1. Introduction
Alzheimer’s disease (AD) is a common neurodegenerative disease in aged population and has become the third leading cause of death after cardiovascular disease and cancer [
1]. According to the world Alzheimer’s report of 2018, there are about 50 million AD patients in the world, and it will increase to 150 million by 2050. Currently, there are no effective drugs that can prevent AD pathogenesis or slow down its progression. Among the different therapeutic strategies of AD, an increase in the acetylcholine level in the brain using acetylcholinesterase inhibitors (AChEI) is considered to be an effective treatment to alleviate some of the symptoms of the disease [
2]. A large number of clinical trials suggest that Huperzine A (HupA) demonstrates the favorable safety profiles at a relatively wide dose range and the potent efficacy in AD patients [
3]. It has been confirmed that HupA is a highly effective, selective, and naturally reversible AChEI, with long-term action, ease in crossing the blood–brain barrier, and small side effects [
4,
5]. In China, HupA has been approved for the treatment of AD and vascular dementia, and phase III clinical trials have been completed in Europe [
6]. So, it is a promising candidate for clinical development as a symptomatic treatment for AD [
7,
8].
Huperzia serrata (
H. serrata) is a traditional Chinese herbal medicine used for the treatment of contusion, strain, swelling, and schizophrenia. Many
Lycopodium alkaloids have been isolated from this plant. Since the highly effective AChEI Hup A was discovered in
H. serrata [
9], there has been more and more attention and research on
H. serrata [
10]. At present,
H. serrata, the main source of HupA, actually possesses a very low content of HupA about 0.007% [
11], grows very slowly, and finds it difficult to reproduce under natural conditions. It takes 6–15 years from spore germination to grow into a complete plant of about 12 cm high. For this reason, many researchers have tried to obtain HupA through chemical synthesis and endophytic fermentation [
12,
13,
14].
In fact, the racemic synthesis of HupA has been achieved. However, regarding the in vitro inhibitory effects on the AChE enzyme, the racemic mixture of HupA is three times less potent than HupA from
H. serrata. If used in large doses, contractions or other symptoms of cholinergic hyperactivity may occur [
11], and this synthesis method requires more expensive reagents, such as phenyl selenium chloride, LDA, etc., which makes it difficult to meet the requirements of large-scale preparation [
11]. Meanwhile, the fermentation yield of endophytic fungi is low and unstable. After several subcultures, the endophytic fungi could not continue to produce HupA [
15,
16]. Therefore, the production of HupA is still completely dependent on the wild
H. serrata or its related species [
10]. This has led to a large number of wild resources being widely harvested, and
H. serrata was even at the risk of being endangered. So, the establishment of a suitable artificial propagation system is an important way to protect and utilize wild
H. serrata.
Plant tissue culture is regarded as a powerful tool for micropropagation and conservation of unique plant germplasms. Studies on the micropropagation of
H. serrata started in 1957, but the progress has been very slow because of the difficult sterilization and regeneration [
17,
18]. Recently, a few research groups have obtained in vitro thallus regenerated from
H. serrata [
19]. Previously, our group had also successfully obtained one genotypic thallus of
H. serrata, which can produce HupA in vitro [
20]. Then we further studied the correlation between HupA accumulation and the morphology of in vitro culture [
21]. However, the contamination rate of
H. serrata in previous studies is still relatively high and the experiment material of
H. serrata is limited to one genotype. Meanwhile, previous studies on bioactive components and antioxidant activities of in vitro
H. serrata are still very limited, which restricted the investigation and potential application of in vitro thallus.
Therefore, the regeneration and micropropagation system of different genotypic H. serrata was investigated in this study. In addition, the potential ability of HupA production and the antioxidant activity for different in vitro H. serrata were comparatively analyzed. These detailed results would be able to relieve the shortage of Chinese medicinal resources for HupA production and lay a very important foundation for germplasm improvement and genetic engineering studies for H. serrata.
2. Results
2.1. The Effect of Sterilization Methods on H. serrata Survival
In order to improve the sterilization effect of
H. serrata explants, different sterilization methods or combinations were used in this study (
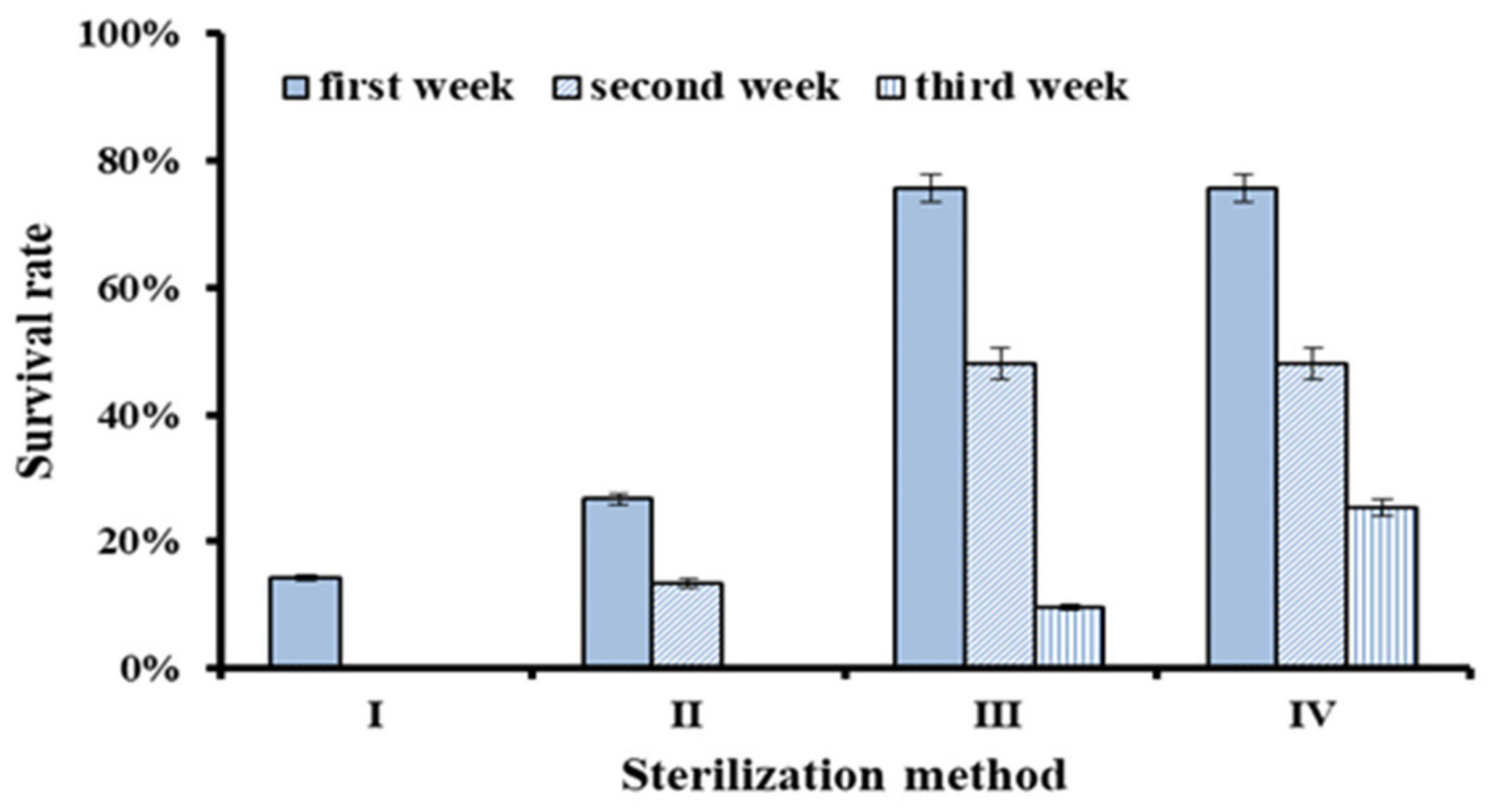
Table 1). The results showed that the survival rate of explants from different sterilization methods was significantly different (
Figure 1). The survival rates in the first week were 14.29% for Method I, 26.67% for Method II, and 75.59% for Method III, respectively. Two weeks later, the survival rates were 0% (Method I), 13.33% (Method II), and 48.03% (Method III), respectively. Therefore, Method III was much better than Method II and I.
Two weeks after inoculation, about 50% of non-polluted explants of Method III were transferred to the regeneration medium containing antibiotics (Method IV). One week later, the survival rate of Method IV (25.37%) was higher than that of Method III (9.52%), which suggested that a certain concentration of antibiotics could inhibit endophytic fungi and would improve the survival rate of explants. Therefore, the best sterilization combination was 75% ethanol for 30 s, 10% hydrogen peroxide for 8 min, and 0.15% mercuric chloride for 5 min. After 2 weeks of regeneration culture, the non-polluted explants were transferred to a new regeneration medium containing 0.5 mg·L−1 malachite green and 100 mg·L−1 AAS (Antibiotic Antimycotic Solution, 100 mg AAS contains 50,000 IU of the penicillin, 50 mg streptomycin and 125 µg amphotericin B).
2.2. The Effects of Hormone Type and Concentration on H. serrata Regeneration
In order to assess the effect of hormone types and concentrations on the
H. serrata regeneration, an orthogonal experiment was designed in the
Table 2. Statistical analysis showed that the range corresponding to factor A (NAA) was the largest (23), followed by factor C (2,4-D) (17.98), and factor B (ZT) (13.70) was the smallest. These data indicated that NAA concentration (factor A) had a significant difference on the thallus regeneration of
H. serrata, followed by 2,4-D (factor C).
Further analysis showed that there are significant differences among the sum of test index (induction rate) corresponding to different levels of each factor. For NAA (factor A), the sum of induction rate reaches a maximum at 0.5 mg/L (Level 2 of factor A), that is KA
2 > KA
3 > KA
1 (
Table 1). For factor B (ZT) and C (2,4-D), the maximum value of the sum of induction rate was at 0 mg·L
−1 (Level 1 of factor B) and 0.1 mg·L
−1 (Level 2 of factor C), respectively. So, the most suitable hormone combination for
H. serrata regeneration was A
2B
1C
2. That is to say, the best induction medium for
H. serrata regeneration was SH medium supplemented with 0.5 mg·L
−1 NAA and 0.1 mg·L
−1 2,4-D [
22,
23], whereby the regeneration rate of the
H. serrata explant was increased to 57.04% (
Table 2).
2.3. The Effects of Hormone Type and Concentration on the H. serrata Proliferation
In order to investigate the effect of hormone types and concentrations on in vitro
H. serrata proliferation, a multi-factor randomized comparative test was designed. As shown in
Table 3, the effects of different hormone concentrations and types were significantly different (
p < 0.05). The relative growth rate of the treatment group (supplemented with hormones group) was significantly higher than that of the control group (No hormones group). When the SH medium was supplemented with 1.0 mg·L
−1 NAA, the relative growth rate achieved the maximum value (164.17 ± 0.41 times), which was about 2 times that of the control group. When the NAA concentration was at 1.5 mg·L
−1, the relative growth rate of in vitro
H. serrata reduced to 114.88 ± 2.13 times, which indicated that the excessive hormone would inhibit the in vitro
H. serrata proliferation. A similar trend was observed among the IAA treatment. The corresponding relative growth rate was less than that of the NAA treatment group with the same concentration. So, the best multiplication medium was SH medium with NAA 1.0 mg·L
−1 for
H. serrata proliferation. The process of in vitro propagation of
H. serrata is shown in
Figure 2.
2.4. Morphological Difference of Different H. serrata Thallus
In this study, some morphology and growth differences of the three genotypes in vitro
H. serrata thallus were observed. As shown in
Table 4, the thallus of the HB genotype grew more rapidly. A 164.17 ± 0.41-fold increase in biomass has increased, followed by the FJ genotype (153.71 ± 0.53 times) and the JX genotype (121.08 ± 1.09 times). For the morphological difference of thallus, the thallus of HB was thin and long, FJ’s were wide and large, and JX’s were thin and short (
Table 4,
Figure 3). As for the size of the thallus, the diameter of HB and JX were 1.74 ± 0.73 cm and 1.18 ± 0.15 cm, respectively, while that of FJ was 2.59 ± 0.21 cm. As shown in
Figure 3, the morphology of the thallus for HB and JX genotype was compact, while that of FJ was loose. In addition, there were some minor differences among the three genotypic in vitro
H. serrata, including the regeneration rate and other morphological characteristics. These results indicated that the genotypes had some effect on micropropagation in vitro
H. serrata.
2.5. HupA Detection in Thallus of H. serrata
In vitro propagation of the plant material that produces HupA has attracted attention in recent years. One of the purposes of this study is to evaluate the ability of HupA production in different genotypic in vitro
H. serrata thallus. HPLC analysis suggested that HupA content of the HB genotype was the highest among the three wild
H. serrata, which was 220.34 ± 3.08 µg·g
−1 (
Table 5). The HupA content of wild
H. serrata from FJ and JX was 198.12 ± 1.71 µg·g
−1 and 186.38 ± 1.84 µg·g
−1, respectively. The retention time for the HupA standard was 10.702 min (
Figure 4A). The chromatogram of the wild FJ genotype is shown in
Figure 4B, and the retention time was 10.682 min.
For the thallus, the HupA content of FJ genotype was the highest, which was 87.17 ± 5.15 µg·g
−1, and in the thallus of the JX and HB genotypes, the HupA content was 76.28 ± 4.26 µg·g
−1 and 53.90 ± 3.96 µg·g
−1, respectively. The chromatogram of FJ’s thallus is shown in
Figure 4C. The content of HupA in the thallus was about one-third of that in the corresponding wild
H. serrata. These results indicated that genotype and environment would significantly affect the accumulation of HupA in
H. serrata. Besides, thallus grew rapidly and was not limited by seasons. The accumulation of HupA in per-unit time was faster than that of wild
H. serrata.
The linear relationship between the peak area (y) and concentration (x, µg·mL−1) of the HupA standard is represented by the regression equation (y = 18182x − 14.312). The calibration curve of this compound shows good linearity (R2 = 1.0000).
2.6. The Antioxidant Activity Analysis
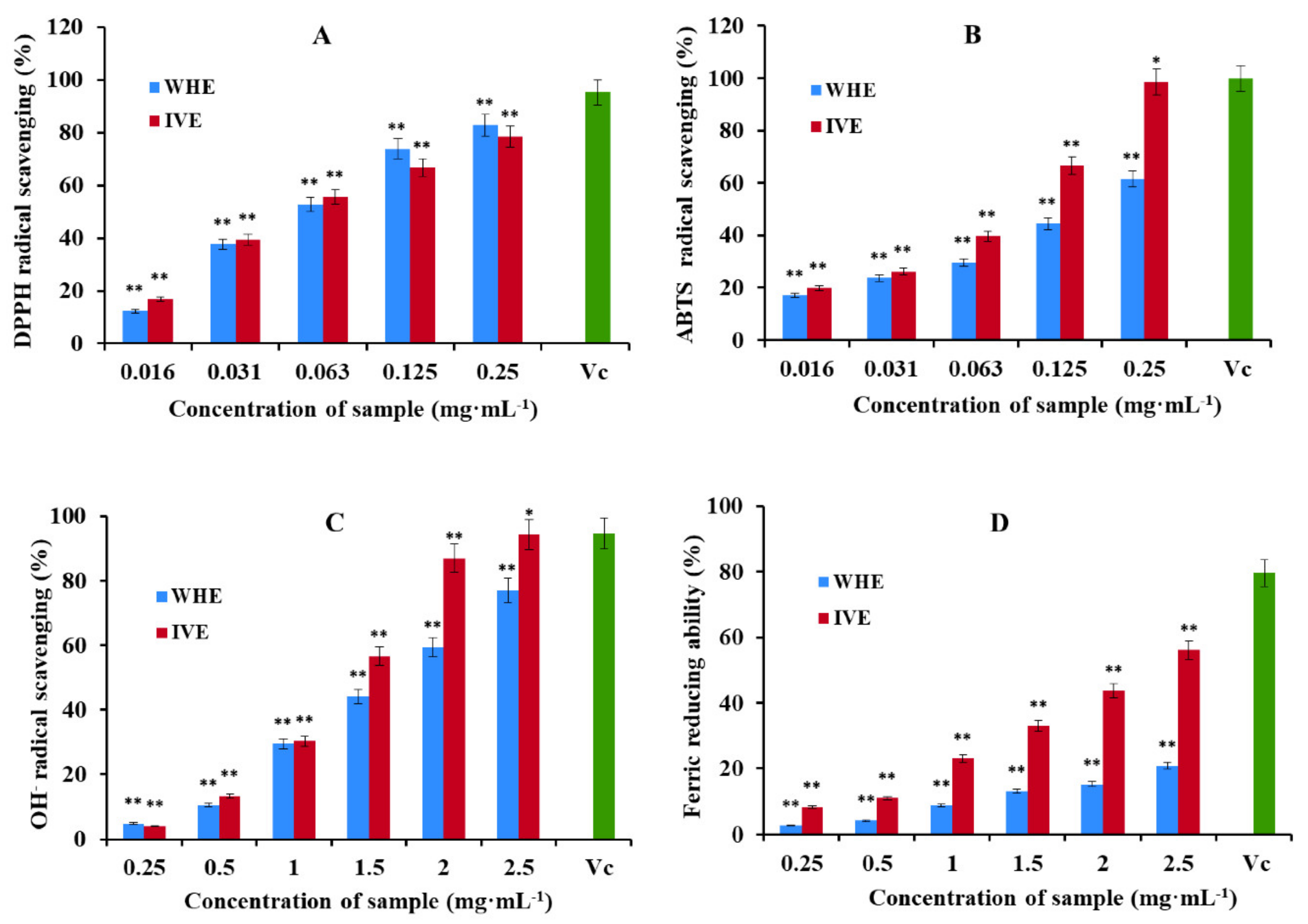
Figure 5 shows the relationship between antioxidant activity and different concentrations of samples in the form of a bar graph. DPPH and ABTS radical scavenging activity is shown in
Figure 5A,B. The results showed that the DPPH and ABTS radical scavenging activity of the in vitro
H. serrata methanol extract (IVE) was significantly greater than that of the wild
H. serrata methanol extract (WHE). The highest scavenging activity of IVE was 82.90 ± 0.56% for DPPH and 98.64 ± 1.00% for ABTS at the concentration of 0.25 mg·mL
−1. However, the DPPH and ABTS radical scavenging activity of WHE was 78.50 ± 0.45% and 61.59 ± 0.68%, respectively. IC50 values of IVE in DPPH and ABTS scavenging assays were 0.051 and 0.061 mg·mL
−1, respectively. IC50 values of WHE in DPPH and ABTS scavenging assays were 0.064 and 0.155 mg·mL
−1, respectively (
Table 6).
The OH
− scavenging activity and the ability to reduce Fe
3+ of WHE and IVE are shown in
Figure 5C,D. The results showed that the hydroxyl radical scavenging activity and the ability to reduce Fe
3+ of IVE was significantly greater than that of WHE at the same tested concentration. The highest scavenging activity of IVE was to 94.29 ± 1.02% for OH
− scavenging capacity and 56.07 ± 0.61% for the ability to reduce Fe
3+ value at the concentration of 2.5 mg·mL
−1. However, it is 77.05 ± 0.72% and 20.77 ± 0.84% in WHE, respectively. IC50 values of IVE in OH
− scavenging activity and the ability to reduce Fe
3+ were 1.37 and 2.52 mg·mL
−1, respectively. IC50 values of WHE in OH
− scavenging activity and the ability to reduce Fe
3+ were 1.67 and 9.73 mg·mL
−1, respectively (
Table 6).
The antioxidant capacity of plants is related to polyphenol compounds. Therefore, experiments were designed to detect the total polyphenol content of WHE and IVE. The linear relationship between the absorbance (y) and concentration (x, µg·mL−1) of the gallic acid standard is expressed by the regression equation y = 6.8x + 0.0597, R2 = 0.9935. The total polyphenol content of WHE was 12.97 ± 0.81 mgGAE/g. The total polyphenol content of IVE is 14.96 ± 0.65 mgGAE/g.
From the experiment of the antioxidant, it shows that in vitro H. serrata has a strong antioxidant effect. The extract of in vitro H. serrata had an excellent scavenging effect on DPPH, ABTS, and OH− and obviously had the ability to reduce Fe3+. Through the detection of total polyphenol content, micropropagation increased the antioxidant capacity by increasing the content of polyphenol compounds.
3. Discussion
H. serrata is an ancient fern plant group with important medicinal value. The fast-growing demand and the high price of the raw material are increasing the pressure on natural habitats, and
H. serrata has become a threatened plant in China due to the over-exploitation and habitat fragmentation. Furthermore, besides not being particularly abundant in wild resources, these plants also grow extremely slowly [
24,
25]. Thus, owing to the unique bioactivity of HupA and its low yield from plants, several research groups have devoted intensive efforts to study the in vitro culture of
H. serrata. However, the progress of these studies has been very slow and cannot be commercialized on a large scale. In order to alleviate the bottleneck of HupA production, it is of great significance to establish an in vitro rapid propagation system for
H. serrata, which would be very important for the sustainable development and utilization of
H. serrata.
Rapid propagation in vitro has the advantages of short growth cycle, high reproduction rate, and artificial control of culture conditions. One of the important reasons for slow progress is that
H. serrata is rich in endophytic fungi, which makes sterilization difficult [
26]. Moreover, the cuticle of
H. serrata leaves is thin, and it is easy to be killed by excessively disinfection. Therefore, the effective sterilization method is the key step for micropropagation of
H. serrata. Some researchers have carried out a series of research and investigations on the sterilization methods for
H. serrata explants [
27,
28]. Shen et al. used
H serrata stems as explants for tissue culture and found that it was very difficult to sterilize although it was sterilized several times [
29]. Szypula et al. found that the antibiotics would be helpful for removing the endophytic fungi of the explant [
30]. However, the survival rate of explants in previous studies was still very low. In this study, multiple sterilization methods were used to sterilize
H. serrata explants. Compared with previous studies, the survival rate of surviving explants without obvious bacterial and endophyte infection after three weeks was significantly improved, which was 25.37%. These results laid an important foundation for the subsequent induction and regeneration of
H. serrata.
During the past years, there were some successful reports on in vitro propagation of
H. serrata and its similar species. Szypula et al. selected the suitable medium for
H. selago explants surviving and growing [
30]. Ma and Gang succeeded in propagating
Phlegmariurus squarrosus (Forst.) in vitro and in detecting HupA produced by the corresponding cultivated plant [
18]. Shoot tips of
H. pinifolia were induced into callus, producing HupA [
31]. We also obtained HupA-producing
H. serrata thallus through in vitro culture [
32]. However, previous regeneration was limited to one genotype, and the micropropagation efficiency is not high. In the present study, an orthogonal experiment and mult-factor experiment were used to establish the in vitro rapid propagation system of
H. serrata. Three different genotypic thallus of
H. serrata in vitro were obtained, of which the regeneration rate (57.04%) and the biomass increased 164.17 ± 0.41 times. In addition, HPLC detection of its thallus could produce HupA. These results are of great significance for solving the resource shortage of
H. serrata for HupA production.
Previous studies suggested that the environmental conditions and genotype would play important roles in controlling HupA production in
Huperzia species [
33]. Ma et al. detected
H. serrata in Yunnan, Hunan, and Sichuan, and the content of HupA in its body was 148 µg·g
−1, 80.2 µg·g
−1, and 182.6 µg·g
−1 respectively [
9,
31]. Li et al. found that in
H. serrata from the Hunan Province (Baiyun Mountain), the Fujian Province (Nanping Municipal county), and the Jiangxi Province (Yifeng county), the levels of HupA are 233.1 µg·g
−1, 148.9 µg·g
−1, and 321.7 µg·g
−1, respectively [
34]. The results of this study showed that the content of HupA in the three different genotypes was significantly different. As for the content of Hup A in cultured tissue, Bao et al. used HPLC to determine the content of HupA in
H. serrata tissue culture and wild sporophytes. The results showed that the content of HupA in cultured tissue was one-fourth of that in wild sporophytes [
19]. Our experimental results show that the HupA content of in vitro
H. serrata was about one-third of that of corresponding wild
H. serrata. We also found that the content of HupA could be increased by adding some metabolic substrates during the proliferation culture [
35,
36]. Since the in vitro
H. serrata thallus grew rapidly and was not limited by seasons, this in vitro propagation system of
H. serrata would be able to relieve the shortage of Chinese medicinal resources for HupA production.
Previous studies have shown that micropropagation exhibited a remarkable influence on phytochemical contents and morphology of plants. Plants that undergo micropropagation have higher contents of phenolics and flavonoids and antioxidant activity compared to those developed from the wild [
37]. The phenols and flavonoids are related to the antioxidant properties of plants. Our results of this study found that the antioxidant activity of thallus was stronger than that of wild
H. serrata, indicating that the advantages of micropropagation of thallus to produce antioxidant metabolites makes it available to fulfill the high pharmaceutical demands. It opens up the practical application value of these rapidly propagating plants. The increased antioxidant capacity of the in vitro
H. serrata may be related to the culture environment and the enrichment of the culture medium during the propagation period. Plant growth regulators contained in micropropagation promote the biosynthesis of antioxidant-related compounds by influencing the expression or up-regulation of genes in the biosynthetic pathway of secondary metabolites.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}