
Biochemical and Physiological Changes during Early Adventitious Root Formation in Chrysanthemum indicum Linné Cuttings





Abstract
1. Introduction
2. Results
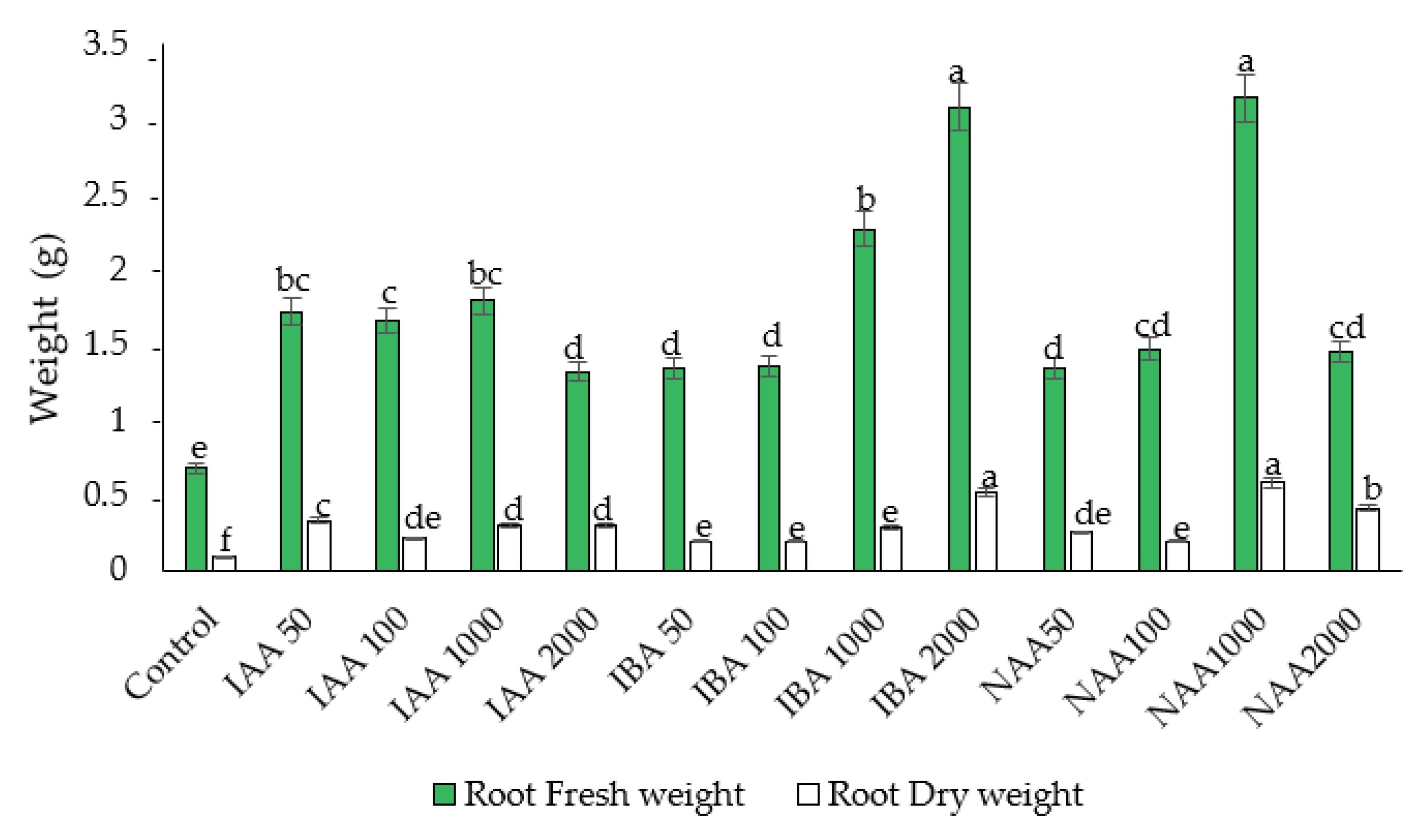
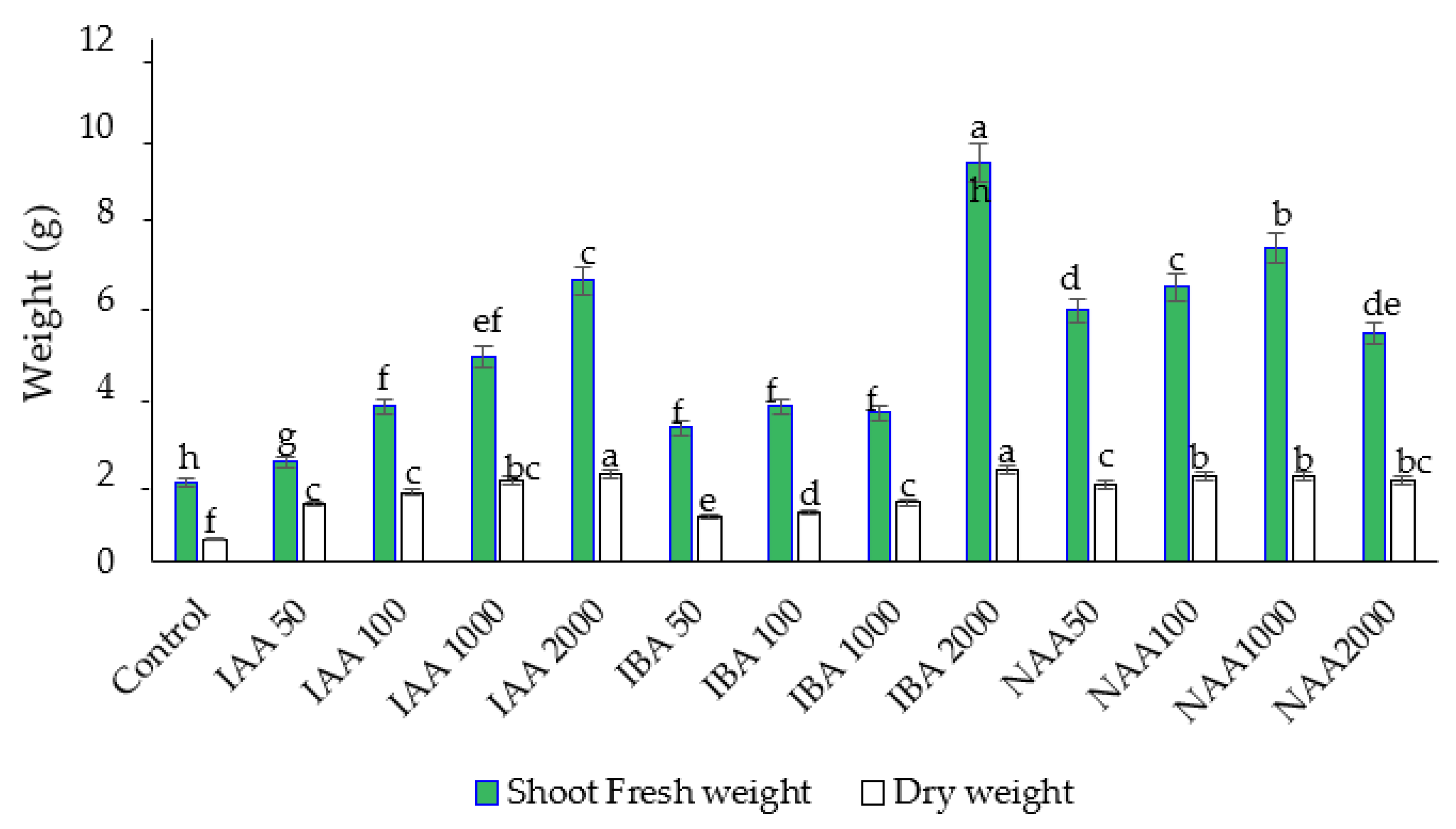
2.1. Effect of Auxins on Fresh Weight (FW) and Dry Weight (DW) of Roots and Shoots
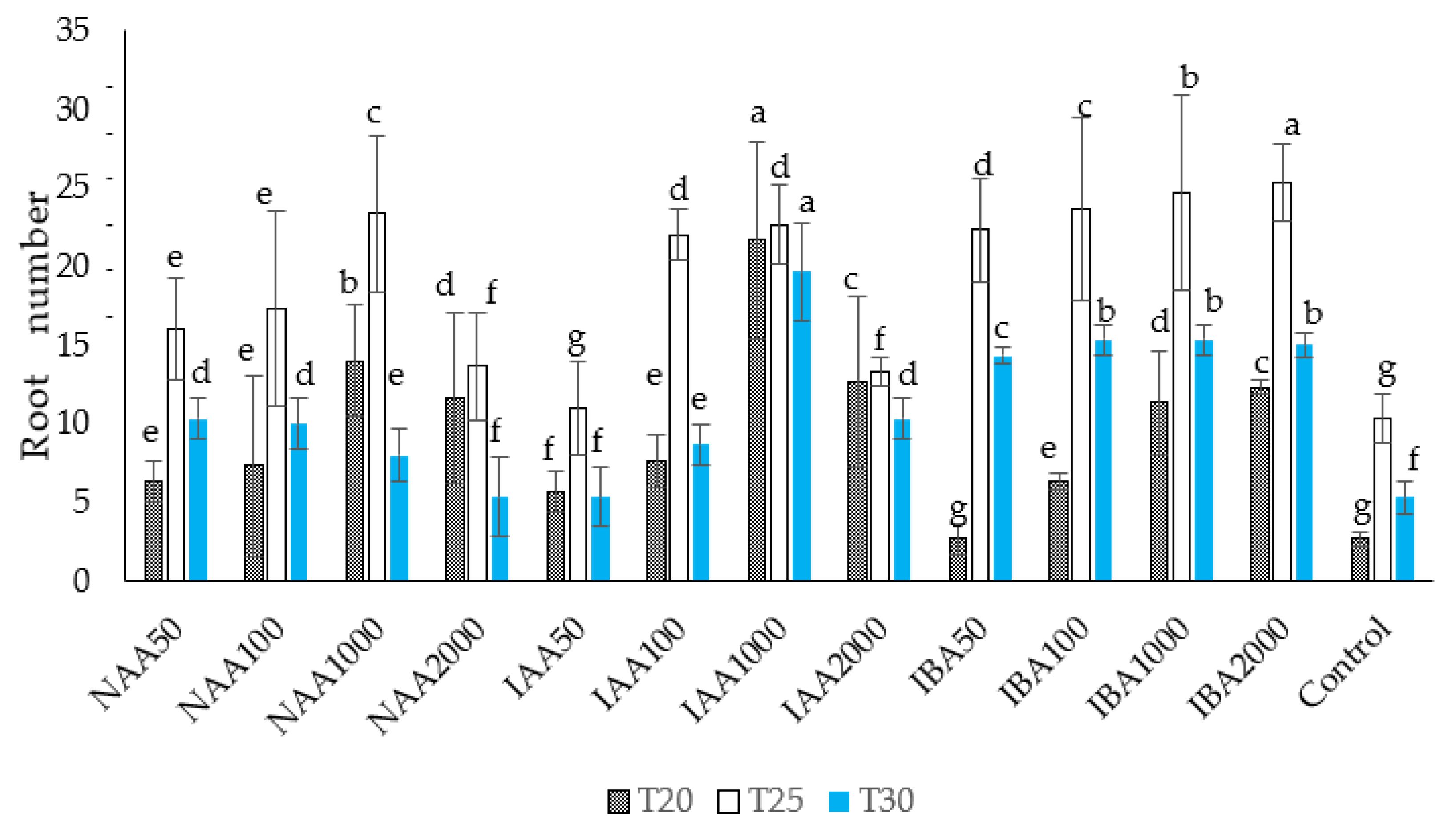
2.2. Effect of Temperature on Adventitious Root Number and Root Length in C. indicum
2.3. Influence of Explant Position and Soil Types on Root Length, Bud Length, and Bud Number
2.4. Influence of Growth Regulators on Chlorophyll Content
2.5. Effect of Auxin Types and Concentration on Total Phenol Content (TPC) and Total Flavonoid Content (TFC)
2.6. Phenolic Compounds
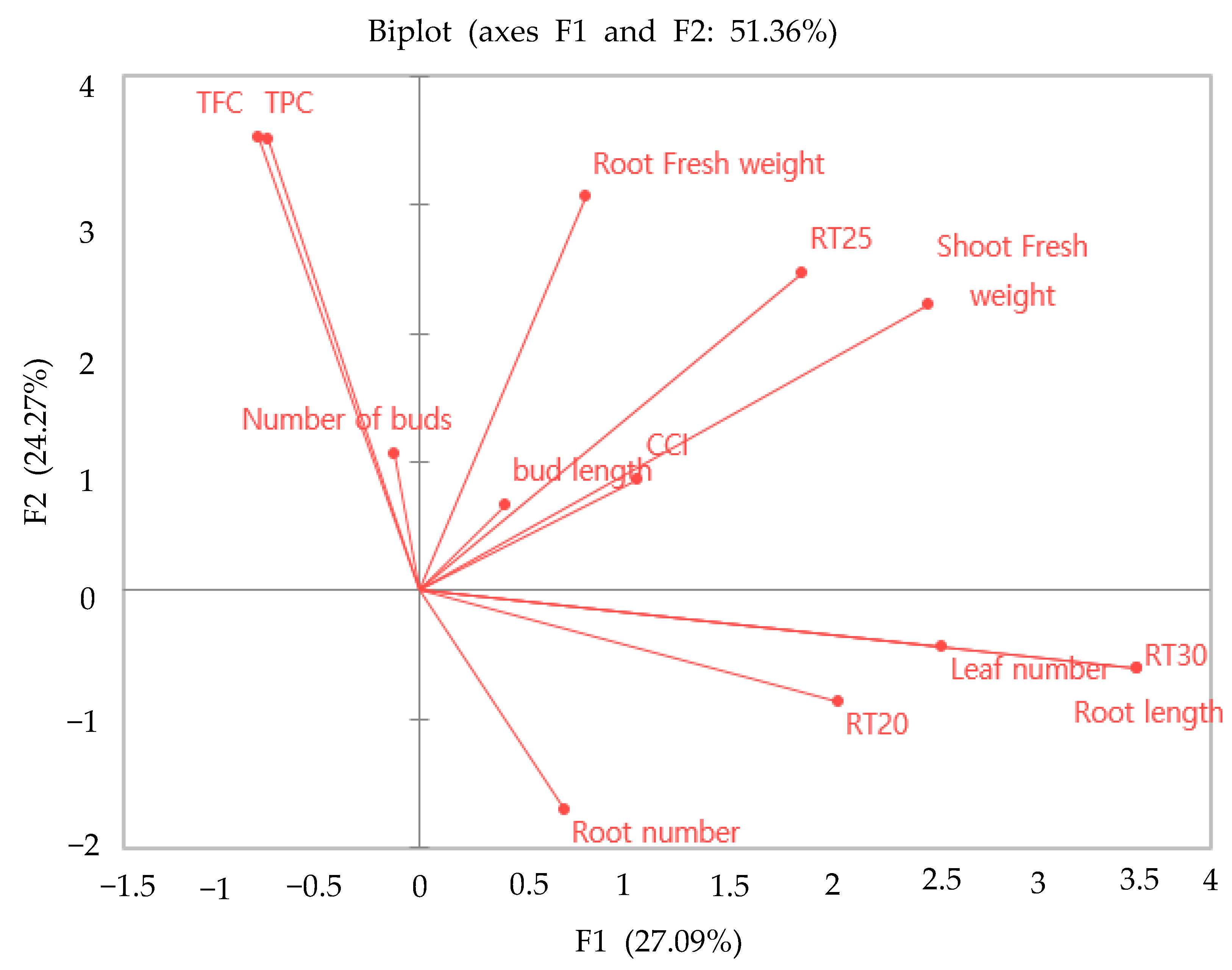
2.7. Principal Component Analysis (PCA) Root System Parameters in C. indicum.
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Experimental Site and Plant Material
4.3. Stem Cutting Preparation and Applying Growth Regulator
4.4. Experimental Design
4.4.1. Sample Collection and Extract Preparation
4.4.2. Determining Total Phenolic Acid Content (TPC)
4.4.3. Determining Total Flavonoid Content (TFC)
4.4.4. Estimation of Phenolic Compounds
4.4.5. Statistical Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuk, C.S. Original Colors Pictorial Book of the Korean Medicinal Use; Academy Books: Seoul, Korea, 2006; p. 537. [Google Scholar]
- Yoshikawa, M.; Morikawa, T.; Toguchida, I.; Harima, S.; Matsuda, H. Inhibitors of nitric oxide production and absolute stereo structures of five new germacrane-type sesquiterpenes, kikkanols D, D monoacetate, E, F, and F mono acetate from the flowers of Chrysanthemum indicum L. Chem. Pharm. Bull. 2000, 48, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Kim, G.H. Volatile Flavor Composition of Gamguk (Chrysanthemum indicum) Flower Essential Oils. Food Sci. Biotechnol. 2011, 20, 319–325. [Google Scholar] [CrossRef]
- Fang, H.L.; Guo, Q.S.; Shen, H.J.; Li, Y.C. Genetic diversity evaluation of Chrysanthemum indicum L. by medicinal compounds and molecular biology tools. Biochem. Syst. Ecol. 2012, 41, 26–34. [Google Scholar] [CrossRef]
- Matsuda, H.; Morikawa, T.; Toguchida, I.; Harima, S.; Yoshikawa, M. Medicinal flowers VI. Absolute stereostructures of two new flavanone glycosides and phenylbutanoid glycoside from the flowers of Chrysanthemum indicum L., there inhibitory activities for rat lens aldose reductase. Chem. Pharm. Bull. 2002, 50, 972–975. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.J.; Jeon, J.R.; Park, G.S. Physicochemical properties of gamgug (Chrysanthemun indicum L.). J. Korean Soc. Food Sci. Nutr. 2004, 33, 146–151. [Google Scholar]
- Kim, H.G.; Kim, Y.E.; Do, J.R.; Lee, Y.C.; Lee, B.Y. Anti oxidative activity and physiological activity of some Korean medicinal plants. Korean J. Food Sci. Technol. 1995, 27, 80–85. [Google Scholar]
- Chang, K.M.; Choi, E.M.; Kim, G.H. Chemical Constituents of Chrysanthemum indicum L. Flower Oil and Effect on Osteoblastic MC3T3-E1 Cells. Food Sci. Biotechnol. 2010, 19, 815–819. [Google Scholar] [CrossRef]
- Jiangsu New Medical College. Dictionary of Chinese Material Medical Science and Technology; Press of Shanghai: Shanghai, China, 1993; pp. 2144–2145. [Google Scholar]
- Arokiyaraj, S.; Kumar, V.D.; Elakya, V.; Kamala, T.; Park, S.K.; Ragam, M.; Saravanan, M.; Bououdina, M.; Arasu, M.V.; Kovendan, K.; et al. Biosynthesized silver nanoparticles using floral extract of Chrysanthemum indicum L.—Potential for malaria vector control. Environ. Sci. Pollut. Res. 2015, 22, 9759–9765. [Google Scholar] [CrossRef]
- Utami, C.V.; Pitinidhipat, N.; Yasurin, P. Antibacterial Activity of Chrysanthemum indicum, Centella asiatica and Andrographis paniculata on Bacillus cereus and Listeria monocytogenes under Low pH Stress. KMITL Sci. Technol. J. 2012, 12, 49–54. [Google Scholar]
- Yan, Y.C.; Lou, X.E.; Jiang, H.D. Experimental studies on the anti-oxidation effects of water extract from Chrysanthemum indicum. Chin. Med. 2013, 8, 7. [Google Scholar]
- Liu, Y.G. Pharmacological study and clinical apply of Chrysanthemum indicum. Shi Zhen Guo Yao 1991, 2, 103. [Google Scholar]
- Ren, A.N.; Wang, Z.G.; Lu, Z.C.; Wang, L.W.; Wu, Y.L. Study on bacteriostasis and antivirotic of flowers Chrysanthemum indicum. Pharm. Biotechnol. 1999, 6, 241–244. [Google Scholar]
- Cheng, W.; Li, J.; You, T.; Hu, C. Anti-inflammatory and immunomodulatory activities of the extracts from the inflorescence of Chrysanthemum indicum Linne. J. Ethnopharmacol. 2005, 101, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Chun, H.S.; Kim, J.M.; Choi, E.H.; Chang, N. Neuroprotective effects of several Korean medicinal plants traditionally used for stroke remedy. J. Med. Food 2008, 11, 246–251. [Google Scholar] [CrossRef]
- Ukiya, M.; Akihisa, T.; Tokuda, H.; Suzuki, H.; Mukainaka, T.; Ichiishi, E.; Yasukawa, K.; Kasahara, Y.; Nishino, H. Constituents of Compositae plants III. Anti-tumor promoting effects and cytotoxic activity against human cancer cell lines of triterpene diols and trios from edible chrysanthemum flowers. Cancer Lett. 2002, 177, 7–12. [Google Scholar] [CrossRef]
- Wang, Z.G.; Ren, A.N.; Xu, L.; Sun, X.J.; Hua, X.B. The experimental study on the immunological and anti-inflammatory activities of Chrysanthemum indicum. Chin. J. Tradit. Med. Sci. Technol. 2000, 2, 92–93. [Google Scholar]
- Debnath, T.; Jin, H.L.; Hasnat, M.A.; Kim, Y.; Samad, N.B.; Park, P.J.; Lim, B.O. Antioxidant potential and oxidative DNA damage preventive activity of Chrysanthemum indicum. J. Food Biochem. 2013, 37, 440–448. [Google Scholar] [CrossRef]
- Lee, B.H.; Nam, T.G.; Park, W.J.; Kang, H.; Heo, H.J.; Chung, D.K.; Kim, G.H.; Kim, D.O. Antioxidative and Neuroprotective Effects of Volatile Components in Essential Oils from Chrysanthemum indicum Linné Flowers. Food Sci. Biotechnol. 2015, 24, 717–723. [Google Scholar] [CrossRef]
- Jeong, S.C.; Kim, S.M.; Jeong, Y.T.; Song, C.H. Hepatoprotective effect of water extract from Chrysanthemum indicum L. flower. Chin. Med. 2013, 8, 7. [Google Scholar] [CrossRef]
- Kato, T.; Noguchi, K.; Miyamoto, Y.; Suekawa, M.; Aburada, M.; Hosoya, E.; Sakanashi, M. Effects of Chrysanthemum indicum Linn. on coronary, vertebral, renal and aortic blood flows of the anesthetized dog. Arch Int. Pharmacodyn. Ther. 1987, 285, 288–300. [Google Scholar]
- Shunying, Z.; Yang, Y.; Huaidong, Y.; Yue, Y.; Guolin, Z. Chemical composition and antimicrobial activity of the essential oils of Chrysanthemum indicum. J. Ethnopharmacol. 2005, 96, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Somashekar, B.S.; Sharma, M. Propagation Techniques of Commercially Important Medicinal Plants; Training Manual; FRLHT: Bangalore, India, 2002; p. 118. [Google Scholar]
- Stefanello, R.; Garcia, D.C.; Menezes, N.L.; Wrasse, C.F. Influência da luz, temperatura e estresse hídrico na germinação de sementes de anis. Rev. Bras. De Agrociência 2006, 12, 45–50. [Google Scholar]
- Kikuti, A.L.P.; Marcos, F.J. Potencial fisiológico de sementes de couve-flor e desenvolvimento das plantas em campo. Rev. Bras. De Sementes 2007, 29, 107–113. [Google Scholar] [CrossRef]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T., Jr.; Geneve, R.L. Hartmann and Kester’s Plant Propagation: Principles and Practices, 7th ed.; Prentice Hall: London, UK, 2002. [Google Scholar]
- Blazich, F.A. Using calendar dates for cutting propagation. Amer. Nurserym. 1987, 165, 140–143. [Google Scholar]
- Halda, J.J.; Waddick, J.W. The Genus Paeonia; Timber Press: Portland, OR, USA, 2004; p. 187. [Google Scholar]
- Henrique, A.; Campinhos, E.N.; Ono, E.O.; de Pinho, S.Z. Effect of plant growth regulators in rooting of Pinus cuttings. Braz. Arch. Biol. Technol. 2006, 49, 189–196. [Google Scholar] [CrossRef]
- Heller, J. Physic Nut. Jatropha curcas-Promoting the Conservation and Use of Underutilized and Neglected Crops; Inst Plant Genet Crop.: Gatersleben, Germany; International plant Genetic Resources Institute: Rome, Italy, 1996. [Google Scholar]
- Gosh, L.; Singh, L. Study of factors influencing vegetative propagation of Jatropha curcas. Indian For. 2010, 136, 1637–1648. [Google Scholar]
- Stoltz, L.P. Factors influencing root initiation in an easy and a difficult-to-root Chrysanthemum. Proc. Am. Soc. Hortic. Sci. 1968, 92, 6. [Google Scholar]
- Vargas, G.; Arellano Ostoa, G.; Soto Hernández, R. Rooting of Icaco stilts (Chrysobalanus icaco L.) subjected to auxin applications. Bioagro 1999, 11, 103–108. [Google Scholar]
- Bhatt, B.B.; Tomar, Y.K. Effect of IBA on rooting performance of Citrus auriantifolia Swingle (Kagzi–lime) in different growing conditions. Nat. Sci. 2010, 8, 8–11. [Google Scholar]
- Zheng, L.; Xiao, Z.; Song, W. Effects of Substrate and Exogenous Auxin on the Adventitious Rooting of Dianthus caryophyllus L. Hortscience 2020, 55, 170–173. [Google Scholar] [CrossRef]
- Woodward, A.W.; Bartel, B.I. Auxin: Regulation, Action, and Interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed]
- Cline, M.G. Exucation of the auxin replacement apical dominance experiment in temperate woody species. Americon J. Bot. 2000, 87, 182–190. [Google Scholar] [CrossRef]
- Al-Saqri, F.; Alderson, P.G. Effects of IBA, cuttings type and rooting media on rooting of Rosa centifolia. J. Hortic. Sci. 1996, 71, 729–737. [Google Scholar] [CrossRef]
- Owuor, B.; Musyimi, D.; Ocaido, M.; Asimwe, J. Vegetative propagation of the large sour plum (Ximenia caffra Sond) by rooting of plagiotropic stem cuttings. ARPN J. Agric. Biol. Sci. 2009, 4, 19–25. [Google Scholar]
- Hartmenn, H.T.; Kestar, D.E.; Davies, F.T., Jr. Plant Propagation: Principles and Practices, 5th ed.; Prentice-Hall: Englewood Cliff, NJ, USA, 1990. [Google Scholar]
- Abu Leila, B.H. Studies on the Effect of Growth Regulators on Vegetative Growth and Pink Flowers of Chrysanthemum. Master’s Thesis, Department of Horticulture, College of Agriculture, University of Cairo, Cairo, Egypt, 1969. [Google Scholar]
- Kochhar, S.; Singh, S.P.; Kochhar, V.K. Effect of auxins and associated biochemical changes during clonal propagation of the biofuel plant—Jatropha curcas. Biomass Bioenergy 2008, 32, 1136–1143. [Google Scholar] [CrossRef]
- Gehlot, A.; Gupta, R.K.; Tripathi, A.; Arya, I.D.; Arya, S. Vegetative propagation of Azadirachta indica: Effect of auxin and rooting media on adventitious root induction in mini-cuttings. Adv. For. Sci. Cuiabá 2014, 1, 1–9. [Google Scholar]
- Anuradha, K.; Sreenivasan, M.S. Studies on rooting ability of Couvery (Catimor) cuttings. J. Coffee Res. 1993, 23, 55–58. [Google Scholar]
- Jaleel, C.A.; Riadh, K.; Gopi, R.; Manivannan, P.; Ine′s, J.; Al-Juburi, H.; Zhao, C.X.; Shao, H.B.; Panneerselvam, R. Antioxidant defense responses: Physiological plasticity in higher plants under abiotic constraints. Acta Physiol. Plant 2009, 31, 427–436. [Google Scholar] [CrossRef]
- Tamagnone, L.; Merida, A.; Stacey, N.; Plaskitt, K.; Parr, A.; Chang, C.F.; Lynn, D.; Dow, J.M.; Roberts, K.; Martin, C. Inhibition of phenolic acid metabolism results in precocious cell death and altered cell morphology in leaves of transgenic tobacco plants. Plant Cell 1998, 10, 1801–1816. [Google Scholar] [CrossRef]
- Murphy, A.; Peer, W.A.; Taiz, L. Regulation of auxin transport by aminopeptidases and endogenous flavonoids. Planta 2000, 211, 315–324. [Google Scholar] [CrossRef]
- Peer, W.A.; Murphy, A.S. Flavonoids and auxin transport: Modulators or regulators? Trends Plant Sci. 2007, 12, 556–563. [Google Scholar] [CrossRef] [PubMed]
- Bandurski, R.S.; Cohen, J.D.; Slovin, J.P.; Reinecke, D.M. Auxin biosynthesis and metabolism. In Plant Hormones; Davies, P.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1995; pp. 39–65. [Google Scholar]
- Wilson, P.J.; Van Staden, J. Rhizocaline, rooting co-factors, and the concept of promoters and inhibitors of adventitious rooting—A review. Ann Bot. 1990, 66, 479–490. [Google Scholar] [CrossRef]
- Haffner, V.; Enjalricf, F.; Lardetl, L.; Carron, M.P. Maturation of woody plants: A review of metabolic and genomic aspects. Ann. Sei. For. 1991, 48, 615–630. [Google Scholar] [CrossRef]
- Curir, P.; Sulis, S.; Mariani, F.; Van Sumere, C.F.; Marchesini, A.; Dolci, M. Influence of endogenous phenols on rootability of Chamaelaucium uncinatum Schauer stem cuttings. Sci. Hortic. 1993, 55, 303–314. [Google Scholar] [CrossRef]
- Bryant, P.H.; Trueman, S.J. Stem anatomy and adventitious root formation in cuttings of Angophora, Corymbia and Eucalyptus. Forests 2015, 6, 1227–1238. [Google Scholar] [CrossRef]
- Gyand, R.R. Effect of auxins on adventitious root development from single node cuttings of Camellia sinensis L. Kuntze and associated biochemical changes. Plant Growth Regul. 2006, 48, 111–117. [Google Scholar]
- Tchoundjeu, Z.; Ngo Mpeck, M.L.; Asaah, E.; Amougou, A. The role of vegetative propagation in the domestication of Pausinystalia johimbe (K. Schum), a highly threatened medicinal species of West and Central Africa. For. Ecol. Manag. 2004, 188, 175–183. [Google Scholar] [CrossRef]
- Negash, L. Successful vegetative propagation techniques for the threatened African pencil cedar (Juniperusprocera hoechst. ex Endl.). For. Ecol. Manag. 2002, 161, 53–64. [Google Scholar] [CrossRef]
- Amri, E.; Lyaruu, H.; Nyomora, A.S.; Kanyeka, Z.L. Vegetative propagation of African Blackwood (Dalbergia melanoxylon Guill. & Perr.): Effects of age of donor plant, IBA treatment and cutting position on rooting ability of stem cuttings. New For. 2010, 39, 183–194. [Google Scholar]
- Sauer, M.; Robert, S.; Kleine-Vehn, J. Auxin: Simply complicated. J. Exp. Bot. 2013, 64, 2565–2577. [Google Scholar] [CrossRef]
- Silva, J.M.; Raposo, A.; Sousa, J.A.; Miranda, E.M. Indução de enraizamento em estacas de joãobrandinho (Piper sp.) com ácido indolbutírico. Rev. Ciência Agronômica 2004, 35, 248–252. [Google Scholar]
- Grossmann, K.; Kwiatkowski, J.; Tresch, S. Auxin herbicides induce H2O2 overproduction and tissue damage in cleavers (Galium aparine L.). J. Exp. Bot. 2001, 52, 1811–1816. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, H.T.; Kester, D.E.; Davies, F.T.; Geneve, R.L. (Eds.) Plant Propagation: Principles and Practices, 8th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2011. [Google Scholar]
- Baul, T.K.; Mezbahuddin, M.; Mohiuddin, M. Vegetative propagation and initial growth performance of Stereospermum suaveolens DC., a wild tropical tree species of medicinal value. New For. 2009, 37, 275–283. [Google Scholar] [CrossRef]
- Lima, D.M.; Silva, C.L.; Ritter, M.; Biasi, L.A.; Zanette, F.; Zuffellato-Ribas, K.C. Substratos e auxinas no enraizamento de estacas caulinares de espinheira-santa. Sci. Agrar. Curitiba 2008, 9, 85–89. [Google Scholar] [CrossRef]
- Alcantara, G.B.; Oliveira, Y.; Lima, D.M.; Fleece, L.A.; Pinto, F.; Biasi, L.A. Efeito dos ácidos naftaleno acético e indolilbutírico no enraizamento de estacas de jambolão [Syzygium cumini (L.) Skeels] cuttings. Rev. Bras. Med. Plants 2010, 12, 317–321. [Google Scholar] [CrossRef]
- Azad, M.S.; Alam, M.J.; Mollick, A.S.; Matin, M.A. Responses of IBA on rooting, biomass production and survival of branch cuttings of Santalum album L., a wild threatened tropical medicinal tree species. J. Sci. Technol. Environ. Inform. 2016, 3, 195–206. [Google Scholar] [CrossRef]
- Kamaluddin, M. Clonal Forestry-Plus Tree Plantation Technology; Gono Unnon Library: Dhaka, Bangladesh, 1999. [Google Scholar]
- Yusnita, Y.; Jamaludin; Agustiansyah; Hapsoro, D. A Combination of IBA and NAA Resulted in Better Rooting and Shoot Sprouting than Single Auxin on Malay Apple [Syzygium malaccense (L.) Merr. & Perry] Stem Cuttings. Agrivita J. Agric. Sci. 2018, 40, 80–90. [Google Scholar]
- Puri, S.; Verma, R.C. Vegetative propagation of Dalbergia sissoo Roxb. Using softwood and hardwood stem cuttings. J. Arid Environ. 1996, 34, 235–245. [Google Scholar] [CrossRef]
- Thatoi, H.; Mishra, P.K.; Ousep, A.; Mohanty, J.R.; Acharjyo, L.N. Vegetative propagation by stem cuttings with auxins of four mangrove (and associate) species of Bhitarkanika, India. J. Trop. For. Sci. 2001, 13, 223–227. [Google Scholar]
- Titon, M.; Xavier, A.; Otoni, W.C.; Reis, G.G. Efeito do AIB no enraizamento de miniestacas e microestacas de clones de Eucalyptus grandis W. Hill Ex Maiden. Rev. Arv. 2003, 27, 1–7. [Google Scholar]
- Wendling, I.; Xavier, A. Influencia do’ acido indolbutírico e ds miniestaquia seriada no enraizamento e vigor de miniestacas de clones de Eucalyptus grandis. Rev. Arvore 2005, 29, 921–930. [Google Scholar] [CrossRef]
- Husen, A.; Pal, M. Metabolic changes during adventitious root primordium development in Tectona grandis Linn. f. (teak) cuttings as affected by age of donor plants and auxin (IBA and NAA) treatment. New For. 2007, 33, 309–323. [Google Scholar] [CrossRef]
- Altman, A.; Wareing, P.F. Effect of IAA on sugar accumulation and basipetal transport of C14-labeled assimilates in relation to root formation in Phaseolus vulgaris cuttings. Physiol. Plant 1975, 33, 32–38. [Google Scholar] [CrossRef]
- Hassig, B.E. Metabolic processes in adventitious rooting. In New Root Formation in Plants and Cuttings; Jackson, M.B., Ed.; Junk Publishers: Boston, MA, USA, 1986; pp. 141–148. [Google Scholar]
- Agullo-Anton, M.A.; Sanchez-Bravo, J.; Acosta, M.; Druege, U. Auxins or sugars: What makes the difference in the adventitious rooting of stored carnation cuttings? J. Plant Growth Regul. 2011, 30, 100–113. [Google Scholar] [CrossRef]
- Mesen, F.; Newton, A.C.; Leaky, R.R.B. Vegetative propagation of Cordial alliodora (Ruiz & Pavon) Oken: The effects of IBA concentration, propagation medium and cutting origin. For. Ecol. Manag. 1997, 92, 45–54. [Google Scholar]
- Husen, A.; Pal, M. Clonal propagation of Tectona grandis (Linn. f.): Effects of IBA and leaf area on carbohydrates drifts and adventitious root regeneration on branch cuttings. Ann. For. 2001, 9, 88–95. [Google Scholar]
- Orlikowska, T. Effect of amino acids on rooting of apple dwarf root stocks in vitro. Biol. Plant. 1992, 34, 39. [Google Scholar] [CrossRef]
- Sankhyan, H.P.; Sehgal, R.N.; Bhrot, N.P. Effect of growth regulators and seasons on the propagation of ifferent sea buckthorn species through cuttings in cold deserts of Himachal Pradesh. Ind. For. 2003, 129, 1300–1302. [Google Scholar]
- Oh, W.; Kim, K.S.; Kweon, Y.Y. Effects of air filled porosity of rooting media on rooting and growth of Chrysanthemum cuttings. J. Korean Soc. Hortic. Sci. 1998, 39, 92–97. [Google Scholar]
- Khayyat, M.; Nazari, F.; Salehi, H. Effects of different pot mixtures on Pothos (Epipremnun aureum Lindl. & Andre “Golden Pothos”) growth and development. Am.-Eurasian J. Agric. Environ. Sci. 2007, 6, 360–371. [Google Scholar]
- Otiende, M.A.; Fricke, K.; Nyabundi, J.O.; Ngamau, K.; Hajirezaei, M.R.; Druege, U. Involvement of the auxin–cytokinin homeostasis in adventitious root formation of rose cuttings as affected by their nodal position in the stock plant. Planta 2021, 254, 65. [Google Scholar] [CrossRef] [PubMed]
- Izhaki, A.; Yitzhak, Y.; Blau, T.; David, I.; Rotbaum, A.; Joseph Riov, J.; Zilkah, S. Rooting of cuttings of selected Diospyros virginiana clonal rootstocks and bud growth in rooted cuttings. Sci. Hortic. 2018, 232, 13–21. [Google Scholar] [CrossRef]
- Nicoloso, F.T.; Fortunato, R.P.; Fogaca, M.A.F. Influência da posição da estaca no ramo sobre o enraizamento de Pfaffia glomerata (Spreng.) Pedersen em dois substratos. Ciênc. Rural 1999, 29, 277–283. [Google Scholar] [CrossRef]
- Garbuio, C.; Biasi, L.A.; Kowalski, A.; Signor, D.; Machado, E.; Deschamps, C. Propagação por estaquia em patchouli com diferentes números de folhas e tipos de estacas. Sci. Agric. 2007, 8, 435–438. [Google Scholar] [CrossRef]
- Amaro, H.T.R.; Silveira, J.R.; David, A.M.S.; Resende, M.A.V.; Andrade, J.A.S. Tipos de estacas e substratos na propagação vegetativa da menta (Mentha arvensis L.). Rev. Bras. De Plantas Med. 2013, 15, 313–318. [Google Scholar] [CrossRef]
- Chagas, J.H.; Pinto, J.E.B.P.; Bertolucci, S.K.V.; Nalon, F.H. Produção de mudas de hortelã-japonesa em função da idade e de diferentes tipos de estacas. Ciênc. Rural 2008, 38, 2157–2163. [Google Scholar] [CrossRef]
- Dias JP, T.; Ono, E.O. Rooting of shoot cuttings derived of root cuttings of blackberry. Rev. Bras. De Frutic. 2011, 33, 649–653. [Google Scholar]
- Ljung, K.; Hull, A.K.; Celenza, J.; Yamada, M.; Estelle, M.; Normanly, J.; Sandberg, G. Sites and regulation of auxin biosynthesis in Arabidopsis roots. Plant Cell 2005, 17, 1090-104. [Google Scholar] [CrossRef]
- Saranga, J.; Cameron, R. Adventitious root formation in Anacardium occidentale L. in response to phytohormones and rem oval of roots. Sci. Hortic. 2007, 111, 164–172. [Google Scholar] [CrossRef]
- Kibbler, H.; Johnston, M.E.; Williams, R.R. Adventitious root formation in cuttings of Backhousia citriodora F. Muell. 1. Plant genotype, juvenility and characteristics of cuttings. Sci. Hortic. 2004, 102, 133–143. [Google Scholar] [CrossRef]
- Corrêa, L.R.; Fett-Neto, A.G. Effects of temperature on adventitious root development in microcuttings of Eucalyptus saligna Smith and Eucalyptus globulus Labill. J. Therm. Biol. 2004, 29, 315–324. [Google Scholar] [CrossRef]
- Garrido, G.; Ramon Guerrero, J.; Angel Cano, E. Origin and basipetal transport of the IAA responsible for rooting of carnation cuttings. Physiol. Plant. 2002, 114, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J. Influence of cutting position and temperature during rooting on adventitious root formation and axillary bud break of Stephenotis floribunda. Sci. Hortic. 1990, 40, 345–354. [Google Scholar] [CrossRef]
- Ling, W.X.; Jine, Q.; Zhong, Z. Effect of cutting medium temperatures on rooting process and root primordium differentiation of Hardwood cuttings of tetraploid Robinia pseudoacacia cutting medium temperatures of Robinia pseudoacacia. Pak. J. Bot. 2014, 46, 2209–2215. [Google Scholar]
- Costa, J.M.; Heuvelink, E.; Pol, P.A.; Put, H.M.C. Anatomy and morphology of rooting in leafy rose stem cuttings and starch dynamics following severance. Acta Hortic. 2007, 751, 495–502. [Google Scholar] [CrossRef]
- Kranner, I.; Zorn, M.; Turk, B.; Wornik, S.; Beckett, R.P.; Batič, F. Biochemical traits of lichens differing in relative desiccation tolerance. New Phytol. 2003, 160, 167–176. [Google Scholar] [CrossRef]
- Cristofori, V.; Rouphael, Y.; Rugini, E. Collection time, cutting age, IBA and putrescine effects on root formation in Corylus avellana L. cuttings. Sci. Hortic. 2010, 124, 189–194. [Google Scholar] [CrossRef]
- Fu, Z.; Xu, P.; He, S.; da Silva, J.A.; Tanaka, M. Dynamic changes in enzyme activities and phenolic content during in vitro rooting of tree peony (Paeonia suffruticosa Andr.) plantlets. Maejo Int. J. Sci. Technol. 2011, 5, 252–265. [Google Scholar]
- Osterc, G.; Štampar, F. Initial cutting length modifies polyphenol role in Castanea cuttings during the root formation process. Eur. J. Hortic. Sci. 2008, 73, 201–204. [Google Scholar]
- Krajnc, A.U.; Turinek, M.; Ivančič, A. Morphological and Physiological Changes during Adventitious Root Formation as Affected by Auxin Metabolism: Stimulatory Effect of Auxin Containing Seaweed Extract Treatment. Agricultura 2013, 10, 17–27. [Google Scholar]
- Osterc, G.; Trobec, M.; Usenik, V.; Solar, A.; Štampar, F. Changes in polyphenols in leafy cuttings during the root initiation phase regarding various cutting types at Castanea. Phyton 2004, 44, 109–119. [Google Scholar]
- Trobec, M.; Štampar, F.; Veberič, R.; Osterc, G. Fluctuations of different endogenous phenolic compounds and cinnamic acid in the rest days of the rooting process of cherry rootstock ‘Gisela 5′ leafy cuttings. J. Plant Physiol. 2005, 162, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Osterc, G.; Štefančič, M.; Solar, A.; Štampar, F. Potential involvement of avonoids in the rooting response of chestnut hybrid (Castanea crenata × Castanea sativa) clones. Aust. J. Exp. Agr. 2007, 47, 96–102. [Google Scholar] [CrossRef]
- Kroin, J. How to improve cutting propagation using water based indole-3-butyric acid rooting solutions. Comb. Proc.-Int. Plant Propagators Soc. 2011, 61, 381–391. [Google Scholar]
- Singleton, V.L.; Rossi, J.A., Jr. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Moreno, M.I.N.; Isla, M.I.; Sampietro, A.R.; Vattuone, M.A. Comparison of the free radical-scavenging activity of propolis from several regions of Argentina. J. Ethnopharmacol. 2000, 7, 109–114. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Yu, C.Y.; Chung, I.M. Assessment of the phenolic profile, antimicrobial activity and oxidative stability of transgenic Perilla frutescens L. overexpressing tocopherol methyltransferase (g-tmt) gene. Plant Physiol. Biochem. 2017, 118, 77–87. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Number of Roots | Root Length (cm) | Number of Leaves | Number of Buds | Bud Length (cm) | |
|---|---|---|---|---|---|---|
| Stem cutting position | Apical | 9.33 ± 0.47 c | 19.33 ± 1.25 a | 10.00 ± 1.63 a | 2.33 ± 0.47 c | 8.66 ± 0.94 b |
| Median | 9.00 ± 0.81 c | 16.00 ± 1.63 b | 8.33 ± 1.24 b | 5.33 ± 0.47 a | 4.33 ± 1.24 c | |
| Basal | 11.00 ± 0.82 b | 16.00 ± 0.82 b | 10.33 ± 1.25 a | 4.00 ± 0.82 b | 1.27 ± 0.17 d | |
| Substrates | V + P | 9.33 ± 0.47 c | 18.6 ± 1.63 a | 6.97 ± 2.49 c | 4.50 ± 1.63 b | 4.67 ± 2.49 c |
| Santro | 13.00 ± 2.16 a | 15.00 ± 0.82 b | 6.67 ± 1.89 c | 2.33 ± 0.47 c | 13.33 ± 3.39 a | |
| Sandy Loam Soil | 5.00 ± 1.10 d | 8.00 ± 1.20 c | 8.00 ± 1.50 b | 4.00 ± 0.06 b | 2.00 ± 0.02 d | |
| Characteristics | Number of Root | Root Length (cm) | Number of Leaves | Number of Buds | Bud Length (cm) |
|---|---|---|---|---|---|
| Leafy cutting | 12.00 ± 1.63 a | 20.00 ± 1.63 a | 6.67 ± 0.94 a | 3.00 ± 0.82 a | 8.00 ± 0.82 a |
| Non-leafy cutting | 9.50 ± 0.81 b | 18.00 ± 0.81 b | 5.33 ± 1.25 b | 2.67 ± 0.47 b | 2.33 ± 0.47 b |
| Treatment | GA | Pro | Gen | p-Hy | Chl | p-C | FA | Bio | Hom | Sal | Van | CA | Vit | Gly | Api | Lut | L-Phe | TPC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (μg/g) | ||||||||||||||||||
| Control | LOD | 162.2 | 9.2 | 26.1 | 5763 | LOQ | ND | 371.0 | LOD | 131.8 | ND | 57.7 | ND | LOQ | ND | LOQ | LOD | 6521 |
| NAA50 | LOQ | 525.0 | 24.7 | LOD | 336 | ND | ND | 137.3 | LOQ | 385.0 | ND | 119.7 | ND | 355.3 | 91.2 | 19.8 | LOD | 1994 |
| NAA 100 | LOQ | 464.0 | 28.2 | LOD | 2672 | ND | ND | 13.4 | LOD | 626.0 | ND | 53.2 | ND | LOQ | ND | 57.2 | LOD | 3914 |
| NAA 1000 | LOD | 375.4 | 18.8 | ND | 19,691 | ND | ND | 87.5 | LOQ | 287.6 | ND | 78.6 | ND | LOQ | ND | 74.9 | ND | 20,614 |
| NAA 2000 | LOQ | 338.3 | 16.8 | LOQ | 18,223 | ND | ND | 975.3 | LOQ | 278.8 | ND | 11.3 | ND | LOQ | LOD | 46.4 | LOD | 19,890 |
| 1AA50 | LOQ | 219.6 | 21.2 | 55.0 | 186 | LOQ | ND | 84.7 | ND | 246.5 | ND | 77.7 | ND | LOQ | ND | LOD | LOQ | 891 |
| 1AA100 | LOQ | 337.3 | 28.4 | 89.5 | 134 | ND | ND | 178.7 | LOQ | 614.7 | LOD | 47.3 | ND | LOQ | 27.7 | 37.2 | LOQ | 1494 |
| 1AA1000 | LOD | 375.3 | 23.6 | 74.1 | 20,095 | LOQ | ND | 544.7 | LOD | 443.3 | ND | 19.0 | ND | LOQ | 73.6 | 19.0 | LOD | 21,667 |
| 1AA2000 | LOD | 498.8 | 25.9 | 77.0 | 22,715 | LOQ | ND | 1288.0 | LOD | 149.5 | ND | 134.9 | ND | LOQ | LOQ | 58.3 | LOQ | 24,947 |
| 1BA50 | 75.3 | 175.5 | 23.4 | LOQ | 2678 | ND | ND | 1364.8 | LOQ | 459.2 | ND | 12.1 | LOQ | LOQ | 153.2 | 31.3 | ND | 4973 |
| 1BA100 | LOQ | 233.3 | 15.8 | LOQ | 5566 | ND | ND | 329.6 | LOQ | 195.4 | ND | 29.7 | ND | LOQ | ND | 15.4 | LOD | 6385 |
| 1BA1000 | ND | 234.0 | 19.7 | LOQ | 1256 | ND | ND | 487.3 | LOD | 293.3 | ND | 68.6 | ND | LOQ | ND | 99.5 | LOQ | 2459 |
| 1BA2000 | LOD | 183.3 | 15.3 | LOQ | 9515 | LOQ | ND | 852.5 | LOQ | 426.3 | ND | 751.5 | ND | 185.2 | ND | LOQ | LOQ | 11,229 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghimire, B.K.; Kim, S.-H.; Yu, C.-Y.; Chung, I.-M. Biochemical and Physiological Changes during Early Adventitious Root Formation in Chrysanthemum indicum Linné Cuttings. Plants 2022, 11, 1440. https://doi.org/10.3390/plants11111440
Ghimire BK, Kim S-H, Yu C-Y, Chung I-M. Biochemical and Physiological Changes during Early Adventitious Root Formation in Chrysanthemum indicum Linné Cuttings. Plants. 2022; 11(11):1440. https://doi.org/10.3390/plants11111440
Chicago/Turabian StyleGhimire, Bimal Kumar, Seung-Hyun Kim, Chang-Yeon Yu, and Ill-Min Chung. 2022. "Biochemical and Physiological Changes during Early Adventitious Root Formation in Chrysanthemum indicum Linné Cuttings" Plants 11, no. 11: 1440. https://doi.org/10.3390/plants11111440
APA StyleGhimire, B. K., Kim, S.-H., Yu, C.-Y., & Chung, I.-M. (2022). Biochemical and Physiological Changes during Early Adventitious Root Formation in Chrysanthemum indicum Linné Cuttings. Plants, 11(11), 1440. https://doi.org/10.3390/plants11111440