Robust Precipitation Changes across Crop Production Areas in the 21st Century. Proc. Natl. Acad. Sci. USA 2019, 116, 6673–6678. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in Plant Growth-Promoting Bacterial Inoculant Technology: Formulations and Practical Perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The Role of Soil Microorganisms in Plant Mineral Nutrition—Current Knowledge and Future Directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef]
- Cornejo, P.; Seguel, A.; Aguilera, P.; Meier, S.; Larsen, J.; Borie, F. Arbuscular Mycorrhizal Fungi Improve Tolerance of Agricultural Plants to Cope Abiotic Stress Conditions BT. In Plant-Microbe Interactions in Agro-Ecological Perspectives: Microbial Interactions and Agro-Ecological Impacts; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: Singapore, 2017; Volume 2, pp. 55–80. ISBN 978-981-10-6593-4. [Google Scholar]
- Campos, P.; Borie, F.; Cornejo, P.; López-Ráez, J.A.; López-García, Á.; Seguel, A. Phosphorus Acquisition Efficiency Related to Root Traits: Is Mycorrhizal Symbiosis a Key Factor to Wheat and Barley Cropping? Front. Plant Sci. 2018, 9, 752. [Google Scholar] [CrossRef] [PubMed]
- Borie, F.; Aguilera, P.; Castillo, C.; Valentine, A.; Seguel, A.; Barea, J.M.; Cornejo, P. Revisiting the Nature of Phosphorus Pools in Chilean Volcanic Soils as a Basis for Arbuscular Mycorrhizal Management in Plant P Acquisition. J. Soil Sci. Plant Nutr. 2019, 19, 390–401. [Google Scholar] [CrossRef]
- Barea, J.M.; Palenzuela, J.; Cornejo, P.; Sánchez-Castro, I.; Navarro-Fernández, C.; Lopéz-García, A.; Estrada, B.; Azcón, R.; Ferrol, N.; Azcón-Aguilar, C. Ecological and Functional Roles of Mycorrhizas in Semi-Arid Ecosystems of Southeast Spain. J. Arid Environ. 2011, 75, 1292–1301. [Google Scholar] [CrossRef]
- Seguel, A.; Cumming, J.R.; Klugh-Stewart, K.; Cornejo, P.; Borie, F. The Role of Arbuscular Mycorrhizas in Decreasing Aluminium Phytotoxicity in Acidic Soils: A Review. Mycorrhiza 2013, 23, 167–183. [Google Scholar] [CrossRef]
- Seguel, A.; Meier, F.; Azcón, R.; Valentine, A.; Meriño-Gergichevich, C.; Cornejo, P.; Aguilera, P.; Borie, F. Showing Their Mettle: Extraradical Mycelia of Arbuscular Mycorrhizae Form a Metal Filter to Improve Host Al Tolerance and P Nutrition. J. Sci. Food Agric. 2020, 100, 803–810. [Google Scholar] [CrossRef]
- Silambarasan, S.; Logeswari, P.; Cornejo, P.; Kannan, V.R. Evaluation of the Production of Exopolysaccharide by Plant Growth Promoting Yeast Rhodotorula sp. Strain CAH2 under Abiotic Stress Conditions. Int. J. Biol. Macromol. 2019, 121, 55–62. [Google Scholar] [CrossRef]
- Durán, P.; Acuña, J.J.; Jorquera, M.A.; Azcón, R.; Borie, F.; Cornejo, P.; Mora, M.L. Enhanced Selenium Content in Wheat Grain by Co-Inoculation of Selenobacteria and Arbuscular Mycorrhizal Fungi: A Preliminary Study as a Potential Se Biofortification Strategy. J. Cereal Sci. 2013, 57, 275–280. [Google Scholar] [CrossRef]
- Durán, P.; Acuña, J.J.; Armada, E.; López-Castillo, O.M.; Cornejo, P.; Mora, M.L.; Azcón, R. Inoculation with Selenobacteria and Arbuscular Mycorrhizal Fungi to Enhance Selenium Content in Lettuce Plants and Improve Tolerance against Drought Stress. J. Soil Sci. Plant Nutr. 2016, 16, 201–225. [Google Scholar] [CrossRef]
- Larsen, J.; Cornejo, P.; Barea, J.M. Interactions between the Arbuscular Mycorrhizal Fungus Glomus intraradices and the Plant Growth Promoting Rhizobacteria Paenibacillus polymyxa and P. macerans in the Mycorrhizosphere of Cucumis sativus. Soil Biol. Biochem. 2009, 41, 286–292. [Google Scholar] [CrossRef]
- Großkopf, T.; Soyer, O.S. Synthetic Microbial Communities. Curr. Opin. Microbiol. 2014, 18, 72–77. [Google Scholar] [CrossRef]
- Guerrero-Molina, M.F.; Lovaisa, N.C.; Salazar, S.M.; Martínez-Zamora, M.G.; Díaz-Ricci, J.C.; Pedraza, R.O. Physiological, Structural and Molecular Traits Activated in Strawberry Plants after Inoculation with the Plant Growth-Promoting Bacterium Azospirillum brasilense REC3. Plant Biol. 2015, 17, 766–773. [Google Scholar] [CrossRef] [PubMed]
- White, R.A.; Rivas-Ubach, A.; Borkum, M.I.; Köberl, M.; Bilbao, A.; Colby, S.M.; Hoyt, D.W.; Bingol, K.; Kim, Y.M.; Wendler, J.P.; et al. The State of Rhizospheric Science in the Era of Multi-Omics: A Practical Guide to Omics Technologies. Rhizosphere 2017, 3, 212–221. [Google Scholar] [CrossRef]
- Reddy, P.P. Plant Growth Promoting Rhizobacteria for Horticultural Crop Protection; Springer India: New Delhi, India, 2014; pp. 978–981. [Google Scholar]
- Gomiero, T. Soil Degradation, Land Scarcity and Food Security: Reviewing a Complex Challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef]
- Godfray, H.C.J.; Crute, I.R.; Haddad, L.; Muir, J.F.; Nisbett, N.; Lawrence, D.; Pretty, J.; Robinson, S.; Toulmin, C.; Whiteley, R. The Future of the Global Food System. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2769–2777. [Google Scholar] [CrossRef]
- Watts, G.; Battarbee, R.W.; Bloomfield, J.P.; Crossman, J.; Daccache, A.; Durance, I.; Elliott, J.A.; Garner, G.; Hannaford, J.; Hannah, D.M.; et al. Climate Change and Water in the UK—Past Changes and Future Prospects. Prog. Phys. Geogr. 2015, 39, 6–28. [Google Scholar] [CrossRef]
- Mansfield, L.A.; Nowack, P.J.; Kasoar, M.; Everitt, R.G.; Collins, W.J.; Voulgarakis, A. Predicting Global Patterns of Long-Term Climate Change from Short-Term Simulations Using Machine Learning. Npj Clim. Atmospheric Sci. 2020, 3, 44–52. [Google Scholar] [CrossRef]
- Molden, D. Water for Food Water for Life: A Comprehensive Assessment of Water Management in Agriculture; International Water Management Institute: London, UK, 2007; 664p, ISBN 9781844073962. [Google Scholar]
- Lobell, D.B.; Schlenker, W.; Costa-Roberts, J. Climate Trends and Global Crop Production since 1980. Science 2011, 333, 616–620. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of Plants to Water Stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Drought Effects on Root and Tuber Production: A Meta-Analysis. Agric. Water Manag. 2016, 176, 122–131. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Shao, H.B.; Chu, L.Y.; Jaleel, C.A.; Zhao, C.X. Water-Deficit Stress-Induced Anatomical Changes in Higher Plants. Comptes Rendus. Biol. 2008, 331, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, S.; Sawant, K. Drought Stress Adaptation: Metabolic Adjustment and Regulation of Gene Expression. Plant Breed. 2013, 132, 21–32. [Google Scholar] [CrossRef]
- dos Santos, T.B.; Ribas, A.F.; de Souza, S.G.H.; Budzinski, I.G.F.; Domingues, D.S. Physiological Responses to Drought, Salinity, and Heat Stress in Plants: A Review. Stresses 2022, 2, 113–135. [Google Scholar] [CrossRef]
- Lisar, S.Y.; Motafakkerazad, R.; Hossain, M.M.; Rahman, I.M. Water Stress in Plants: Causes, Effects and Responses. In Water Stress; InTech: Rijeka, Croatia, 2012; pp. 1–14. ISBN 978-953-307-963-9. [Google Scholar]
- Fang, Y.; Xiong, L. General Mechanisms of Drought Response and Their Application in Drought Resistance Improvement in Plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer: Dordrecht, The Netherlands, 2009; pp. 153–188. [Google Scholar] [CrossRef]
- Khan, H.A.; Ziaf, K.; Amjad, M.; Iqbal, Q. Exogenous Application of Polyamines Improves Germination and Early Seedling Growth of Hot Pepper. Chil. J. Agric. Res. 2012, 72, 429–433. [Google Scholar] [CrossRef]
- Nakashima, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. The Transcriptional Regulatory Network in the Drought Response and Its Crosstalk in Abiotic Stress Responses Including Drought, Cold, and Heat. Front. Plant Sci. 2014, 5, 170. [Google Scholar] [CrossRef] [PubMed]
- Naylor, D.; Coleman-Derr, D. Drought Stress and Root-Associated Bacterial Communities. Front. Plant Sci. 2018, 8, 2223. [Google Scholar] [CrossRef] [PubMed]
- Cavicchioli, R.; Ripple, W.J.; Timmis, K.N.; Azam, F.; Bakken, L.R.; Baylis, M.; Behrenfeld, M.J.; Boetius, A.; Boyd, P.W.; Classen, A.T.; et al. Scientists’ Warning to Humanity: Microorganisms and Climate Change. Nat. Rev. Microbiol. 2019, 17, 569–586. [Google Scholar] [CrossRef]
- Molina-Montenegro, M.A.; Ballesteros, G.I.; Castro-Nallar, E.; Meneses, C.; Gallardo-Cerda, J.; Torres-Díaz, C. A First Insight into the Structure and Function of Rhizosphere Microbiota in Antarctic Plants Using Shotgun Metagenomic. Polar Biol. 2019, 42, 1825–1835. [Google Scholar] [CrossRef]
- Adedayo, A.A.; Babalola, O.O.; Prigent-Combaret, C.; Cruz, C.; Stefan, M.; Kutu, F.; Glick, B.R. The Application of Plant Growth-Promoting Rhizobacteria in Solanum lycopersicum Production in the Agricultural System: A Review. PeerJ 2022, 10, e13405. [Google Scholar] [CrossRef]
- Kong, Z.; Liu, H. Modification of Rhizosphere Microbial Communities: A Possible Mechanism of Plant Growth Promoting Rhizobacteria Enhancing Plant Growth and Fitness. Front. Plant Sci. 2022, 13, 920813. [Google Scholar] [CrossRef] [PubMed]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere Engineering with Plant Growth-Promoting Microorganisms for Agriculture and Ecological Sustainability. Front. Sustain. Food Syst. 2021, 5, 617157. [Google Scholar] [CrossRef]
- Hassan, M.K.; McInroy, J.A.; Kloepper, J.W. The Interactions of Rhizodeposits with Plant Growth-Promoting Rhizobacteria in the Rhizosphere: A Review. Agriculture 2019, 9, 142. [Google Scholar] [CrossRef]
- Jamil, F.; Mukhtar, H.; Fouillaud, M.; Dufossé, L. Rhizosphere Signaling: Insights into Plant–Rhizomicrobiome Interactions for Sustainable Agronomy. Microorganisms 2022, 10, 899. [Google Scholar] [CrossRef] [PubMed]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The Chemistry of Plant–microbe Interac-tions in the Rhizosphere and the Potential for Metabolomics to Reveal Signaling Related to Defense Priming and Induced Systemic Resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 871, 1473. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Marchesi, J.R.; Mougel, C.; Selosse, M. Host-Microbiota Interactions: From Holobiont Theory to Analysis. Microbiome 2019, 7, 5. [Google Scholar] [CrossRef]
- Li, E.; de Jonge, R.; Liu, C.; Jiang, H.; Friman, V.P.; Pieterse, C.M.J.; Bakker, P.A.H.M.; Jousset, A. Rapid Evolution of Bacte-rial Mutualism in the Plant Rhizosphere. Nat. Commun. 2021, 12, 3829. [Google Scholar] [CrossRef]
- Asghari, B.; Khademian, R.; Sedaghati, B. Plant Growth Promoting Rhizobacteria (PGPR) Confer Drought Resistance and Stimulate Biosynthesis of Secondary Metabolites in Pennyroyal (Mentha pulegium L.) under Water Shortage Condition. Sci. Hortic. 2020, 263, 109132. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. Soil Microbiomes and Climate Change. Nat. Rev. Microbiol. 2020, 18, 35–46. [Google Scholar] [CrossRef]
- Bettenfeld, P.; Cadena i Canals, J.; Jacquens, L.; Fernandez, O.; Fontaine, F.; van Schaik, E.; Courty, P.E.; Trouvelot, S. The Microbiota of the Grapevine Holobiont: A Key Component of Plant Health. J. Adv. Res. 2022, 40, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wahdan, S.F.M.; Tanunchai, B.; Wu, Y.T.; Sansupa, C.; Schädler, M.; Dawoud, T.M.; Buscot, F.; Purahong, W. Deciphering Trifolium Pratense, L. Holobiont Reveals a Microbiome Resilient to Future Climate Changes. MicrobiologyOpen 2021, 10, e1217. [Google Scholar] [CrossRef] [PubMed]
- Govindasamy, V.; George, P.; Kumar, M.; Aher, L.; Raina, S.K.; Rane, J.; Annapurna, K.; Minhas, P.S. Multi-Trait PGP Rhi-zobacterial Endophytes Alleviate Drought Stress in a Senescent Genotype of Sorghum [Sorghum bicolor (L.) Moench]. 3 Biotech 2020, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Scagliola, M.; Valentinuzzi, F.; Mimmo, T.; Cesco, S.; Crecchio, C.; Pii, Y. Bioinoculants as Promising Complement of Chem-ical Fertilizers for a More Sustainable Agricultural Practice. Front. Sustain. Food Syst. 2021, 4, 622169. [Google Scholar] [CrossRef]
- Jansson, C.; Faiola, C.; Wingler, A.; Zhu, X.G.; Kravchenko, A.; de Graaff, M.A.; Ogden, A.J.; Handakumbura, P.P.; Werner, C.; Beckles, D.M. Crops for Carbon Farming. Front. Plant Sci. 2021, 12, 636709. [Google Scholar] [CrossRef]
- Tang, A.; Haruna, A.O.; Majid, N.M.A.; Jalloh, M.B. Potential PGPR Properties of Cellulolytic, Nitrogen-Fixing, Phos-phate-Solubilizing Bacteria in Rehabilitated Tropical Forest Soil. Microorganisms 2020, 8, 442. [Google Scholar] [CrossRef]
- Alzandi, A.A.; Naguib, D.M. Effect of Yeast Application on Soil Health and Root Metabolic Status of Corn Seedlings under Drought Stress. Arch. Microbiol. 2022, 204, 233. [Google Scholar] [CrossRef]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G.A. An Underground Revolution: Biodiversity and Soil Ecological Engineering for Agricultural Sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef]
- Cornejo, P.; Pérez-Tienda, J.; Meier, S.; Valderas, A.; Borie, F.; Azcón-Aguilar, C.; Ferrol, N. Copper Compartmentalization in Spores as a Survival Strategy of Arbuscular Mycorrhizal Fungi in Cu-Polluted Environments. Soil Biol. Biochem. 2013, 57, 925–928. [Google Scholar] [CrossRef]
- Santander, C.; Ruiz, A.; García, S.; Aroca, R.; Cumming, J.; Cornejo, P. Efficiency of Two Arbuscular Mycorrhizal Fungal Inocula to Improve Saline Stress Tolerance in Lettuce Plants by Changes of Antioxidant Defense Mechanisms. J. Sci. Food Agric. 2020, 100, 1577–1587. [Google Scholar] [CrossRef]
- Aponte, H.; Herrera, W.; Cameron, C.; Black, H.; Meier, S.; Paolini, J.; Tapia, Y.; Cornejo, P. Alteration of Enzyme Activities and Functional Diversity of a Soil Contaminated with Copper and Arsenic. Ecotoxicol. Environ. Saf. 2020, 192, 110264. [Google Scholar] [CrossRef] [PubMed]
- Durán, P.; Viscardi, S.; Acuña, J.J.; Cornejo, P.; Azcón, R.; de la Luz Mora, M. Endophytic Selenobacteria and Arbuscular Mycorrhizal Fungus for Selenium Biofortification and Gaeumannomyces graminis Biocontrol. J. Soil Sci. Plant Nutr. 2018, 18, 1021–1035. [Google Scholar] [CrossRef]
- Ma, Y.; Rajkumar, M.; Zhang, C.; Freitas, H. Inoculation of Brassica Oxyrrhina with Plant Growth Promoting Bacteria for the Improvement of Heavy Metal Phytoremediation under Drought Conditions. J. Hazard. Mater. 2016, 320, 36–44. [Google Scholar] [CrossRef]
- Banik, A.; Pandya, P.; Patel, B.; Rathod, C.; Dangar, M. Characterization of Halotolerant, Pigmented, Plant Growth Promot-ing Bacteria of Groundnut Rhizosphere and Its in-Vitro Evaluation of Plant-Microbe Protocooperation to Withstand Salinity and Metal Stress. Sci. Total Environ. 2018, 630, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Meier, S.; Alvear, M.; Borie, F.; Aguilera, P.; Ginocchio, R.; Cornejo, P. Influence of Copper on Root Exudate Patterns in Some Metallophytes and Agricultural Plants. Ecotoxicol. Environ. Saf. 2012, 75, 8–15. [Google Scholar] [CrossRef]
- de la Luz Mora, M.; Demanet, R.; Acuña, J.J.; Viscardi, S.; Jorquera, M.; Rengel, Z.; Durán, P. Aluminum-Tolerant Bacteria Improve the Plant Growth and Phosphorus Content in Ryegrass Grown in a Volcanic Soil Amended with Cattle Dung Manure. Appl. Soil Ecol. 2017, 115, 19–26. [Google Scholar] [CrossRef]
- Sarabia, M.; Cornejo, P.; Azcón, R.; Carreón-Abud, Y.; Larsen, J. Mineral Phosphorus Fertilization Modulates Interactions between Maize, Rhizosphere Yeasts and Arbuscular Mycorrhizal Fungi. Rhizosphere 2017, 4, 89–93. [Google Scholar] [CrossRef]
- Aguilera, P.; Larsen, J.; Borie, F.; Berríos, D.; Tapia, C.; Cornejo, P. New Evidences on the Contribution of Arbuscular My-corrhizal Fungi Inducing Al Tolerance in Wheat. Rhizosphere 2018, 5, 43–50. [Google Scholar] [CrossRef]
- Chávez, D.; Machuca, Á.; Fuentes-Ramirez, A.; Fernandez, N.; Cornejo, P. Shifts in Soil Traits and Arbuscular Mycorrhizal Symbiosis Represent the Conservation Status of Araucaria araucana Forests and the Effects after Fire Events. For. Ecol. Manag. 2020, 458, 117806. [Google Scholar] [CrossRef]
- Ademar Avelar Ferreira, P.; Ceretta, C.A.; Hildebrand Soriani, H.; Luiz Tiecher, T.; Fonsêca Sousa Soares, C.R.; Rossato, L.V.; Nicoloso, F.T.; Brunetto, G.; Paranhos, J.T.; Cornejo, P. Rhizophagus clarus and Phosphate Alter the Physiological Re-sponses of Crotalaria juncea Cultivated in Soil with a High Cu Level. Appl. Soil Ecol. 2015, 91, 37–47. [Google Scholar] [CrossRef]
- Griebenow, S.; Zuniga-Feest, A.; Muñoz, G.; Cornejo, P.; Kleinert, A.; Valentine, A. Photosynthetic Metabolism during Phos-phate Limitation in a Legume from the Mediterranean-Type Fynbos Ecosystem. J. Plant Physiol. 2019, 243, 153051. [Google Scholar] [CrossRef]
- Cornejo, P.; Rubio, R.; Castillo, C.; Azcón, R.; Borie, F. Mycorrhizal Effectiveness on Wheat Nutrient Acquisition in an Acidic Soil from Southern Chile as Affected by Nitrogen Sources. J. Plant Nutr. 2008, 31, 1555–1569. [Google Scholar] [CrossRef]
- Seguel, A.; Cornejo, P.; Ramos, A.; Von Baer, E.; Cumming, J.; Borie, F. Phosphorus Acquisition by Three Wheat Cultivars Contrasting in Aluminium Tolerance Growing in an Aluminium-Rich Volcanic Soil. Crop Pasture Sci. 2017, 68, 305–316. [Google Scholar] [CrossRef]
- Meier, S.; Azcón, R.; Cartes, P.; Borie, F.; Cornejo, P. Alleviation of Cu Toxicity in Oenothera picensis by Copper-Adapted Arbuscular Mycorrhizal Fungi and Treated Agrowaste Residue. Appl. Soil Ecol. 2011, 48, 117–124. [Google Scholar] [CrossRef]
- Parada, J.; Valenzuela, T.; Gómez, F.; Tereucán, G.; García, S.; Cornejo, P.; Winterhalter, P.; Ruiz, A. Effect of Fertilization and Arbuscular Mycorrhizal Fungal Inoculation on Antioxidant Profiles and Activities in Fragaria ananassa Fruit. J. Sci. Food Agric. 2019, 99, 1397–1404. [Google Scholar] [CrossRef]
- Ruiz, A.; Sanhueza, M.; Gómez, F.; Tereucán, G.; Valenzuela, T.; García, S.; Cornejo, P.; Hermosín-Gutiérrez, I. Changes in the Content of Anthocyanins, Flavonols, and Antioxidant Activity in Fragaria ananassa Var. Camarosa Fruits under Tradi-tional and Organic Fertilization. J. Sci. Food Agric. 2019, 99, 2404–2410. [Google Scholar] [CrossRef]
- Aguilera, A.; Tereucán, G.; Ercoli, S.; Cornejo, P.; Gomez, M.R.; Uhlmann, L.; Guigas, C.; Esatbeyoglu, T.; Ruiz, A. Influence of Organic and Chemical Fertilisation on Antioxidant Compounds Profiles and Activities in Fruits of Fragaria ananassa Var. Camarosa. J. Soil Sci. Plant Nutr. 2020, 20, 715–724. [Google Scholar] [CrossRef]
- Bandeppa; Paul, S.; Aggarwal, C.; Manjunatha, B.S.; Rathi, M.S. Characterization of Osmotolerant Rhizobacteria for Plant Growth Promoting Activities in Vitro and during Plant-Microbe Association under Osmotic Stress. Indian J. Exp. Biol 2018, 56, 582–589. [Google Scholar]
- Singh, N.P.; Singh, R.K.; Meena, V.S.; Meena, R.K. Can We Use Maize (Zea mays) Rhizobacteria as Plant Growth Promoter? Vegetos 2015, 28, 86–99. [Google Scholar] [CrossRef]
- Yadav, A.N. Agriculturally Important Micro Biomes: Biodiversity and Multifarious PGP Attributes for Amelioration of Di-verse Abiotic Stresses in Crops for Sustainable Agriculture. Biomed. J. Sci. Tech. Res. 2017, 1, 861–864. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kumar, R.; Kumar, S.; Kumar, V.; Sugitha, T.C.K.; Singh, B.; Chauahan, V.S.; Dhaliwal, H.S.; Saxena, A.K. Beneficial Microbiomes: Biodiversity and Potential Biotechnological Applications for Sustainable Agriculture and Human Health. J. Appl. Biol. Biotechnol. 2017, 5, 45–57. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kumar, V.; Dhaliwal, H.S.; Prasad, R.; Saxena, A.K. Microbiome in Crops: Diversity, Distribution, and Poten-tial Role in Crop Improvement. In New and Future Developments in Microbial Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2018; pp. 305–332. ISBN 9780444639882. [Google Scholar]
- Kour, D.; Rana, K.L.; Kaur, T.; Sheikh, I.; Yadav, A.N.; Kumar, V.; Dhaliwal, H.S.; Saxena, A.K. Microbe-Mediated Allevia-tion of Drought Stress and Acquisition of Phosphorus in Great Millet (Sorghum bicolor L.) by Drought-Adaptive and Phos-phorus-Solubilizing Microbes. Biocatal. Agric. Biotechnol. 2020, 23, 101501. [Google Scholar] [CrossRef]
- Chukwuneme, C.F.; Babalola, O.O.; Kutu, F.R.; Ojuederie, O.B. Characterization of Actinomycetes Isolates for Plant Growth Promoting Traits and Their Effects on Drought Tolerance in Maize. J. Plant Interact. 2020, 15, 93–105. [Google Scholar] [CrossRef]
- Ansari, F.A.; Jabeen, M.; Ahmad, I. Pseudomonas Azotoformans FAP5, a Novel Biofilm-Forming PGPR Strain, Alleviates Drought Stress in Wheat Plant. Int. J. Environ. Sci. Technol. 2021, 18, 3855–3870. [Google Scholar] [CrossRef]
- Dubey, A.; Saiyam, D.; Kumar, A.; Hashem, A.; Abduallah, E.F.; Khan, M.L. Bacterial Root Endophytes: Characterization of Their Competence and Plant Growth Promotion in Soybean (Glycine max (L.) Merr.) under Drought Stress. Int. J. Environ. Res. Public Health 2021, 18, 931. [Google Scholar] [CrossRef] [PubMed]
- Devarajan, A.K.; Muthukrishanan, G.; Truu, J.; Truu, M.; Ostonen, I.; Subramanian Kizhaeral, S.; Panneerselvam, P.; Go-palasubramanian, S.K. The Foliar Application of Rice Phyllosphere Bacteria Induces Drought-Stress Tolerance in Oryza sativa (L.). Plants 2021, 10, 387. [Google Scholar] [CrossRef]
- Sood, G.; Kaushal, R.; Sharma, M. Alleviation of Drought Stress in Maize (Zea mays L.) by Using Endogenous Endophyte Bacillus subtilis in North West Himalayas. Acta Agric. Scand. Sect. B Soil Plant Sci. 2020, 70, 361–370. [Google Scholar] [CrossRef]
- Abbasi, S.; Sadeghi, A.; Safaie, N. Streptomyces Alleviate Drought Stress in Tomato Plants and Modulate the Expression of Transcription Factors ERF1 and WRKY70 Genes. Sci. Hortic. 2020, 265, 109206. [Google Scholar] [CrossRef]
- Khan, A.; Singh, A.V. Multifarious Effect of ACC Deaminase and EPS Producing Pseudomonas sp. and Serratia marcescens to Augment Drought Stress Tolerance and Nutrient Status of Wheat. World J. Microbiol. Biotechnol. 2021, 37, 198. [Google Scholar] [CrossRef]
- Karimzadeh, J.; Alikhani, H.A.; Etesami, H.; Pourbabaei, A.A. Improved Phosphorus Uptake by Wheat Plant (Triticum aes-tivum L.) with Rhizosphere Fluorescent Pseudomonads Strains Under Water-Deficit Stress. J. Plant Growth Regul. 2021, 40, 162–178. [Google Scholar] [CrossRef]
- Pang, Z.; Zhao, Y.; Xu, P.; Yu, D. Microbial Diversity of Upland Rice Roots and Their Influence on Rice Growth and Drought Tolerance. Microorganisms 2020, 8, 1329. [Google Scholar] [CrossRef] [PubMed]
- Chandra, D.; Srivastava, R.; Glick, B.R.; Sharma, A.K. Rhizobacteria Producing ACC Deaminase Mitigate Water-Stress Re-sponse in Finger Millet (Eleusine coracana (L.) Gaertn.). 3 Biotech 2020, 10, 65. [Google Scholar] [CrossRef] [PubMed]
- Silambarasan, S.; Logeswari, P.; Vangnai, A.S.; Kamaraj, B.; Cornejo, P. Plant Growth-Promoting Actinobacterial Inoculant Assisted Phytoremediation Increases Cadmium Uptake in Sorghum bicolor under Drought and Heat Stresses. Environ. Pollut. 2022, 307, 119489. [Google Scholar] [CrossRef] [PubMed]
- Mathur, S.; Tomar, R.S.; Jajoo, A. Arbuscular Mycorrhizal Fungi (AMF) Protects Photosynthetic Apparatus of Wheat under Drought Stress. Photosynth. Res. 2019, 139, 227–238. [Google Scholar] [CrossRef]
- Mathur, S.; Sharma, M.P.; Jajoo, A. Improved Photosynthetic Efficacy of Maize (Zea mays) Plants with Arbuscular Mycor-rhizal Fungi (AMF) under High Temperature Stress. J. Photochem. Photobiol. B Biol. 2018, 180, 149–154. [Google Scholar] [CrossRef]
- Cornejo, P.; Meier, S.; Borie, G.; Rillig, M.C.; Borie, F. Glomalin-Related Soil Protein in a Mediterranean Ecosystem Affected by a Copper Smelter and Its Contribution to Cu and Zn Sequestration. Sci. Total Environ. 2008, 406, 154–160. [Google Scholar] [CrossRef]
- Santander, C.; Vidal, G.; Ruiz, A.; Vidal, C.; Cornejo, P. Salinity Eustress Increases the Biosynthesis and Accumulation of Phenolic Compounds That Improve the Functional and Antioxidant Quality of Red Lettuce. Agronomy 2022, 12, 598. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Ali, D.F.I.; Xiong, Y.C.; Brestic, M.; Skalicky, M.; Hamoud, Y.A.; Ulhassan, Z.; Shaghaleh, H.; AbdElgawad, H.; Farooq, M.; et al. Physiological and Biochemical Responses of Soybean Plants Inoculated with Arbuscular Mycorrhizal Fungi and Bradyrhizobium under Drought Stress. BMC Plant Biol. 2021, 21, 195. [Google Scholar] [CrossRef]
- Cheng, H.Q.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Arbuscular Mycorrhizal Fungi Alleviate Drought Stress in Trifoliate Orange by Regulating H+-ATPase Activity and Gene Expression. Front. Plant Sci. 2021, 12, 659694. [Google Scholar] [CrossRef]
- Zhang, F.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Arbuscular Mycorrhizas Modulate Root Polyamine Metabolism to Enhance Drought Tolerance of Trifoliate Orange. Environ. Exp. Bot. 2020, 171, 103926. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Abd Elgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, H.; Alhaj Hamoud, Y.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and Mycorrhiza Confers Tolerance to Drought Stress and Improve Seed Yield and Quality of Soybean Plant. Physiol. Plant. 2021, 172, 2153–2169. [Google Scholar] [CrossRef] [PubMed]
- Al-Arjani, A.B.F.; Hashem, A.; Abd_Allah, E.F. Arbuscular Mycorrhizal Fungi Modulates Dynamics Tolerance Expression to Mitigate Drought Stress in Ephedra foliata Boiss. Saudi J. Biol. Sci. 2020, 27, 380–394. [Google Scholar] [CrossRef]
- Begum, N.; Ahanger, M.A.; Zhang, L. AMF Inoculation and Phosphorus Supplementation Alleviates Drought Induced Growth and Photosynthetic Decline in Nicotiana tabacum by Up-Regulating Antioxidant Metabolism and Osmolyte Accumulation. Environ. Exp. Bot. 2020, 176, 104088. [Google Scholar] [CrossRef]
- Tahiri, A.-I.; Meddich, A.; Raklami, A.; Alahmad, A.; Bechtaoui, N.; Anli, M.; Göttfert, M.; Heulin, T.; Achouak, W.; Oufdou, K. Assessing the Potential Role of Compost, PGPR, and AMF in Improving Tomato Plant Growth, Yield, Fruit Quality, and Water Stress Tolerance. J. Soil Sci. Plant Nutr. 2022, 22, 743–764. [Google Scholar] [CrossRef]
- Liu, C.Y.; Wang, Y.J.; Wu, Q.S.; Yang, T.Y.; Kuča, K. Arbuscular Mycorrhizal Fungi Improve the Antioxidant Capacity of Tea (Camellia sinensis) Seedlings under Drought Stress. Not. Bot. Horti. Agrobot. Cluj-Napoca 2020, 48, 1993–2005. [Google Scholar] [CrossRef]
- Gui, L.X.; Lu, S.S.; Chen, Q.; Yang, L.; Xiao, J.X. ITRAQ-Based Proteomic Analysis Reveals Positive Impacts of Arbuscular Mycorrhizal Fungi Inoculation on Photosynthesis and Drought Tolerance in Blueberry. Trees—Struct. Funct. 2021, 35, 81–92. [Google Scholar] [CrossRef]
- Huang, D.; Ma, M.; Wang, Q.; Zhang, M.; Jing, G.; Li, C.; Ma, F. Arbuscular Mycorrhizal Fungi Enhanced Drought Re-sistance in Apple by Regulating Genes in the MAPK Pathway. Plant Physiol. Biochem. 2020, 149, 245–255. [Google Scholar] [CrossRef]
- Han, Y.; Lou, X.; Zhang, W.; Xu, T.; Tang, M. Arbuscular Mycorrhizal Fungi Enhanced Drought Resistance of Populus cathayana by Regulating the 14-3-3 Family Protein Genes. Microbiol. Spectr. 2022, 10, e02456-21. [Google Scholar] [CrossRef]
- Akensous, F.Z.; Anli, M.; Boutasknit, A.; Ben-Laouane, R.; Ait-Rahou, Y.; Ben Ahmed, H.; Nasri, N.; Hafidi, M.; Meddich, A. Boosting Date Palm (Phoenix dactylifera L.) Growth under Drought Stress: Effects of Innovative Biostimulants. Gesunde Pflanz. 2022; in press. [Google Scholar] [CrossRef]
- Haddidi, I.; Duc, N.H.; Tonk, S.; Rápó, E.; Posta, K. Defense Enzymes in Mycorrhizal Tomato Plants Exposed to Combined Drought and Heat Stresses. Agronomy 2020, 10, 1657. [Google Scholar] [CrossRef]
- Arpanahi, A.A.; Feizian, M.; Mehdipourian, G.; Khojasteh, D.N. Arbuscular Mycorrhizal Fungi Inoculation Improve Essen-tial Oil and Physiological Parameters and Nutritional Values of Thymus daenensis Celak and Thymus vulgaris L. under Normal and Drought Stress Conditions. Eur. J. Soil Biol. 2020, 100, 103217. [Google Scholar] [CrossRef]
- Liang, S.M.; Jiang, D.J.; Xie, M.M.; Zou, Y.N.; Wu·, Q.S.; Kuca, K. Physiological Responses of Mycorrhizal Symbiosis to Drought Stress in White Clover. Not. Bot. Horti Agrobot. Cluj-Napoca 2021, 49, 1–10. [Google Scholar] [CrossRef]
- Gupta, A.; Bano, A.; Rai, S.; Mishra, R.; Singh, M.; Sharma, S.; Pathak, N. Mechanistic Insights of Plant-Microbe Interaction towards Drought and Salinity Stress in Plants for Enhancing the Agriculture Productivity. Plant Stress 2022, 4, 100073. [Google Scholar] [CrossRef]
- Kour, D.; Khan, S.S.; Kaur, T.; Kour, H.; Singh, G.; Yadav, A.; Yadav, A.N. Drought Adaptive Microbes as Bioinoculants for the Horticultural Crops. Heliyon 2022, 8, e09493. [Google Scholar] [CrossRef] [PubMed]
- Ahkami, A.H.; Allen White, R.; Handakumbura, P.P.; Jansson, C. Rhizosphere Engineering: Enhancing Sustainable Plant Ecosystem Productivity. Rhizosphere 2017, 3, 233–243. [Google Scholar] [CrossRef]
- Foster, K.R.; Bell, T. Competition, Not Cooperation, Dominates Interactions among Culturable Microbial Species. Curr. Biol. 2012, 22, 1845–1850. [Google Scholar] [CrossRef] [PubMed]
- Arzanesh, M.H.; Alikhani, H.A.; Khavazi, K.; Rahimian, H.A.; Miransari, M. Wheat (Triticum aestivum L.) Growth En-hancement by Azospirillum Sp. under Drought Stress. World J. Microbiol. Biotechnol. 2011, 27, 197–205. [Google Scholar] [CrossRef]
- Bresson, J.; Varoquaux, F.; Bontpart, T.; Touraine, B.; Vile, D. The PGPR Strain Phyllobacterium brassicacearum STM196 Induc-es a Reproductive Delay and Physiological Changes That Result in Improved Drought Tolerance in Arabidopsis. New Phytol. 2013, 200, 558–569. [Google Scholar] [CrossRef]
- Liu, F.; Xing, S.; Ma, H.; Du, Z.; Ma, B. Cytokinin-Producing, Plant Growth-Promoting Rhizobacteria That Confer Resistance to Drought Stress in Platycladus orientalis Container Seedlings. Appl. Microbiol. Biotechnol. 2013, 97, 9155–9164. [Google Scholar] [CrossRef]
- Kang, S.M.; Radhakrishnan, R.; Khan, A.L.; Kim, M.J.; Park, J.M.; Kim, B.R.; Shin, D.H.; Lee, I.J. Gibberellin Secreting Rhizo-bacterium, Pseudomonas putida H-2-3 Modulates the Hormonal and Stress Physiology of Soybean to Improve the Plant Growth under Saline and Drought Conditions. Plant Physiol. Biochem. 2014, 84, 115–124. [Google Scholar] [CrossRef]
- Bharti, N.; Pandey, S.S.; Barnawal, D.; Patel, V.K.; Kalra, A. Plant Growth Promoting Rhizobacteria Dietzia natronolimnaea Modulates the Expression of Stress Responsive Genes Providing Protection of Wheat from Salinity Stress. Sci. Rep. 2016, 6, 34768. [Google Scholar] [CrossRef]
- Ortíz-Castro, R.; Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; López-Bucio, J. The Role of Microbial Signals in Plant Growth and Development. Plant Signal. Behav. 2009, 4, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Dangi, A.K.; Sharma, B.; Khangwal, I.; Shukla, P. Combinatorial Interactions of Biotic and Abiotic Stresses in Plants and Their Molecular Mechanisms: Systems Biology Approach. Mol. Biotechnol. 2018, 60, 636–650. [Google Scholar] [CrossRef] [PubMed]
- Imam, J.; Singh, P.K.; Shukla, P. Plant Microbe Interactions in Post Genomic Era: Perspectives and Applications. Front. Microbiol. 2016, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pinu, F.R.; Beale, D.J.; Paten, A.M.; Kouremenos, K.; Swarup, S.; Schirra, H.J.; Wishart, D. Systems Biology and Multi-Omics Integration: Viewpoints from the Metabolomics Research Community. Metabolites 2019, 9, 76. [Google Scholar] [CrossRef]
- Llorens, E.; Sharon, O.; Camañes, G.; García-Agustín, P.; Sharon, A. Endophytes from Wild Cereals Protect Wheat Plants from Drought by Alteration of Physiological Responses of the Plants to Water Stress. Environ. Microbiol. 2019, 21, 3299–3312. [Google Scholar] [CrossRef]
- Omar, S.A.; Fetyan, N.A.H.; Eldenary, M.E.; Abdelfattah, M.H.; Abd-Elhalim, H.M.; Wrobel, J.; Kalaji, H.M. Alteration in Expression Level of Some Growth and Stress-Related Genes after Rhizobacteria Inoculation to Alleviate Drought Tolerance in Sensitive Rice Genotype. Chem. Biol. Technol. Agric. 2021, 8, 41. [Google Scholar] [CrossRef]
- Jayakumar, A.; Padmakumar, P.; Nair, I.C.; Radhakrishnan, E.K. Drought Tolerant Bacterial Endophytes with Potential Plant Probiotic Effects from Ananas Comosus. Biologia 2020, 75, 1769–1778. [Google Scholar] [CrossRef]
- Saikia, J.; Sarma, R.K.; Dhandia, R.; Yadav, A.; Bharali, R.; Gupta, V.K.; Saikia, R. Alleviation of Drought Stress in Pulse Crops with ACC Deaminase Producing Rhizobacteria Isolated from Acidic Soil of Northeast India. Sci. Rep. 2018, 8, 3560. [Google Scholar] [CrossRef]
- Nephali, L.; Moodley, V.; Piater, L.; Steenkamp, P.; Buthelezi, N.; Dubery, I.; Burgess, K.; Huyser, J.; Tugizimana, F. A Metabolomic Landscape of Maize Plants Treated with a Microbial Biostimulant Under Well-Watered and Drought Conditions. Front. Plant Sci. 2021, 12, 676632. [Google Scholar] [CrossRef]
- Lephatsi, M.; Nephali, L.; Meyer, V.; Piater, L.A.; Buthelezi, N.; Dubery, I.A.; Opperman, H.; Brand, M.; Huyser, J.; Tugizimana, F. Molecular Mechanisms Associated with Microbial Biostimulant-Mediated Growth Enhancement, Priming and Drought Stress Tolerance in Maize Plants. Sci. Rep. 2022, 12, 10450. [Google Scholar] [CrossRef]
- Adeleke, B.S.; Babalola, O.O. Meta-Omics of Endophytic Microbes in Agricultural Biotechnology. Biocatal. Agric. Biotechnol. 2022, 42, 102332. [Google Scholar] [CrossRef]
- Chen, X.L.; Sun, M.C.; Chong, S.L.; Si, J.P.; Wu, L.S. Transcriptomic and Metabolomic Approaches Deepen Our Knowledge of Plant–Endophyte Interactions. Front. Plant Sci. 2022, 12, 700200. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, T.; Shukla, P. Bioinoculant Capability Enhancement through Metabolomics and Systems Biology Approaches. Brief. Funct. Genom. 2018, 18, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.M.; Zhang, F.; Zou, Y.N.; Kuča, K.; Wu, Q.S. Metabolomics Analysis Reveals Drought Responses of Trifoliate Orange by Arbuscular Mycorrhizal Fungi with a Focus on Terpenoid Profile. Front. Plant Sci. 2021, 12, 102332. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A.; Rahman, M.A.; Guo, J.; Kang, Z.; Babar, M.A. Comparative Physiological and Metabolic Analysis Reveals a Complex Mechanism Involved in Drought Tolerance in Chickpea (Cicer arietinum L.) Induced by PGPR and PGRs. Sci. Rep. 2019, 9, 2097. [Google Scholar] [CrossRef]
- Tyers, M.; Mann, M. From Genomics to Proteomics: One Approach. Am. Lab. 2003, 34, 28. [Google Scholar]
- Alberton, D.; Valdameri, G.; Moure, V.R.; Monteiro, R.A.; de Pedrosa, F.O.; Müller-Santos, M.; de Souza, E.M. What Did We Learn from Plant Growth-Promoting Rhizobacteria (PGPR)-Grass Associations Studies Through Proteomic and Metabolomic Approaches? Front. Sustain. Food Syst. 2020, 4, 607343. [Google Scholar] [CrossRef]
- De Cremer, K.; Mathys, J.; Vos, C.; Froenicke, L.; Michelmore, R.W.; Cammue, B.P.A.; De Coninck, B. RNAseq-Based Tran-scriptome Analysis of Lactuca sativa Infected by the Fungal Necrotroph Botrytis cinerea. Plant Cell Environ. 2013, 36, 1992–2007. [Google Scholar] [CrossRef]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From Saprophytes to Endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef]
- Belbahri, L.; Bouket, A.C.; Rekik, I.; Alenezi, F.N.; Vallat, A.; Luptakova, L.; Petrovova, E.; Oszako, T.; Cherrad, S.; Vacher, S.; et al. Comparative Genomics of Bacillus amyloliquefaciens Strains Reveals a Core Genome with Traits for Habitat Adapta-tion and a Secondary Metabolites Rich Accessory Genome. Front. Microbiol. 2017, 8, 1438. [Google Scholar] [CrossRef]
- Nordstedt, N.P.; Jones, M.L. Genomic Analysis of Serratia plymuthica MBSA-MJ1: A Plant Growth Promoting Rhizobacteria That Improves Water Stress Tolerance in Greenhouse Ornamentals. Front. Microbiol. 2021, 12, 653556. [Google Scholar] [CrossRef] [PubMed]
- Suarez, C.; Ratering, S.; Hain, T.; Fritzenwanker, M.; Goesmann, A.; Blom, J.; Chakraborty, T.; Bunk, B.; Spröer, C.; Overmann, J.; et al. Complete Genome Sequence of the Plant Growth-Promoting Bacterium Hartmannibacter diazotrophicus Strain E19T. Int. J. Genomics 2019, 2019, 7586430. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Franken, P.; Schneider, C.; Schwarz, D.; Giovannetti, M.; Agnolucci, M.; De Pascale, S.; Bonini, P.; Colla, G. Arbuscular Mycorrhizal Fungi Act as Biostimulants in Horticultural Crops. Sci. Hortic. 2015, 196, 91–108. [Google Scholar] [CrossRef]
- Broadhurst, D.I.; Kell, D.B. Statistical Strategies for Avoiding False Discoveries in Metabolomics and Related Experiments. Metabolomics 2006, 2, 171–196. [Google Scholar] [CrossRef]
- Hultman, J.; Waldrop, M.P.; Mackelprang, R.; David, M.M.; McFarland, J.; Blazewicz, S.J.; Harden, J.; Turetsky, M.R.; McGuire, A.D.; Shah, M.B.; et al. Multi-Omics of Permafrost, Active Layer and Thermokarst Bog Soil Microbiomes. Nature 2015, 521, 208–212. [Google Scholar] [CrossRef]
- Kuo, T.C.; Tian, T.F.; Tseng, Y.J. 3Omics: A Web-Based Systems Biology Tool for Analysis, Integration and Visualization of Human Transcriptomic, Proteomic and Metabolomic Data. BMC Syst. Biol. 2013, 7, 64. [Google Scholar] [CrossRef]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards More Trans-parent and Integrative Metabolomics Analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef]
- FAO. The State of the World’s Biodiversity for Food and Agriculture; Bélanger, J., Pilling, D., Eds.; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2019; 572p, Available online: http://www.fao.org/3/CA3129EN/CA3129EN.pdf (accessed on 17 August 2022).

| Methods Used | Approach | Crop | Microorganism | Crop Effects | Reference |
|---|---|---|---|---|---|
| UPLC-QTOF MS | Metabolomics | Tritricum aestivum | Acremonium sclerotigenum, Sarocladium implicatum | + Proline − ABA − Lipid peroxidation − Malondialdehide − Ferulic acid | [133] |
| RT-QPCR | Genomic | Oryza sativa L. | Diverse PGP microorganism | + Soil enzyme activities (dehydrogenase, nitrogenase, urease, and alkaline phosphatase) + Regulation of growth and stress-related genes (COX1, AP2-EREBP, GRAM, NRAMP6, NAM, GST, and DHN) | [134] |
| LS-MS/MS | Metabolomics | Ananas comosus | Consortium of Staphylococcus sp. | + Indole acetic acid + ACC deaminase + Promotion of plant growth | [135] |
| RT-QPCR | Genomic | Vigna mungo L. | Ochrobactrum pseudogrignonense RJ12, Pseudomonas sp. RJ15, and Bacillus subtilis RJ46 | + Seed germination, root length, shoot length, and dry weight of treated plants − Regulation of ACC deaminase gene | [136] |
| RT-QPCR | Genomic | Pisum sativum L. | Ochrobactrum pseudogrignonense RJ12, Pseudomonas sp. RJ15, and Bacillus subtilis RJ46 | + Seed germination, root length, shoot length, and dry weight of treated plants − Regulation of ACC deaminase gene | [136] |
| UHPLC-HDMS | Metabolomics | Zea mays | Bacillus licheniformis, Brevibacillus laterosporus, and Bacillus amyloliquefaciens | + Phenylpropanoid biosynthesis + Glycine, serine, and threonine metabolism + Tyrosine metabolism + TCA cycle metabolism | [137] |
| LC-ESI-QqQ-MS | Metabolomics | Zea mays | Bacillus licheniformis, Brevibacillus laterosporus, and Bacillus amyloliquefaciens | + Salicylic acid + Indole-3-carboxylic acid + Glycine, cysteine, and tyrosine + Apigenin, apigetrin, and vicenin | [138] |
| RT-QPCR and Elisa | Genomic | Zea mays | Bacillus licheniformis, Brevibacillus laterosporus, and Bacillus amyloliquefaciens | + Global DNA methylation + Regulation of PAL and FSNII gene | [138] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidal, C.; González, F.; Santander, C.; Pérez, R.; Gallardo, V.; Santos, C.; Aponte, H.; Ruiz, A.; Cornejo, P. Management of Rhizosphere Microbiota and Plant Production under Drought Stress: A Comprehensive Review. Plants 2022, 11, 2437. https://doi.org/10.3390/plants11182437
Vidal C, González F, Santander C, Pérez R, Gallardo V, Santos C, Aponte H, Ruiz A, Cornejo P. Management of Rhizosphere Microbiota and Plant Production under Drought Stress: A Comprehensive Review. Plants. 2022; 11(18):2437. https://doi.org/10.3390/plants11182437
Chicago/Turabian StyleVidal, Catalina, Felipe González, Christian Santander, Rodrigo Pérez, Víctor Gallardo, Cledir Santos, Humberto Aponte, Antonieta Ruiz, and Pablo Cornejo. 2022. "Management of Rhizosphere Microbiota and Plant Production under Drought Stress: A Comprehensive Review" Plants 11, no. 18: 2437. https://doi.org/10.3390/plants11182437
APA StyleVidal, C., González, F., Santander, C., Pérez, R., Gallardo, V., Santos, C., Aponte, H., Ruiz, A., & Cornejo, P. (2022). Management of Rhizosphere Microbiota and Plant Production under Drought Stress: A Comprehensive Review. Plants, 11(18), 2437. https://doi.org/10.3390/plants11182437