
Advanced Applications for Protein and Compounds from Microalgae




Abstract
:1. Introduction
2. Microalgae Engineering to Obtain Platforms for Biocompounds Productions

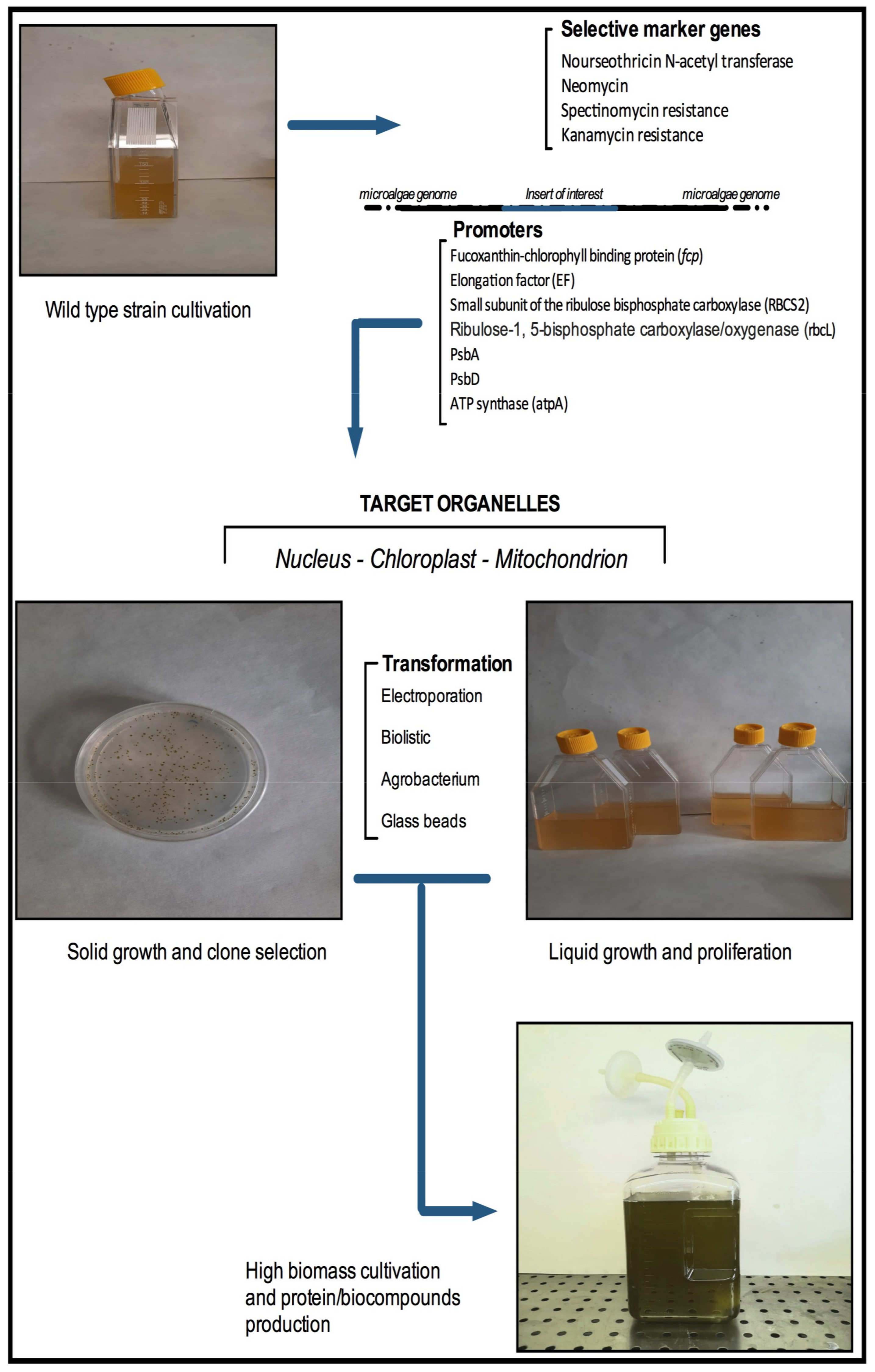{kind=link}
| Host Organisms | Gene | Donor Organism | Enzyme | Approach | Effects | References |
|---|---|---|---|---|---|---|
| Phaeodactylum tricornutum | PtGPAT2 | P. tricornutum | Glycerol-3-phosphate acyltransferase 2 | Overexpression | Hyperaccumulation of TAG | [20] |
| Phaeodactylum tricornutum | PtPGM | P. tricornutum | Phosphoglucomutase | Overexpression | Increased synthesis of chrysolaminarin | [12] |
| Phaeodactylum tricornutum | OsElo5 | O. tauri | Δ5-elongase | Endogenous expression | Improved accumulation of EPA and DHA—dark cultivation | [46] |
| Phaeodactylum tricornutum | PpGT | P. patens | Glucose transporter | Endogenous expression | Improved accumulation of EPA and DHA—dark cultivation | [46] |
| Phaeodactylum tricornutum | PtTL | _ | TAG lipase | Knockdown | Hyperaccumulation of TAG | [27] |
| Thalassiosira pseudonana | TpTL | _ | TAG lipase | Knockdown | Hyperaccumulation of TAG | [26] |
| Phaeodactylum tricornutum | PtDGAT2B | P. tricornutum | 2 acyl-CoA:diacylglycerol acyltransferase | Endogenous overexpression | Increased DHA and TAG content | [53] |
| Phaeodactylum tricornutum | PtG6PDH | P. tricornutum | Glucose-6-phosphate dehydrogenase | Overexpression | Enhanced lipid and w-3 accumulation | [39] |
| Phaeodactylum tricornutum | AnPhyA | A. niger | Phytase | Endogenous expression | Improved accumulation of EPA and DHA | [48] |
| Phaeodactylum tricornutum | EcAppA | E. coli | Phytase | Endogenous expression | Improved accumulation of EPA and DHA | [48] |
| Phaeodactylum tricornutum | PtMCAT | P. tricornutum | Malonyl CoA-acyl carrier protein transacylase | Overexpression | Improved accumulation of EPA | [45] |
| Phaeodactylum tricornutum | PtFAD5b | P. tricornutum | Fatty acid desaturase 5b | Overexpression | Improved accumulation of EPA | [45] |
| Phaeodactylum tricornutum | PtGPAT1 | P. tricornutum | Glycerol-3-phosphate acyltransferase | Overexpression | Increased lipid content | [19] |
| Phaeodactylum tricornutum | PtLPAAT1 | P. tricornutum | Lysophosphatidic acid acyltransferase | Overexpression | Increased lipid content | [19] |
| Phaeodactylum tricornutum | PtPTP | P. tricornutum | Plastidial pyruvate transporter | Overexpression | Increased production of biomass and lipids | [54] |
| Phaeodactylum tricornutum | PtTES1 | _ | Hotdog-fold thioesterase | TALEN- mutagenesis | Hyperaccumulation of TAG | [28] |
| Chlorella ellipsoidea | AtLEC1 | A. thaliana | Leafy cotyledon 1 transcription factor | Endogenous expression | Lipid overexpression | [33] |
| Synechocystis sp. | SaACC | S. alba | Acetyl-CoA carboxylase | Endogenous expression | Lipid overexpression | [55] |
| Chlamydomonas reinhardtii | CrGAPDH | Ch. reinhardtii | Glyceraldehyde-3-phosphate dehydrogenase | Overexpression | Enhanced carbon fixation | [56] |
| Chlorella vulgaris | CvNR | _ | Nitrate reductase | CRISP-cas9 editing | Reduced growth upon specific conditions | [57] |
| Chlorella vulgaris | CvAPT | _ | Adenine phosphoribosyltransferase | CRISP-cas9 editing | Reduced growth upon specific conditions | [57] |
| Nannochloropsis oceanica | AtDXS | A. thaliana | 1-deoxy-D-xylulose 5-phosphate synthase | Endogenous expression | Improved CO2 absorption, biomass and lipids | [34] |
| Nannochloropsis salina | NsME | N. salina | Malic enzyme | Overexpression | Increased production of lipids and fatty acids | [35] |
| Nannochloropsis salina | CrLCIA | Ch. reinhardtii | Anion transporter | Endogenous expression | Increased production of fatty acids | [36] |
| Nannochloropsis oceanica | NoAPL | - | Apetala 2 like transcription factor | Insertional mutagenesis | Increased production of lipids | [37] |
| Chlamydomonas reinhardtii | CrSBP1 | C. reinhardtii | Sedoheptulose-1,7-bisphosphatase | Overexpression | Photosynthetic and growth rates improvement | [58] |
| Neochloris oleoabundans | NoGPAT | _ | Glycerol-3-phosphate acyltransferase | Overexpression | Increased lipid content | [25] |
| Neochloris oleoabundans | NoLPAAT | _ | Lysophosphatidic acid acyltransferase | Overexpression | Increased lipid content | [25] |
| Porphyridium purpureum | PpCHS1 | _ | Chlorophyll synthase | CRISP-cas9 editing | Increased phycoerythrin content | [50] |
| Haematococcus pluvialis | PkHUP1 | Parachlorella kesslerii | Hexose uptake protein | Endogenous overexpression | Dark cultivation | [52] |
| Chlorella pyrenoidosa | PtG6PDH | P. tricornutum | Glucose-6-phosphate dehydrogenase | Endogenous overexpression | Increased lipid content | [38] |
| Chlorella pyrenoidosa | NoG6PDH | N. oceanica | Glucose-6-phosphate dehydrogenase | Endogenous overexpression | Increased lipid content | [38] |
3. Microalgae Engineering for the Production of Pharmacological Proteins
| Organism host | Product | Application | Transformation Method | Localization | Outcome | Expression Yields | References |
|---|---|---|---|---|---|---|---|
| Ch. reinhardtii | E2 protein | Swine fever virus vaccine | Biolistic | Chloroplast | Strong immunogenic response in mice | 1.5–2% TSP | [74] |
| Ch. reinhardtii | VP28 | White spot syndrome virus vaccine | Glass bead | Chloroplast | Shrimp survival up to 87% | ND | [75] |
| D. salina | VP28 | White spot syndrome virus vaccine | Glass bead | Chloroplast | 59% protection rate | 78 mg/100 culture | [76] |
| Ch. reinhardtii | VP28 | Chloroplast | Up to 10% TSP | [69] | |||
| Ch. reinhardtii | Antiviral dsRNA | Yellow head virus RNAi-based vaccine | Glass beads | Chloroplast | Reduced mortality | Up to 16 ng dsRNA/L culture | [77] |
| Ch. reinhardtii | dsRNA-YHV | Yellow head virus antiviral | Glass beads | Nucleus | 22% Shrimp survival | 45 ng/100-mL culture | [78] |
| Chlorella sp. | AMPs Scy-hepc | A. hydrophila bacteria oral antibiotics | Electroporation | Nucleus | In vitro inhibitory effects on A. hydrophila; in vivo S. macrocephalus | Up to 0.078%TSP | [79] |
| H. pluvialis | Piscidin-4 peptide | Antibacterial activity | Biolistic | Chloroplast | Stable expression | ND | [81] |
| Nannochloropsis sp | OmpK fragment gene | Vibrio species oral vaccine | Nucleus | Fifth generation stable immunogenic peptide production | ND | [80] | |
| Ch. reinhardtii | Mytichitin-A peptide | Antibacterial activity | Electroporation | Nucleus | High inhibition of bacteria growth (MIC assays); No toxicity on HEK293 cells. | 0.28% TSP | [60] |
| Ch. reinhardtii | SARS-CoV-2-RBD | Antigen proteins against SARS-CoV-2 | Geminiviral vector | Transient | ELISA assay showed specific binding with the anti-RBD antibody | 1.14 µg/g | [83] |
| Ch. vulgaris | SARS-CoV-2-RBD | Antigen proteins against SARS-CoV-2 | Geminiviral vector | Transient | ELISA assay showed specific binding with the anti-RBD antibody | 1.161 µg/g | [83] |
| Ch. reinhardtii | SARS-CoV-2-RBD | Antigen proteins against SARS-CoV-2 | Electroporation | Transient | Bind human ACE2 receptor | 0.1% TSP | [84] |
| Ch. reinhardtii | Human Interferon-α | Chronic viral diseases and cancers treat | Agrobacterium | Nucleus | In vivo e in vitro antitumoral activity, anticancer proprieties, antiviral activity | ND | [82] |
| Ch. reinhardtii, | Human interleukin-2 | Interleukin production | Agrobacterium | Nucleus | ELISA assay showed biological activity, high stability | Up to 0.94% TSP | [72] |
| D. salina; C. vulgaris | Human interleukin-2 | Interleukin production | Agrobacterium | Nucleus | ELISA assay showed biological activity, high stability | Up to 0.59% TSP | [72] |
| Schizochytrium sp. | ZK antigen | Zika virus oral vaccine | Algevir technology | Transient | IgG and IgA production | Up to 365.3 μg g−1 FW | [85] |
| Schizochytrium sp. | LTB:RAGE antigen | Alzheimer’s disease vaccine | Algevir technology | Transient | ELISA assay showed high stability up to of 60 °C | Up to 380 μg g−1 FW | [86] |
| Ch. reinhardtii, | PfCelTOS antigen | Malaria antigen for diagnosis tests | Biolistic | Chloroplast | Stable expression | ND | [64] |
| Schizochytrium sp. | Multiepitope protein (BCB) | Breast cancer vaccine | Algevir technology | Transient | Tumor cell line 4T1 reactivity; IgG production in mice immunized with BCB | Up to 637 μg/g | [87] |
| P. tricornutum | Hepatitis B Antibody | Antibodies against Hepatitis B | Biolistic | Nucleus | Binding FcγRI | 2 mg/L | [71] |
| P. tricornutum | Monoclonal antibodies | Antibodies against Marburg virus | Biolistic | Nucleus | Elisa assay showed binding efficiency | 1300 ng/ml | [88] |
| T. pseudonana | Antibody for EA1 | Biosensor for anthracis detection | Biolistic | Nucleus | Detection of detected EA1 epitope in lysed spores | ND | [61] |
4. Extremophilic Microalgae as Bioreactors
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fischer, R.; Emans, N.; Schuster, F.; Hellwig, S.; Drossard, J. Towards molecular farming in the future: Using plant-cell-suspension cultures as bioreactors. Biotechnol. Appl. Biochem. 1999, 30, 109–112. [Google Scholar]
- Fischer, R.; Stoger, E.; Schillberg, S.; Christou, P.; Twyman, R.M. Plant-based production of biopharmaceuticals. Curr. Opin. Plant Biol. 2004, 7, 152–158. [Google Scholar] [CrossRef]
- Lau, O.S.; Sun, S.S.M. Plant seeds as bioreactors for recombinant protein production. Biotechnol. Adv. 2009, 27, 1015–1022. [Google Scholar] [CrossRef]
- Liang, Z.C.; Liang, M.H.; Jiang, J.G. Transgenic microalgae as bioreactors. Crit. Rev. Food Sci. Nutr. 2020, 60, 3195–3213. [Google Scholar] [CrossRef]
- De Zoeten, G.; Penswick, J.; Horisberger, M.; Ahl, P.; Schultze, M.; Hohn, T. The expression, localization, and effect of a human interferon in plants. Virology 1989, 172, 213–222. [Google Scholar] [CrossRef]
- Hiatt, A.; Cafferkey, R.; Bowdish, K. Production of antibodies in transgenic plants. Nature 1989, 342, 76–78. [Google Scholar] [CrossRef]
- Shanmugaraj, B.; Bulaon, C.J.I.; Phoolcharoen, W. Plant molecular farming: A viable platform for recombinant biopharmaceutical production. Plants 2020, 9, 842. [Google Scholar] [CrossRef]
- Foley, J.A.; Defries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamed, I. The evolution and versatility of microalgal biotechnology: A review. Compr. Rev. Food Sci. Food Saf. 2016, 15, 1104–1123. [Google Scholar] [CrossRef] [PubMed]
- Hashemian, M.; Ahmadzadeh, H.; Hosseini, M.; Lyon, S.; Pourianfar, H. Production of microalgae-derived high-protein biomass to enhance food for animal feedstock and human consumption. Adv. Bioprocess. Altern. Fuels Biobased Chem. Bioprod. 2019, 393–405. [Google Scholar] [CrossRef]
- Gallo, C.; Landi, S.; d’Ippolito, G.; Nuzzo, G.; Manzo, E.; Sardo, A.; Fontana, A. Diatoms synthesize sterols by inclusion of animal and fungal genes in the plant pathway. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Yang, R.; Wei, D.; Xie, J. Diatoms as cell factories for high-value products: Chrysolaminarin, eicosapentaenoic acid, and fucoxanthin. Crit. Rev. Biotechnol. 2020, 40, 993–1009. [Google Scholar] [CrossRef]
- Zhao, C.; Yaschenko, A.; Alonso, J.M.; Stepanova, A.N. Leveraging synthetic biology approaches in plant hormone research. Curr. Opin. Plant. Biol. 2021, 60. [Google Scholar] [CrossRef]
- George, J.; Kahlke, T.; Abbriano, R.M.; Kuzhiumparambil, U.; Ralph, P.J.; Fabris, M. Metabolic engineering strategies in diatoms reveal unique phenotypes and genetic configurations with implications for algal genetics and synthetic biology. Front. Bioeng. Biotechnol. 2020, 8. [Google Scholar] [CrossRef] [PubMed]
- Rosales-Mendoza, S.; García-Silva, I.; González-Ortega, O.; Sandoval-Vargas, J.M.; Malla, A.; Vimolmangkang, S. The potential of algal biotechnology to produce antiviral compounds and biopharmaceuticals. Molecules 2020, 25, 49. [Google Scholar] [CrossRef] [PubMed]
- Vella, F.M.; Sardo, A.; Gallo, C.; Landi, S.; Fontanta, A.; d’Ippolito, G. Annual outdoor cultivation of the diatom Thalassiosira weissflogii: Productivity, limits and perspectives. Algal Res. 2019, 42. [Google Scholar] [CrossRef]
- Landi, S.; Esposito, S. Bioinformatic characterization of sulfotransferase provides new insights for the exploitation of sulfated polysaccharides in Caulerpa. Int. J. Mol. Sci. 2020, 21, 681. [Google Scholar] [CrossRef] [PubMed]
- Fabris, M.; Abbriano, R.M.; Pernice, M.; Sutherland, D.L.; Commault, A.S.; Hall, C.C.; Labeeuw, L.; McCauley, J.I.; Kuzhiuparambil, U.; Ray, P.; et al. Emerging technologies in algal biotechnology: Toward the establishment of a sustainable, algae-based bioeconomy. Front. Plant Sci. 2020, 11, 279. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Dong, H.P.; Wei, W.; Balamurugan, S.; Yang, W.D.; Liu, J.S.; Li, H.Y. Dual expression of plastidial GPAT1 and LPAT1 regulates triacylglycerol production and the fatty acid profile in Phaeodactylum tricornutum. Biotechnol. Biofuels 2018, 11. [Google Scholar] [CrossRef]
- Wang, X.; Liu, S.F.; Li, R.Y.; Yang, W.D.; Liu, J.S.; Lin, C.S.K.; Balamurugan, S.; Li, H.Y. TAG pathway engineering via GPAT2 concurrently potentiates abiotic stress tolerance and oleaginicity in Phaeodactylum tricornutum. Biotechnol. Biofuels 2020, 13. [Google Scholar] [CrossRef]
- Sun, X.M.; Ren, L.J.; Zhao, Q.Y.; Ji, X.J.; Huang, H. Enhancement of lipid accumulation in microalgae by metabolic engineering. BBA Lipids 2019, 4, 552–566. [Google Scholar] [CrossRef] [PubMed]
- Li-Beisson, Y.; Shorrosh, B.; Beisson, F.; Andersson, M.X.; Arondel, V.; Bates, P.D.; Baud, S.; Bird, D.; Debono, A.; Durrett, T.P.; et al. Acyl-lipid metabolism. Arab. Book 2010, 8, e0133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zulu, N.N.; Zienkiewicz, K.; Vollheyde, K.; Feussner, I. Current trends to comprehend lipid metabolism in diatoms. Prog. Lipid Res. 2018, 70, 1–16. [Google Scholar] [CrossRef]
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog. Lipid Res. 2019, 74, 31–68. [Google Scholar] [CrossRef] [PubMed]
- Chungjatupornchai, W.; Fa-Aroonsawat, S. Enhanced triacylglycerol production in oleaginous microalga Neochloris oleoabundans by co-overexpression of lipogenic genes: Plastidial LPAAT1 and ER-located DGAT2. J. Biosci. Bioeng. 2021, 131, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Trentacoste, E.M.; Shrestha, R.P.; Smith, S.R.; Glé, C.; Hartmann, A.; Hildebrand, M.; Gerwick, W.H. Metabolic engineering of lipid catabolism increases microalgal lipid accumulation without compromising growth. Proc. Natl. Acad. Sci. USA 2013, 110, 19748–19753. [Google Scholar] [CrossRef] [Green Version]
- Barka, F.; Angstenberger, M.; Ahrendt, T.; Lorenzen, W.; Bode, H.B.; Büchel, C. Identification of a triacylglycerol lipase in the diatom Phaeodactylum tricornutum. Biochim. Biophys. Acta 2016, 1861, 239–248. [Google Scholar] [CrossRef]
- Hao, X.; Luo, L.; Jouhet, J.; Rébeillé, F.; Maréchal, E.; Hu, H.; Pan, Y.; Tan, X.; Chen, Z.; You, L.; et al. Enhanced triacylglycerol production in the diatom Phaeodactylum tricornutum by inactivation of a Hotdog-fold thioesterase gene using TALEN-based targeted mutagenesis. Biotechnol. Biofuels 2018, 11, 312. [Google Scholar] [CrossRef]
- Stukenberg, D.; Zauner, S.; Dell’Aquila, G.; Maier, U.G. Optimizing CRISPR/Cas9 for the diatom Phaeodactylum tricornutum. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopes, A.; Nekrasov, V.; Kamoun, S.; Mock, T. Editing of the urease gene by CRISPR-Cas in the diatom Thalassiosira pseudonana. Plant Methods. 2016, 12. [Google Scholar] [CrossRef] [Green Version]
- Moosburner, M.A.; Gholami, P.; McCarthy, J.K.; Tan, M.; Bielinski, V.A.; Allen, A.E. Multiplexed knockouts in the model diatom Phaeodactylum by episomal delivery of a selectable Cas9. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [Green Version]
- Mu, J.; Tan, H.; Zheng, Q.; Fu, F.; Liang, Y.; Zhang, J.; Yang, X.; Wang, T.; Chong, K.; Wang, X.-J.; et al. LEAFY COTYLEDON1 is a key regulator of fatty acid biosynthesis in Arabidopsis. Plant Physiol. 2008, 148, 1042–1054. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zhang, D.; Zhang, J.; Chen, Y.; Liu, X.; Fan, C.; Wang, R.R.; Hou, Y.; Hu, Z. Overexpression of the transcription factor AtLEC1 significantly improved the lipid content of Chlorella ellipsoidea. Front. Bioeng. Biotechnol. 2021, 9. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Song, X.; Li, F.; Lu, Y. Improving lipid productivity by engineering a control-knob gene in the oleaginous microalga Nannochloropsis oceanica. Metab. Eng. Commun. 2020, 11, e00142. [Google Scholar] [CrossRef]
- Jeon, S.; Koh, H.G.; Cho, J.M.; Kang, N.K.; Chang, Y.K. Enhancement of lipid production in Nannochloropsis salina by overexpression of endogenous NADP-dependent malic enzyme. Algal Res. 2021. [Google Scholar] [CrossRef]
- Vikramathithan, J.; Hwangbo, K.; Lim, J.M.; Lim, K.M.; Kang, D.Y.; Park, Y.; Jeong, W.J. Overexpression of Chlamydomonas reinhardtii LCIA (CrLCIA) gene increases growth of Nannochloropsis salina CCMP1776. Algal Res. 2020. [Google Scholar] [CrossRef]
- Südfeld, C.; Hubáček, M.; Figueiredo, D.; Naduthodi, M.I.S.; van der Oost, J.; Wijffels, R.H.; Barbosa, M.J.; D’Adamo, S. High-throughput insertional mutagenesis reveals novel targets for enhancing lipid accumulation in Nannochloropsis oceanica. Metab. Eng. 2021, 66, 239–258. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.; Chen, T.T.; Zheng, J.W.; Balamurugan, S.; Liu, Y.H.; Yang, W.D.; Liu, J.S.; Li, H.Y. Glucose-6-phosphate dehydrogenase from the oleaginous microalga Nannochloropsis uncovers its potential role in promoting lipogenesis. Biotechnol. J. 2020, 15, e1900135. [Google Scholar] [CrossRef]
- Wu, S.; Gu, W.; Huang, A.; Li, Y.; Kumar, M.; Lim, P.E.; Huan, L.; Gao, S.; Wang, G. Elevated CO2 improves both lipid accumulation and growth rate in the glucose-6-phosphate dehydrogenase engineered Phaeodactylum tricornutum. Microb. Cell Fact. 2019, 18, 161. [Google Scholar] [CrossRef]
- Esposito, S. Nitrogen assimilation, abiotic stress and glucose 6-phosphate dehydrogenase: The full circle of ductants. Plants 2016, 11, 24. [Google Scholar] [CrossRef]
- Landi, S.; De Lillo, A.; Nurcato, R.; Grillo, S.; Esposito, S. In-field study on traditional Italian tomato landraces: The constitutive activation of the ROS scavenging machinery reduces effects of drought stress. Plant Physiol. Biochem. 2017, 118, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Borowitzka, M.A. High-value products from microalgae-their development and commercialisation. J. Appl. Phycol. 2013, 25, 743–756. [Google Scholar] [CrossRef]
- Rodolfi, L.; Zittelli, G.C.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection. Induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Cupo, A.; Landi, S.; Morra, S.; Nuzzo, G.; Gallo, C.; Manzo, E.; Fontana, A.; d’Ippolito, G. Autotrophic vs. Heterotrophic cultivation of the marine diatom cyclotella cryptica for EPA production. Mar. Drugs 2021, 19, 355. [Google Scholar] [CrossRef]
- Wang, X.; Luo, S.W.; Luo, W.; Yang, W.D.; Liu, J.S.; Li, H.Y. Adaptive evolution of microalgal strains empowered by fulvic acid for enhanced polyunsaturated fatty acid production. Bioresour. Technol. 2019, 277, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.L.; Powers, S.; Napier, J.A.; Sayanova, O. Heterotrophic production of omega-3 long-chain polyunsaturated fatty acids by trophically converted marine diatom Phaeodactylum tricornutum. Mar. Drugs 2016, 14, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nasopoulou, C.; Zabetakis, I. Benefits of fish oil replacement by plant originated oils in compounded fish feeds. A review. LWT Food Sci. Technol. 2012, 47, 217–224. [Google Scholar] [CrossRef]
- Pudney, A.; Gandini, C.; Economou, C.K.; Smith, R.; Goddard, P.; Napier, J.A.; Spicer, A.; Sayanova, O. Multifunctionalizing the marine diatom Phaeodactylum tricornutum for sustainable co-production of omega-3 long chain polyunsaturated fatty acids and recombinant phytase. Sci. Rep. 2019, 9, 11444. [Google Scholar] [CrossRef]
- Li, S.; Ji, L.; Shi, Q.; Wu, H.; Fan, J. Advances in the production of bioactive substances from marine unicellular microalgae Porphyridium spp. Bioresour. Technol. 2019, 292, 122048. [Google Scholar] [CrossRef]
- Jeon, M.S.; Han, S.I.; Jeon, M.; Choi, Y.E. Enhancement of phycoerythrin productivity in Porphyridium purpureum using the clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9 ribonucleoprotein system. Bioresour. Technol. 2021, 330, 124974. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, X.; Duan, C.; Yi, S.; Gao, Z.; Xiao, C.; Agathos, S.N.; Wang, G.; Li, J. Biotechnological production of astaxanthin from the microalga Haematococcus pluvialis. Biotechnol. Adv. 2021, 43, 107602. [Google Scholar] [CrossRef]
- Waissman-Levy, N.; Leu, S.; Khozin-Goldberg, I.; Boussiba, S. Manipulation of trophic capacities in Haematococcus pluvialis enables low-light mediated growth on glucose and astaxanthin formation in the dark. Algal Res. 2019, 40, 101497. [Google Scholar] [CrossRef]
- Haslam, R.P.; Hamilton, M.L.; Economou, C.K.; Smith, R.; Hassall, K.L.; Napier, J.A.; Sayanova, O. Overexpression of an endogenous type 2 diacylglycerol acyltransferase in the marine diatom Phaeodactylum tricornutum enhances lipid production and omega-3 long-chain polyunsaturated fatty acid content. Biotechnol. Biofuels 2020, 13, 87. [Google Scholar] [CrossRef] [PubMed]
- Seo, S.; Kim, J.; Lee, J.W.; Nam, O.; Chang, K.S.; Jin, E. Enhanced pyruvate metabolism in plastids by overexpression of putative plastidial pyruvate transporter in Phaeodactylum tricornutum. Biotechnol. Biofuels 2020, 13, 120. [Google Scholar] [CrossRef]
- Fathy, W.; Essawy, E.; Tawfik, E.; Khedr, M.; Abdelhameed, M.S.; Hammouda, O.; Elsayed, K. Recombinant overexpression of the Escherichia coli acetyl-CoA carboxylase gene in Synechocystis sp. boosts lipid production. J. Basic Microbiol. 2021. [Google Scholar] [CrossRef]
- Zhu, Z.; Cao, H.; Li, X.; Rong, J.; Cao, X.; Tian, J. A carbon fixation enhanced Chlamydomonas reinhardtii strain for achieving the double-win between growth and biofuel production under non-stressed conditions. Front. Bioeng. Biotechnol. 2021, 8, 603513. [Google Scholar] [CrossRef]
- Kim, J.; Chang, K.S.; Lee, S.; Jin, E. Establishment of a genome editing tool using CRISPR-Cas9 in Chlorella vulgaris UTEX395. Int. J. Mol. Sci. 2021, 22, 480. [Google Scholar] [CrossRef] [PubMed]
- Hammel, A.; Sommer, F.; Zimmer, D.; Stitt, M.; Mühlhaus, T.; Schroda, M. Overexpression of sedoheptulose-1,7-bisphosphatase enhances photosynthesis in Chlamydomonas reinhardtii and has no effect on the abundance of other calvin-benson cycle enzymes. Front. Plant Sci. 2020, 11, 868. [Google Scholar] [CrossRef]
- He, Y.; Zheng, Z.; An, M.; Chen, H.; Qu, C.; Liu, F.; Wang, Y.; Miao, J.; Hou, X. Molecular cloning and functional analysis of a Δ12-fatty acid desaturase from the Antarctic microalga Chlamydomonas sp. ICE-L. 3 Biotech 2019, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Cheng, R.Q.; Liu, Q.Y.; Wang, J.; Fan, Z.C. Multimer of the antimicrobial peptide mytichitin-A expressed in Chlamydomonas reinhardtii exerts a broader antibacterial spectrum and increased potency. J. Biosci. Bioeng. 2018, 125, 175–179. [Google Scholar] [CrossRef]
- Ford, N.R.; Xiong, Y.; Hecht, K.A.; Squier, T.C.; Rorrer, G.L.; Roesijadi, G. Optimizing the design of diatom biosilica-targeted fusion proteins in biosensor construction for Bacillus anthracis detection. Biology 2020, 9, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sproles, A.E.; Fields, F.J.; Smalley, T.N.; Le, C.H.; Badary, A.; Mayfield, S.P. Recent advancements in the genetic engineering of microalgae. Algal Res. 2021. [Google Scholar] [CrossRef]
- Barolo, L.; Abbriano, R.M.; Commault, A.S.; George, J.; Kahlke, T.; Fabris, M.; Padula, M.P.; Lopez, A.; Ralph, P.J.; Pernice, M. Perspectives for glyco-engineering of recombinant biopharmaceuticals from microalgae. Cells 2020, 9, 633. [Google Scholar] [CrossRef] [Green Version]
- Shamriz, S.; Ofoghi, H. Expression of recombinant PfCelTOS antigen in the chloroplast of Chlamydomonas reinhardtii and its potential use in detection of malaria. Mol. Biotechnol. 2019, 61, 102–110. [Google Scholar] [CrossRef]
- Yan, N.; Fan, C.; Chen, Y.; Hu, Z. The potential for microalgae as bioreactors to produce pharmaceuticals. Int. J. Mol. Sci. 2016, 17, 962. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.E.; Scharff, L.B. Engineering of plastids to optimize the production of high-value metabolites and proteins. Curr. Opin. Biotechnol. 2019, 59, 8–15. [Google Scholar] [CrossRef]
- Barrera, D.J.; Rosenberg, J.N.; Chiu, J.G.; Chang, Y.N.; Debatis, M.; Ngoi, S.M.; Chang, J.T.; Shoemaker, C.B.; Oyler, G.A.; Mayfield, S.P. Algal chloroplast produced camelid VH H antitoxins are capable of neutralizing botulinum neurotoxin. Plant Biotechnol. J. 2015, 13, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Hempel, F.; Lau, J.; Klingl, A.; Maier, U.G. Algae as protein factories: Expression of a human antibody and the respective antigen in the diatom Phaeodactylum tricornutum. PLoS ONE 2011, 6, e28424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surzycki, R.; Greenham, K.; Kitayama, K.; Dibal, F.; Wagner, R.; Rochaix, J.D.; Ajam, T.; Surzycki, S. Factors effecting expression of vaccines in microalgae. Biologicals 2009, 37, 133–138. [Google Scholar] [CrossRef]
- Sun, M.; Qian, K.; Su, N.; Chang, H.; Liu, J.; Shen, G. Foot-and-mouth disease virus VP1 protein fused with cholera toxin B subunit expressed in Chlamydomonas reinhardtii chloroplast. Biotechnol. Lett. 2003, 25, 1087–1092. [Google Scholar] [CrossRef] [PubMed]
- Vanier, G.; Stelter, S.; Vanier, J.; Hempel, F.; Maier, U.G.; Lerouge, P.; Ma, J.; Bardor, M. Alga-made anti-hepatitis B antibody binds to human Fcγ receptors. Biotechnol. J. 2018, 13, e1700496. [Google Scholar] [CrossRef]
- Dehghani, J.; Adibkia, K.; Movafeghi, A.; Pourseif, M.M.; Omidi, Y. Designing a new generation of expression toolkits for engineering of green microalgae; robust production of human interleukin-2. Bioimpacts 2020, 10, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Velmurugan, N.; Deka, D. Transformation techniques for metabolic engineering of diatoms and haptophytes: Current state and prospects. Appl. Microbiol. Biotechnol. 2018, 102, 4255–4267. [Google Scholar] [CrossRef]
- He, D.M.; Qian, K.X.; Shen, G.F.; Zhang, Z.F.; Li, Y.N.; Su, Z.L.; Shao, H.B. Recombination and expression of classical swine fever virus (CSFV) structural protein E2 gene in Chlamydomonas reinhardtii chroloplasts. Colloids Surf. B Biointerfaces 2007, 55, 26–30. [Google Scholar] [CrossRef]
- Kiataramgul, A.; Maneenin, S.; Purton, S.; Areechon, N.; Hirono, I.; Brocklehurst, T.W.; Unajak, S. An oral delivery system for controlling white spot syndrome virus infection in shrimp using transgenic microalgae. Aquaculture 2020, 521, 735022. [Google Scholar] [CrossRef]
- Feng, S.; Feng, W.; Zhao, L.; Gu, H.; Li, Q.; Shi, K.; Guo, S.; Zhang, N. Preparation of transgenic Dunaliella salina for immunization against white spot syndrome virus in crayfish. Arch. Virol. 2014, 159, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Charoonnart, P.; Worakajit, N.; Zedler, J.A.Z.; Meetam, M.; Robinson, C.; Saksmerprome, V. Generation of microalga Chlamydomonas reinhardtii expressing shrimp antiviral dsRNA without supplementation of antibiotics. Sci. Rep. 2019, 9, 3164. [Google Scholar] [CrossRef] [Green Version]
- Somchai, P.; Jitrakorn, S.; Thitamadee, S.; Meetam, M.; Saksmerprome, V. Use of microalgae Chlamydomonas reinhardtii for production of double-stranded RNA against shrimp virus. Aquac. Rep. 2016, 3, 178–183. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Peng, H.; Liu, J.; Chen, F.; Zhou, Y.; Ma, X.; Chen, H.; Wang, K. Chlorella sp. transgenic with Scy-hepc enhancing the survival of Sparus macrocephalus and hybrid grouper challenged with Aeromonas hydrophila. Fish Shellfish Immunol. 2018, 73, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Abidin, A.A.Z.; Othman, N.A.; Yusoff, F.M.; Yusoff, Z.N.B. Determination of transgene stability in Nannochloropsis sp. transformed with immunogenic peptide for oral vaccination against vibriosis. Aquac. Int. 2021, 29, 477–486. [Google Scholar] [CrossRef]
- Wang, K.; Cui, Y.; Wang, Y.; Gao, Z.; Liu, T.; Meng, C.; Qin, S. Chloroplast genetic engineering of a unicellular green alga Haematococcus pluvialis with expression of an antimicrobial peptide. Mar. Biotechnol. 2020, 22, 572–580. [Google Scholar] [CrossRef] [PubMed]
- El-Ayouty, Y.; El-Manawy, I.; Nasih, S.; Hamdy, E.; Kebeish, R. Engineering Chlamydomonas reinhardtii for expression of functionally active human interferon-α. Mol. Biotechnol. 2019, 61, 134–144. [Google Scholar] [CrossRef]
- Malla, A.; Rosales-Mendoza, S.; Phoolcharoen, W.; Vimolmangkang, S. Efficient transient expression of recombinant proteins using DNA viral vectors in freshwater microalgal species. Front. Plant. Sci. 2021, 12, 650820. [Google Scholar] [CrossRef]
- Berndt, A.; Smalley, T.; Ren, B.; Badary, A.; Sproles, A.; Fields, F.; Torres-Tiji, Y.; Heredia, V.; Mayfield, S. Recombinant production of a functional SARS- CoV-2 spike receptor binding domain in the green algae Chlamydomonas reinhardtii. bioRxiv 2021. [Google Scholar] [CrossRef]
- Márquez-Escobar, V.A.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Expression of a Zika virus antigen in microalgae: Towards mucosal vaccine development. J. Biotechnol. 2018, 282, 86–91. [Google Scholar] [CrossRef]
- Ortega-Berlanga, B.; Bañuelos-Hernández, B.; Rosales-Mendoza, S. Efficient expression of an Alzheimer’s disease vaccine candidate in the microalga Schizochytrium sp. using the Algevir system. Mol. Biotechnol. 2018, 60, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ramírez, J.; Wong-Arce, A.; González-Ortega, O.; Rosales-Mendoza, S. Expression in algae of a chimeric protein carrying several epitopes from tumor associated antigens. Int. J. Biol. Macromol. 2020, 147, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Hempel, F.; Maurer, M.; Brockmann, B.; Mayer, C.; Biedenkopf, N.; Kelterbaum, A.; Becker, S.; Maier, U.G. From hybridomas to a robust microalgal-based production platform: Molecular design of a diatom secreting monoclonal antibodies directed against the Marburg virus nucleoprotein. Microb. Cell Fact. 2017, 16, 131. [Google Scholar] [CrossRef] [PubMed]
- Pedrioli, G.; Piovesana, E.; Vacchi, E.; Balbi, C. Extracellular vesicles as promising carriers in drug delivery: Considerations from a cell biologist’s perspective. Biology 2021, 10, 376. [Google Scholar] [CrossRef]
- Doyle, L.M.; Wang, M.Z. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [Green Version]
- Villata, S.; Canta, M.; Cauda, V. EVs and bioengineering: From cellular products to engineered nanomachines. Int. J. Mol. Sci. 2020, 21, 6048. [Google Scholar] [CrossRef]
- Picciotto, S.; Barone, M.E.; Fierli, D.; Aranyos, A.; Adamo, G.; Božič, D.; Romancino, D.P.; Stanly, C.; Parkes, R.; Morsbach, S.; et al. Isolation of extracellular vesicles from microalgae: Towards the production of sustainable and natural nanocarriers of bioactive compounds. Biomater. Sci. 2021, 9, 2917–2930. [Google Scholar] [CrossRef]
- Raddadi, N.; Cherif, A.; Daffonchio, D.; Neifar, M.; Fava, F. Biotechnological applications of extremophiles, extremozymes and extremolytes. Appl. Microbiol. Biotechnol. 2015, 99, 7907–7913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malavasi, V.; Soru, S.; Cao, G. Extremophile microalgae: The potential for biotechnological application. J. Phycol. 2020, 56, 559–573. [Google Scholar] [CrossRef]
- Vinogradova, O.M.; Darienko, T.M. Terrestrial algae of hypersaline environments of the Central Syvash islands (Kherson Region, Ukraine). Biologia 2008, 63, 813–823. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, C.; Murakami, M.; Niwa, A.; Takeya, M.; Osanai, T. Efficient extraction and preservation of thermotolerant phycocyanins from red alga Cyanidioschyzon merolae. J. Biosci. Bioeng. 2021, 131, 161–167. [Google Scholar] [CrossRef]
- Puzorjov, A.; McCormick, A.J. Phycobiliproteins from extreme environments and their potential applications. J. Exp. Bot. 2020, 71, 3827–3842. [Google Scholar] [CrossRef]
- Varshney, P.; Mikulic, P.; Vonshak, A.; Beardall, J.; Wangikar, P.P. Extremophilic micro-algae and their potential contribution in biotechnology. Bioresour. Technol. 2015, 184, 363–372. [Google Scholar] [CrossRef]
- Chen, G.Q.; Jiang, X.R. Next generation industrial biotechnology based on extremophilic bacteria. Curr. Opin. Biotechnol. 2018, 50, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Matsakas, L.; Rova, U.; Christakopoulos, P. A perspective on biotechnological applications of thermophilic microalgae and cyanobacteria. Bioresour. Technol. 2019, 278, 424–434. [Google Scholar] [CrossRef]
- Souza, L.D.; Simioni, C.; Bouzon, Z.L.; Schneider, R.C.; Gressler, P.; Miotto, M.C.; Rossi, M.J.; Rörig, L.R. Morphological and ultrastructural characterization of the acidophilic and lipid-producer strain Chlamydomonas acidophila LAFIC-004 (Chlorophyta) under different culture conditions. Protoplasma 2017, 254, 1385–1398. [Google Scholar] [CrossRef]
- Soru, S.; Malavasi, V.; Caboni, P.; Concas, A.; Cao, G. Behavior of the extremophile green alga Coccomyxa melkonianii SCCA 048 in terms of lipids production and morphology at different pH values. Extremophiles 2019, 23, 79–89. [Google Scholar] [CrossRef]
- Abu-Ghosh, S.; Dubinsky, Z.; Banet, G.; Iluz, D. Optimizing photon dose and frequency to enhance lipid productivity of thermophilic algae for biofuel production. Bioresour. Technol. 2018, 260, 374–379. [Google Scholar] [CrossRef]
- de Jaeger, L.; Springer, J.; Wolbert, E.J.H.; Martens, D.E.; Eggink, G.; Wijffels, R.H. Gene silencing of stearoyl-ACP desaturase enhances the stearic acid content in Chlamydomonas reinhardtii. Bioresour. Technol. 2017, 245, 1616–1626. [Google Scholar] [CrossRef]
- Zorin, B.; Pal-Nath, D.; Lukyanov, A.; Smolskaya, S.; Kolusheva, S.; Didi-Cohen, S.; Boussiba, S.; Cohen, Z.; Khozin-Goldberg, I.; Solovchenko, A. Arachidonic acid is important for efficient use of light by the microalga Lobosphaera incisa under chilling stress. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2017, 1862, 853–868. [Google Scholar] [CrossRef]
- Hulatt, C.J.; Berecz, O.; Egeland, E.S.; Wijffels, R.H.; Kiron, V. Polar snow algae as a valuable source of lipids? Bioresour. Technol. 2017, 235, 338–347. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Chaiklahan, R.; Chirasuwan, N.; Bunnag, B. Stability of phycocyanin extracted from Spirulina sp.: Influence of temperature, pH and preservatives. Process Biochem. 2012, 47, 659–664. [Google Scholar] [CrossRef]
- Rahman, D.Y.; Sarian, F.D.; van Wijk, A.; Martinez-Garcia, M.; van der Maarel, M.J.E.C. Thermostable phycocyanin from the red microalga Cyanidioschyzon merolae, a new natural blue food colorant. J. Appl. Phycol. 2017, 29, 1233–1239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remias, D.; Lütz-Meindl, U.; Lütz, C. Photosynthesis, pigments and ultrastructure of the alpine snow alga Chlamydomonas nivalis. Eur. J. Phycol. 2005, 40, 259–268. [Google Scholar] [CrossRef]
- Leya, T.; Rahn, A.; Lütz, C.; Remias, D. Response of arctic snow and permafrost algae to high light and nitrogen stress by changes in pigment composition and applied aspects for biotechnology. FEMS Microbiol. Ecol. 2009, 67, 432–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seckbach, J.; Oren, A.; Stan-lotter, H. Polyextremophiles Life under Multiple Forms of Stress, Cellular Origin, Life in Extreme Habitats and Astrobiology; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Sloth, J.K.; Wiebe, M.G.; Eriksen, N.T. Accumulation of phycocyanin in heterotrophic and mixotrophic cultures of the acidophilic red alga Galdieria sulphuraria. Enzym. Microb. Technol. 2006, 38, 168–175. [Google Scholar] [CrossRef]
- Xie, Y.; Ho, S.H.; Chen, C.N.N.; Chen, C.Y.; Ng, I.S.; Jing, K.J.; Chang, J.S.; Lu, Y. Phototrophic cultivation of a thermo-tolerant Desmodesmus sp. for lutein production: Effects of nitrate concentration, light intensity and fed-batch operation. Bioresour. Technol. 2013, 144, 435–444. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Castiglia, D.; Landi, S.; Esposito, S. Advanced Applications for Protein and Compounds from Microalgae. Plants 2021, 10, 1686. https://doi.org/10.3390/plants10081686
Castiglia D, Landi S, Esposito S. Advanced Applications for Protein and Compounds from Microalgae. Plants. 2021; 10(8):1686. https://doi.org/10.3390/plants10081686
Chicago/Turabian StyleCastiglia, Daniela, Simone Landi, and Sergio Esposito. 2021. "Advanced Applications for Protein and Compounds from Microalgae" Plants 10, no. 8: 1686. https://doi.org/10.3390/plants10081686
APA StyleCastiglia, D., Landi, S., & Esposito, S. (2021). Advanced Applications for Protein and Compounds from Microalgae. Plants, 10(8), 1686. https://doi.org/10.3390/plants10081686