
Topical Gynura procumbens as a Novel Therapeutic Improves Wound Healing in Diabetic Mice

 ,
, 
Abstract

1. Introduction
2. Results
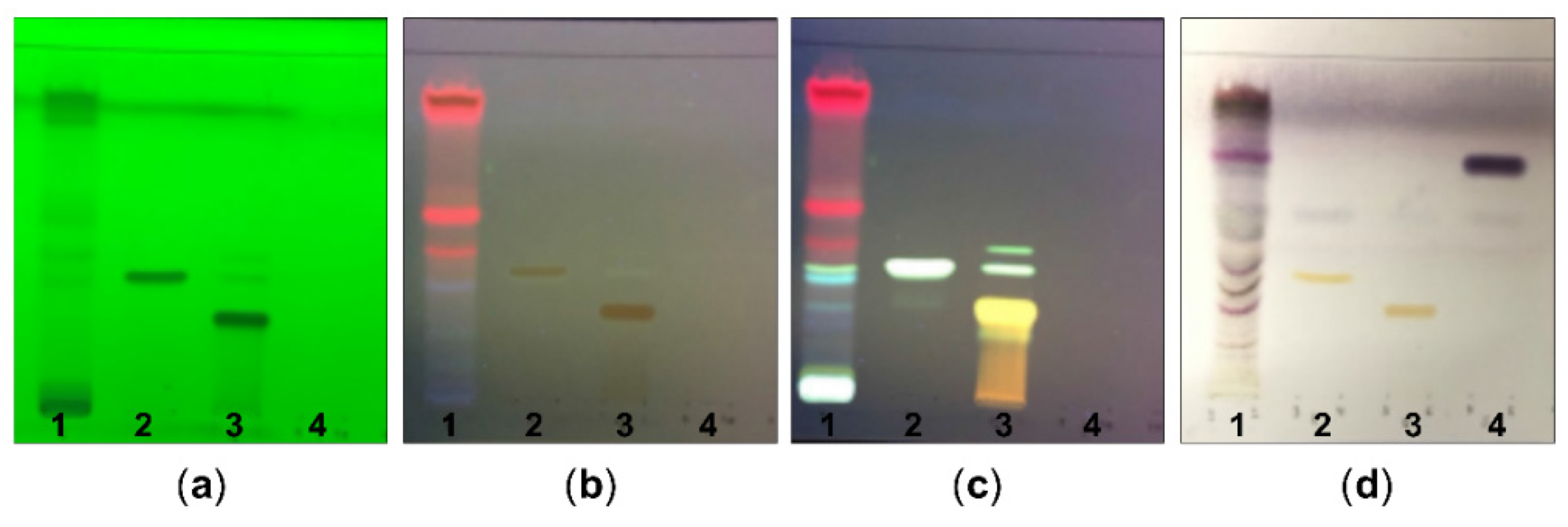
2.1. Chemical Constituents of G. procumbens as Determined by TLC and Phytochemical Screening
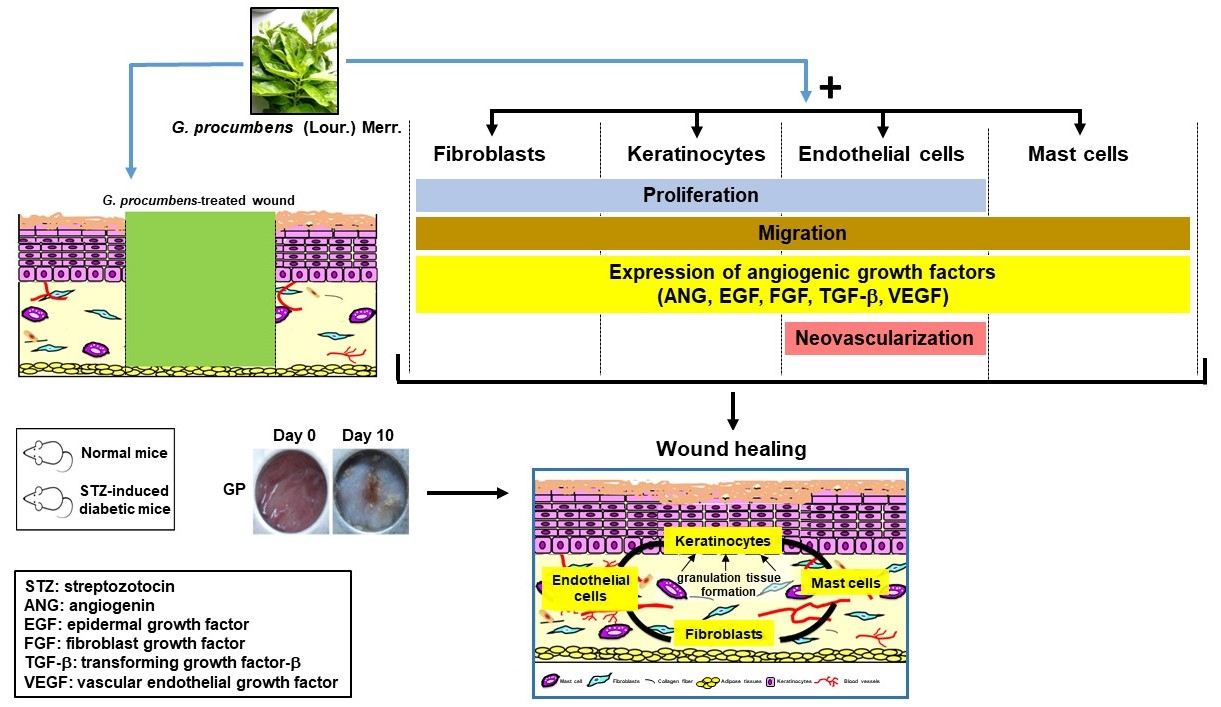
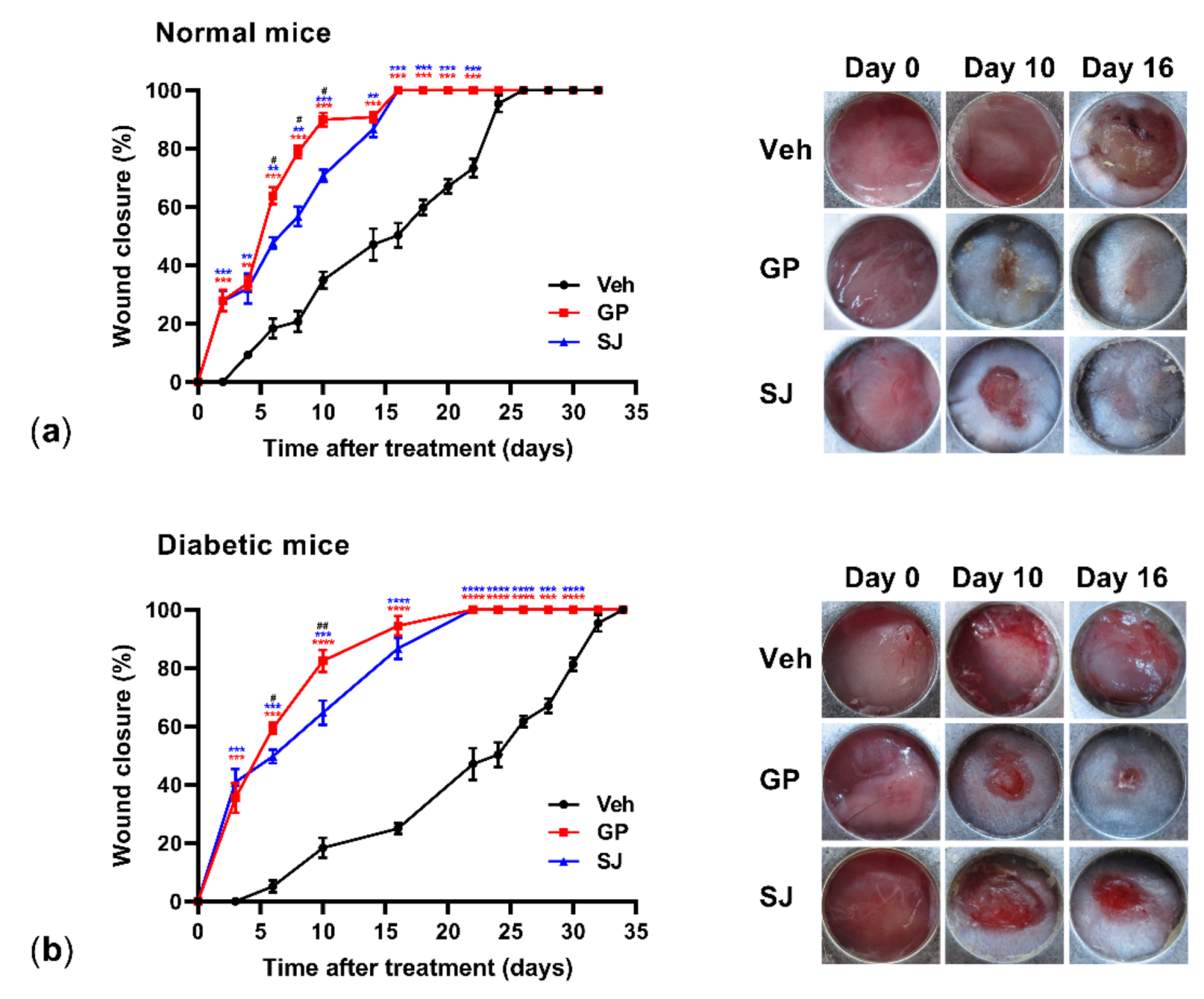
2.2. Ethanolic G. procumbens Extract Promotes Wound Healing in Both Normal and Diabetic Mice
2.3. G. procumbens Induces the Expression of Various Angiogenic Factors
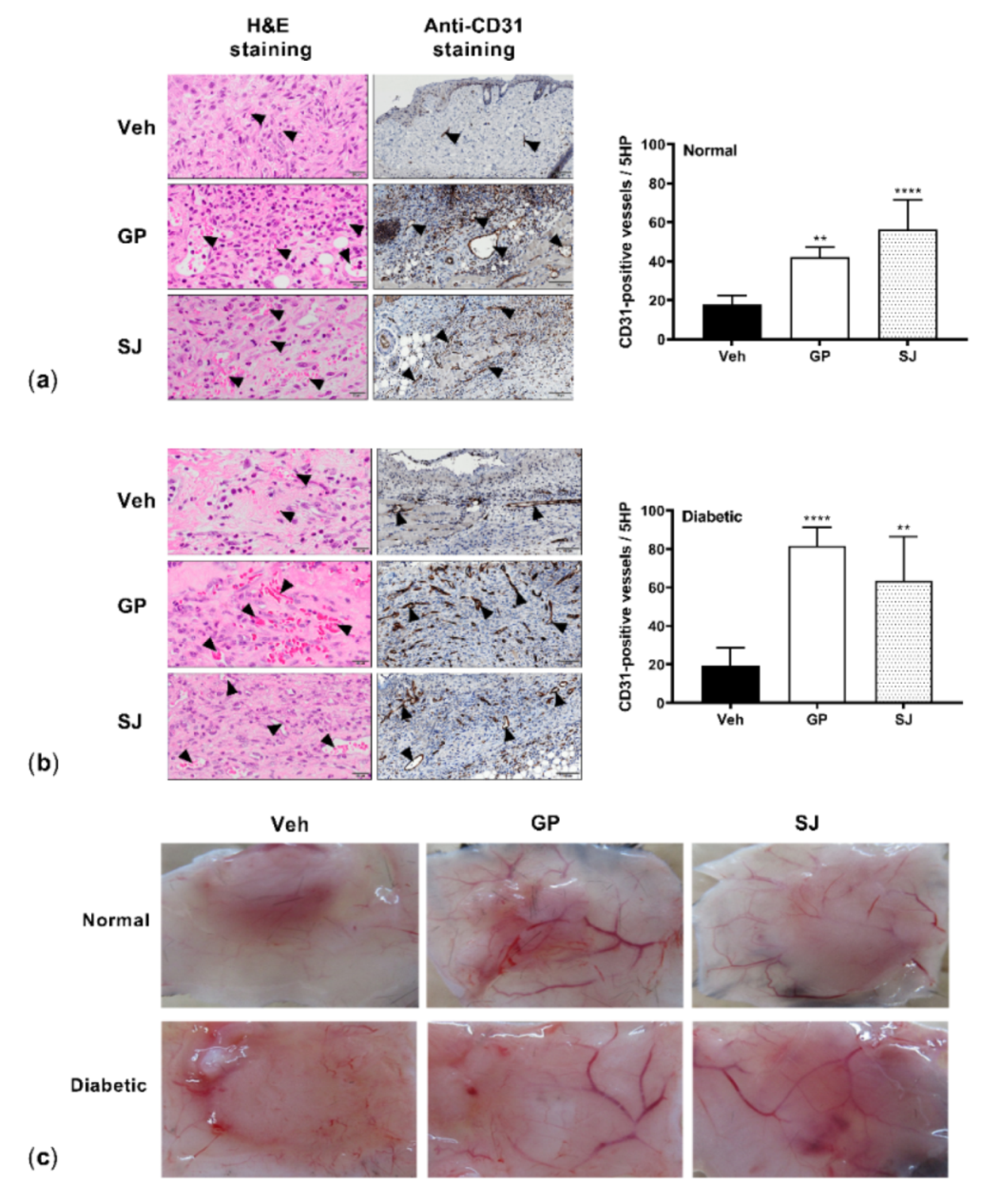
2.4. G. procumbens Promotes Vascular Formation in Normal and Diabetic Mice
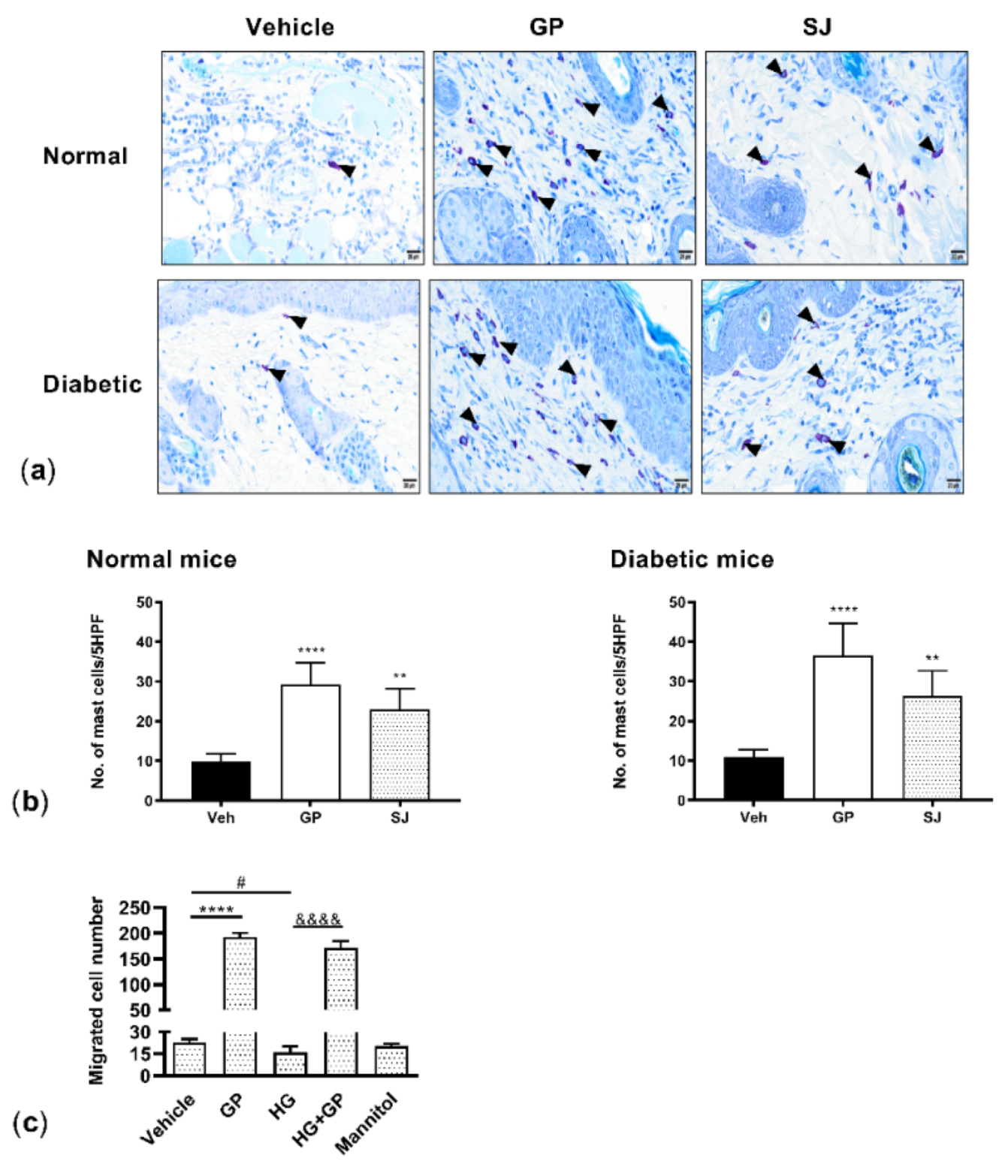
2.5. G. procumbens Enhances Mast Cell Accumulation and Migration
2.6. G. procumbens Promotes the Proliferation of Keratinocytes, Fibroblasts and Endothelial Cells
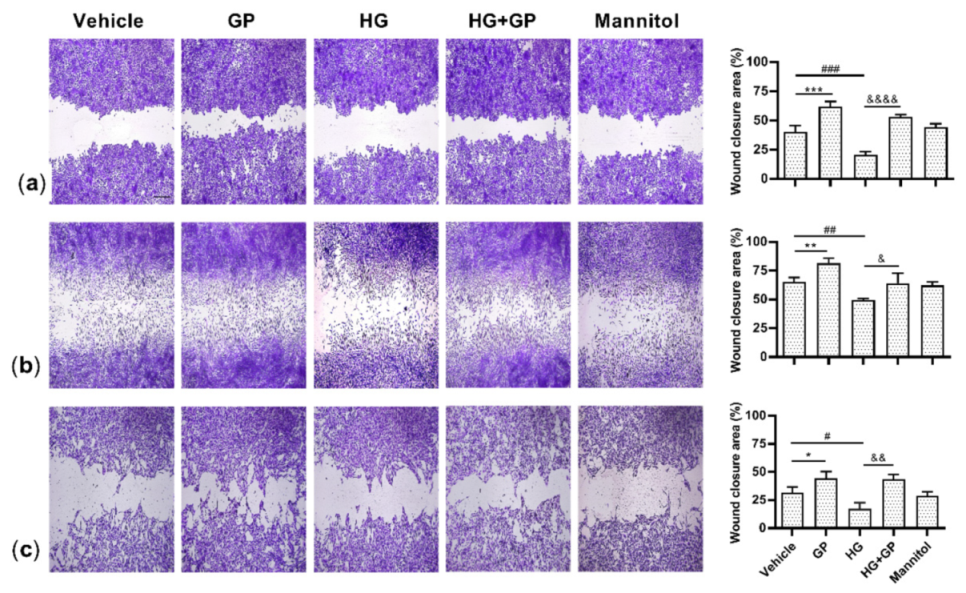
2.7. G. procumbens Accelerates Wound Healing In Vitro
3. Discussion
4. Materials and Methods
4.1. Extraction of G. procumbens
4.2. Phytochemical Screening of the G. procumbens Extract
4.3. Animals
4.4. In Vivo Wound Model and Treatment
4.5. Cell Culture
4.6. Cell Migration
4.7. Cell Proliferation
4.8. Total RNA Extraction and Real-Time Quantitative PCR
4.9. Histologic and Immunohistochemical Analyses and Visualization of Vascular Formation at the Wound Site
4.10. Statistical Analysis
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANG | Angiogenin |
| EGF | Epidermal growth factor |
| FGF | Fibroblast growth factor |
| G. procumbens | Gynura procumbens |
| PBS | Phosphate-buffered saline |
| PDGF | Platelet-derived growth factor |
| TGF | Transforming growth factor |
| VEGF | Vascular endothelial growth factor |
References
- Nussbaum, S.R.; Carter, M.J.; Fife, C.E.; DaVanzo, J.; Haught, R.; Nusgart, M.; Cartwright, D. An economic evaluation of the impact, cost, and medicare policy implications of chronic nonhealing wounds. Value Health 2018, 21, 27–32. [Google Scholar] [CrossRef]
- Brem, H.; Tomic-Canic, M. Cellular and molecular basis of wound healing in diabetes. J. Clin. Investig. 2007, 117, 1219–1222. [Google Scholar] [CrossRef]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [CrossRef]
- Kavitha, K.V.; Tiwari, S.; Purandare, V.B.; Khedkar, S.; Bhosale, S.S.; Unnikrishnan, A.G. Choice of wound care in diabetic foot ulcer: A practical approach. World J. Diabetes 2014, 5, 546–556. [Google Scholar] [CrossRef]
- Tan, H.L.; Chan, K.G.; Pusparajah, P.; Lee, L.H.; Goh, B.H. Gynura procumbens: An overview of the biological activities. Front. Pharmacol. 2016, 7, 52. [Google Scholar] [CrossRef]
- Ng, H.K.; Poh, T.F.; Lam, S.K.; Hoe, S.Z. Potassium channel openers and prostacyclin play a crucial role in mediating the vasorelaxant activity of Gynura procumbens. BMC Complement. Altern. Med. 2013, 13, 188. [Google Scholar] [CrossRef]
- Zahra, A.A.; Kadir, F.A.; Mahmood, A.; Suzy, S.; Sabri, S.; Latif, I.; Ketuly, K. Acute toxicity study and wound healing potential of Gynura procumbens leaf extract in rats. J. Med. Plants Res. 2011, 5, 2551–2558. [Google Scholar]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Monaco, J.L.; Lawrence, W.T. Acute wound healing an overview. Clin. Plast. Surg. 2003, 30, 1–12. [Google Scholar] [CrossRef]
- Barrientos, S.; Brem, H.; Stojadinovic, O.; Tomic-Canic, M. Clinical application of growth factors and cytokines in wound healing. Wound Repair Regen. 2014, 22, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Algariri, K.; Meng, K.Y.; Atangwho, I.J.; Asmawi, M.Z.; Sadikun, A.; Murugaiyah, V.; Ismail, N. Hypoglycemic and anti-hyperglycemic study of Gynura procumbens leaf extracts. Asian Pac. J. Trop. Biomed. 2013, 3, 358–366. [Google Scholar] [CrossRef]
- Wilmink, J.M.; Stolk, P.W.; van Weeren, P.R.; Barneveld, A. The effectiveness of the haemodialysate Solcoseryl for second-intention wound healing in horses and ponies. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2000, 47, 311–320. [Google Scholar] [CrossRef]
- El-Mesallamy, H.O.; Diab, M.R.; Hamdy, N.M.; Dardir, S.M. Cell-based regenerative strategies for treatment of diabetic skin wounds, a comparative study between human umbilical cord blood-mononuclear cells and calves’ blood haemodialysate. PLoS ONE 2014, 9, e89853. [Google Scholar] [CrossRef]
- Okano, J.; Kojima, H.; Katagi, M.; Nakagawa, T.; Nakae, Y.; Terashima, T.; Kurakane, T.; Kubota, M.; Maegawa, H.; Udagawa, J. Hyperglycemia induces skin barrier dysfunctions with impairment of epidermal integrity in non-wounded skin of type 1 diabetic mice. PLoS ONE 2016, 11, e0166215. [Google Scholar] [CrossRef]
- El-Remessy, A.B.; Abou-Mohamed, G.; Caldwell, R.W.; Caldwell, R.B. High glucose-induced tyrosine nitration in endothelial cells: Role of eNOS uncoupling and aldose reductase activation. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3135–3143. [Google Scholar] [CrossRef]
- Galiano, R.D.; Tepper, O.M.; Pelo, C.R.; Bhatt, K.A.; Callaghan, M.; Bastidas, N.; Bunting, S.; Steinmetz, H.G.; Gurtner, G.C. Topical vascular endothelial growth factor accelerates diabetic wound healing through increased angiogenesis and by mobilizing and recruiting bone marrow-derived cells. Am. J. Pathol. 2004, 164, 1935–1947. [Google Scholar] [CrossRef]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Singh, N.; Armstrong, D.G.; Lipsky, B.A. Preventing foot ulcers in patients with diabetes. JAMA 2005, 293, 217–228. [Google Scholar] [CrossRef]
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic foot ulcers and their recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef]
- Rosidah; Yam, M.F.; Sadikun, A.; Ahmad, M.; Akowuah, G.A.; Asmawi, M.Z. Toxicology evaluation of standardized methanol extract of Gynura procumbens. J. Ethnopharmacol. 2009, 123, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Kaewseejan, N.; Sutthikhum, V.; Siriamornpun, S. Potential of Gynura procumbens leaves as source of flavonoid-enriched fractions with enhanced antioxidant capacity. J. Funct. Foods 2015, 12, 120–128. [Google Scholar] [CrossRef]
- Kim, J.; Lee, C.W.; Kim, E.K.; Lee, S.J.; Park, N.H.; Kim, H.S.; Kim, H.K.; Char, K.; Jang, Y.P.; Kim, J.W. Inhibition effect of Gynura procumbens extract on UV-B-induced matrix-metalloproteinase expression in human dermal fibroblasts. J. Ethnopharmacol. 2011, 137, 427–433. [Google Scholar] [CrossRef]
- Tatsimo, S.J.; Tamokou, J.D.; Havyarimana, L.; Csupor, D.; Forgo, P.; Hohmann, J.; Kuiate, J.R.; Tane, P. Antimicrobial and antioxidant activity of kaempferol rhamnoside derivatives from Bryophyllum pinnatum. BMC Res. Notes 2012, 5, 158–164. [Google Scholar] [CrossRef]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and inflammation: From chemistry to medicine. Pharmacol. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Özay, Y.; Güzel, S.; Yumrutaş, Ö.; Pehlivanoğlu, B.; Erdoğdu, İ.H.; Yildirim, Z.; Türk, B.A.; Darcan, S. Wound healing effect of kaempferol in diabetic and nondiabetic rats. J. Surg. Res. 2019, 233, 284–296. [Google Scholar] [CrossRef] [PubMed]
- Petpiroon, N.; Suktap, C.; Pongsamart, S.; Chanvorachote, P.; Sukrong, S. Kaempferol-3-O-rutinoside from Afgekia mahidoliae promotes keratinocyte migration through FAK and Rac1 activation. J. Nat. Med. 2015, 69, 340–348. [Google Scholar] [CrossRef]
- Chirumbolo, S. Flavonoids in propolis acting on mast cell-mediated wound healing. Inflammopharmacology 2012, 20, 99–101. [Google Scholar] [CrossRef]
- Ahmad, M.; Sultana, M.; Raina, R.; Pankaj, N.K.; Verma, P.K.; Prawez, S. Hypoglycemic, Hypolipidemic, and Wound Healing Potential of Quercetin in Streptozotocin-Induced Diabetic Rats. Pharmacogn. Mag. 2017, 13, S633–S639. [Google Scholar] [CrossRef]
- Hujiahemaiti, M.; Sun, X.; Zhou, J.; Lv, H.; Li, X.; Qi, M.; Chi, M.; Li, C.; Zhou, Y. Effects of quercetin on human oral keratinocytes during re-epithelialization: An in vitro study. Arch. Oral Biol. 2018, 95, 187–194. [Google Scholar] [CrossRef]
- Kaur, N.; Chaudhary, J.; Jain, A.; Kishore, L. Stigmasterol: A comprehensive review. Int. J. Pharm. Sci. Res. 2011, 2, 2259–2265. Available online: https://ijpsr.com/bft-article/stigmasterol-a-comprehensive-review/?view=fulltext (accessed on 7 March 2020).
- Viswanathan, M.B.; Ananthi, J.D.J.; Raja, N.L.; Venkateshan, N. Wound healing activity of Jatropha tanjorensis leaves. J. Pharm. Res. 2018, 3, 24–30. [Google Scholar] [CrossRef]
- Chaudhari, M.; Mengi, S. Evaluation of phytoconstituents of Terminalia arjuna for wound healing activity in rats. Phytother. Res. 2006, 20, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Barreto, R.S.; Albuquerque-Júnior, R.L.; Araújo, A.A.; Almeida, J.R.; Santos, M.R.; Barreto, A.S.; DeSantana, J.M.; Siqueira-Lima, P.S.; Quintans, J.S.; Quintans-Júnior, L.J. A systematic review of the wound-healing effects of monoterpenes and iridoid derivatives. Molecules 2014, 19, 846–862. [Google Scholar] [CrossRef]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Encinar, J.A.; Rodríguez-Díaz, J.C.; Micol, V. Antimicrobial capacity of plant polyphenols against gram-positive bacteria: A comprehensive review. Curr. Med. Chem. 2020, 27, 2576–2606. [Google Scholar] [CrossRef]
- Ashraf, K.; Halim, H.; Lim, S.M.; Ramasamy, K.; Sultan, S. In vitro antioxidant, antimicrobial and antiproliferative studies of four different extracts of Orthosiphon stamineus, Gynura procumbens and Ficus deltoidea. Saudi J. Biol. Sci. 2020, 27, 417–432. [Google Scholar] [CrossRef]
- Afandi, A.; Sadikun, A.; Ismail, S. Antioxidant properties of Gynura procumbens extracts and their inhibitory effects on two major human recombinant cytochrome P450s using a high throughout luminescence assay. Asian J. Pharm. Clin. Res. 2014, 7, 36–41. Available online: https://innovareacademics.in/journals/index.php/ajpcr/article/viewFile/1162/1476 (accessed on 10 May 2021).
- Abrika, O.S.; Yam, M.F.; Asmawi, M.Z.; Sadikun, A.; Dieng, H.; Hussain, E.A. Effects of extracts and fractions of Gynura procumbens on rat atrial contraction. J. Acupunct. Meridian Stud. 2013, 6, 199–207. [Google Scholar] [CrossRef]
- Okonkwo, U.A.; DiPietro, L.A. Diabetes and wound angiogenesis. Int. J. Mol. Sci. 2017, 18, 1419. [Google Scholar] [CrossRef]
- Nishikori, Y.; Shiota, N.; Okunishi, H. The role of mast cells in cutaneous wound healing in streptozotocin-induced diabetic mice. Arch. Dermatol. Res. 2014, 306, 823–835. [Google Scholar] [CrossRef]
- Borena, B.M.; Martens, A.; Broeckx, S.Y.; Meyer, E.; Chiers, K.; Duchateau, L.; Spaas, J.H. Regenerative skin wound healing in mammals: State-of-the-art on growth factor and stem cell-based treatments. Cell. Physiol. Biochem. 2015, 36, 1–23. [Google Scholar] [CrossRef]
- Steed, D.L. Clinical evaluation of recombinant human platelet-derived growth factor for the treatment of lower extremity ulcers. Plast. Reconstr. Surg. 2006, 117, 143S–149S. [Google Scholar] [CrossRef] [PubMed]
- Mäkinen, K.; Manninen, H.; Hedman, M.; Matsi, P.; Mussalo, H.; Alhava, E.; Ylä-Herttuala, S. Increased vascularity detected by digital subtraction angiography after VEGF gene transfer to human lower limb artery: A randomized, placebo-controlled, double-blinded phase II study. Mol. Ther. 2002, 6, 127–133. [Google Scholar] [CrossRef]
- Sheng, J.; Xu, Z. Three decades of research on angiogenin: A review and perspective. Acta Biochim. Biophys. Sin. 2016, 48, 399–410. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, M.; Chen, S.; Wang, X.; Tian, Z.; Chen, Y.; Xu, P.; Zhang, L.; Zhang, L.; Zhang, L. Peptide-modified chitosan hydrogels accelerate skin wound healing by promoting fibroblast proliferation, migration, and secretion. Cell Transpl. 2017, 26, 1331–1340. [Google Scholar] [CrossRef]
- Moenner, M.; Gusse, M.; Hatzi, E.; Badet, J. The widespread expression of angiogenin in different human cells suggests a biological function not only related to angiogenesis. Eur. J. Biochem. 1994, 226, 483–490. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial RNases of human skin. J. Investig. Dermatol. 2009, 129, 2091–2093. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Haigh, B.J.; Griffin, F.J.; Wheeler, T.T. The mammalian secreted RNases: Mechanisms of action in host defence. Innate Immun. 2013, 19, 86–97. [Google Scholar] [CrossRef]
- Behm, B.; Babilas, P.; Landthaler, M.; Schreml, S. Cytokines, chemokines and growth factors in wound healing. J. Eur. Acad. Dermatol. Venereol. 2012, 26, 812–820. [Google Scholar] [CrossRef]
- Serra, M.B.; Barroso, W.A.; da Silva, N.N.; Silva, S.D.N.; Borges, A.C.R.; Abreu, I.C.; Borges, M. From inflammation to current and alternative therapies involved in wound healing. Int. J. Inflamm. 2017, 2017, 1–17. [Google Scholar] [CrossRef]
- Noli, C.; Miolo, A. The mast cell in wound healing. Vet. Dermatol. 2001, 12, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Antsiferova, M.; Martin, C.; Huber, M.; Feyerabend, T.B.; Förster, A.; Hartmann, K.; Rodewald, H.R.; Hohl, D.; Werner, S. Mast cells are dispensable for normal and activin-promoted wound healing and skin carcinogenesis. J. Immunol. 2013, 191, 6147–6155. [Google Scholar] [CrossRef]
- Willenborg, S.; Eckes, B.; Brinckmann, J.; Krieg, T.; Waisman, A.; Hartmann, K.; Roers, A.; Eming, S.A. Genetic ablation of mast cells redefines the role of mast cells in skin wound healing and bleomycin-induced fibrosis. J. Investig. Dermatol. 2014, 134, 2005–2015. [Google Scholar] [CrossRef] [PubMed]
- Jaradat, N.; Hussen, F.; Al Ali, A. Preliminary phytochemical screening, quantitative estimation of total flavonoids, total phenols and antioxidant activity of Ephedra alata Decne. J. Mater. Environ. Sci. 2015, 6, 1771–1778. [Google Scholar] [CrossRef]
- Zhang, X.F.; Tan, B.K. Effects of an ethanolic extract of Gynura procumbens on serum glucose, cholesterol and triglyceride levels in normal and streptozotocin-induced diabetic rats. Singap. Med. J. 2000, 41, 9–13. Available online: https://www.sma.org.sg/SMJ/4101/Articles/4101a2.htm (accessed on 16 April 2020).
- Cragg, G.; Newman, D. Drugs from nature: Past achievements, future prospects. Adv. Phyto Med. 2002, 1, 23–37. [Google Scholar] [CrossRef]
- Chieosilapatham, P.; Yue, H.; Ikeda, S.; Ogawa, H.; Niyonsaba, F. Involvement of the lipoprotein receptor LRP1 in AMP-IBP5-mediated migration and proliferation of human keratinocytes and fibroblasts. J. Dermatol. Sci. 2020, 99, 158–167. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Song, P.; Yue, H.; Sutthammikorn, N.; Umehara, Y.; Okumura, K.; Ogawa, H. Antimicrobial peptide derived from insulin-like growth factor-binding protein 5 activates mast cells via Mas-related G protein-coupled receptor X2. Allergy 2020, 75, 203–207. [Google Scholar] [CrossRef]
- Yanashima, K.; Chieosilapatham, P.; Yoshimoto, E.; Okumura, K.; Ogawa, H.; Niyonsaba, F. Innate defense regulator IDR-1018 activates human mast cells through G protein-, phospholipase C-, MAPK- and NF-ĸB-sensitive pathways. Immunol. Res. 2017, 65, 920–931. [Google Scholar] [CrossRef]
- Chieosilapatham, P.; Niyonsaba, F.; Kiatsurayanon, C.; Okumura, K.; Ikeda, S.; Ogawa, H. The antimicrobial peptide derived from insulin-like growth factor-binding protein 5, AMP-IBP5, regulates keratinocyte functions through Mas-related gene X receptors. J. Dermatol. Sci. 2017, 88, 117–125. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test | Result |
|---|---|
| 1. Alkaloid | |
| 1.1. Dragendorff’s reagent | − |
| 1.2. Wagner’s reagent | − |
| 1.3. Marme’s reagent | − |
| 1.4. Mayer’s reagent | − |
| 2. Tannins (General test) | |
| 2.1. Ferric chloride TS. | + |
| 2.2. 0.5% Gelatin solution | + |
| 2.3. 1% Lead acetate solution | − |
| 2.4. 1% Quinine sulfate | − |
| 3. Tannins (hydrolysable tannin) | |
| 3.1. Ferric chloride TS. | − |
| 3.2. Lime water | − |
| 4. Tannins (condensed tannin) | |
| 4.1. Vanillin reagent | − |
| 4.2. Formalin-HCl reagent | − |
| 5. Glycoside | |
| 5.1. Antraquinone | − |
| 5.2. Flavonoid | + |
| 5.3. Saponin | − |
| 5.4. Cyanogenic glycoside | − |
| 5.5. Cardiac glycoside (steroid) | − |
| 5.6. Cardiac glycoside (unsaturated lactone) | − |
| 5.7. Cardiac glycoside (deoxy sugar) | − |
| 6. Terpenoids | + |
| 7. Protein | + |
| 7.1. Xanthoproteic | + |
| 7.2. Ninhydrin | + |
| 7.3. Lead sulfide | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutthammikorn, N.; Supajatura, V.; Yue, H.; Takahashi, M.; Chansakaow, S.; Nakano, N.; Song, P.; Ogawa, T.; Ikeda, S.; Okumura, K.; et al. Topical Gynura procumbens as a Novel Therapeutic Improves Wound Healing in Diabetic Mice. Plants 2021, 10, 1122. https://doi.org/10.3390/plants10061122
Sutthammikorn N, Supajatura V, Yue H, Takahashi M, Chansakaow S, Nakano N, Song P, Ogawa T, Ikeda S, Okumura K, et al. Topical Gynura procumbens as a Novel Therapeutic Improves Wound Healing in Diabetic Mice. Plants. 2021; 10(6):1122. https://doi.org/10.3390/plants10061122
Chicago/Turabian StyleSutthammikorn, Nutda, Volaluck Supajatura, Hainan Yue, Miho Takahashi, Sunee Chansakaow, Nobuhiro Nakano, Pu Song, Takasuke Ogawa, Shigaku Ikeda, Ko Okumura, and et al. 2021. "Topical Gynura procumbens as a Novel Therapeutic Improves Wound Healing in Diabetic Mice" Plants 10, no. 6: 1122. https://doi.org/10.3390/plants10061122
APA StyleSutthammikorn, N., Supajatura, V., Yue, H., Takahashi, M., Chansakaow, S., Nakano, N., Song, P., Ogawa, T., Ikeda, S., Okumura, K., Ogawa, H., & Niyonsaba, F. (2021). Topical Gynura procumbens as a Novel Therapeutic Improves Wound Healing in Diabetic Mice. Plants, 10(6), 1122. https://doi.org/10.3390/plants10061122