
Mitigating the Negative Effect of Drought Stress in Oat (Avena sativa L.) with Silicon and Sulphur Foliar Fertilization

 ,
, 
 ,
,  and
and 
Abstract
:1. Introduction
2. Results
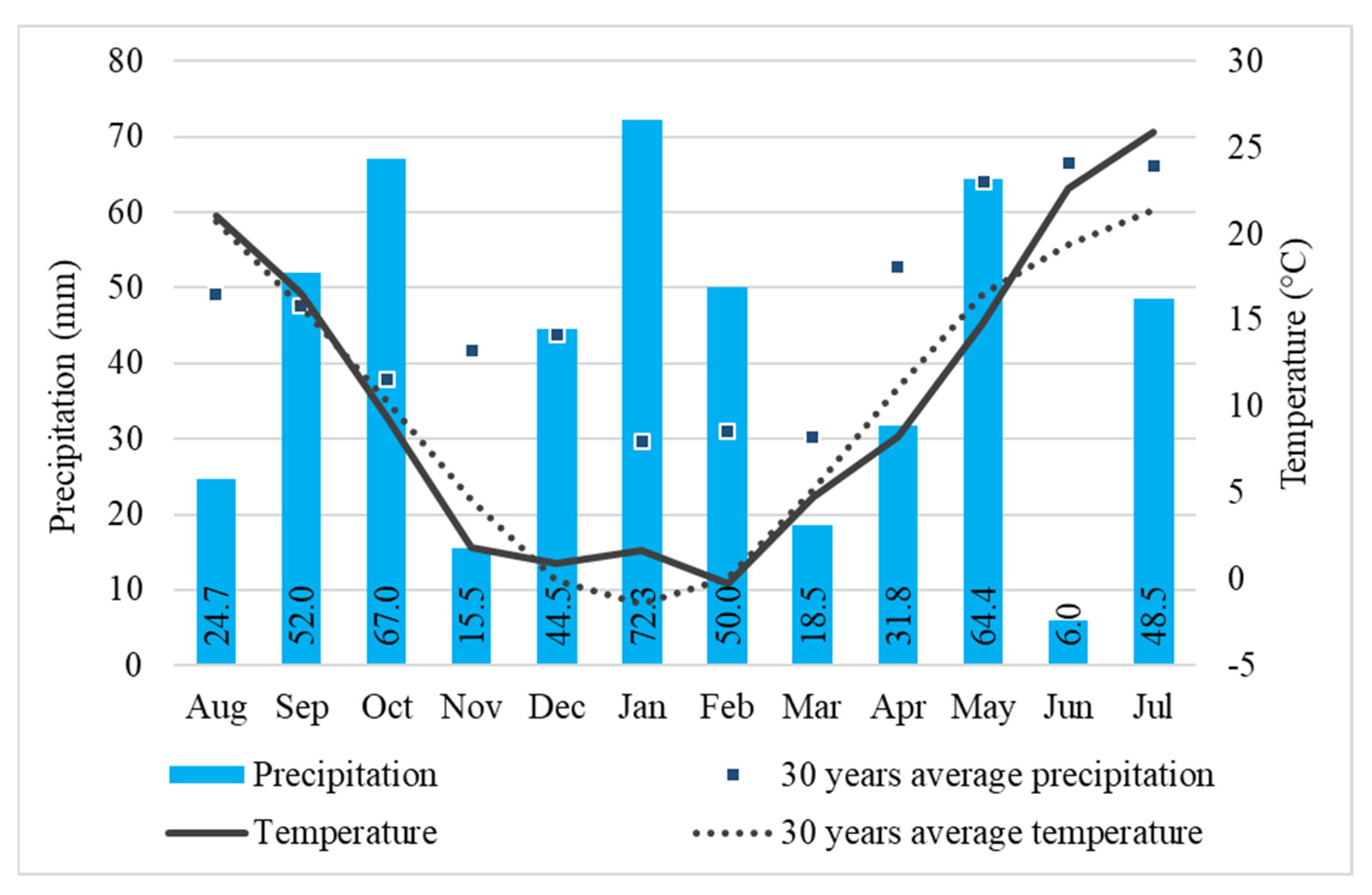
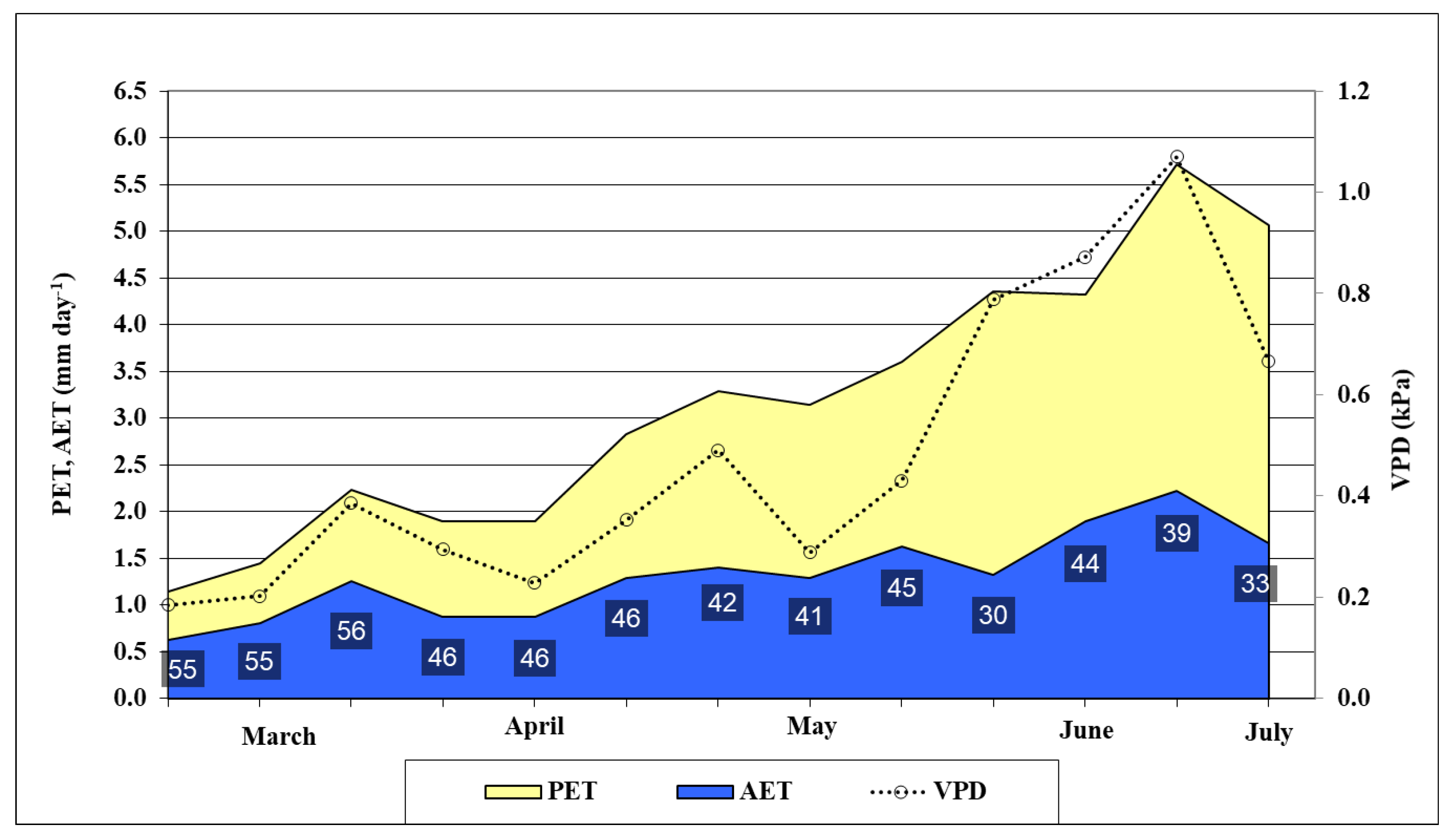
2.1. Evaluation of Water Status for Winter Oat in the Experiment Site
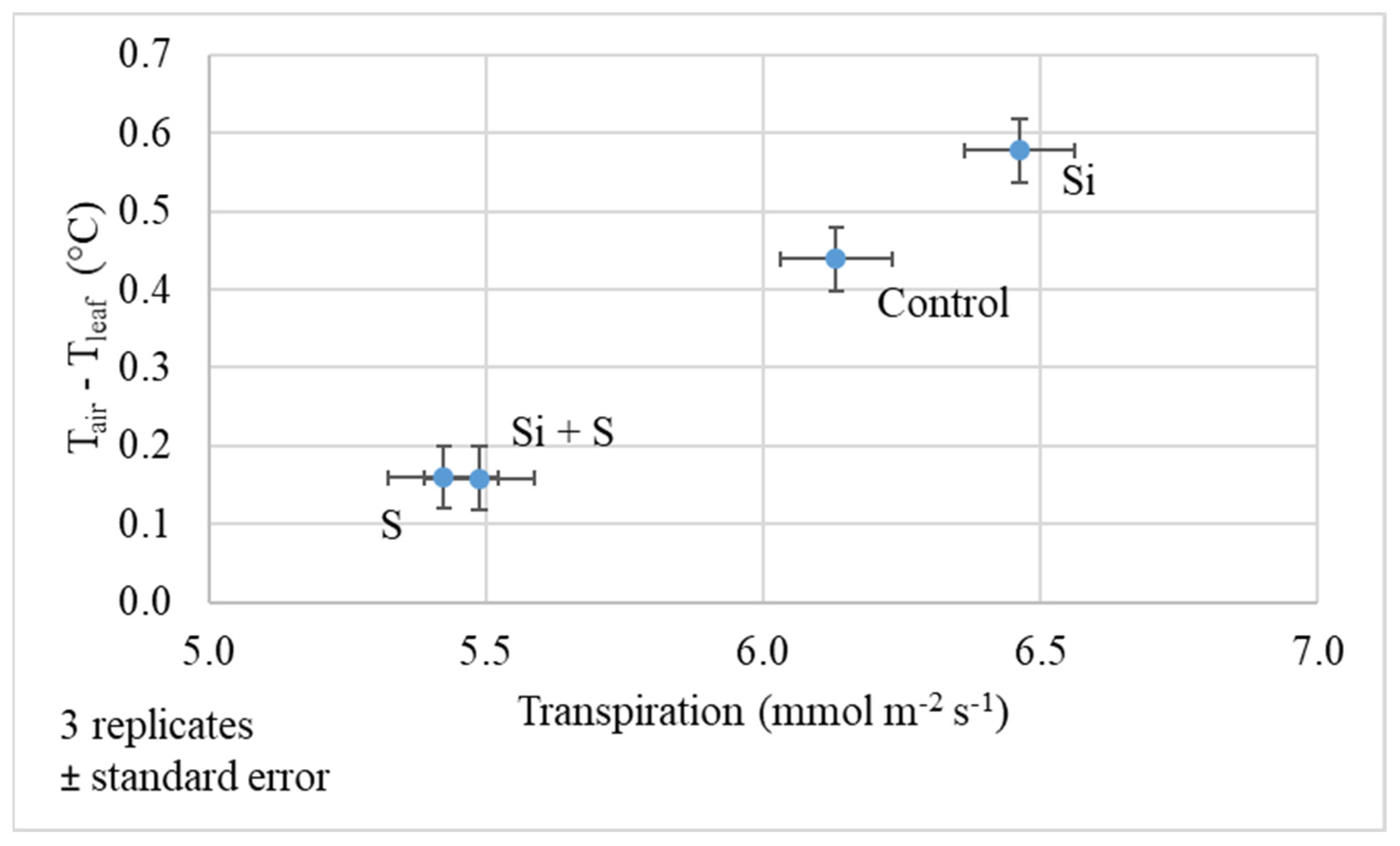
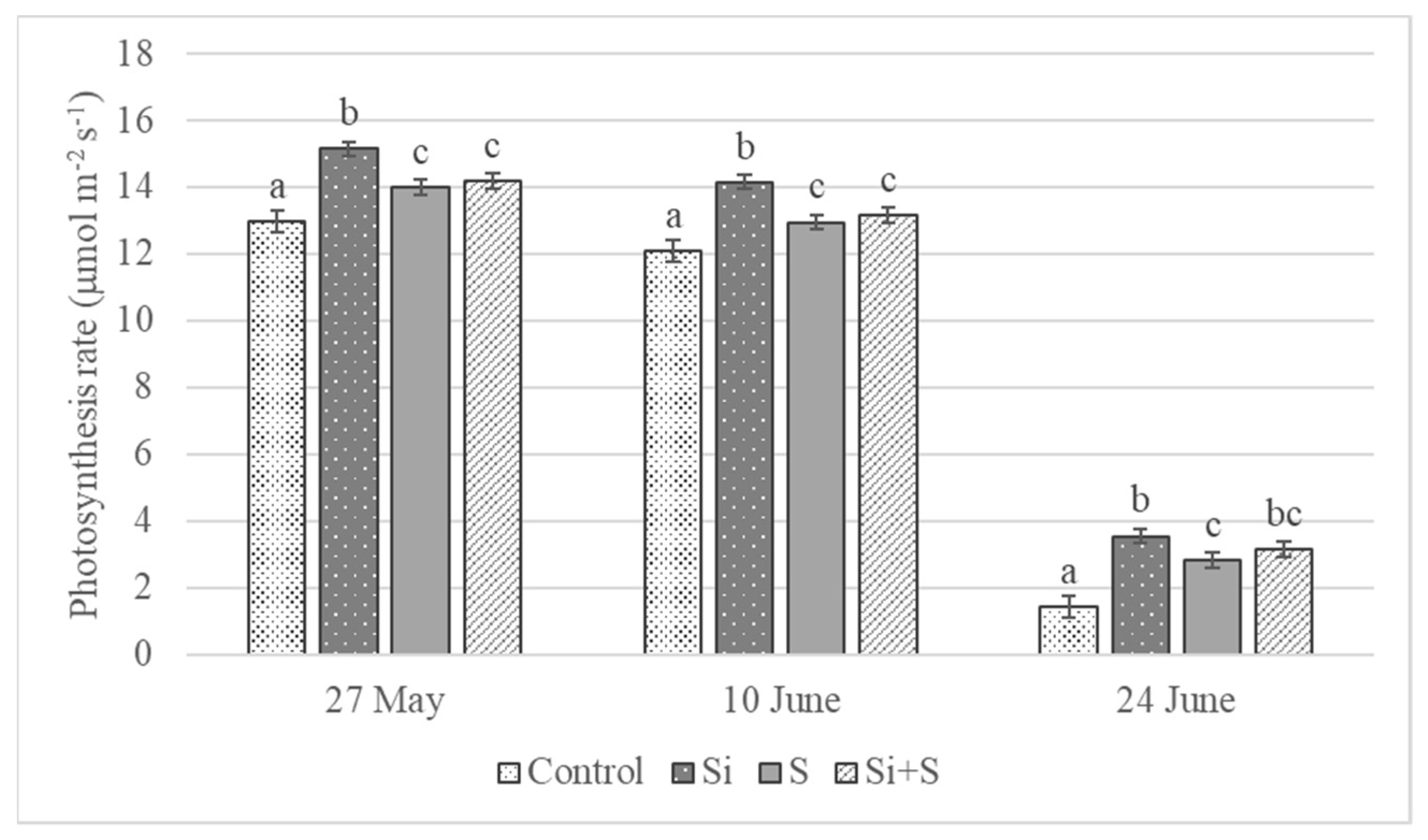
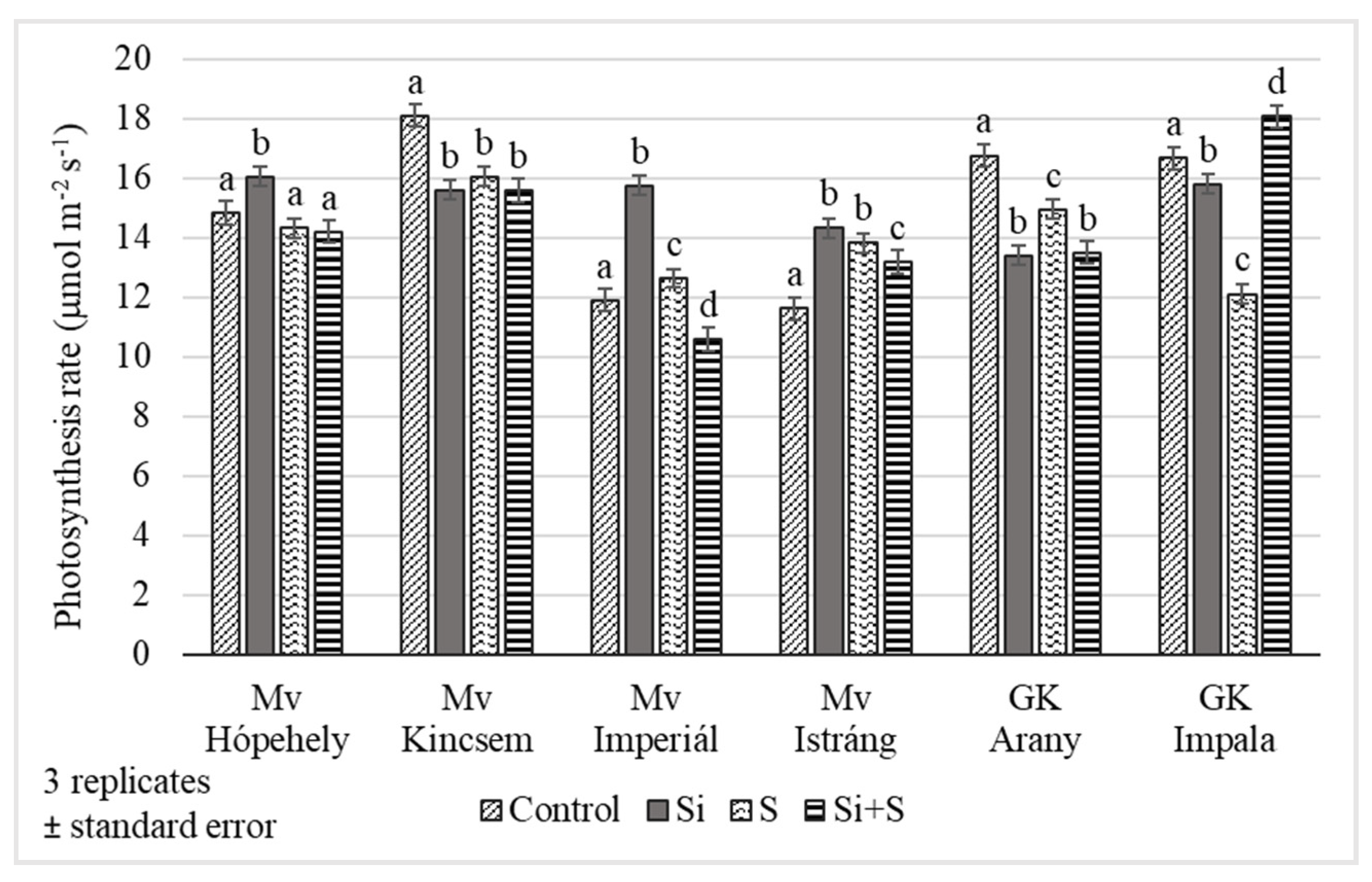
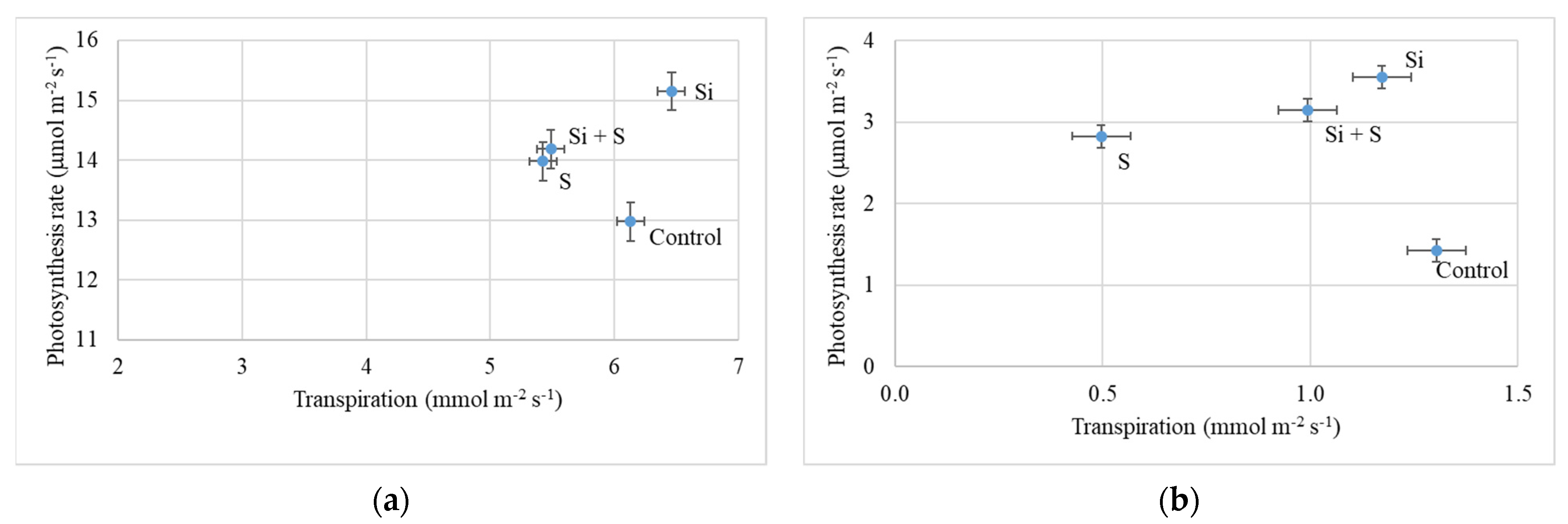
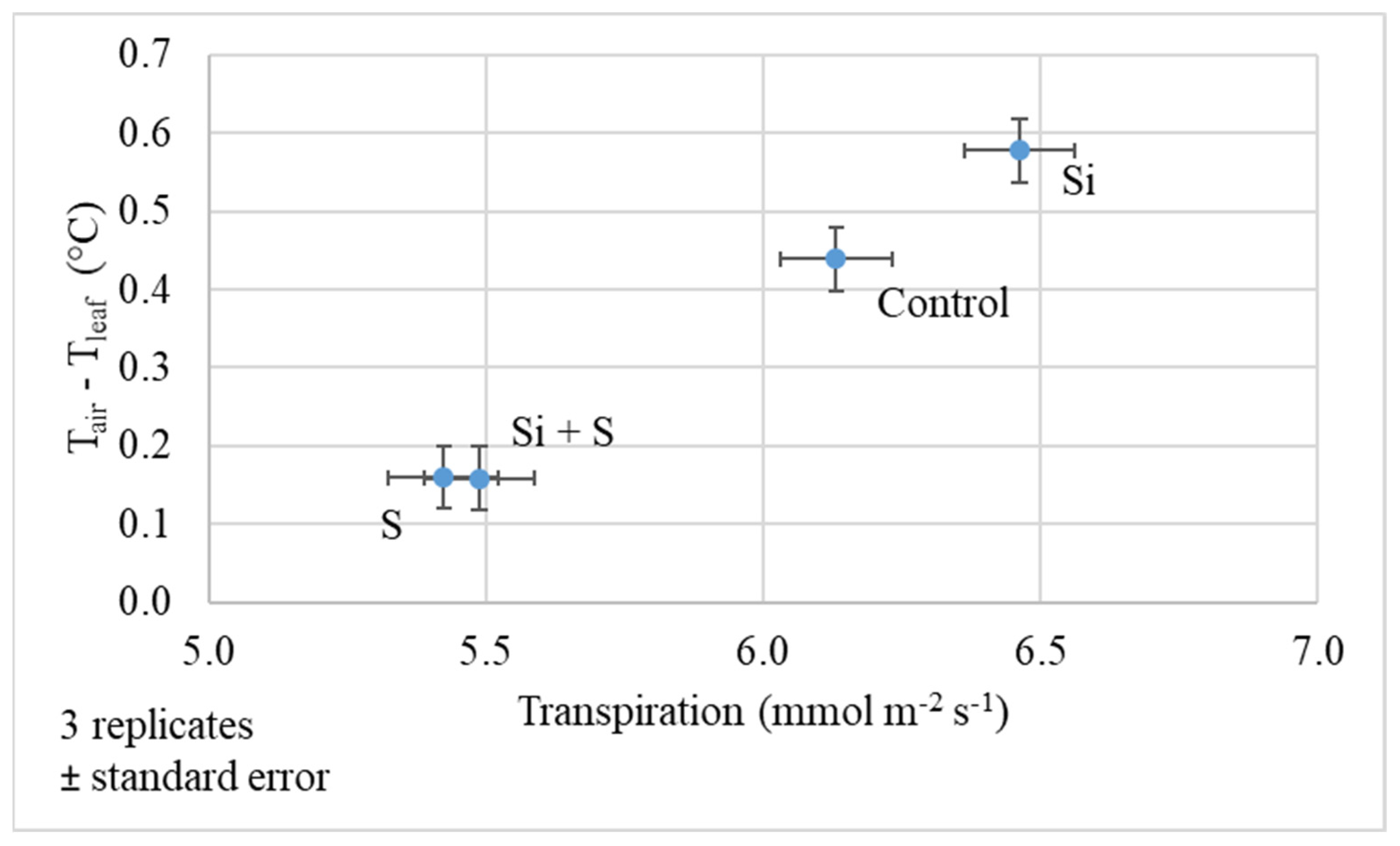
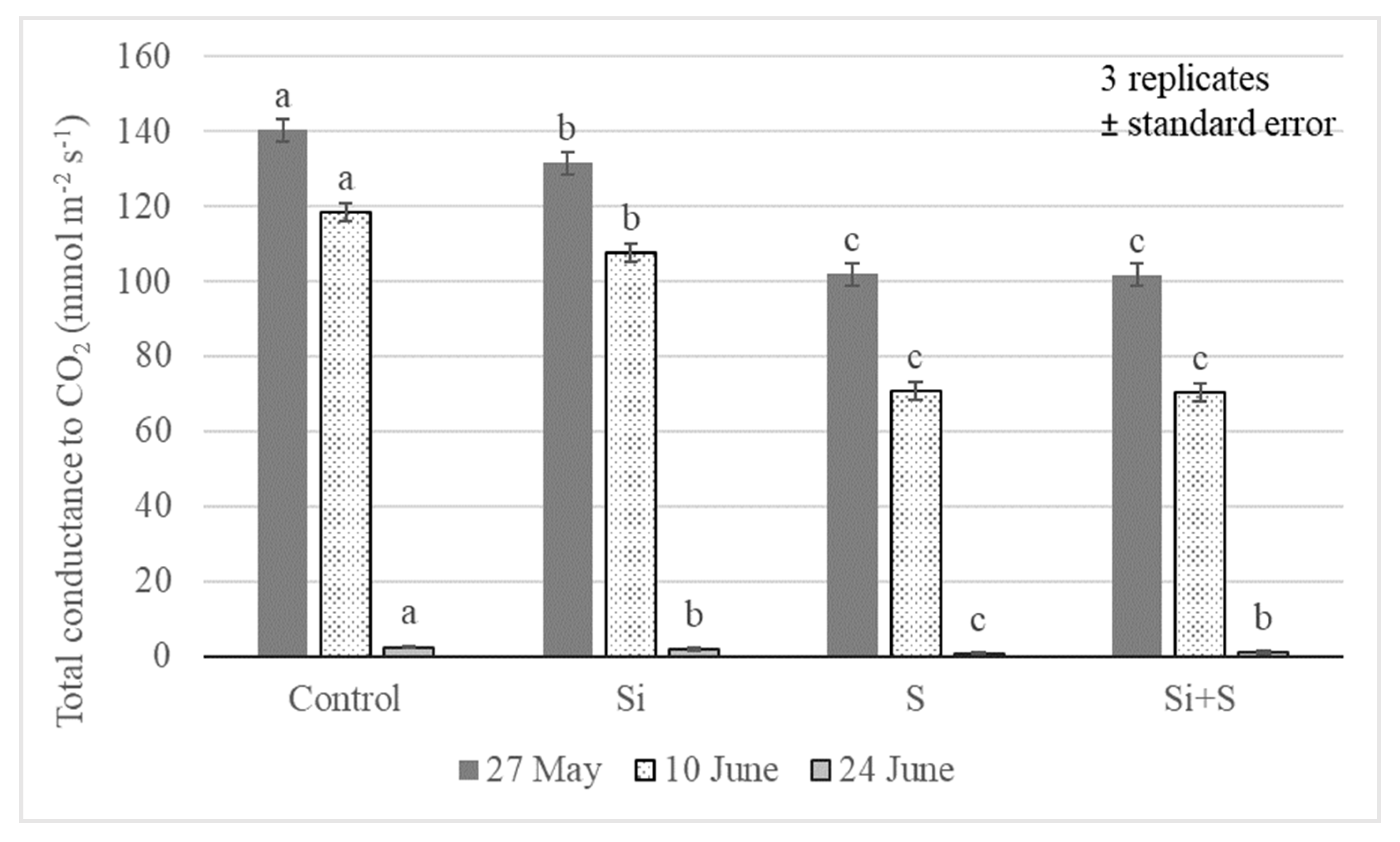
2.2. Response of Winter Oat Varieties to Drought Stress and the Effect of Foliar Fertilization through the Photosynthesis Parameters and Transpiration
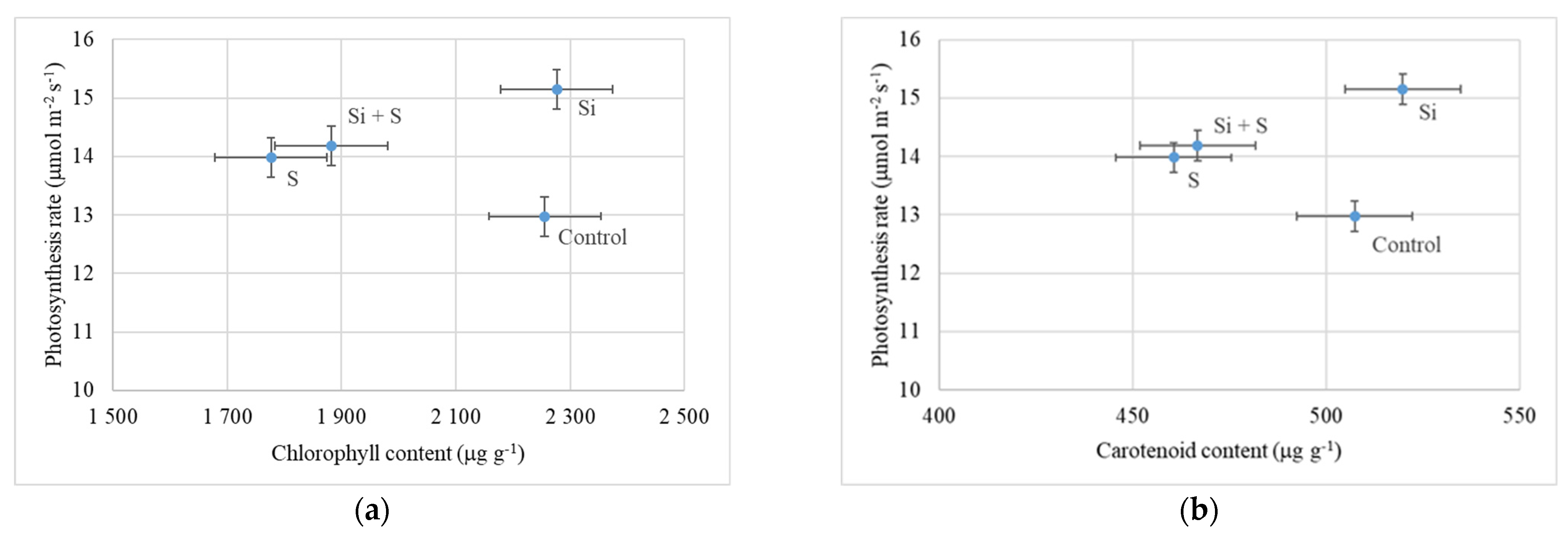
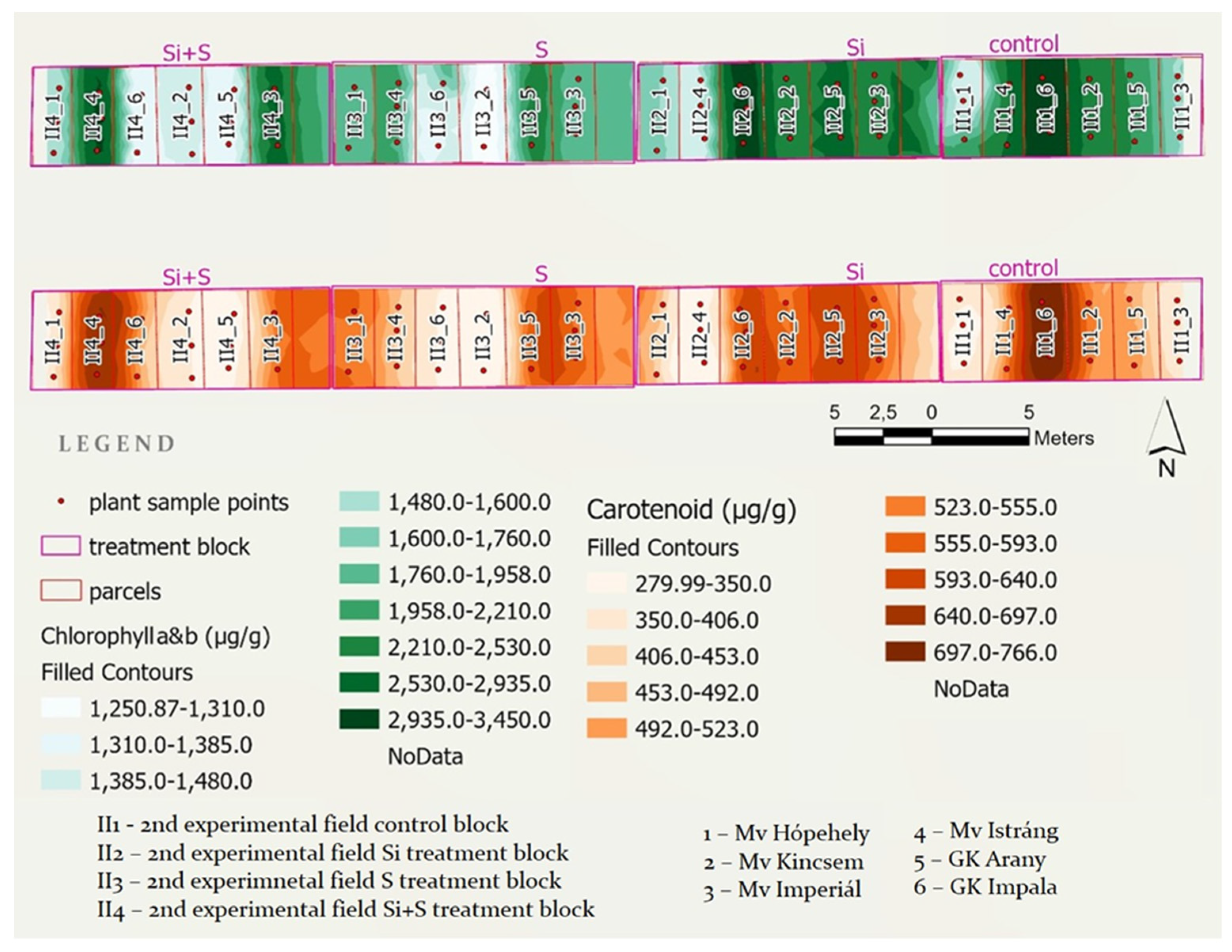
2.3. Effects of the Treatments on the Chlorophyll and Carotenoid Content of Oat
2.4. Water Use Efficiency of Winter Oat Varieties
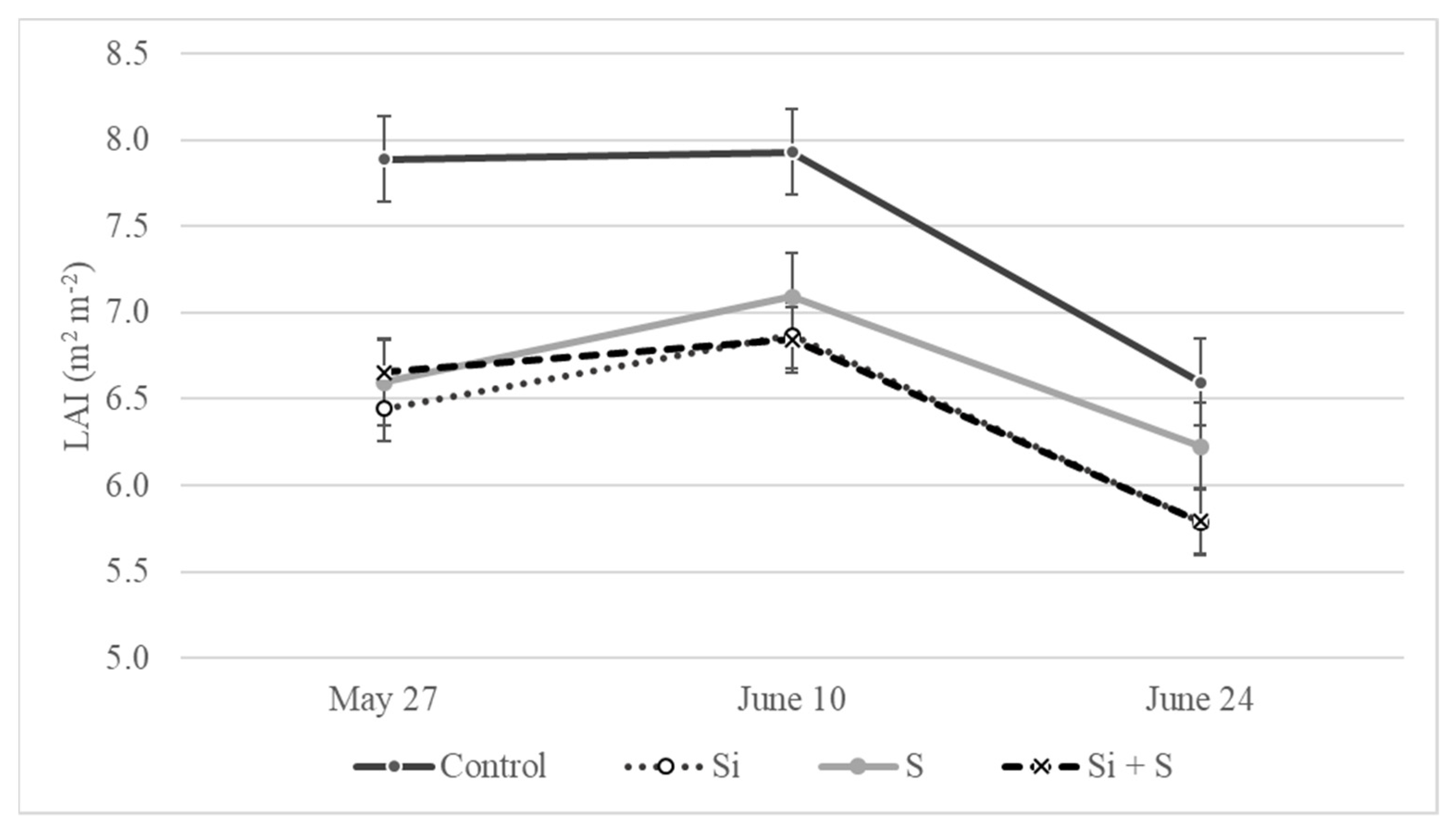
2.5. Leaf Area Changes as an Effect of the Treatments
2.6. Evaluation of the Yield and TKW of the Winter Oat Varieties
3. Discussion
4. Materials and Methods
4.1. Soil Characteristics of the Experimental Site
4.2. Experimental Setup
- Control, without foliar fertilization
- Silicon fertilization (Si) 3.0 L ha−1
- Sulphur fertilization (S) 5.0 L ha−1
- Silicon plus sulphur fertilization (Si + S) 3.0 + 5.0 L ha−1
- Sulphur fertilizer: liquid foliar fertilizer with high sulphur content (lignosulfonate formulation) 1000 g L−1 SO3, 30 g L−1 N, 30 g L−1 MgO, 27 g L−1 B, 0.003 g L−1 Mo
- Silicon fertilizer: (potassium silicate formulation) 1.4 m/m% Si, 10.5 m/m% K2O
- 1 December 2020
- BBCH13 (3 leaves unfolded)
- 10 May 2021
- BBCH39 (flag leaf stage)
- 18 June 2021
- BBCH73 (early milk)
4.3. Measurements, Calculations and, Their Methodology
- WUE: water use efficiency (kg m−3)
- Emm: transpirated H2O (mmol m2 s−1)
- Ass: assimilated CO2 (μmol m2 s−1)
- PET: potential evapotranspiration (mm day−1),
- Rn: net radiation at the crop surface (MJ m−2 day−1),
- G: soil heat flux density (MJ m−2 day−1),
- T: mean daily air temperature at 2 m height (°C),
- u2: wind speed at 2 m height (m s−1),
- es: saturation vapor pressure (kPa),
- ea: actual vapor pressure (kPa),
- Δ: slope vapor pressure curve (kPa °C−1),
- γ: psychrometric constant (kPa °C−1).
- AET: actual evapotranspiration (mm day−1),
- w: relative soil moisture content in the 0–100 cm layer (mm)
- b: crop coefficient factor of oat
- PET: potential evapotranspiration (mm day−1)
- w: relative soil moisture content in the 0–100 cm layer (mm)
- WP: permanent wilting point of the soil (mm)
- Wc: field capacity of the soil (mm)
- VPsat: saturated vapor pressure of the air (kPa),
- T: air temperature (°C)
- VPair: actual vapor pressure of the air (kPa)
- Chla + b: chlorophyll a and b content (µg/g)
- V: volume of extracted plant tissue (mL)
- A644: spectral absorbance value at wavelength of 644 nm (unitless)
- A663: spectral absorbance value at wavelength of 663 nm (unitless)
- w: weight of fresh plant tissue sample (g)
- Car: carotenoid content (µg/g)
- V: volume of extracted plant tissue (mL)
- A470: spectral absorbance value at wavelength of 470 nm (unitless)
- A644: spectral absorbance value at wavelength of 644 nm (unitless)
- A663: spectral absorbance value at wavelength of 663 nm (unitless)
- w: weight of fresh plant tissue sample (g)
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Marshall, A.H.; Cowan, S.; Edwards, S.; Griffiths, I.; Howarth, C.J.; Langdon, T.; White, E. Crops that feed the world 9. Oats—A cereal crop for human and livestock feed with industrial applications. Food Secur. 2013, 5, 13–33. [Google Scholar] [CrossRef]
- Allwood, J.W.; Martinez-Martin, P.; Xu, Y.; Cowan, A.; Pont, S.; Griffiths, I.; Sungurtas, J.; Clarke, S.; Goodacre, R.; Marshall, A.; et al. Assessing the impact of nitrogen supplementation in oats across multiple growth locations and years with targeted phenotyping and high-resolution metabolite profiling approaches. Food Chem. 2021, 355, 129585. [Google Scholar] [CrossRef] [PubMed]
- Stewart, D.; McDougall, G. Oat agriculture, cultivation and breeding targets: Implications for human nutrition and health. Br. J. Nutr. 2014, 112, 50–57. [Google Scholar] [CrossRef] [Green Version]
- Buerstmayr, H.; Krenn, N.; Stephan, U.; Grausgruber, H.; Zechner, E. Agronomic performance and quality of oat (Avena sativa L.) genotypes of worldwide origin produced under central European growing conditions. Field Crops Res. 2007, 101, 343–351. [Google Scholar] [CrossRef]
- Palágyi, A.; Andrea, P.; Móroczné, S. Different Species of Legumes Grown in Combination with Oats as Green Forage. Cereal Res. Commun. 2012, 40, 436–447. [Google Scholar] [CrossRef]
- Achleitner, A.; Tinker, N.A.; Zechner, E.; Hermann, B. Genetic diversity among oat varieties of worldwide origin and associations of AFLP markers with quantitative traits. Theor. Appl. Genet. 2008, 117, 1041–1053. [Google Scholar] [CrossRef]
- Cristina, M.V.; Peñas, E. Health benefits of oat: Current evidence and molecular mechanisms. Curr. Opin. Food Sci. 2017, 14, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.; Ziya, D.; Tevrican, D.; Aydin, A. Response of Six Oat (Avena Sativa L.) Cultivars to Nitrogen Fertilization for Agronomical Traits. Turk. J. Field Crops 2013, 18, 254–259. [Google Scholar]
- Wilson, M.L.; Evans, E.E.; Klossner, L.; Pagliari, P.H. Organic Oat Response to Variety, Seeding Rate, and Nutrient Source and Rate. Agronomy 2021, 11, 1418. [Google Scholar] [CrossRef]
- Andersson, A.A.; Börjesdottir, D. Effect of environment and variety on content and molecular weight of β-glucan in oats. J. Cereal Sci. 2011, 54, 122–128. [Google Scholar] [CrossRef]
- Ning, D.; Song, A.; Fan, F.; Li, Z.; Liang, Y. Effects of slag-based Silicon fertilizer on rice growth and brown-spot resistance. PLoS ONE 2014, 9, e102681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meena, V.; Dotaniya MCoumar, V.; Rajendiran Kumar, A.; Kundu, S.; Rao, A. A Case for Silicon Fertilization to Improve Crop Yields in Tropical Soils. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2014, 84, 508–518. [Google Scholar] [CrossRef] [Green Version]
- Artyszak, A. Effect of Silicon Fertilization on Crop Yield Quantity and Quality—A Literature Review in Europe. Plants 2018, 7, 54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, K.K.; Song, X.-P.; Tian, D.-D.; Guo, D.-J.; Chen, Z.-L.; Zhong, C.-S.; Nikpay, A.; Singh, M.; Rajput, V.D.; Singh, R.K.; et al. Influence of Silicon on Biocontrol Strategies to Manage Biotic Stress for Crop Protection, Performance, and Improvement. Plants 2021, 10, 2163. [Google Scholar] [CrossRef]
- Botta, A.; Rodrigues, F.A.; Sierras, N.; Marin, C.; Cerda, J.M.; Brossa, R. Evaluation of Armurox® (Complex of Peptides with soluble Silicon) on Mechanical and Biotic Stresses in Gramineae. In Proceedings of the 6th International Conference on Silicon in Agriculture, Stockholm, Sweden, 26–30 August 2014; p. 46. [Google Scholar]
- Dallagnol, L.J.; Roman, A.E.; Dorneles, K.d.R. Silicon Use in the Integrated Disease Management of Wheat: Current Knowledge. Available online: https://www.intechopen.com/online-first/74509 (accessed on 10 November 2021).
- Soratto, R.P.; Crusciol, C.A.; Castro, G.S.; Costa, C.H.; Ferrari, N.J. Leaf application of silicic acid to white oat and wheat. Rev. Bras. Ciênc. Solo 2012, 36, 1538–1544. [Google Scholar] [CrossRef]
- White, B.; Tubana, B.S.; Babu, T.; Mascagni, H., Jr.; Agostinho, F.; Datnoff, L.E.; Harrison, S. Effect of silicate slag application on wheat grown under two nitrogen rates. Plants 2017, 6, 47. [Google Scholar] [CrossRef] [Green Version]
- Hanafy Ahmed, A.H.; Harb, E.M.; Higazy, M.A.; Morgan, S.H. Effect of Silicon and boron foliar applications on wheat plants grown under saline soil conditions. Int. J. Agric. Res. 2008, 3, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Csajbók, J.; Kutasy, E.; Hunyadi Borbélyné, É.; Lesznyák, M.; Futó, Z.; Jakab, P. Effects of nutrient supply on the photosynthesis of maize hybrids. Cereal Res. Commun. 2005, 33, 169–172. [Google Scholar] [CrossRef]
- Kobra, M.; Yahya, E.; Muhammad, A. Influence of foliar application of Silicon on chlorophyll fluorescence, photosynthetic pigments, and growth in water-stressed wheat cultivars differing in drought tolerance. Turk. J. Bot. 2015, 39, 625–634. [Google Scholar] [CrossRef]
- Augustyniak, A.; Pawłowicz, I.; Lechowicz, K.; Izbiańska-Jankowska, K.; Arasimowicz-Jelonek, M.; Rapacz, M.; Perlikowski, D.; Kosmala, A. Freezing Tolerance of Lolium multiflorum/Festuca arundinacea Introgression Forms is Associated with the High Activity of Antioxidant System and Adjustment of Photosynthetic Activity under Cold Acclimation. Int. J. Mol. Sci. 2020, 21, 5899. [Google Scholar] [CrossRef]
- Ahmad, M.; El-Saeid, M.H.; Akram, M.A.; Ahmad, H.R.; Haroon, H.; Hussain, A. Silicon fertilization—A tool to boost up drought tolerance in wheat (Triticum aestivum L.) crop for better yield. J. Plant Nutr. 2016, 39, 1283–1291. [Google Scholar] [CrossRef]
- Csajbók, J.; Pepó, P.; Kutasy, E. Photosynthetic and Agronomic Traits of Winter Barley (Hordeum vulgare L.) Varieties. Agronomy 2020, 10, 1999. [Google Scholar] [CrossRef]
- Barczak, B.; Klikocka, H.; Kozera, W.; Knapowski, T. Assessment of the effect of Sulphur fertilisation on oat grain yield and micronutrient uptake. J. Elem. 2018, 23, 45–56. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Miralles, D.J. Radiation interception, biomass production and grain yield as affected by the interaction of nitrogen and sulfur fertilization in wheat. Eur. J. Agron. 2008, 28, 282–290. [Google Scholar] [CrossRef]
- Leilei, L.; Hongting, J.; Junpeng, A.; Kejia, S.; Jifeng, M.; Bing, L.; Liang, T.; Weixing, C.; Yan, Z. Response of biomass accumulation in wheat to low-temperature stress at jointing and booting stages. Environ. Exp. Bot. 2019, 157, 46–57. [Google Scholar] [CrossRef]
- Schaffer, A.A. Photoassimilate Distribution in Plants and Crops: Source–Sink Relationships; Werf, A., van der Zamski, E., Schaffer, A., Eds.; Marcel Dekker Inc.: New York, NY, USA, 1996; pp. 1–20. [Google Scholar]
- Sinclair, T.R.; Muchow, R.C. Radiation Use Efficiency. In Advances in Agronomy; Donald, L., Ed.; Academic Press: Cambridge, MA, USA, 1999; pp. 215–265. [Google Scholar] [CrossRef]
- Csajbók, J.; Kutasy, E. A transzspiráció és a fotoszintetikus aktivitás összefüggései kukoricában (Correlations between transpiration and photosynthetic activity in maize). In Talajtani, Vízgazdálkodási és Növénytermesztési Tudományos Nap; Lehoczky, É., Ed.; MTA ATK TAKI: Budapest, Hungary, 2012; pp. 151–154. (In Hungarian) [Google Scholar]
- Flint, L.E.; Flint, A.L. Downscaling future climate scenarios to fine scales for hydrologic and ecological modeling and analysis. Ecol. Process 2012, 1, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Yves, T.; Koutroulis, A.; Samaniego, L.; Vicente-Serrano, S.M.; Volaire, F.; Boone, A.; Le Page, M.; Llasat, M.C.; Albergel, C.; Burak, S.; et al. Challenges for drought assessment in the Mediterranean region under future climate scenarios. Earth-Sci. Rev. 2020, 210, 103348. [Google Scholar] [CrossRef]
- Collados-Lara, A.-J.; Pulido-Velazquez, D.; Pardo-Igúzquiza, E. An Integrated Statistical Method to Generate Potential Future Climate Scenarios to Analyse Droughts. Water 2018, 10, 1224. [Google Scholar] [CrossRef] [Green Version]
- Juhász, C.; Gálya, B.; Kovács, E.; Nagy, A.; Tamás, J.; Huzsvai, L. Seasonal predictability of weather and crop yield in regions of Central European continental climate. Comput. Electron. Agric. 2020, 173. [Google Scholar] [CrossRef]
- Jayawardana, H.A.R.K.; Weerahewa, H.L.D.; Saparamadu, M.D.J.S. Effect of root or foliar application of soluble Silicon on plant growth. Trop. Agric. Res. 2014, 26, 74–81. [Google Scholar] [CrossRef] [Green Version]
- Dinesh, J.; Shiva, D.; Anchal, D.; Sharma, V.K. Silicon and Phosphorus Fertilization in Aerobic Rice-Wheat System. In Proceedings of the 7th International Conference on Silicon in Agriculture, Bengaluru, India, 24–28 October 2017; p. 130. [Google Scholar]
- Frew, A.; Weston, L.A.; Reynolds, O.L.; Gurr, G.M. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef] [Green Version]
- Ciecierski, W. Effect of Silicon on Biotic and Abiotic Stress Mitigation in Horticultural and Field Crops. In Proceedings of the International Symposium “Mikroelementy w Rolnictwie i ’Srodowisku”, Kudowa-Zdrój, Poland, 21–24 June 2016; p. 25. [Google Scholar]
- Bocharnikova, E.A.; Loginov, S.V.; Matychenkov, V.V.; Storozhenko, P.A. Silicon fertilizer efficiency. Russ. Agric. Sci. 2010, 36, 446–448. [Google Scholar] [CrossRef]
- Farkas, Z.; Anda, A.; Vida, G.; Veisz, O.; Varga, B. CO2 Responses of Winter Wheat, Barley and Oat Cultivars under Optimum and Limited Irrigation. Sustainability 2021, 13, 9931. [Google Scholar] [CrossRef]
- Pan, L.; George-Jaeggli, B.; Borrell, A.; Jordan, D.; Koller, F.; Al-Salman, Y.; Ghannoum, O.; Cano, F.J. Coordination of stomata and vein patterns with leaf width underpins water use efficiency in a C4 crop. Plant Cell Environ. 2021. [CrossRef]
- Pandey, M. Effect of integrated nutrient management on yield, quality and uptake of nutrients in oat (Avena sativa) in alluvial soil. Ann. Plant Soil Res. 2018, 20, 1–6. [Google Scholar]
- Kurmanbayeva, M.; Sekerova, T.; Tileubayeva, Z.; Kaiyrbekov, T.; Kusmangazinov, A.; Shapalov, S.; Madenova, A.; Burkitbayev, M.; Bachilova, N. Influence of new sulfur-containing fertilizers on performance of wheat yield. Saudi J. Biol. Sci. 2021, 28, 4644–4655. [Google Scholar] [CrossRef]
- Klikocka, H.; Cybulska, M.; Barczak, B.; Narolski, B.; Szostak, B.; Kobiałka, A.; Nowak, A.; Wójcik, E. The effect of sulphur and nitrogen fertilization on grain yield and technological quality of spring wheat. Plant Soil Environ. 2016, 62, 230–236. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Song, F.; Xu, H.; Shao, H.; Song, R. Effects of Silicon on photosynthetic characteristics of maize (Zea mays L.) on alluvial soil. Sci. World J. 2014, 2014, 718716. [Google Scholar] [CrossRef] [Green Version]
- Amin, M.; Ahmad, R.; Ali, A.; Hussain, I.; Mahmood, R.; Aslam, M.; Lee, D.J. Influence of Silicon Fertilization on Maize Performance Under Limited Water Supply. Silicon 2018, 10, 177–183. [Google Scholar] [CrossRef]
- Sivakumar, D.; Jifon, J. Influence of Photoselective Shade Nettings on Postharvest Quality of Vegetables. In Preharvest Modulation of Postharvest Fruit and Vegetable Quality; Siddiqui, M.W., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 121–138. [Google Scholar]
- USS Working Group WRB. World Reference Base for Soil Resources 2014, International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; World Soil Resources Reports: Rome, Italy, 2015. [Google Scholar]
- Meier, U. Growth Stages of Mono and Dicotyledonous Plants, BBCH Monograph; Julius Kühn-Institut (JKI): Quedlinburg, Germany, 2018. [Google Scholar] [CrossRef]
- Von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- LI-COR, Inc. Li-6800 Portable Photosynthesis System, Software Version 1.2; LI-COR, Inc.: Lincoln, NE, USA, 2017. [Google Scholar]
- Tanner, C.B.; Sinclair, T.R. Efficient Water Use in Crop Production: Research or Re-Search. In Limitations to Efficient Water Use in Crop Production; Taylor, H.M., Jordan, W.R., Sinclair, T.R., Eds.; ASA: Madison, WI, USA, 1983; pp. 1–27. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration—Guidelines for Computing Crop Water Requirements—FAO Irrigation and Drainage Paper 56; FAO—Food and Agriculture Organization of the United Nations: Rome, Italy, 1998. [Google Scholar]
- Antal, E. Az éghajlatváltozás hatása a magyarországi aszályokra (Effect of climate change on drought in Hungary). Acta Geogr. Debrecina 1991, 28, 17–18. (In Hungarian) [Google Scholar]
- Szabó, A.; Tamás, J.; Nagy, A. The influence of hail net on the water balance and leaf pigment content of apple orchards. Sci. Hortic. 2021, 283, 110112. [Google Scholar] [CrossRef]
- Droppa, M.; Erdei, S.; Horváth, G.; Kissimom, J.; Mészáros, A.; Szalai, J.; Kosáry, J. Növénybiokémiai és élettani gyakorlatok (Plant Biochemistry and Physiology Practices); Budapest University of Economic Sciences and Public Administration: Budapest, Hungary, 2003; p. 88. (In Hungarian) [Google Scholar]
- Lichtenthaler, H.K.; Wellbum, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 603, 591–592. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ci | GSW | Transp | Tleaf-Tair | WUE | VPDleaf | |
|---|---|---|---|---|---|---|
| Ass | −0.355 ** | 0.876 ** | 0.888 ** | −0.667 ** | 0.447 ** | −0.698 ** |
| Ci | 1 | 0.440 ** | 0.328 ** | −0.461 ** | −0.745 ** | −0.548 ** |
| GSW | 0.440 ** | 1 | 0.937 ** | −0.801 ** | 0.058 | −0.873 ** |
| Transp | 0.328 ** | 0.937 ** | 1 | −0.789 ** | 0.013 | −0.706 ** |
| Tleaf-Tair | −0.461 ** | −0.801 ** | −0.789 ** | 1 | 0.103 * | 0.686 ** |
| WUE | −0.745 ** | 0.058 | 0.013 | 0.103 * | 1 | −0.108 * |
| Treatments | WUE (kg m−3) 27 May | WUE (kg m−3) 10 June | WUE (kg m−3) 24 June |
|---|---|---|---|
| Control | 5.27 ± 0.049 a | 10.55 ± 0.166 a | 2.37 ± 0.145 a |
| Si | 5.75 ± 0.042 b | 12.14 ± 0.152 b | 7.01 ± 0.176 b |
| S | 6.32 ± 0.051 c | 15.19 ± 0.167 c | 8.82 ± 0.177 c |
| Si + S | 6.25 ± 0.049 c | 14.41 ± 0.160 d | 6.83 ± 0.168 d |
| Varieties | WUE (kg m−3) 27 May | WUE (kg m−3) 10 June | WUE (kg m−3) 24 June |
|---|---|---|---|
| Mv Hópehely | 6.32 ± 0.059 a | 12.67 ± 0.199 a | 5.61 ± 0.201 a |
| Mv Kincsem | 6.27 ± 0.059 a | 11.83 ± 0.199 b | 5.28 ± 0.201 b |
| Mv Imperiál | 5.87 ± 0.051 b | 12.39 ± 0.173 c | 7.26 ± 0.174 c |
| Mv Istráng | 5.92 ± 0.059 c | 12.38 ± 0.198 c | 6.46 ± 0.201 d |
| GK Arany | 5.75 ± 0.058 d | 14.67 ± 0.195 d | 7.55 ± 0.198 e |
| GK Impala | 6.30 ± 0.072 a | 14.51 ± 0.244 d | 6.88 ± 0.247 f |
| Treatments | Control | Si | S | Si + S | ||||
|---|---|---|---|---|---|---|---|---|
| Varieties | Yield (kg ha−1) | TKW (g) | Yield (kg ha−1) | TKW (g) | Yield (kg ha−1) | TKW (g) | Yield (kg ha−1) | TKW (g) |
| GK Arany | 6662.5 a | 32.0 a | 7331.8 a | 34.7 a | 7041.5 a | 31.7 a | 7026.8 a | 34.7 a |
| GK Impala | 5593.0 b | 26.3 b | 6323.3 b | 24.3 b | 6228.6 b | 21.3 b | 6750.8 b | 23.7 b |
| Mv Hópehely | 6421.6 c | 35.0 c | 6469.2 b | 34.3 a | 6718.7 c | 32.7 a | 6366.1 c | 33.7 a |
| Mv Imperiál | 5487.0 b | 26.3 b | 6523.7 b | 28.0 c | 6198.2 b | 27.0 c | 6470.0 c | 30.3 c |
| Mv Istráng | 6747.9 a | 33.0 a | 6821.7 c | 34.3 a | 6382.8 b | 31.3 a | 6757.3 b | 34.0 a |
| Mv Kincsem | 5112.2 d | 27.0 b | 6222.2 b | 29.7 c | 5807.7 d | 29.0 c | 5520.5 d | 32.0 c |
| Layer 0–20 cm | Layer 20–40 cm | Layer 40–60 cm | |
|---|---|---|---|
| pH (H2O) | 8.30 | 8.36 | 8.43 |
| KA | 38 | 38 | 38 |
| CaCO3 (%) | 8.1 | 8.1 | 8.1 |
| Humus (%) | 3.66 | 2.92 | 2.70 |
| NO3 + NO2 (mg kg−1) | 1.71 | 2.95 | 3.18 |
| NH4 (mg kg−1) | 0.836 | 1.023 | 3.18 |
| P2O5 (AL) (mg kg−1) | 1671.6 | 1376.1 | 1076.8 |
| K2O (AL) (mg kg−1) | 658.9 | 648.2 | 525.5 |
| SO4 (mg kg−1) | 3.07 | 6.00 | 7.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutasy, E.; Buday-Bódi, E.; Virág, I.C.; Forgács, F.; Melash, A.A.; Zsombik, L.; Nagy, A.; Csajbók, J. Mitigating the Negative Effect of Drought Stress in Oat (Avena sativa L.) with Silicon and Sulphur Foliar Fertilization. Plants 2022, 11, 30. https://doi.org/10.3390/plants11010030
Kutasy E, Buday-Bódi E, Virág IC, Forgács F, Melash AA, Zsombik L, Nagy A, Csajbók J. Mitigating the Negative Effect of Drought Stress in Oat (Avena sativa L.) with Silicon and Sulphur Foliar Fertilization. Plants. 2022; 11(1):30. https://doi.org/10.3390/plants11010030
Chicago/Turabian StyleKutasy, Erika, Erika Buday-Bódi, István Csaba Virág, Fanni Forgács, Anteneh Agezew Melash, László Zsombik, Attila Nagy, and József Csajbók. 2022. "Mitigating the Negative Effect of Drought Stress in Oat (Avena sativa L.) with Silicon and Sulphur Foliar Fertilization" Plants 11, no. 1: 30. https://doi.org/10.3390/plants11010030
APA StyleKutasy, E., Buday-Bódi, E., Virág, I. C., Forgács, F., Melash, A. A., Zsombik, L., Nagy, A., & Csajbók, J. (2022). Mitigating the Negative Effect of Drought Stress in Oat (Avena sativa L.) with Silicon and Sulphur Foliar Fertilization. Plants, 11(1), 30. https://doi.org/10.3390/plants11010030