
Over 30 Years of Misidentification: A New Nothospecies Lycoris × jinzheniae (Amaryllidaceae) in Eastern China, Based on Molecular, Morphological, and Karyotypic Evidence


Abstract
1. Introduction
2. Results
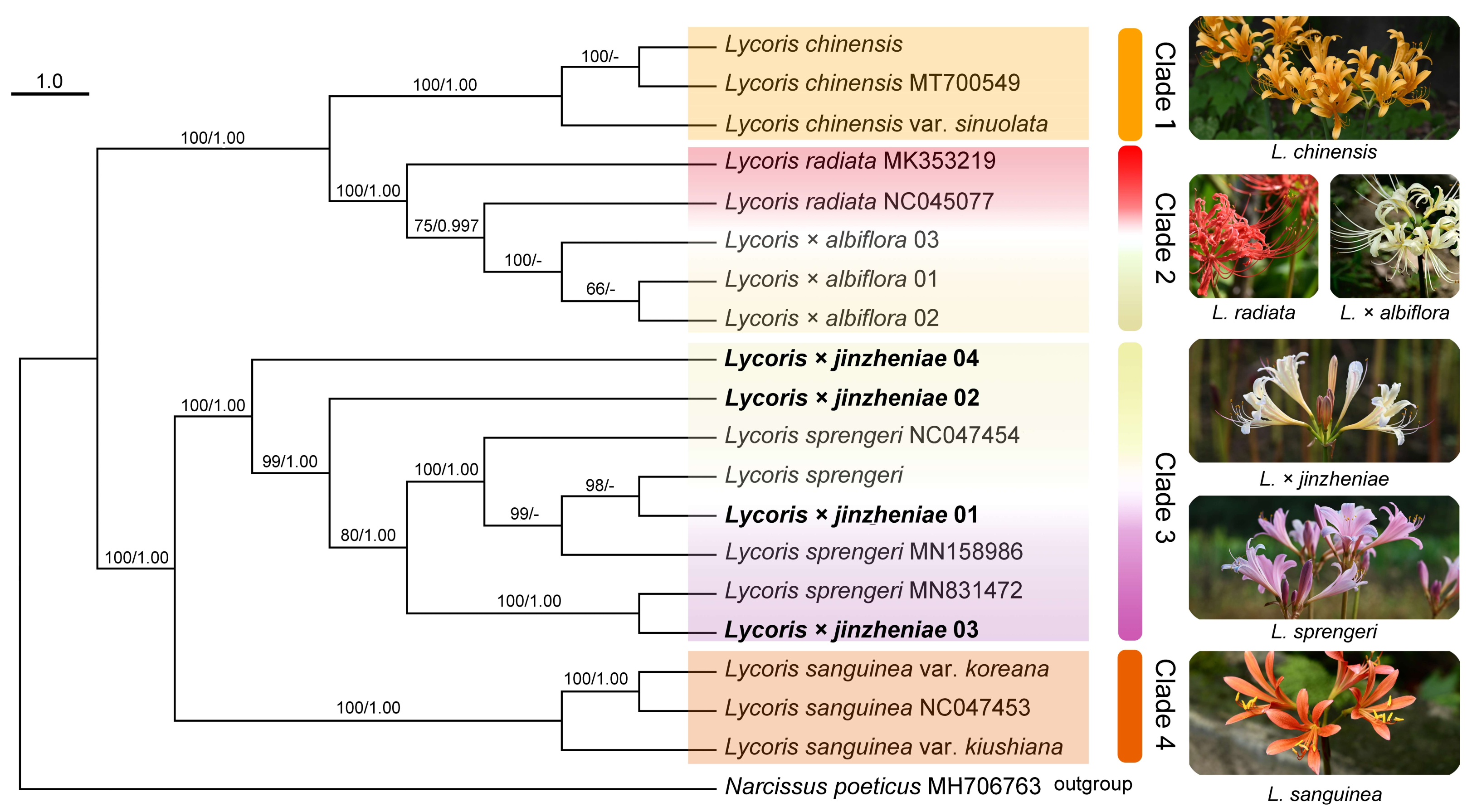
2.1. Molecular Phylogenetics
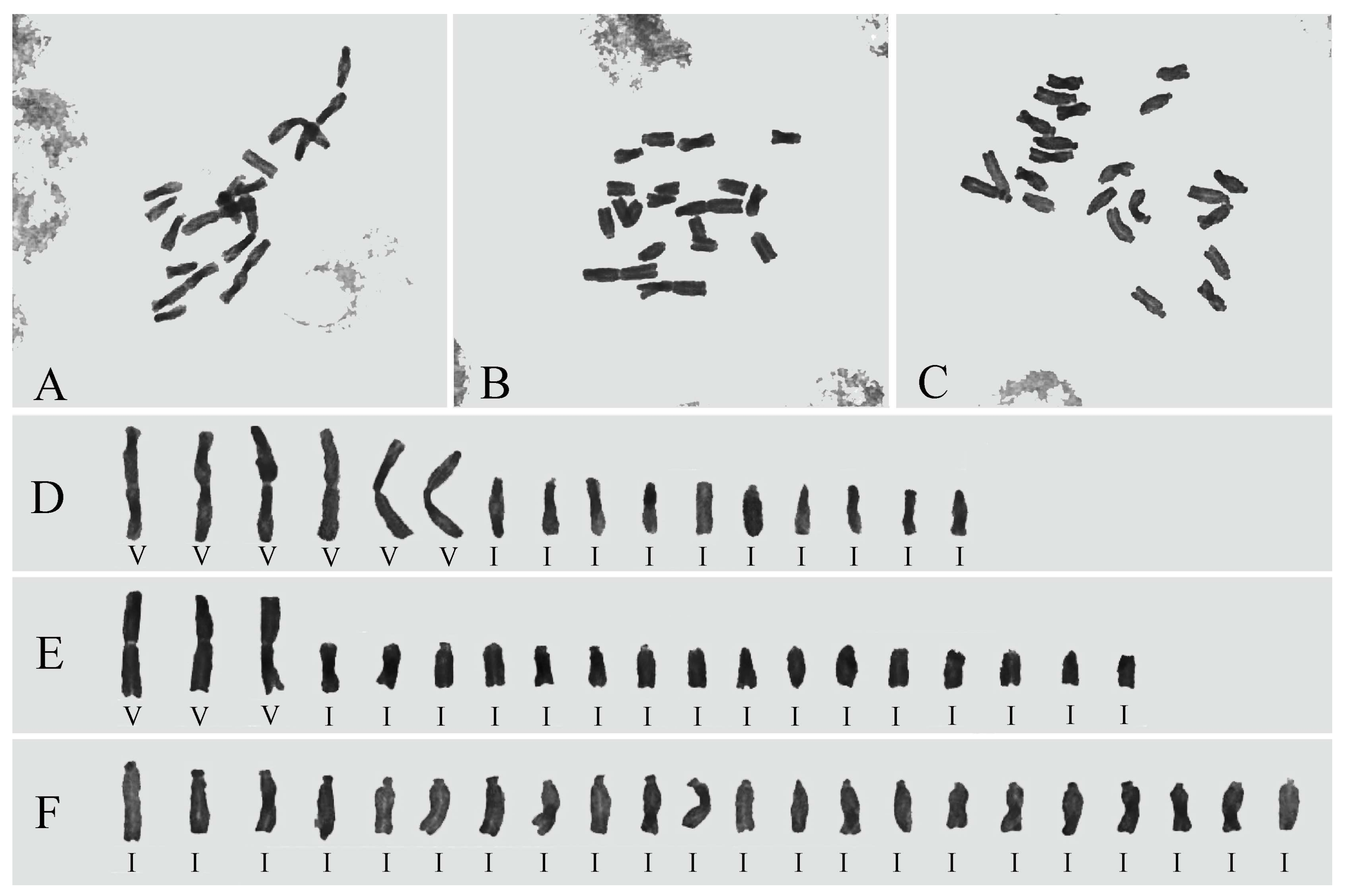
2.2. Karyotype
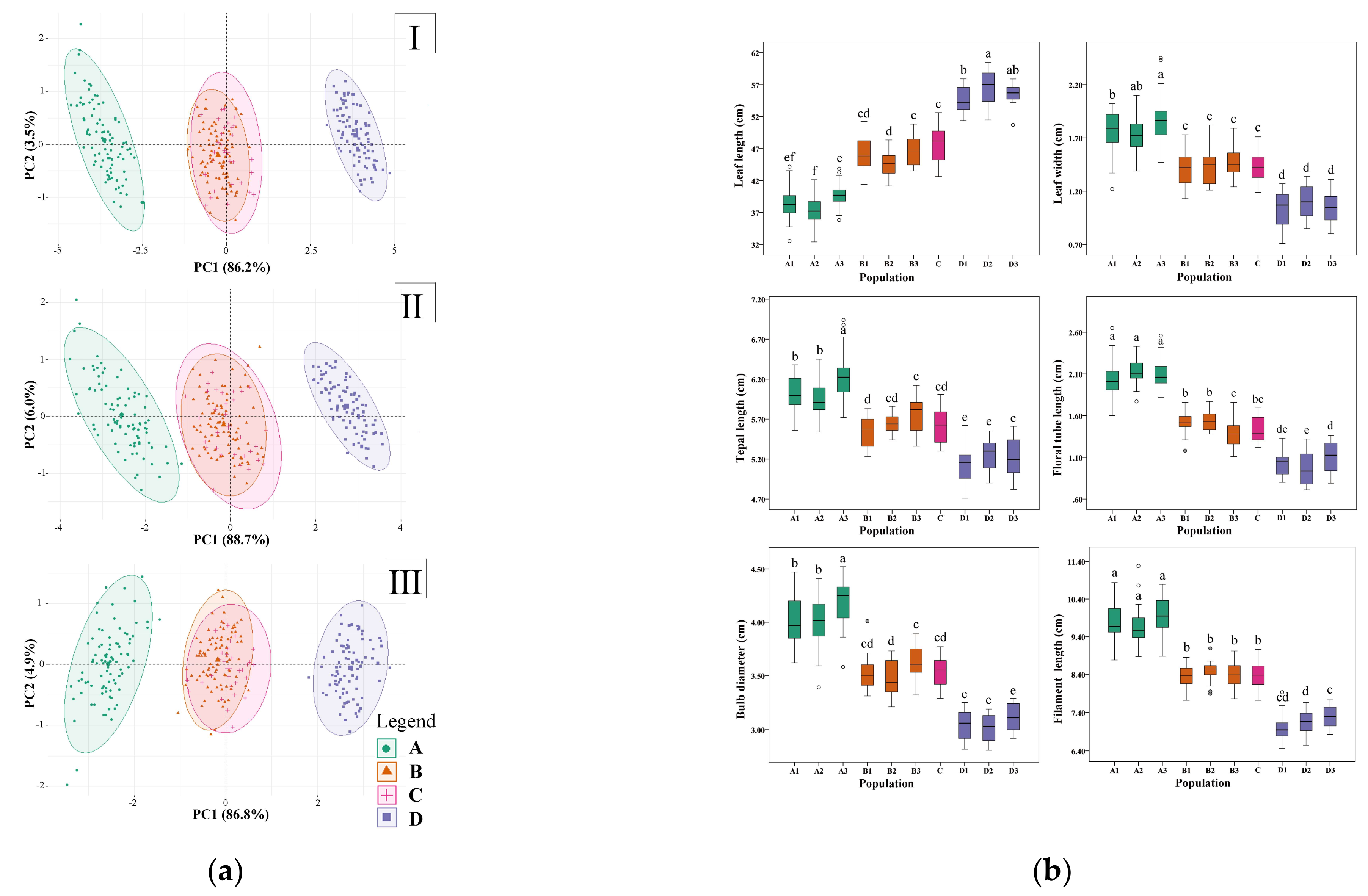
2.3. Morphological Characteristics and Fertility
2.4. Taxonomic Treatment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Samples and Code | Total |
|---|---|---|
| China, Jiangsu Province, Wuxi City, Yixing County, Shanjuan Cave | A1: L. chinensis *30, B1: L. × jinzheniae *30, D1: L. sprengeri *30 | 300 |
| China, Zhejiang Province, Cixi City, Longshan Town, Dapon Mountain | A2: L. chinensis *30, B2: L. × jinzheniae *30, D2: L. sprengeri *30 | |
| China, Anhui Province, Chuzhou City, Langya County, Langya Mountain | A3: L. chinensis *30, B3: L. × jinzheniae *30, D3: L. sprengeri *30 | |
| China, Zhejiang Province, Hangzhou City, Hangzhou Botanical Garden | C: L. × jinzheniae (artificial hybridization) *30 |
3. Discussion
4. Materials and Methods
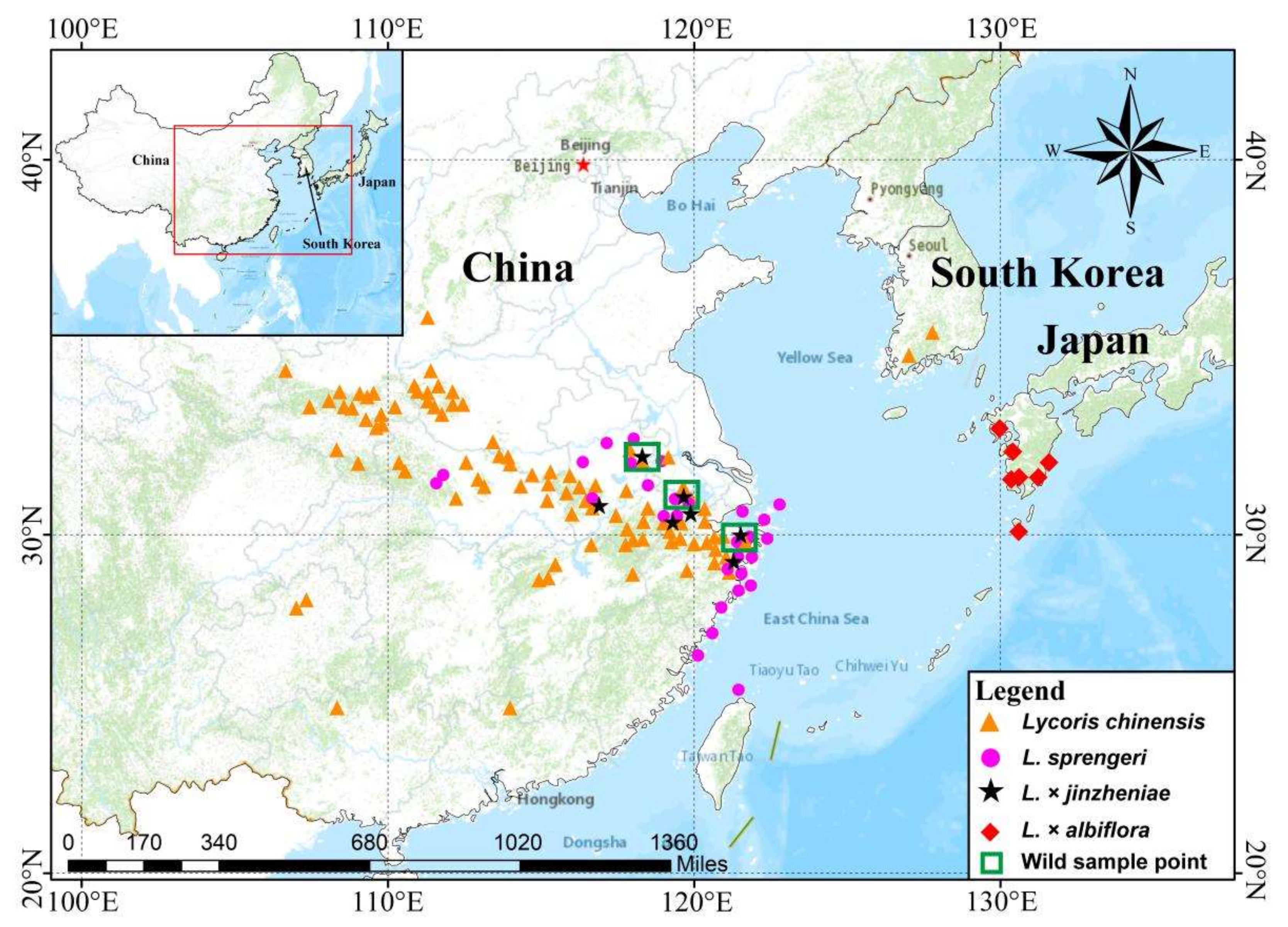
4.1. Plant Material
4.2. Chloroplast Genome Acquisition and Phylogenetic Analysis
4.3. Karyotype Observation
4.4. Morphological Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hsu, P.S.; Kurita, S.; Yu, Z.Z.; Lin, J.Z. Synopsis of the genus Lycoris. SIDA Contrib. Bot. 1994, 16, 301–331. [Google Scholar]
- Kim, M.Y. A taxonomic review of Korean Lycoris (Amaryllidaceae). Korean J. Plant Taxon. 2004, 34, 9–26. [Google Scholar] [CrossRef]
- Hori, T.A.; Hayashi, A.; Sasanuma, T.; Kurita, S. Genetic variations in the chloroplast genome and phylogenetic clustering of Lycoris species. Genes Genet. Syst. 2006, 81, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.D.; Qiu, Y.X.; Li, E.X.; Wu, L.; Fu, C.X. Phylogenetic Relationships and Possible Hybrid Origin of Lycoris Species (Amaryllidaceae) Revealed by ITS Sequences. Biochem. Genet. 2006, 44, 198–208. [Google Scholar] [CrossRef]
- Shi, S.D.; Qiu, Y.X.; Wu, L.; Fu, C.X. Interspecific relationships of Lycoris (Amaryllidaceae) inferred from inter-simple sequence repeat data. Sci. Hortic. 2006, 110, 285–291. [Google Scholar] [CrossRef]
- Yoo, Y.K.; Yuan, T.; Lee, J.S.; Lee, A.K.; Roh, M.S.; Kurita, S.; Suh, J.K. Species relationships of Lycoris endemic to Korea evaluated by RAPD and SNPs of nrDNA-ITS regions. Hortic. Environ. Biotechnol. 2011, 52, 145. [Google Scholar] [CrossRef]
- Quan, M.H.; Ou, L.J.; She, C.W. A New Species of Lycoris (Amaryllidaceae) from Hunan, China. Novon 2013, 22, 307–310. [Google Scholar] [CrossRef]
- Meng, W.Q.; Zheng, L.; Shao, J.W.; Zhou, S.B.; Liu, K. A new natural allotriploid, Lycoris × hubeiensis hybr. nov. (Amaryllidaceae), identified by morphological, karyological andmolecular data. Nord. J. Bot. 2018, 36, e01780. [Google Scholar] [CrossRef]
- Lu, J.Y.; Wang, T.; Wang, Y.C.; Zhang, P.C. Lycoris tsinlingensis (Amaryllidaceae), a new species from shaanxi, China. Ann. Bot. Fenn. 2020, 57, 193–196. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Huang, Y.; Zhang, P.; Chen, Y.B.; Shao, J.W. Lycoris wulingensis, a dwarf new species of Amaryllidaceae from Hunan, China. PhytoKeys 2021, 177, 1–9. [Google Scholar] [CrossRef]
- Kurita, S. Variation and Evolution in the Karyotype of Lycoris, Amaryllidaceae I. General Karyomorphological Characteristics of the Genus. Cytologia 1986, 51, 803–815. [Google Scholar] [CrossRef]
- Kurita, S. Variation and evolution on the karyotype of Lycoris, Amaryllidaceae II. Karyotype analysis of ten taxa among which seven are native to China. Cytologia 1987, 52, 19–40. [Google Scholar] [CrossRef]
- Rieseberg, L.H. Hybrid origins of plant species. Annu. Rev. Ecol. Evol. Syst. 1997, 28, 359–389. [Google Scholar] [CrossRef]
- Schumer, M.; Rosenthal, G.G.; Andolfatto, P. How common is homoploid hybrid speciation? Evolution 2014, 68, 1553–1560. [Google Scholar] [CrossRef]
- Turland, N.J.; Wiersema, J.H.; Barrie, F.R.; Greuter, W.; Hawksworth, D.L.; Herendeen, P.S.; Knapp, S.; Kusber, W.H.; Li, D.Z.; Marhold, K.; et al. (Eds.) Names of Hybirds. In International Code of Nomenclature for Algae, Fungi, and Plants (Shenzhen Code), Proceedings of the Nineteenth International Botanical Congress, Shenzhen, China, 23–29 July 2017; Koeltz Botanical Books: Glashütten, Germany, 2018; Volume H, pp. H1–H12. [Google Scholar] [CrossRef]
- Xu, Y.; Hu, B.Z.; Huang, X.L.; Fan, G.J. Amaryllidaceae. In Flora Reipublicae Popularis Sinicae; Pei, J., Ding, Z.Z., Eds.; Science Press: Beijing, China, 1985; Volume 16, pp. 22–24. [Google Scholar]
- Koidzumi, G. Contributiones ad Cognitionem Floræ Asiæ Orientalis (continued from Vol. XXXVII p. 59). Bot. Mag. 1924, 449, 100. [Google Scholar] [CrossRef][Green Version]
- Lin, J.Z.; Yu, Z.Z.; Xu, B.S. Hybridization and breeding of Lycoris. In Proceedings of the International Symposium Botanical, Nanjin, China, 25–28 September 1988. [Google Scholar]
- IUCN. Guidelines for Using the IUCN Red List Categories and Criteria. Version 15. Prepared by the Standards and Petitions Committee. 2022. Available online: https://www.iucnredlist.org/resources/redlistguidelines (accessed on 7 June 2022).
- Liu, K.; Sun, L.; Meng, W.Q.; Zhu, H.; Zhang, D.; Wang, J.X. Comparative genomics and phylogenetic perspectives of sixfertile Lycoris species endemic to East Asia based on plastome characterization. Nord. J. Bot. 2022, 2022, e03412. [Google Scholar] [CrossRef]
- Tiffin, P.; Olson, S.; Moyle, L.C. Asymmetrical crossing barriers in angiosperms. Proc. R. Soc. Lond. B 2001, 268, 861–867. [Google Scholar] [CrossRef]
- Zhou, R.C.; Gong, X.; Boufford, D.; Wu, C.I.; Shi, S.H. Testing a hypothesis of unidirectional hybridization in plants: Observations on Sonneratia, Bruguiera and Ligularia. BMC Evol. Biol. 2008, 8, 149. [Google Scholar] [CrossRef]
- Ma, Y.P.; Xie, W.J.; Tian, X.L.; Sun, W.B.; Wu, Z.K.; Milne, R. Unidirectional hybridization and reproductive barriers between two heterostylous primrose species in north-west Yunnan, China. Ann. Bot. 2014, 113, 763–775. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar] [CrossRef]
- Larridon, I.; Walter, H.E.; Guerrero, P.C.; Duarte, M.; Cisternas, M.A.; Hernández, C.P.; Bauters, K.; Asselman, P.; Goetghebeur, P.; Samain, M. An integrative approach to understanding the evolution and diversity of Copiapoa (Cactaceae), a threatened endemic Chilean genus from the Atacama Desert. Am. J. Bot. 2015, 102, 1506–1520. [Google Scholar] [CrossRef]
- Jin, J.J.; Yu, W.B.; Yang, J.B.; Song, Y.; Claude, W.; Yi, T.S.; Li, D.Z. GetOrganelle: A fast and versatile toolkit for accurate de novo assembly of organelle genomes. Genome Biol. 2020, 21, 241. [Google Scholar] [CrossRef]
- Qu, X.J.; Moore, M.J.; Li, D.Z.; Yi, T.S. PGA: A software package for rapid, accurate, and flexible batch annotation of plastomes. Plant Methods 2019, 15, 50. [Google Scholar] [CrossRef]
- Könyves, K.; Bilsborrow, J.; David, J.; Culham, A. The complete chloroplast genome of Narcissus poeticus L. (Amaryllidaceae: Amaryllidoideae). Mitochondrial DNA Part B 2018, 3, 1137–1138. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.J.; Shu, X.C.; Wang, T.; Zhuang, W.B.; Wang, Z. The complete chloroplast genome sequence of Lycoris radiata. Mitochondrial DNA Part B 2019, 4, 2886–2887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.J.; Zhuang, W.B.; Shu, X.C.; Wang, T.; Wang, Z. Complete chloroplast genome of Lycoris sprengeri (Amaryllidaceae) and genetic comparison. Mitochondrial DNA Part B 2019, 4, 3782–3783. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.J.; Wang, T.; Shu, X.C.; Wang, N.; Zhuang, W.B.; Wang, Z. Complete Chloroplast Genomes and Comparative Analyses of L. chinensis, L. anhuiensis, and L. aurea (Amaryllidaceae). Int. J. Mol. Sci. 2020, 21, 5729. [Google Scholar] [CrossRef] [PubMed]
- Ranwez, V.; Douzery, E.J.P.; Cambon, C.; Chantret, N.; Delsuc, F. MACSE v2: Toolkit for the Alignment of Coding Sequences Accounting for Frameshifts and Stop Codons. Mol. Biol. Evol. 2018, 35, 2582–2584. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseleret, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- He, X.; Cao, J.J.; Zhang, W.; Li, Y.Q.; Zhang, C.; Li, X.H.; Xia, G.H.; Shao, J.W. Integrative taxonomy of herbaceous plants with narrow fragmented distributions: A case study on Primula merrilliana species complex. J. Syst. Evol. 2021. accepted. [Google Scholar] [CrossRef]
- Chen, Y.H.; Li, M.X. Karytype Analyses of four species (varieties) of Lycoris Herb. Acta Hortic. Sin. 1985, 12, 57–60. [Google Scholar]
- Levan, L.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]






| Characters | L. × jinzheniae | L. chinensis | L. sprengeri | L. × albiflora |
|---|---|---|---|---|
| Emerging period of leaf | Spring | Spring | Spring | Autumn |
| Degree of leaf twist | Slight | No | Strong | No |
| Corolla color | White, sometimes pale pink or yellowish | Yellow | Pink-blue | Milky white or yellowish |
| Degree of tepal undulation | Slight | Strong | No | Strong |
| Degree of tepal recurve | Slight | Strong | Slight | Strong |
| Type of flower symmetry | Central or bilateral symmetry | Bilateral symmetry | Centrosymmetric | Bilateral symmetry |
| Approximate ratio of filament to corolla length | 1.2:1 | 1.3:1 | 1.2:1 | 2:1 |
| Fertility | Infertile | Fertile | Fertile | Infertile |
| Karyotype | 2n = 3V + 16I | 2n = 6V + 10I | 2n = 22I | 2n = 5V + 12I/6V + 12I |
| Distribution | Eastern China | China and South Korea | Eastern and Central China | Japan |
| Individual | Locality | GenBank acc. no |
|---|---|---|
| L. × jinzheniae 1 | China, Jiangsu Province, Wuxi City, Yixing County, Shanjuan Cave | ON611628 |
| L. × jinzheniae 2 | China, Zhejiang Province, Cixi City, Longshan Town, Dapeng Mountain | ON611629 |
| L. × jinzheniae 3 | ★China, Zhejiang Province, Hangzhou City, Hangzhou Botanical Garden | ON611630 |
| L. × jinzheniae 4 | ★China, Zhejiang Province, Hangzhou City, Hangzhou Botanical Garden | ON611631 |
| L. sprengeri | China, Jiangsu Province, Wuxi City, Yixing County, Shanjuan Cave | ON611639 |
| L. sanguinea var. kiushinan | Japan, Kyushu Island, Kagoshima Prefecture, Kirishima City | ON611637 |
| L. sanguinea var. koreana | South Korea, Jeju Special Self-Governing Province, Hallim-eup | ON611638 |
| L. chinensis | China, Jiangsu Province, Wuxi City, Yixing County, Shanjuan Cave | ON611636 |
| L. chinensis var. sinulata | South Korea, Jeollanam-do, Gangjin-gun | ON611635 |
| L. × albiflora 1 | Japan, Kyushu Region, Kagoshima Prefecture, Kirishima City | ON611632 |
| L. × albiflora 2 | Japan, Kyushu Region, Kagoshima Prefecture, Amami City | ON611633 |
| L. × albiflora 3 | Japan, Honshu Region, Hyogo Prefecture, Akashi City | ON611634 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.-Y.; Hu, Y.-F.; Wang, H.-T.; Zhang, P.-C.; Shao, J.-W. Over 30 Years of Misidentification: A New Nothospecies Lycoris × jinzheniae (Amaryllidaceae) in Eastern China, Based on Molecular, Morphological, and Karyotypic Evidence. Plants 2022, 11, 1730. https://doi.org/10.3390/plants11131730
Zhang S-Y, Hu Y-F, Wang H-T, Zhang P-C, Shao J-W. Over 30 Years of Misidentification: A New Nothospecies Lycoris × jinzheniae (Amaryllidaceae) in Eastern China, Based on Molecular, Morphological, and Karyotypic Evidence. Plants. 2022; 11(13):1730. https://doi.org/10.3390/plants11131730
Chicago/Turabian StyleZhang, Si-Yu, Ying-Feng Hu, Hao-Tian Wang, Peng-Chong Zhang, and Jian-Wen Shao. 2022. "Over 30 Years of Misidentification: A New Nothospecies Lycoris × jinzheniae (Amaryllidaceae) in Eastern China, Based on Molecular, Morphological, and Karyotypic Evidence" Plants 11, no. 13: 1730. https://doi.org/10.3390/plants11131730
APA StyleZhang, S.-Y., Hu, Y.-F., Wang, H.-T., Zhang, P.-C., & Shao, J.-W. (2022). Over 30 Years of Misidentification: A New Nothospecies Lycoris × jinzheniae (Amaryllidaceae) in Eastern China, Based on Molecular, Morphological, and Karyotypic Evidence. Plants, 11(13), 1730. https://doi.org/10.3390/plants11131730