
In Vitro Polyploid Induction of Highbush Blueberry through De Novo Shoot Organogenesis


 ,
,  ,
,  and
and 
Abstract
:1. Introduction
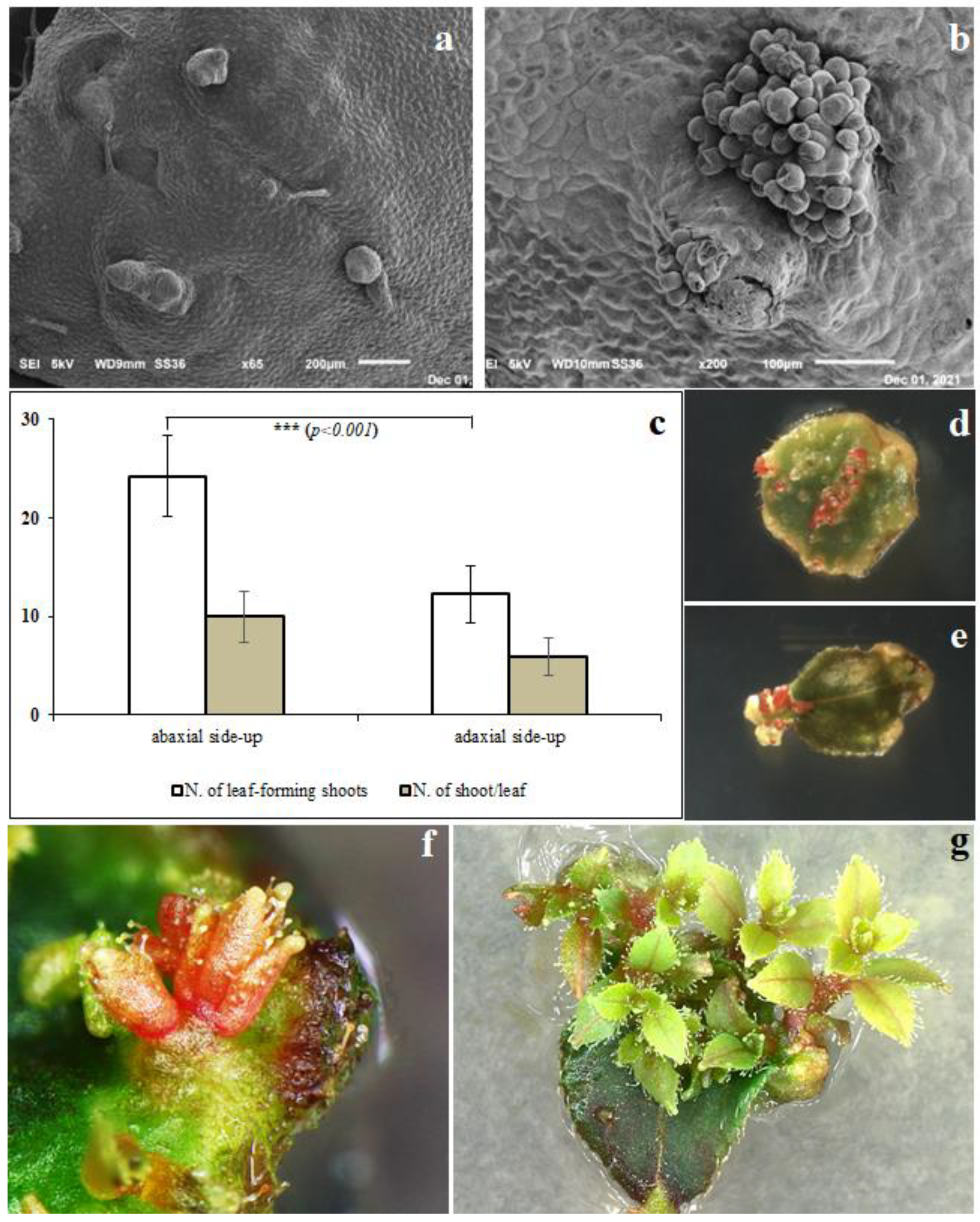
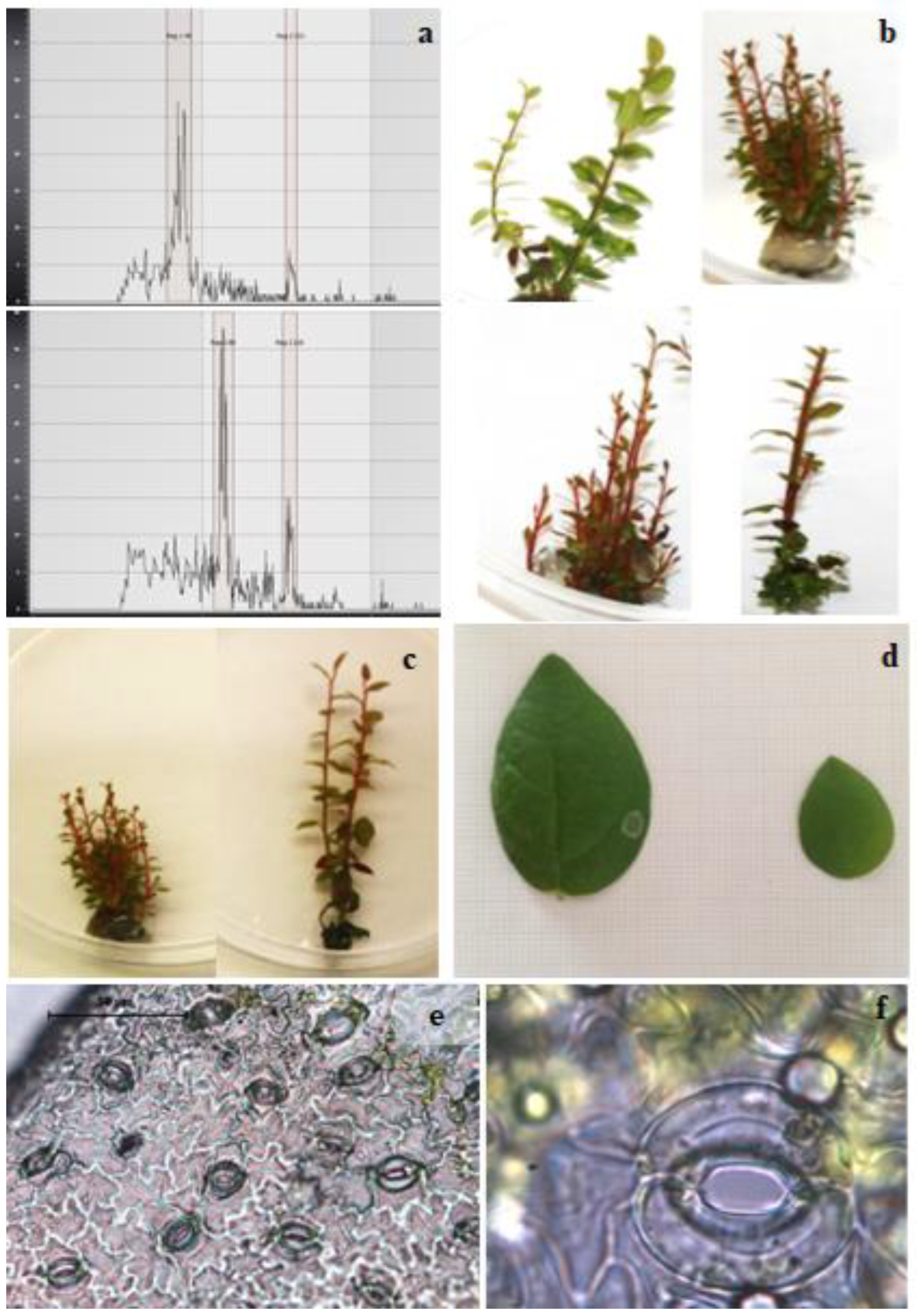
2. Results and Discussions
3. Materials and Methods
3.1. Plant Material
3.2. Adventitious Shoot Organogenesis and Polyploid Induction I
3.3. Media Preparation and Sterilization Conditions
3.4. Ploidy Evaluation
3.5. Electron Microscopy and Stomata Traits
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrao, L.F.V.; Amadeu, R.R.; Benvenuto, J.; De Bem Oliveira, I.; Munoz, P.R. Genomic selection in an outcrossing auto-tetraploid fruit crop: Lesson from blueberry breeding. Front. Plant Sci. 2021, 12, 676326. [Google Scholar] [CrossRef]
- Han, X.; Yang, Y.; Han, X.; Ryner, J.T.; Ahmed, E.A.H.; Qi, Y.; Zhong, G.; Song, G. CRISPR Cas9- and Cas12a-me-diated gusA editing in transgenic blueberry. Plant Cell Tiss. Organ Cult. 2022, 148, 217–229. [Google Scholar] [CrossRef]
- Gallardo, R.K.; Zhang, Q.; Dossett, M.; Polashock, J.J.; Rodriguez-Saona, C.; Vorsa, N.; Edger, P.P.; Ashrafi, H.; Babiker, E.; Finn, C.E.; et al. Breeding trait priorities of the blueberry industry in the United States and Canada. HortScience 2018, 53, 1021. [Google Scholar] [CrossRef]
- Ballington, J.R. The role of interspecific hybridization in blueberry improvement. Acta Hortic. 2009, 810, 49–59. [Google Scholar] [CrossRef]
- Brevis, P.; Bassil, N.; Ballington, J.; Hancock, J. Impact of wide hybridization on highbush blueberry breeding. J. Am. Soc. Hortic. Sci. 2008, 133, 11. [Google Scholar] [CrossRef]
- Schaart, J.G.; Van de Wiel, C.C.M.; Smulders, M.J.M. Genome editing of polyploid crops: Prospects, achievements, and bottlenecks. Transgenic Res. 2021, 30, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Song, G.Q. Blueberry (Vaccinium corymbosum L.). Methods Mol. Biol. 2015, 1224, 121–131. [Google Scholar] [PubMed]
- Walworth, A.; Song, G.Q. The cold-regulated genes of blueberry and their response to overexpression of VcDDF1 in several tissues. Int. J. Mol. Sci. 2018, 19, 1553. [Google Scholar] [CrossRef] [PubMed]
- Song, G.Q.; Walworth, A.; Lin, T.; Chen, Q.; Han, X.; Irina Zaharia, L.; Zhong, G.Y. VcFT-induced mobile florigenic signals in transgenic and transgrafted blueberries. Hortic. Res. 2019, 6, 105. [Google Scholar] [CrossRef]
- Song, G.Q.; Chen, Q. Overexpression of the MADS-box gene K-domain increases the yield potential of blueberry. Plant Sci. 2018, 276, 10. [Google Scholar] [CrossRef]
- Lin, T.Y.; Walworth, A.; Zong, X.J.; Danial, G.H.; Tomaszewski, E.M.; Callow, P.; Han, X.M.; Zaharia, L.I.; Edger, P.P.; Zhong, G.Y.; et al. VcRR2 regulates chilling-mediated flowering through expression of hormone genes in a trans-genic blueberry mutant. Hortic. Res. 2019, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- Surridge, C. Blueberry fooled into flowering. Nat. Plants 2019, 5, 910. [Google Scholar] [CrossRef] [PubMed]
- Omori, M.; Yamane, H.; Osakabe, K.; Osakabe, Y.; Tao, R. Targeted mutagenesis of CENTRORADIALIS using CRISPR/Cas9 system through the improvement of genetic transformation efficiency of tetraploid highbush blueberry. J. Hortic. Sci. Biotechnol. 2021, 96, 153–161. [Google Scholar] [CrossRef]
- Rugini, E.; Silvestri, C.; Ceccarelli, M.; Muleo, R.; Cristofori, V. Mutagenesis, and biotechnology techniques as tools for selecting new stable diploid and tetraploid olive genotypes and their dwarfing agronomical characterization. HortScience 2016, 51, 799–804. [Google Scholar] [CrossRef]
- Rowland, L.J.; Ogden, E.L. Use of a cytokinin conjugate for efficient shoot regeneration from leaf sections of highbush blueberry. HortScience 1992, 27, 1127–1129. [Google Scholar] [CrossRef]
- Cao, X.; Hammerschlag, F.A. Improved shoot organogenesis from leaf explants of highbush blueberry. HortScience 2000, 35, 945–947. [Google Scholar] [CrossRef]
- Manzoor, A.; Ahmad, T.; Bashir, M.A.; Hafiz, I.A.; Silvestri, C. Studies on colchicine induced chromosome doubling for enhancement of quality traits in ornamental plants. Plants 2019, 8, 194. [Google Scholar] [CrossRef]
- Frank, M.H.; Chitwood, D.H. Plant chimeras: The good, the bad and the “Bizzarria”. Dev. Biol. 2016, 419, 42–53. [Google Scholar] [CrossRef]
- Malabarba, J.; Chevreau, E.; Dousset, N.; Veillet, F.; Moizan, J.; Vergne, E. New strategies to overcome present CRISPR/Cas9 limitation in apple and pear: Efficient dechimerization and base editing. Int. J. Mol. Sci. 2021, 22, 319. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.Q.; Tu, H.; Liang, W.J.; Long, J.M.; Wu, X.M.; Zhang, H.Y.; Guo, W.W. Comparative metabolic and transcriptional analysis of a doubled diploid and its diploid citrus rootstock (C. junos cv. Ziyang xiangcheng) suggests its potential value for stress resistance improvement. BMC Plant Biol. 2015, 15, 89. [Google Scholar] [CrossRef] [Green Version]
- Stupar, R.M.; Bhaskar, P.B.; Yandell, B.S.; Rensink, W.A.; Hart, A.L.; Ouyang, S.; Veilleux, R.E.; Busse, J.S.; Erhardt, R.J.; Buell, C.R.; et al. Phenotypic and transcriptomic changes associated with potato autopolyploidization. Genetics 2007, 176, 2055–2067. [Google Scholar] [CrossRef] [PubMed]
- Riddle, N.C.; Jiang, H.; An, L.; Doerge, R.W.; Birchler, J.A. Gene expression analysis at the intersection of ploidy and hybridity in maize. Theor. Appl. Genet. 2010, 120, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Allario, T.; Brumos, J.; Colmenero-Flores, J.M.; Tadeo, F.; Froelicher, Y.; Talon, M.; Navarro, L.; Ollitrault, P.; Morrison, R. Large changes in anatomy and physiology between diploid rangpure lime (Citrus limonia) and its autotetraploid are not associated with large changes in leaf gene expression. J. Exp. Bot. 2011, 62, 2507–2509. [Google Scholar] [CrossRef] [PubMed]
- Rugini, E.; Pannelli, G.; Ceccarelli, M.; Muganu, M. Isolation of triploid and tetraploid olive (Olea europaea L.) plants from mixoploid cv. ‘Frantoio’and ‘Leccino’mutants by in vivo and in vitro selection. Plant Breed. 1996, 115, 23–27. [Google Scholar] [CrossRef]
- Allario, T.; Brumos, J.; Colmenero-Flores, J.M.; Iglesias, D.J.; Pina, J.A.; Navarro, L.; Morillon, R. Tetraploid rangpur lime rootstocks increase drought tolerance via enhanced constitutive root abscisic acid production. Plant Cell Environ. 2013, 36, 856–868. [Google Scholar] [CrossRef]
- Meiners, J.; Schwab, M.; Szankowski, I. Efficient in vitro regeneration systems for Vaccinium species. Plant Cell Tiss. Org. Cult. 2007, 89, 169–176. [Google Scholar] [CrossRef]
- Debnath, S.C.; Mcrae, K.B. An efficient adventitious shoot regeneration system on excised leaves of micropropagated lingonberry (Vaccinium vitis-idaea L.). J. Hortic. Sci. Biotechnol. 2002, 77, 744–752. [Google Scholar] [CrossRef]
- Debnath, S.C. Strawberry sepal: Another explant for thidiazuron-induced adventitious shoot regeneration. Vitr. Cell Dev. Biol. Plant 2005, 41, 671–676. [Google Scholar] [CrossRef]
- Schuchovski, C.; Sant’Anna Santos, B.F.; Marra, R.C.; Biasi, L.A. Morphological and anatomical insights into de novo shoot organogenesis of in vitro “Delite” rabbiteye blueberries. Helyon 2020, 6, e05468. [Google Scholar] [CrossRef]
- Liu, C.; Callow, P.; Rowland, L.J.; Hancock, J.F.; Song, G.Q. Adventitious shoot regeneration from leaf explants of southern highbush blueberry cultivars. Plant Cell Tiss. Organ Cult. 2010, 103, 137–144. [Google Scholar] [CrossRef]
- Rauf, S.; Ortiz, R.; Malinowski, D.P.; Clarindo, W.R.; Kainat, W.; Shehzad, M.; Hassan, S.W. Induced polyploidy: A tool for forage species improvement. Agriculture 2021, 11, 210. [Google Scholar] [CrossRef]
- Touchell, D.H.; Palmer, I.E.; Ranney, T.G. In vitro ploidy manipulation for crop improvement. Front. Plant Sci. 2020, 11, 722. [Google Scholar] [CrossRef]
- Eng, W.; Ho, W. Polyploidization using colchicine in horticultural plants: A review. Sci. Hortic. 2019, 246, 604–617. [Google Scholar] [CrossRef]
- Rêgo, M.M.; Rêgo, E.R.; Bruckner, C.H.; Finger, F.L.; Otoni, W.C. In vitro induction of autotetraploids from diploid yellow passion fruit mediated by colchicine and oryzalin. Plant Cell Tiss. Organ Cult. 2011, 107, 451–459. [Google Scholar] [CrossRef]
- Chalak, L.; Legave, J.M. Oryzalin combined with adventitious regeneration for an efficient chromosomoe doubling of trihaploid kiwifruit. Plant Cell Rep. 1996, 16, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Vainola, A. Polyploidization and early screening of Rhododendron hybrids. Euphytica 2000, 112, 239–244. [Google Scholar] [CrossRef]
- Carvalho, J.F.R.; Carvalho, C.R.; Otoni, W.C. In vitro induction of polyploidy in annatto (Bixa orellana). Plant Cell Tiss. Organ Cult. 2005, 80, 69–75. [Google Scholar] [CrossRef]
- Van Duren, M.; Morpurgo, R.; Dolezel, J.; Afza, R. Induction, and verification of autotetraploids in diploid banana (Musa acuminata) by in vitro techniques. Euphytica 1996, 88, 25–34. [Google Scholar] [CrossRef]
- Sajjad, Y.; Jaskani, M.J.; Mehmood, A.; Ahmad, I.; Abbas, H. Effect of colchicine on in vitro polyploidy induction in African marigold (Tagetes erecta). Pak. J. Bot. 2013, 45, 1255–1258. [Google Scholar]
- Eng, W.H.; Ho, W.S.; Ling, K.H. In vitro induction and identification of polyploid Neolamarckia cadamba plants by colchicine treatment. Peer J. 2021, 9, e12399. [Google Scholar] [CrossRef]
- Pirkoohi, M.H.; Keyvanloo, M.; Hassanpur, M. Colchicine induced polyploidy in mint by seed treatment. Int. J. Agric. Crop Sci. 2011, 3, 102–104. [Google Scholar]
- Aleza, P.; Juárez, J.; Ollitrault, P.; Navarro, L. Production of tetraploid plants of non-apomictic citrus genotypes. Plant Cell Rep. 2009, 28, 1837–1846. [Google Scholar] [CrossRef]
- Regalado, J.J.; Carmona-Martín, E.; Castro, P.; Moreno, R.; Gil, J.; Encina, C.L. Study of the somaclonal variation produced by different methods of polyploidization in Asparagus officinalis L. Plant Cell Tiss. Organ Cult. 2015, 122, 31–44. [Google Scholar] [CrossRef]
- Zhou, H.W.; Zeng, W.D.; Yan, H.B. In vitro induction of tetraploids in cassava variety Xinxuan 048′using colchicine. Plant Cell Tiss. Organ Cult. 2017, 28, 723–729. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, S.; Hu, J.; He, G.; Liu, Y.; Chen, X.; Lei, T.; Li, Q.; Yang, L.; Li, W.; et al. Polyploidization of Plumbago auriculata Lam, in vitro and its characterization including cold tolerance. Plant Cell Tiss. Organ Cult. 2020, 140, 315–325. [Google Scholar] [CrossRef]
- Comai, L. The advantages and disadvantages of being polyploid. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Predieri, S. Mutation induction and tissue culture in improving fruits. Plant Cell Tiss. Organ Cult. 2001, 64, 185–210. [Google Scholar] [CrossRef]
- Tan, F.Q.; Zhang, M.; Xie, K.D.; Fan, Y.J.; Song, X.; Wang, R.; Wu, X.M.; Zhang, H.Y.; Guo, W.W. Polyploidy remodels fruit metabolism by modifying carbon source utilization and metabolic flux in Ponkan mandarin. Plant Sci. 2019, 289, 110276. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.Q.; Tu, H.; Wang, R.; Wu, X.M.; Xie, K.D.; Chen, J.J.; Zhang, H.Y.; Xu, J.; Guo, W.W. Metabolic adaptation following genome doubling in citrus doubled diploids revealed by non-targeted metabolomics. Metabolomics 2017, 13, 143. [Google Scholar] [CrossRef]
- Silvestri, C.; Rugini, E.; Cristofori, V. The effect of CuSO4 for establishing in vitro culture, and the role nitrogen and iron sources in in vitro multiplication of corylus avellana L. cv. tonda gentile romana. Plant Biosyst. 2020, 154, 17–23. [Google Scholar] [CrossRef]
- Lloyd, G.; McCown, B. Commercially feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot tip culture. Intl. Plant Prop. Soc. Proc. 1980, 30, 421–427. [Google Scholar]
- Doležel, J.; Binarová, P.; Lucretti, S. Analysis of nuclear DNA content in plant cells by flow cytometry. Biol. Plant 1989, 31, 113–120. [Google Scholar] [CrossRef]
- Kuzminsky, E.; Meschini, R.; Terzoli, S.; Pavani, L.; Silvestri, C.; Choury, Z.; Scarascia-Mugnozza, G. Isolation of mesophyll protoplasts from Mediterranean woody plants for the study of DNA integrity under abiotic stress. Front. Plant Sci. 2016, 7, 1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demurtas, O.C.; Frusciante, S.; Ferrante, P.; Diretto, G.; Azad, N.H.; Pietrella, M.; Aprea, G.; Taddei, A.R.; Romano, E.; Mi, J.; et al. Candidate enzymes for saffron crocin biosynthesis are localized in multiple cellular compartments. Plant Physiol. 2018, 177, 990–1006. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Treatment Colchicine (µM) | Leaf-Forming Buds (%) | No. of Buds per Leaf | No. of Regenerated Shoots | No. of Tetraploids x | No. of Mixoploids y | Tetraploid Induction Rate (%) z |
|---|---|---|---|---|---|---|
| 0 (Control) | 81.5 ± 6.1 a | 8.2 ± 1.6 a | 244 | 0 | 0 | - |
| 25 | 36.5 ± 7.8 b | 4.2 ± 0.7 b | 102 | 8 | 31 | 7.8 ± 1.2 b |
| 250 | 4.5 ± 3.2 c | 1.3 ± 0.4 c | 63 | 7 | 3 | 11.0 ± 1.4 a |
| 500 | - | - | - | - | - | - |
| Ploidy Level | Stomata Length (µm) | Stomata Length (µm) | Stomata Density (N/mm2) |
|---|---|---|---|
| Diploid | 11.91 ± 0.63 a | 5.2 ± 0.56 a | 854.6 ± 22.1 a |
| Tetraploid | 18.59 ± 0.51 b | 12.45 ± 1.12 b | 491.2 ± 52.1 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marangelli, F.; Pavese, V.; Vaia, G.; Lupo, M.; Bashir, M.A.; Cristofori, V.; Silvestri, C. In Vitro Polyploid Induction of Highbush Blueberry through De Novo Shoot Organogenesis. Plants 2022, 11, 2349. https://doi.org/10.3390/plants11182349
Marangelli F, Pavese V, Vaia G, Lupo M, Bashir MA, Cristofori V, Silvestri C. In Vitro Polyploid Induction of Highbush Blueberry through De Novo Shoot Organogenesis. Plants. 2022; 11(18):2349. https://doi.org/10.3390/plants11182349
Chicago/Turabian StyleMarangelli, Federico, Vera Pavese, Giuseppe Vaia, Michela Lupo, Muhammad Ajmal Bashir, Valerio Cristofori, and Cristian Silvestri. 2022. "In Vitro Polyploid Induction of Highbush Blueberry through De Novo Shoot Organogenesis" Plants 11, no. 18: 2349. https://doi.org/10.3390/plants11182349
APA StyleMarangelli, F., Pavese, V., Vaia, G., Lupo, M., Bashir, M. A., Cristofori, V., & Silvestri, C. (2022). In Vitro Polyploid Induction of Highbush Blueberry through De Novo Shoot Organogenesis. Plants, 11(18), 2349. https://doi.org/10.3390/plants11182349