
Selenium Nanoparticles (Se-NPs) Alleviates Salinity Damages and Improves Phytochemical Characteristics of Pineapple Mint (Mentha suaveolens Ehrh.)

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
2.1. Growth Characteristics
2.2. Relative Water Content and Chlorophyll Index
2.3. Essential Oil Content and Compositions
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Experimental Design and Plants Material
4.3. Growth Parameters
4.4. Relative Water Content (RWC)
4.5. Chlorophyll Index
4.6. Essential Oil Extraction
4.7. Essential Oil Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El-Kashoury, E.-S.A.; El-Askary, H.I.; Kandil, Z.A.; Salem, M.A. Botanical and genetic characterization of Mentha suaveolens Ehrh. cultivated in Egypt. Pharmacogn. J. 2013, 5, 228–237. [Google Scholar] [CrossRef]
- Božović, M.; Pirolli, A.; Ragno, R. Mentha suaveolens Ehrh. (Lamiaceae) essential oil and its main constituent piperitenone oxide: Biological activities and chemistry. Molecules 2015, 20, 8605–8633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmadi, H.; Morshedloo, M.R.; Emrahi, R.; Javanmard, A.; Rasouli, F.; Maggi, F.; Kumar, M.; Lorenzo, J.M. Introducing Three New Fruit-Scented Mints to Farmlands: Insights on Drug Yield, Essential-Oil Quality and Antioxidant Properties. Antioxidants 2022, 11, 866. [Google Scholar] [CrossRef]
- Civitelli, L.; Panella, S.; Marcocci, M.E.; de Petris, A.; Garzoli, S.; Pepi, F.; Vavala, E.; Ragno, R.; Nencioni, L.; Palamara, A.T. In vitro inhibition of herpes simplex virus type 1 replication by Mentha suaveolens essential oil and its main component piperitenone oxide. Phytomedicine 2014, 21, 857–865. [Google Scholar] [CrossRef]
- Ezzat, S.M.; El-Kashoury, A.E.-S.; Kandil, A.Z.; Salem, A.M.; Sleem, A.A. Chemical and biological study of Mentha suaveolens Ehrh. cultivated in Egypt. J. Med. Plant Res. 2014, 8, 747–755. [Google Scholar]
- Oumzil, H.; Ghoulami, S.; Rhajaoui, M.; Ilidrissi, A.; Fkih-Tetouani, S.; Faid, M.; Benjouad, A. Antibacterial and antifungal activity of essential oils of Mentha suaveolens. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2002, 16, 727–731. [Google Scholar] [CrossRef]
- Aziz, E.E.; Al-Amier, H.; Craker, L.E. Influence of salt stress on growth and essential oil production in peppermint, pennyroyal, and apple mint. J. Herbs Spices Med. Plants 2008, 14, 77–87. [Google Scholar] [CrossRef]
- Sutour, S.; Bradesi, P.; Casanova, J.; Tomi, F. Composition and chemical variability of Mentha suaveolens ssp. suaveolens and M. suaveolens ssp. insularis from Corsica. Chem. Biodivers. 2010, 7, 1002–1008. [Google Scholar]
- Benayad, N.; Ebrahim, W.; Hakiki, A.; Mosaddak, M. Chemical characterization and insecticidal evaluation of the essential oil of Mentha suaveolens L. and Mentha pulegium L. growing in Morocco. Sci. Study Res. Chem. Chem. Eng. Biotechnol. Food Ind. 2012, 13, 27. [Google Scholar]
- Morshedloo, M.R.; Mumivand, H.; Craker, L.E.; Maggi, F. Chemical composition and antioxidant activity of essential oils in Origanum vulgare subsp. gracile at different phenological stages and plant parts. J. Food Process. Preserv. 2018, 42, e13516. [Google Scholar]
- Emrahi, R.; Morshedloo, M.R.; Ahmadi, H.; Javanmard, A.; Maggi, F. Intraspecific divergence in phytochemical characteristics and drought tolerance of two carvacrol-rich Origanum vulgare subspecies: Subsp. hirtum and subsp. gracile. Ind. Crops Prod. 2021, 168, 113557. [Google Scholar] [CrossRef]
- Jamal Omidi, F.; Mohajjel Shoja, H.; Sariri, R. Effect of water-deficit stress on secondary metabolites of Melissa officinalis L.: Role of exogenous salicylic acid. Casp. J. Environ. Sci. 2018, 16, 121–134. [Google Scholar]
- Bistgani, Z.E.; Hashemi, M.; DaCosta, M.; Craker, L.; Maggi, F.; Morshedloo, M.R. Effect of salinity stress on the physiological characteristics, phenolic compounds and antioxidant activity of Thymus vulgaris L. and Thymus daenensis Celak. Ind. Crops Prod. 2019, 135, 311–320. [Google Scholar] [CrossRef]
- Khalvandi, M.; Amerian, M.; Pirdashti, H.; Keramati, S.; Hosseini, J. Essential oil of peppermint in symbiotic relationship with Piriformospora indica and methyl jasmonate application under saline condition. Ind. Crops Prod. 2019, 127, 195–202. [Google Scholar] [CrossRef]
- Kasrati, A.; Jamali, C.A.; Bekkouche, K.; Wohlmuth, H.; Leach, D.; Abbad, A. Plant growth, mineral nutrition and volatile oil composition of Mentha suaveolens subsp. timija (Briq.) Harley cultivated under salt stress conditions. Ind. Crops Prod. 2014, 59, 80–84. [Google Scholar]
- Djanaguiraman, M.; Prasad, P. Effects of salinity on ion transport, water relations and oxidative damage. In Ecophysiology and Responses of Plants under Salt Stress; Springer: Berlin/Heidelberg, Germany, 2013; pp. 89–114. [Google Scholar]
- Csiszár, J.; Lantos, E.; Tari, I.; Madosa, E.; Wodala, B.; Vashegyi, Á.; Horváth, F.; Pécsváradi, A.; Szabó, M.; Bartha, B. Antioxidant enzyme activities in Allium species and their cultivars under water stress. Plant Soil Environ. 2007, 53, 517. [Google Scholar] [CrossRef] [Green Version]
- Mirza, H.; Hossain, M.A.; Fujita, M. Selenium in higher plants: Physiological role, antioxidant metabolism and abiotic stress tolerance. J. Plant Sci. 2010, 5, 354–375. [Google Scholar]
- Hasanuzzaman, M.; Bhuyan, M.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Selmar, D. Potential of salt and drought stress to increase pharmaceutical significant secondary compounds in plants. Landbauforsch. Volkenrode 2008, 58, 139. [Google Scholar]
- Hasani, A.; Omid, B.R. Effects of water stress on some morphological, physiological and metabolical characteristics of basil (Ocimum basilicum). J. Agric. Sci. 2002, 12, 47–59. [Google Scholar]
- Banerjee, A.; Roychoudhury, A. Effect of salinity stress on growth and physiology of medicinal plants. In Medicinal Plants and Environmental Challenges; Springer: Berlin/Heidelberg, Germany, 2017; pp. 177–188. [Google Scholar]
- Ostadi, A.; Javanmard, A.; Machiani, M.A.; Morshedloo, M.R.; Nouraein, M.; Rasouli, F.; Maggi, F. Effect of different fertilizer sources and harvesting time on the growth characteristics, nutrient uptakes, essential oil productivity and composition of Mentha × piperita L. Ind. Crops Prod. 2020, 148, 112290. [Google Scholar] [CrossRef]
- Semida, W.M.; Abdelkhalik, A.; Mohamed, G.; El-Mageed, A.; Taia, A.; El-Mageed, A.; Shimaa, A.; Rady, M.M.; Ali, E.F. Foliar application of zinc oxide nanoparticles promotes drought stress tolerance in eggplant (Solanum melongena L.). Plants 2021, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.-S.P. Alleviation of the effect of salinity on growth and yield of strawberry by foliar spray of selenium-nanoparticles. Environ. Pollut. 2019, 253, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, F.; Ahmad, R.; Ashraf, M.; Waraich, E.; Khan, S. Effect of selenium foliar spray on physiological and biochemical processes and chemical constituents of wheat under drought stress. Ecotoxicol. Environ. Saf. 2015, 113, 191–200. [Google Scholar] [CrossRef]
- Ning, N.; Yuan, X.-Y.; Dong, S.-Q.; Wen, Y.-Y.; Gao, Z.-P.; Guo, M.-J.; Guo, P.-Y. Increasing selenium and yellow pigment concentrations in foxtail millet (Setaria italica L.) grain with foliar application of selenite. Biol. Trace Elem. Res. 2016, 170, 245–252. [Google Scholar] [CrossRef]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Çakir, Ö.; Turgut-Kara, N.; Ari, Ş. Selenium induced selenocysteine methyltransferase gene expression and antioxidant enzyme activities in Astragalus chrysochlorus. Acta Bot. Croat. 2016, 75, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Cartes, P.; Gianfreda, L.; Paredes, C.; Mora, M. Selenium uptake and its antioxidant role in ryegrass cultivars as affected by selenite seed pelletization. J. Soil Sci. Plant Nutr. 2011, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Germ, M.; Stibilj, V.; Kreft, I. Metabolic importance of selenium for plants. Eur. J. Plant Sci. Biotechnol. 2007, 1, 91–97. [Google Scholar]
- Xia, X.; Ling, L.; Zhang, W.-X. Genesis of pure Se (0) nano-and micro-structures in wastewater with nanoscale zero-valent iron (nZVI). Environ. Sci. Nano 2017, 4, 52–59. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Prasad, P.V.; Seppanen, M. Selenium protects sorghum leaves from oxidative damage under high temperature stress by enhancing antioxidant defense system. Plant Physiol. Biochem. 2010, 48, 999–1007. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; Volume 456. [Google Scholar]
- Morshedloo, M.R.; Maggi, F.; Neko, H.T.; Aghdam, M.S. Sumac (Rhus coriaria L.) fruit: Essential oil variability in Iranian populations. Ind. Crops Prod. 2018, 111, 1–7. [Google Scholar] [CrossRef]
- Amzallag, G.; Lerner, H.; Poljakoff-Mayber, A. Exogenous ABA as a modulator of the response of sorghum to high salinity. J. Exp. Bot. 1990, 41, 1529–1534. [Google Scholar] [CrossRef]
- Greenway, H.; Munns, R. Mechanisms of salt tolerance in nonhalophytes. Annu. Rev. Plant Physiol. 1980, 31, 149–190. [Google Scholar] [CrossRef]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef]
- Zhu, J.-K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Tanji, K.K. Agricultural Salinity Assessment and Management; Scientific Publishers: Singapore, 2012. [Google Scholar]
- Lopez, C.; Takahashi, H.; Yamazaki, S. Plant–water relations of kidney bean plants treated with NaCl and foliarly applied glycinebetaine. J. Agron. Crop Sci. 2002, 188, 73–80. [Google Scholar] [CrossRef]
- Weisany, W.; Sohrabi, Y.; Heidari, G.; Siosemardeh, A.; Ghassemi-Golezani, K. Physiological responses of soybean (‘Glycine max’ L.) To zinc application under salinity stress. Aust. J. Crop Sci. 2011, 5, 1441–1447. [Google Scholar]
- Nedjimi, B. Salt tolerance strategies of Lygeum spartum L.: A new fodder crop for Algerian saline steppes. Flora-Morphol. Distrib. Funct. Ecol. Plants 2009, 204, 747–754. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 17672. [Google Scholar] [CrossRef]
- Yeo, A.; Lee, Λ.-S.; Izard, P.; Boursier, P.; Flowers, T. Short-and long-term effects of salinity on leaf growth in rice (Oryza sativa L.). J. Exp. Bot. 1991, 42, 881–889. [Google Scholar] [CrossRef]
- Yamane, K.; Kawasaki, M.; Taniguchi, M.; Miyake, H. Differential effect of NaCl and polyethylene glycol on the ultrastructure of chloroplasts in rice seedlings. J. Plant Physiol. 2003, 160, 573–575. [Google Scholar] [CrossRef] [PubMed]
- El-Kinany, R.; Brengi, S.; Nassar, A.; El-Batal, A. Enhancement of Plant Growth, Chemical Composition and Secondary Metabolites of Essential Oil of Salt-Stressed Coriander (Coriandrum Sativum L.) Plants Using Selenium, Nano-Selenium, and Glycine Betaine. Sci. J. Flowers Ornam. Plants 2019, 6, 151–173. [Google Scholar] [CrossRef] [Green Version]
- Morshedloo, M.R.; Craker, L.E.; Salami, A.; Nazeri, V.; Sang, H.; Maggi, F. Effect of prolonged water stress on essential oil content, compositions and gene expression patterns of mono-and sesquiterpene synthesis in two oregano (Origanum vulgare L.) subspecies. Plant Physiol. Biochem. 2017, 111, 119–128. [Google Scholar] [CrossRef]
- Rehman, R.; Hanif, M.A.; Mushtaq, Z.; Mochona, B.; Qi, X. Biosynthetic factories of essential oils: The aromatic plants. Nat. Prod. Chem. Res. 2016, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Heydari, F.; Zehtab, S.S.; Javanshir, A.; Aliari, H.; Dadpour, M. The effects of application microelements and plant density on yield and essential oil of peppermint (Mentha piperita L.). Iran. J. Med. Aromat. Plants 2008, 24, 1–9. [Google Scholar]
- Tavakoli, S.; Enteshari, S.; Yousefifard, M. Investigation of the effect of selenium on growth, antioxidant capacity and secondary metabolites in Melissa officinalis. Iran. J. Plant Physiol. 2020, 10, 3125–3134. [Google Scholar]
- Lorenzo, D.; Paz, D.; Dellacassa, E.; Davies, P.; Vila, R.; Cañigueral, S. Essential oils of Mentha pulegium and Mentha rotundifolia from Uruguay. Braz. Arch. Biol. Technol. 2002, 45, 519–524. [Google Scholar] [CrossRef]
- Aziz, E.E.; Craker, L. Essential oil constituents of peppermint, pennyroyal, and apple mint grown in a desert agrosystem. J. Herbs Spices Med. Plants 2010, 15, 361–367. [Google Scholar] [CrossRef]
- El-Keltawi, N.E.; Croteau, R. Salinity depression of growth and essential oil formation in spearmint and marjoram and its reversal by foliar applied cytokinin. Phytochemistry 1987, 26, 1333–1334. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Prajapati, V.; Ahmad, A.; Aggarwal, K.K.; Khanuja, S.P. Piperitenone oxide as toxic, repellent, and reproduction retardant toward malarial vector Anopheles stephensi (Diptera: Anophelinae). J. Med. Entomol. 2004, 41, 691–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
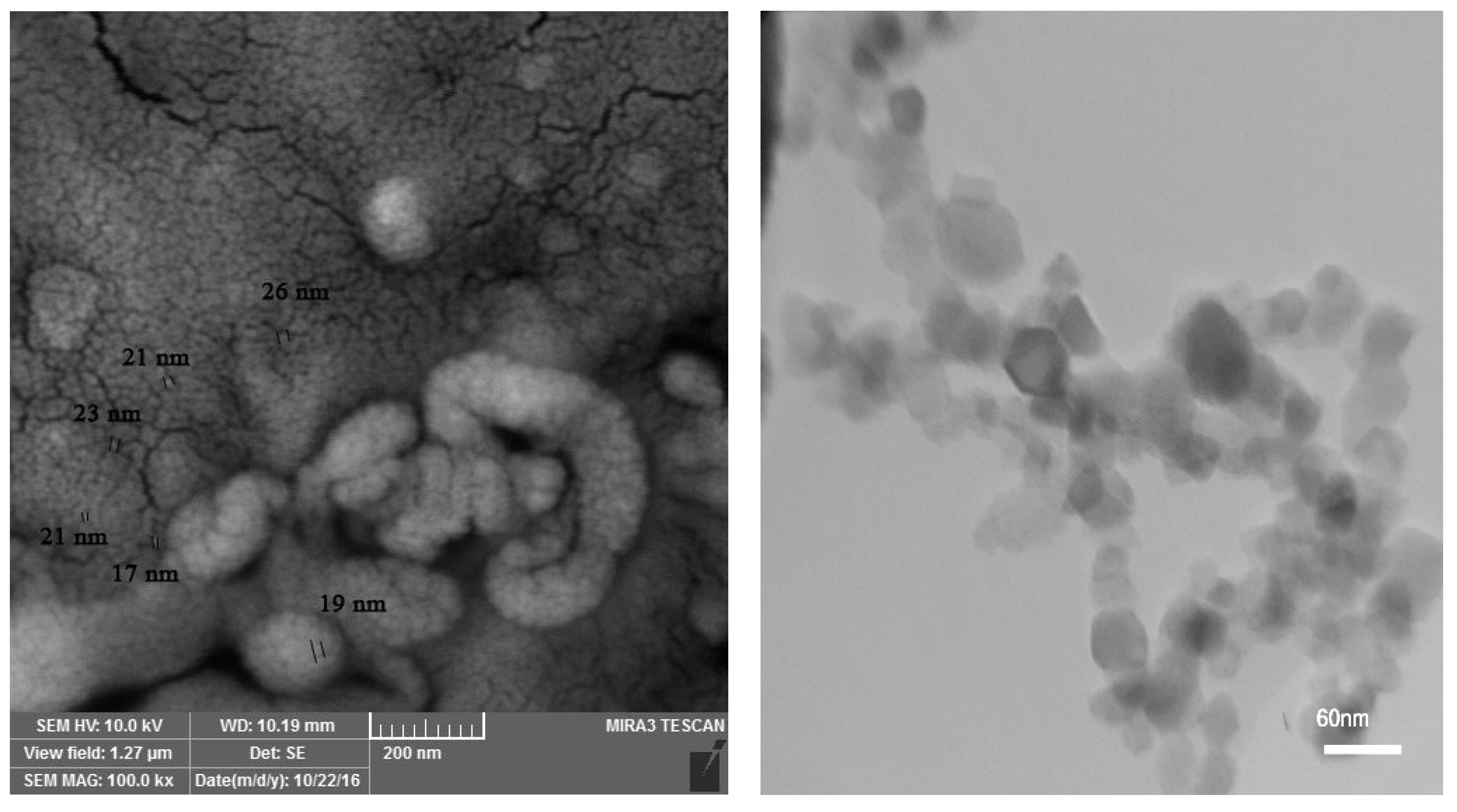

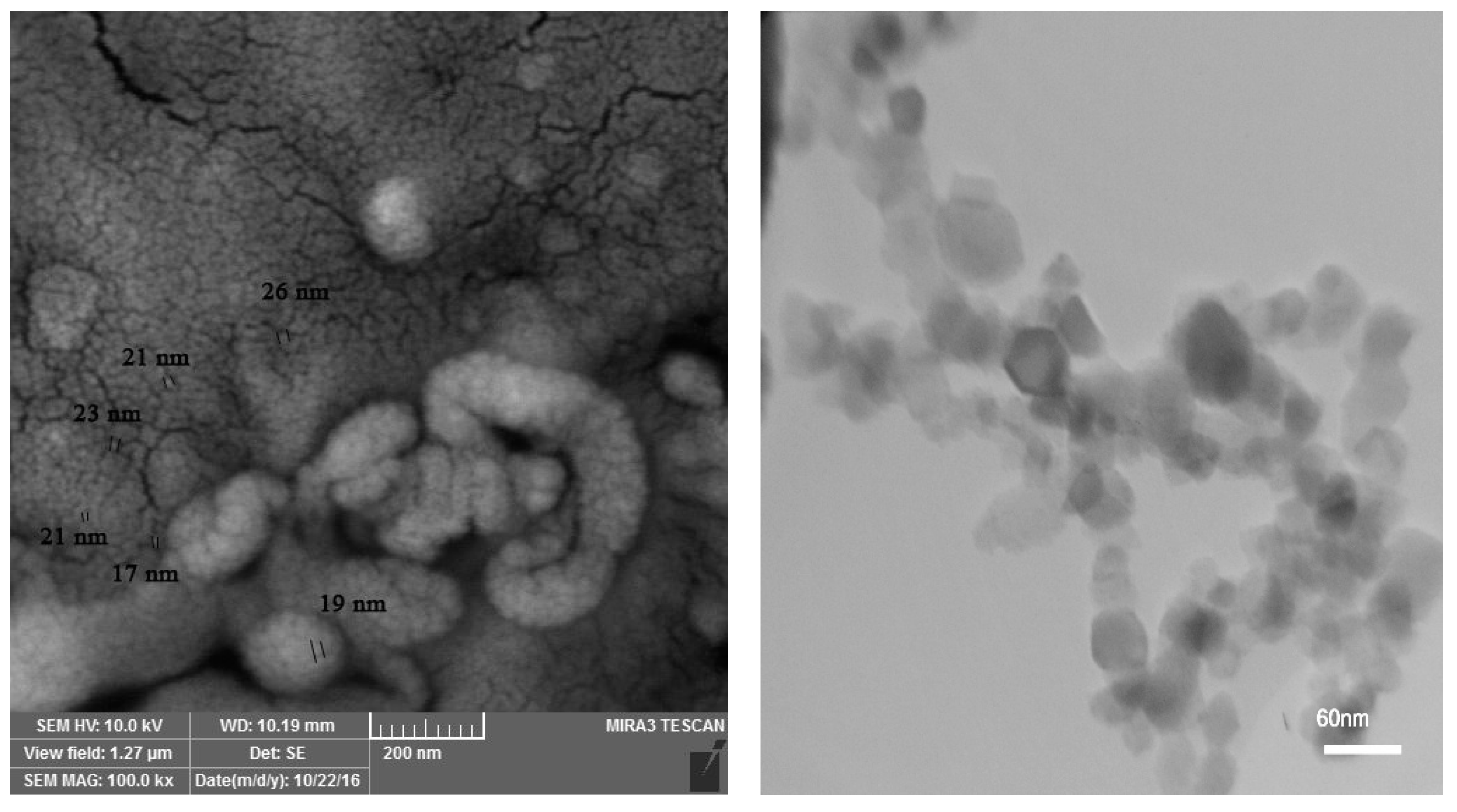{kind=link}
| FW | DW | PH | RWC | Chl Index | Essential Oil | |
|---|---|---|---|---|---|---|
| g | g | cm | % | % | ||
| Salinity (S) | ** | Ns | ** | ** | ns | Ns |
| 0 | 86.6 a | 18.86 | 31.66 a | 71.02 a | 37.75 | 0.36 |
| 30 | 74.92 b | 18.77 | 28.44 b | 65.98 ab | 36.74 | 0.32 |
| 60 | 73.1 b | 19.54 | 28 b | 64.3 bc | 37.78 | 0.31 |
| 90 | 72.36 b | 19.01 | 25.61 b | 58.2 c | 36.42 | 0.29 |
| Se (NPs) | ns | Ns | ns | ns | ns | Ns |
| 0 | 77.43 | 18.64 | 28.58 | 63.18 | 36.88 | 0.30 |
| 10 | 78.26 | 19.35 | 28.25 | 65.76 | 37.34 | 0.34 |
| 20 | 74.54 | 19.15 | 28.45 | 65.05 | 37.29 | 0.32 |
| S × NPs | ns | Ns | ns | ns | * | Ns |
| 0 × 0 | 83.33 | 17.43 | 32.66 | 67.22 | 36.49 bac | 0.35 |
| 0 × 10 | 95.46 | 20.76 | 31.83 | 67.20 | 36.89 bac | 0.36 |
| 0 × 20 | 81.10 | 18.40 | 30.50 | 78.64 | 39.87 a | 0.37 |
| 30 × 0 | 74.50 | 19.03 | 28.50 | 67.18 | 36.67 bac | 0.27 |
| 30 × 10 | 73.86 | 18.76 | 28.33 | 61.63 | 37.55 bac | 0.34 |
| 30 × 20 | 68.73 | 18.53 | 28.50 | 64.09 | 36 bc | 0.35 |
| 60 × 0 | 72.36 | 19.00 | 28.50 | 55.30 | 39.94 a | 0.29 |
| 60 × 10 | 68.23 | 18.50 | 25.16 | 62.34 | 36.25 bc | 0.34 |
| 60 × 20 | 78.70 | 21.13 | 30.33 | 56.96 | 37.16 bac | 0.32 |
| 90 × 0 | 79.63 | 19.10 | 24.66 | 65.57 | 34.42 c | 0.32 |
| 90 × 10 | 75.50 | 19.36 | 27.66 | 71.86 | 38.68 ba | 0.31 |
| 90 × 20 | 69.63 | 18.56 | 24.50 | 60.53 | 36.15 bc | 0.25 |
| Fresh Weight | Dry Weight | Plant Height | RWC | Chl Index | Essential Oil Content | Piperitenone Oxide | β-Myrcene | Limonene | (Z)-Jasmone | (E)-β-Farnesene | Germacrene D | Viridiflorol | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fresh weight | 1.00 | ||||||||||||
| Dry weight | 0.64 ** | 1.00 | |||||||||||
| Plant height | 0.31 | 0.07 | 1.00 | ||||||||||
| RWC | 0.31 | 0.02 | 0.14 | 1.00 | |||||||||
| Chl index | 0.12 | 0.18 | 0.27 | 0.07 | 1.00 | ||||||||
| Essential oil content | 0.21 | 0.01 | 0.20 | 0.29 | 0.02 | 1.00 | |||||||
| Piperitenone oxide | 0.29 | 0.03 | −0.05 | 0.06 | −0.23 | 0.19 | 1.00 | ||||||
| β-Myrcene | −0.36 * | 0.09 | −0.14 | −0.39 * | 0.17 | −0.42 * | −0.78 ** | 1.00 | |||||
| Limonene | −0.23 | 0.01 | 0.04 | −0.21 | 0.29 | −0.366 * | −0.86 ** | 0.92 ** | 1.00 | ||||
| (Z)-Jasmone | −0.17 | −0.07 | 0.07 | −0.07 | 0.15 | −0.11 | −0.68 ** | 0.73 ** | 0.78 ** | 1.00 | |||
| (E)-β-Farnesene | −0.22 | 0.05 | 0.06 | −0.05 | 0.13 | −0.16 | −0.90 ** | 0.75 ** | 0.77 ** | 0.70 ** | 1.00 | ||
| Germacrene D | −0.06 | −0.05 | 0.27 | 0.05 | 0.17 | 0.01 | −0.83 ** | 0.62 ** | 0.75 ** | 0.79 ** | 0.89 ** | 1.00 | |
| Viridiflorol | −0.31 | −0.09 | −0.04 | 0.02 | 0.13 | −0.13 | −0.72 ** | 0.30 | 0.365 * | 0.13 | 0.55 ** | 0.38 * | 1.00 |
| Constituents | RI * | Treatments | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 (mM) | 30 (mM) | 60 (mM) | 90 (mM) | ||||||||||
| 0 | 10 | 20 | 0 | 10 | 20 | 0 | 10 | 20 | 0 | 10 | 20 | ||
| α-Pinene | 930 | 0.53 | 0.53 | 0.56 | 0.73 | 0.68 | 0.75 | 0.73 | 0.62 | 0.68 | 0.56 | 0.64 | 0.62 |
| Sabinene | 969 | 0.29 | 0.29 | 0.32 | 0.44 | 0.39 | 0.42 | 0.44 | 0.36 | 0.39 | 0.36 | 0.39 | 0.4 |
| β-Pinene | 972 | 0.67 | 0.69 | 0.75 | 0.97 | 0.86 | 0.93 | 0.97 | 0.82 | 0.89 | 0.80 | 0.89 | 0.89 |
| β-Myrcene | 989 | 0.51 | 0.52 | 0.59 | 0.82 | 0.72 | 0.74 | 0.84 | 0.62 | 0.65 | 0.55 | 0.52 | 0.69 |
| n-Decane | 998 | 0.07 | 0.03 | 0.03 | 0.03 | 0.06 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Limonene | 1025 | 2.75 | 2.45 | 3.09 | 4.01 | 3.44 | 3.32 | 4.09 | 2.6 | 2.61 | 2.44 | 2.54 | 2.98 |
| (Z)-β-Ocimene | 1035 | 0.17 | 0.16 | 0.21 | 0.25 | 0.22 | 0.22 | 0.23 | 0.09 | 0.13 | 0.09 | 0.12 | 0.13 |
| n-Undecane | 1098 | 0.08 | 0.06 | 0.09 | 0.14 | 0.13 | 0.12 | 0.13 | 0.05 | 0.05 | 0.09 | 0.11 | 0.05 |
| 1-Octen-3-yl acetate | 1111 | 1.89 | 1.77 | 2.45 | 3.0 | 2.75 | 2.75 | 3.01 | 2.59 | 2.57 | 2.69 | 3.25 | 2.66 |
| Borneol | 1160 | 0.1 | 0.17 | 0.37 | 0.43 | 0.25 | 0.32 | 0.7 | 0.54 | 0.56 | 0.75 | 0.84 | 0.65 |
| n-Dodecane | 1198 | 0.53 | 0.74 | 0.8 | 1.25 | 1.24 | 1.69 | 0.79 | 1.11 | 1.08 | 1.16 | 1.19 | 1.02 |
| Piperitenone oxide | 1368 | 76.69 | 79.45 | 72.97 | 67.74 | 69.26 | 67.85 | 70.47 | 76.85 | 77.39 | 78.69 | 75.72 | 76.35 |
| β-Bourbonene | 1380 | 0.22 | 0.22 | 0.27 | 0.32 | 0.33 | 0.32 | 0.29 | 0.24 | 0.34 | 0.19 | 0.12 | 0.39 |
| β-Elemene | 1388 | 0.37 | 0.33 | 0.48 | 0.58 | 0.83 | 0.91 | 0.66 | 0.65 | 0.6 | 0.55 | 0.68 | 0.63 |
| (Z)-Jasmone | 1394 | 0.92 | 0.9 | 1.37 | 1.34 | 1.15 | 1.26 | 1.32 | 0.99 | 0.95 | 0.89 | 0.6 | 0.01 |
| trans-Caryophyllene | 1413 | 0.52 | 0.49 | 0.65 | 0.72 | 0.75 | 0.86 | 0.62 | 0.52 | 0.48 | 0.43 | 0.39 | 0.52 |
| (E)-β-Farnesene | 1455 | 0.99 | 0.9 | 1.22 | 1.37 | 1.41 | 1.53 | 1.17 | 1.02 | 1.0 | 0.91 | 0.88 | 1.07 |
| Germacrene D | 1476 | 6.65 | 5.4 | 7.48 | 8.0 | 8.06 | 8.65 | 6.81 | 4.99 | 4.2 | 3.37 | 3.17 | 4.49 |
| Viridiflorol | 1585 | 1.69 | 1.34 | 1.78 | 2.11 | 2.19 | 2.35 | 1.78 | 1.73 | 1.54 | 1.67 | 2.26 | 1.73 |
| Treatments | Piperitenone Oxide | Limonene | Viridiflorol | Jasmone | β-Myrcene | |
|---|---|---|---|---|---|---|
| Salinity (mM) | Se-NPs-(mg L−1) | |||||
| 0 | 0 | 76.69 ba | 2.75 dc | 1.69 edc | 0.92 c | 0.51 e |
| 10 | 79.45 a | 2.45 d | 1.34 e | 0.9 c | 0.52 e | |
| 20 | 72.97 bc | 3.09 dc | 1.78 bdc | 1.37 a | 0.59 ecd | |
| 30 | 0 | 67.74 d | 4.01 ba | 2.11 bac | 1.34 a | 0.82 a |
| 10 | 69.26 dc | 3.44 bac | 2.19 ba | 1.15 ba | 0.72 ba | |
| 20 | 67.85 d | 3.32 bc | 2.35 a | 1.26 ba | 0.74 ba | |
| 60 | 0 | 70.47 dc | 4.09 a | 1.78 bdc | 1.32 a | 0.84 a |
| 10 | 76.85 ba | 2.6 d | 1.73 edc | 0.99 c | 0.62 becd | |
| 20 | 77.39 a | 2.61 d | 1.54 ed | 0.95 c | 0.65 bcd | |
| 90 | 0 | 78.69 a | 2.44 d | 1.67 ed | 0.89 c | 0.55 ed |
| 10 | 75.72 ba | 2.54 d | 2.26 a | 0.6 d | 0.52 e | |
| 20 | 76.35 ba | 2.98 dc | 1.73 edc | 1 bc | 0.69 bc | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kiumarzi, F.; Morshedloo, M.R.; Zahedi, S.M.; Mumivand, H.; Behtash, F.; Hano, C.; Chen, J.-T.; Lorenzo, J.M. Selenium Nanoparticles (Se-NPs) Alleviates Salinity Damages and Improves Phytochemical Characteristics of Pineapple Mint (Mentha suaveolens Ehrh.). Plants 2022, 11, 1384. https://doi.org/10.3390/plants11101384
Kiumarzi F, Morshedloo MR, Zahedi SM, Mumivand H, Behtash F, Hano C, Chen J-T, Lorenzo JM. Selenium Nanoparticles (Se-NPs) Alleviates Salinity Damages and Improves Phytochemical Characteristics of Pineapple Mint (Mentha suaveolens Ehrh.). Plants. 2022; 11(10):1384. https://doi.org/10.3390/plants11101384
Chicago/Turabian StyleKiumarzi, Fatemeh, Mohammad Reza Morshedloo, Seyed Morteza Zahedi, Hasan Mumivand, Farhad Behtash, Christophe Hano, Jen-Tsung Chen, and Jose M. Lorenzo. 2022. "Selenium Nanoparticles (Se-NPs) Alleviates Salinity Damages and Improves Phytochemical Characteristics of Pineapple Mint (Mentha suaveolens Ehrh.)" Plants 11, no. 10: 1384. https://doi.org/10.3390/plants11101384
APA StyleKiumarzi, F., Morshedloo, M. R., Zahedi, S. M., Mumivand, H., Behtash, F., Hano, C., Chen, J.-T., & Lorenzo, J. M. (2022). Selenium Nanoparticles (Se-NPs) Alleviates Salinity Damages and Improves Phytochemical Characteristics of Pineapple Mint (Mentha suaveolens Ehrh.). Plants, 11(10), 1384. https://doi.org/10.3390/plants11101384