
Chemical Composition, Production of Secondary Metabolites and Antioxidant Activity in Coffee Cultivars Susceptible and Partially Resistant to Bacterial Halo Blight

 ,
, 
Abstract
1. Introduction
2. Results
2.1. Chemical Composition and Antioxidant Activity of Coffee Leaf Extracts from Mundo Novo 376/4 and Iapar-59 Cultivars
2.2. Production of Secondary Metabolites in Iapar-59 Coffee Leaves Inoculated with P. syringae pv. garcae
3. Discussion
4. Material and Methods
4.1. Chemical Composition and Antioxidant Activity of Coffee Leaf Extracts from Mundo Novo 376/4 and Iapar-59 Cultivars
4.1.1. Preparation of Coffee Seedlings
4.1.2. Preparation of Aqueous Extracts from Coffee Leaves
4.1.3. Determination of the Contents of Total Phenolics and Flavonoids
4.1.4. Determination of the Phenolic Compound Profile by High Performance Liquid Chromatography
4.1.5. Antioxidant Activity: Free Radical Scavenging Activity (DPPH) and Ferric Reducing Antioxidant Power (FRAP)
4.2. Production of Secondary Metabolites in Iapar-59 Coffee Leaves Inoculated with P. syringae pv. garcae
4.2.1. Inoculation with P. syringae pv. garcae
4.2.2. Quantification of Total Phenolic and Flavonoid Compounds, Reducing Sugars and Determination of Phenolic Profile in Leaves
4.2.3. Severity Disease in Mundo Novo and Iapar-59 Cultivars
4.3. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taiz, L. Fisiologia Vegetal; Artmed: Porto Alegre, Brazil, 2010; ISBN 9788536316147. [Google Scholar]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic Compounds and Their Role in Disease Resistance. Annu. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Riaz, U.; Kharal, M.A.; Murtaza, G.; Zaman, Q.; Javaid, S.; Malik, H.A.; Aziz, H.; Abbas, Z. Prospective Roles and Mechanisms of Caffeic Acid in Counter Plant Stress: A Mini Review. PJAR 2018, 32, 8–19. [Google Scholar] [CrossRef]
- Treutter, D. Significance of Flavonoids in Plant Resistance and Enhancement of Their Biosynthesis. Plant. Biol. 2005, 7, 581–591. [Google Scholar] [CrossRef]
- Vargas, P.; Farias, G.A.; Nogales, J.; Prada, H.; Carvajal, V.; Barón, M.; Rivilla, R.; Martín, M.; Olmedilla, A.; Gallegos, M.-T. Plant Flavonoids Target Pseudomonas Syringae Pv. Tomato DC3000 Flagella and Type III Secretion System: Flavonoids Affect Pto DC3000 Virulence. Environ. Microbiol. Rep. 2013, 5, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Morkunas, I.; Ratajczak, L. The Role of Sugar Signaling in Plant Defense Responses against Fungal Pathogens. Acta Physiol. Plant 2014, 36, 1607–1619. [Google Scholar] [CrossRef]
- Moghaddam, M.R.B.; Van den Ende, W. Sugars and Plant Innate Immunity. J. Exp. Bot. 2012, 63, 3989–3998. [Google Scholar] [CrossRef]
- Rojas, C.M.; Senthil-Kumar, M.; Tzin, V.; Mysore, K.S. Regulation of Primary Plant Metabolism during Plant-Pathogen Interactions and Its Contribution to Plant Defense. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef]
- Kang, H.-M.; Saltveit, M.E. Reduced Chilling Tolerance in Elongating Cucumber Seedling Radicles Is Related to Their Reduced Antioxidant Enzyme and DPPH-Radical Scavenging Activity. Physiol. Plant. 2002, 115, 244–250. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Devaki, M. The ferric reducing/antioxidant power (FRAP) assay for non-enzymatic antioxidant capacity: Concepts, procedures, limitations and applications. In Measurement of Antioxidant Activity & Capacity; Apak, R., Capanoglu, E., Shahidi, F., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2017; pp. 77–106. ISBN 9781119135388. [Google Scholar]
- Thite, S.V.; Aparadht, V.T.; Kore, S.A. Effect of powdery mildew infection on DPPH radical scavenging activity and ferric-re ducing antioxidant power of plants. World J. Pharm. Res. 2013, 2, 1–6. [Google Scholar]
- Ramiro, D.A.; Guerreiro-Filho, O.; Mazzafera, P. Phenol Contents, Oxidase Activities, and the Resistance of Coffee to the Leaf Miner Leucoptera Coffeella. J. Chem. Ecol. 2006, 32, 1977–1988. [Google Scholar] [CrossRef]
- Melo, G.A.; Shimizu, M.M.; Mazzafera, P. Polyphenoloxidase Activity in Coffee Leaves and Its Role in Resistance against the Coffee Leaf Miner and Coffee Leaf Rust. Phytochemistry 2006, 67, 277–285. [Google Scholar] [CrossRef] [PubMed]
- Kempthorne, C.J.; Nielsen, A.J.; Wilson, D.C.; McNulty, J.; Cameron, R.K.; Liscombe, D.K. Metabolite Profiling Reveals a Role for Intercellular Dihydrocamalexic Acid in the Response of Mature Arabidopsis thaliana to Pseudomonas syringae. Phytochemistry 2021, 187, 112747. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, X.; Zeng, Y.; Liu, P. Metabolic Profiling Reveals Local and Systemic Responses of Kiwifruit to Pseudomonas Syringae Pv. Actinidiae. Plant Direct 2020, 4, e00297. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Langlois-Meurinne, M.; Bellvert, F.; Garmier, M.; Didierlaurent, L.; Massoud, K.; Saindrenan, P. The differential spatial distribution of secondary metabolites in Arabidopsis leaves reacting hypersensitively to Pseudomonas syringae pv. tomato is dependent on the oxidative burst. J. Exp. Bot. 2010, 61, 3355–3370. [Google Scholar] [CrossRef] [PubMed]
- Badel, J.L.; Zambolim, L. Coffee Bacterial Diseases: A Plethora of Scientific Opportunities. Plant Pathol. 2019, 68, 411–425. [Google Scholar] [CrossRef]
- Amaral, J.F.; Teixeira, C.G.; Pinheiro, E.D. Obacterio causador da Mancha Aureolada Do Cafeeiro. Arq. Inst. Biológico 1956, 23, 151–155. [Google Scholar]
- Young, J.M.; Dye, D.W.; Bradbury, J.F.; Panagopoulos, C.G.; Robbs, C.F. A Proposed Nomenclature and Classification for Plant Pathogenic Bacteria. N. Z. J. Agric. Res. 1978, 21, 153–177. [Google Scholar] [CrossRef]
- Zoccoli, D.M.; Takatsu, A.; Uesugi, C.H. Ocorrência de Mancha Aureolada Em Cafeeiros Na Região Do Triângulo Mineiro e Alto Paranaíba. Bragantia 2011, 70, 843–849. [Google Scholar] [CrossRef]
- Sera, G.H.; Sera, T.; Fazuoli, L.C. IPR 102—Dwarf Arabica Coffee Cultivar with Resistance to Bacterial Halo Blight. Crop. Breed. Appl. Biotechnol. 2017, 17, 403–407. [Google Scholar] [CrossRef]
- Andreazi, E.; Sera, G.H.; Sera, T.; Fonseca, I.C.d.B.; Carducci, F.C.; Shigueoka, L.H.; dos Santos, W.G.; Pereira, C.T.M. Resistance to Bacterial Halo Blight in Arabica Coffee Lines Derivative from the Genotype C1195-5-6-2 under Natural Infection Conditions. Crop. Breed. Appl. Biotechnol. 2018, 18, 110–115. [Google Scholar] [CrossRef]
- Petek, M.R.; Sera, T.; Sera, G.H.; Fonseca, I.C.d.B.; Ito, D.S. Seleção de Progênies de Coffea Arabica Com Resistência Simultânea à Mancha Aureolada e à Ferrugem Alaranjada. Bragantia 2006, 65, 65–73. [Google Scholar] [CrossRef]
- Ito, D.S.; Sera, T.; Sera, G.H.; Grossi, L.D.; Kanayama, F.S. Resistance to Bacterial Blight in Arabica Coffee Cultivars. CBAB 2008, 8, 99–103. [Google Scholar] [CrossRef]
- Sera, G.H.; Sera, T.; Ito, D.S.; Fonseca, I.C.d.B.; Kanayama, F.S.; Grossi, L.D.; Shigueoka, L.H. Seleção para a resistência à ferrugem em progênies das cultivares de café IPR 99 e IPR 107. Bragantia 2010, 69, 547–554. [Google Scholar] [CrossRef][Green Version]
- Karamać, M.; Kosińska, A.; Pegg, R.B. Content of gallic acid in selected plant extracts. Pol. J. Food Nutr. Sci. 2006, 15, 55–58. [Google Scholar]
- Ngamsuk, S.; Huang, T.-C.; Hsu, J.-L. Determination of Phenolic Compounds, Procyanidins, and Antioxidant Activity in Processed Coffea Arabica L. Leaves. Foods 2019, 8, 389. [Google Scholar] [CrossRef] [PubMed]
- Lashermes, P.; Andrzejewski, S.; Bertrand, B.; Combes, M.C.; Dussert, S.; Graziosi, G.; Trouslot, P.; Anthony, F. Molecular Analysis of Introgressive Breeding in Coffee (Coffea Arabica L.). Theor. Appl. Genet. 2000, 100, 139–146. [Google Scholar] [CrossRef]
- Carvalho, F.G.; Sera, G.H.; Andreazi, E.; Sera, T.; Fonseca, I.C.D.B.; Carducci, F.C.; Shigueoka, L.H.; Holderbaum, M.M.; Costa, K.C. Tolerância Ao Déficit Hídrico Em Mudas de Genótipos de Café Portadores de Genes de Diferentes Espécies. C. Sci. 2017, 12, 156. [Google Scholar] [CrossRef]
- Herrera, J.C.; Lambot, C. The Coffee Tree—Genetic Diversity and Origin. In The Craft and Science of Coffee; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–16. ISBN 9780128035207. [Google Scholar]
- Roussos, P.A.; Pontikis, C.A.; Tsantili, E. Root Promoting Compounds Detected in Olive Knot Extract in High Quantities as a Response to Infection by the Bacterium Pseudomonas Savastanoi Pv. Savastanoi. Plant. Sci. 2002, 163, 533–541. [Google Scholar] [CrossRef]
- Cayuela, J.A.; Rada, M.; Rios, J.J.; Albi, T.; Guinda, A. Changes in Phenolic Composition Induced by Pseudomonas Savastanoi Pv. Savastanoi Infection in Olive Tree: Presence of Large Amounts of Verbascoside in Nodules of Tuberculosis Disease. J. Agric. Food Chem. 2006, 54, 5363–5368. [Google Scholar] [CrossRef] [PubMed]
- Heath, M.C. The Absence of Active Defense Mechanisms in Compatible Hostpathogen Interactions. In Active Defense Mechanisms in Plants; Wood, R.K.S., Ed.; Plenum Press: New York, NY, USA, 1982; pp. 143–156. [Google Scholar]
- Markakis, E.A.; Tjamos, S.E.; Antoniou, P.P.; Roussos, P.A.; Paplomatas, E.J.; Tjamos, E.C. Phenolic responses of resistant and susceptible olive cultivars induced by defoliating and nondefoliating Verticillium dahliae pathotypes. Plant Dis. 2010, 94, 1156–1162. [Google Scholar] [CrossRef]
- Rodrigues, F.Á.; Carré-Missio, V.; Jham, G.N.; Berhow, M.; Schurt, D.A. Chlorogenic acid levels in leaves of coffee plants supplied with silicon and infected by Hemileia vastatrix. Trop. Plant. Pathol. 2011, 36. [Google Scholar] [CrossRef]
- Maddox, C.E.; Laur, L.M.; Tian, L. Antibacterial Activity of Phenolic Compounds against the Phytopathogen Xylella Fastidiosa. Curr. Microbiol. 2010, 60, 53–58. [Google Scholar] [CrossRef]
- Dadáková, K.; Heinrichová, T.; Lochman, J.; Kašparovský, T. Production of Defense Phenolics in Tomato Leaves of Different Age. Molecules 2020, 25, 4952. [Google Scholar] [CrossRef]
- Zhou, X.; Jia, H.; Ge, X.; Wu, F. Effects of Vanillin on the Community Structures and Abundances of Fusarium and Trichoderma Spp. in Cucumber Seedling Rhizosphere. J. Plant Interact. 2018, 13, 45–50. [Google Scholar] [CrossRef]
- Moreira, M.E.d.C.; Pereira, R.G.F.A.; Dias, D.F.; Gontijo, V.S.; Vilela, F.C.; de Moraes, G.d.O.I.; Giusti-Paiva, A.; dos Santos, M.H. Anti-Inflammatory Effect of Aqueous Extracts of Roasted and Green Coffea Arabica L. J. Funct. Foods 2013, 5, 466–474. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1999; Volume 299, pp. 152–178. ISBN 9780121822002. [Google Scholar]
- Kalia, K.; Sharma, K.; Singh, H.P.; Singh, B. Effects of Extraction Methods on Phenolic Contents and Antioxidant Activity in Aerial Parts of Potentilla Atrosanguinea Lodd. and Quantification of Its Phenolic Constituents by RP-HPLC. J. Agric. Food Chem. 2008, 56, 10129–10134. [Google Scholar] [CrossRef]
- Aquino, F.W.B.; Rodrigues, S.; do Nascimento, R.F.; Casimiro, A.R.S. Simultaneous Determination of Aging Markers in Sugar Cane Spirits. Food Chem. 2006, 98, 569–574. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a Free Radical Method to Evaluate Antioxidant Activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and Mechanisms of Antioxidant Activity Using the DPPH. Free Radical Method. LWT—Food Sci. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Belan, L.L.; Pozza, E.A.; Freitas, M.L.d.O.; de Souza, R.M.; Junior, W.C.d.J.; Oliveira, J.M. Diagrammatic Scale for Assessment of Bacterial Blight in Coffee Leaves. J. Phytopathol. 2014, 162, 801–810. [Google Scholar] [CrossRef]


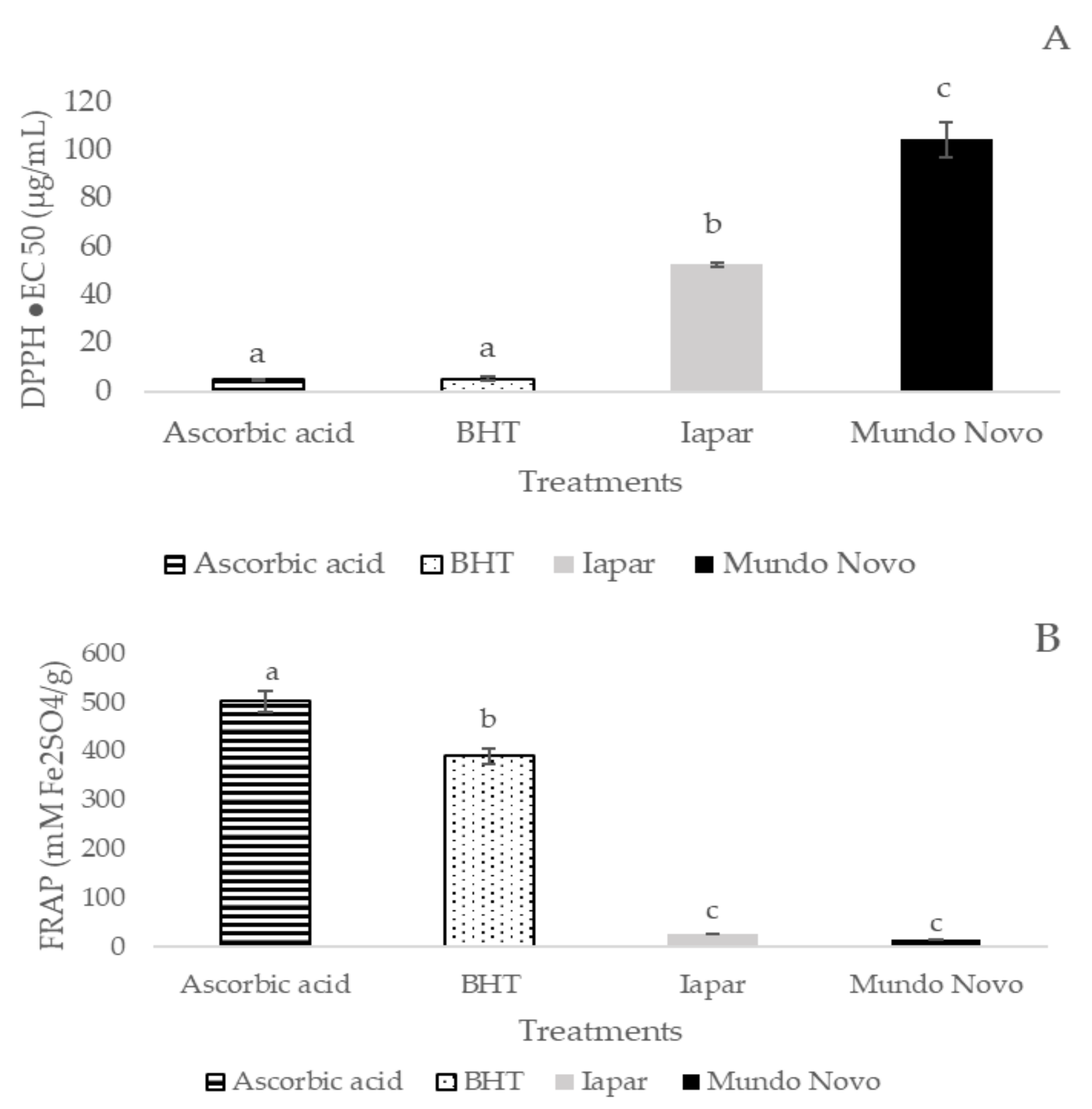{kind=link}
| Cultivar | Phenolic Compounds | Flavonoids | Gallic Acid | Chlorogenic Acid | Caffeic Acid |
|---|---|---|---|---|---|
| Iapar-59 | 597.84 a | 31.87 a | 47.24 a | 1.93 a | 1487.42 a |
| Mundo Novo 376/4 | 394.54 b | 17.11 b | 27.14 b | 1.51 a | 975.11 b |
| Treatments | Reducing Sugars | Flavonoids | Phenolic Compound | Gallic Acid | Chlorogenic Acid | Caffeic Acid | Vanillin |
|---|---|---|---|---|---|---|---|
| Inoculated | 53.86 b | 52.50 b | 397.38 b | 72.99 a | 3.04 a | 1697.29 a | 20.19 |
| Non-inoculated | 89.29 a | 120.19 a | 755.52 a | 47.24 b | 1.93 b | 1482.42 b | Nd |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, J.A.G.; de Resende, M.L.V.; Ribeiro, I.S.; Lima, A.R.; Albuquerque, L.R.M.; Monteiro, A.C.A.; Pereira, M.H.B.; Botelho, D.M.d.S. Chemical Composition, Production of Secondary Metabolites and Antioxidant Activity in Coffee Cultivars Susceptible and Partially Resistant to Bacterial Halo Blight. Plants 2021, 10, 1915. https://doi.org/10.3390/plants10091915
da Silva JAG, de Resende MLV, Ribeiro IS, Lima AR, Albuquerque LRM, Monteiro ACA, Pereira MHB, Botelho DMdS. Chemical Composition, Production of Secondary Metabolites and Antioxidant Activity in Coffee Cultivars Susceptible and Partially Resistant to Bacterial Halo Blight. Plants. 2021; 10(9):1915. https://doi.org/10.3390/plants10091915
Chicago/Turabian Styleda Silva, Joyce Alves Goulart, Mário Lúcio Vilela de Resende, Ingridy Simone Ribeiro, Adriene Ribeiro Lima, Luiz Roberto Marques Albuquerque, Ana Cristina Andrade Monteiro, Matheus Henrique Brito Pereira, and Deila Magna dos Santos Botelho. 2021. "Chemical Composition, Production of Secondary Metabolites and Antioxidant Activity in Coffee Cultivars Susceptible and Partially Resistant to Bacterial Halo Blight" Plants 10, no. 9: 1915. https://doi.org/10.3390/plants10091915
APA Styleda Silva, J. A. G., de Resende, M. L. V., Ribeiro, I. S., Lima, A. R., Albuquerque, L. R. M., Monteiro, A. C. A., Pereira, M. H. B., & Botelho, D. M. d. S. (2021). Chemical Composition, Production of Secondary Metabolites and Antioxidant Activity in Coffee Cultivars Susceptible and Partially Resistant to Bacterial Halo Blight. Plants, 10(9), 1915. https://doi.org/10.3390/plants10091915