
Effects of Elevated CO2 and Heat on Wheat Grain Quality



Abstract
1. Introduction
2. Wheat Quality and Grain Protein
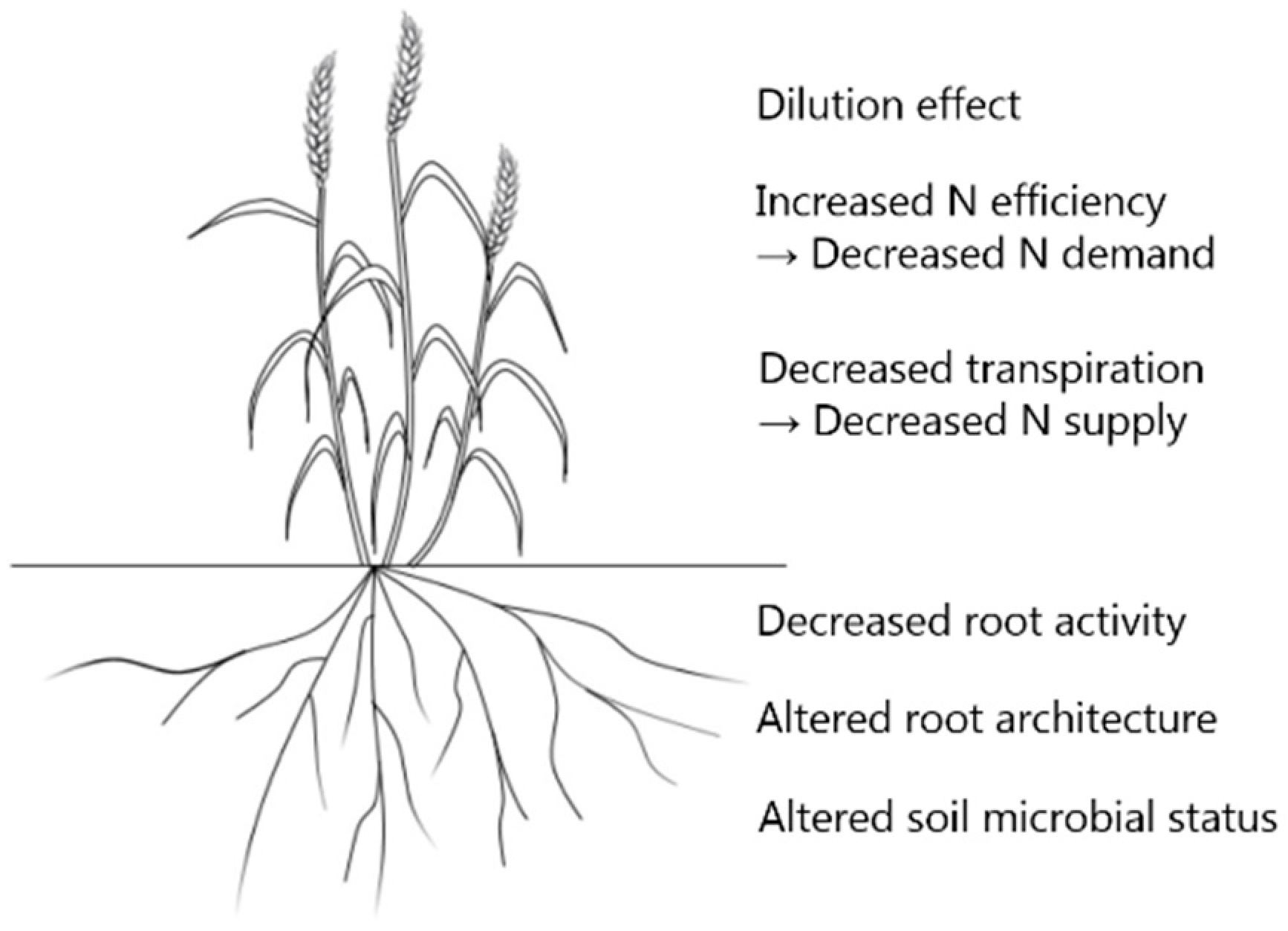
3. The Effects of eCO2 on Wheat Plants and the Mechanisms Behind
4. The Effects of Heat Stress on Wheat Grain Quality
5. The Interactive Effects of eCO2 and Heat Stress on Wheat Plants
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mirza, M.M.Q. Climate change and extreme weather events: Can developing countries adapt? Clim. Policy 2003, 3, 233–248. [Google Scholar] [CrossRef]
- Ray, D.K.; West, P.C.; Clark, M.; Gerber, J.S.; Prishchepov, A.V.; Chatterjee, S. Climate change has likely already affected global food production. PLoS ONE 2019, 14, e0217148. [Google Scholar] [CrossRef]
- Shewry, P.R.; Hey, S.J. The contribution of wheat to human diet and health. Food Energy Secur. 2015, 4, 178–202. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Schjoerring, J.K. Effects of elevated atmospheric CO2 on physiology and yield of wheat (Triticum aestivum L.): A meta-analytic test of current hypotheses. Agric. Ecosyst. Environ. 2013, 178, 57–63. [Google Scholar] [CrossRef]
- Högy, P.; Wieser, H.; Köhler, P.; Schwadorf, K.; Breuer, J.; Franzaring, J.; Muntifering, R.; Fangmeier, A. Effects of elevated CO2 on grain yield and quality of wheat: Results from a 3-year free-air CO2 enrichment experiment. Plant Biol. 2009, 11, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Huybers, P.; Leakey, A.D.B.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; et al. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Loladze, I. Rising atmospheric CO2 and human nutrition: Toward globally imbalanced plant stoichiometry? Trends Ecol. Evol. 2002, 17, 457–461. [Google Scholar] [CrossRef]
- Loladze, I. Hidden shift of the ionome of plants exposed to elevated CO2 depletes minerals at the base of human nutrition. Elife 2014, 2014, 1–29. [Google Scholar]
- Cossani, C.M.; Reynolds, M.P. Physiological Traits for Improving Heat Tolerance in Wheat 1 [W]. Plant Physiol. 2020, 160, 1710–1718. [Google Scholar] [CrossRef]
- Melaku, T.A. Environmental Impact on Processing Quality of Wheat Grain. Int. J. Food Sci. Nutr. Diet. 2019, 1–8. [Google Scholar] [CrossRef]
- Farooq, M.; Bramley, H.; Palta, J.A.; Siddique, K.H.M.M. Heat stress in wheat during reproductive and grain-filling phases. Crit. Rev. Plant Sci. 2011, 30, 491–507. [Google Scholar] [CrossRef]
- McDonald, G.K.; Sutton, B.G.; Ellison, F.W. The effect of time of sowing on the grain yield of irrigated wheat in the Namoi Valley, New South Wales. Aust. J. Agric. Res. 1983, 34, 229–240. [Google Scholar] [CrossRef]
- Mullarkey, M.; Jones, P. Isolation and analysis of thermotolerant mutants of wheat. J. Exp. Bot. 2000, 51, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Wardlaw, I.F.; Wrigley, C.W. Heat Tolerance in Temperate Cereals: An Overview. Funct. Plant Biol. 1994, 21, 695. [Google Scholar] [CrossRef]
- Saini, H.S.; Aspinall, D. Abnormal sporogenesis in wheat (Triticum aestivum L.) induced by short periods of high temperature. Ann. Bot. 1982, 49, 835–846. [Google Scholar] [CrossRef]
- Kaše, M.; Čatský, J. Maintenance and growth components of dark respiration rate in leaves of C3 and C4 plants as affected by leaf temperature. Biol. Plant. 1984, 26, 461–470. [Google Scholar] [CrossRef]
- Blumenthal, C.S.; Barlow, E.W.R.; Wrigley, C.W. Growth environment and wheat quality: The effect of heat stress on dough properties and gluten proteins. J. Cereal Sci. 1993, 18, 3–21. [Google Scholar] [CrossRef]
- Stone, P.; Nicolas, M. Wheat Cultivars Vary Widely in Their Responses of Grain Yield and Quality to Short Periods of Post-Anthesis Heat Stress. Funct. Plant Biol. 1994, 21, 887. [Google Scholar] [CrossRef]
- Fleitas, M.C.; Mondal, S.; Gerard, G.S.; Hernández-Espinosa, N.; Singh, R.P.; Crossa, J.; Guzmán, C. Identification of CIMMYT spring bread wheat germplasm maintaining superior grain yield and quality under heat-stress. J. Cereal Sci. 2020, 93, 102981. [Google Scholar] [CrossRef]
- Williams, R.M.; O’Brien, L.; Eagles, H.A.; Solah, V.A.; Jayasena, V. The influences of genotype, environment, and genotype ¾ environment interaction on wheat quality. Aust. J. Agric. Res. 2008, 59, 95–111. [Google Scholar] [CrossRef]
- Hernández-Espinosa, N.; Mondal, S.; Autrique, E.; Gonzalez-Santoyo, H.; Crossa, J.; Huerta-Espino, J.; Singh, R.P.; Guzmán, C. Milling, processing and end-use quality traits of CIMMYT spring bread wheat germplasm under drought and heat stress. Field Crop. Res. 2018, 215, 104–112. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Y.; Hernandez-Espinosa, N.; Peña, R.J. The influence of drought and heat stress on the expression of end-use quality parameters of common wheat. J. Cereal Sci. 2013, 57, 73–78. [Google Scholar] [CrossRef]
- Kimball, B.A.; Pinter, P.J.; Garcia, R.L.; Lamorte, R.L.; Wall, G.W.; Hunsaker, D.J.; Wechsung, G.; Wechsung, F.; Kartschall, T. Productivity and water use of wheat under free-air CO2 enrichment. Glob. Chang. Biol. 1995, 1, 429–442. [Google Scholar] [CrossRef]
- Morison, J.I.L. Response of plants to CO2 under water limited conditions. Vegetatio 1993, 104, 193–209. [Google Scholar] [CrossRef]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Peña, R.J. Wheat for bread and other foods. In Bread Wheat Improvement and Production; Food and Agriculture Organization of the United Nations: Rome, Italy, 2002; pp. 483–542. [Google Scholar]
- Battenfield, S.D.; Guzmán, C.; Gaynor, R.C.; Singh, R.P.; Peña, R.J.; Dreisigacker, S.; Fritz, A.K.; Poland, J.A. Genomic Selection for Processing and End-Use Quality Traits in the CIMMYT Spring Bread Wheat Breeding Program. Plant Genome 2016, 9. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, K.P.; Liu, B.; Deng, Z.Y.; Qu, H.L.; Tian, J.C. A comparison of grain protein content QTLs and flour protein content QTLs across environments in cultivated wheat. Euphytica 2010, 174, 325–335. [Google Scholar] [CrossRef]
- Coles, G.D.; Hartunian-Sowa, S.M.; Jamieson, P.D.; Hay, A.J.; Atwell, W.A.; Fulcher, R.G. Environmentally-induced variation in starch and non-starch polysaccharide content in wheat. J. Cereal Sci. 1997, 26, 47–54. [Google Scholar] [CrossRef]
- Arena, S.; D’Ambrosio, C.; Vitale, M.; Mazzeo, F.; Mamone, G.; Di Stasio, L.; Maccaferri, M.; Curci, P.L.; Sonnante, G.; Zambrano, N.; et al. Differential representation of albumins and globulins during grain development in durum wheat and its possible functional consequences. J. Proteom. 2017, 162, 86–98. [Google Scholar] [CrossRef] [PubMed]
- Wieser, H.; Seilmeier, W. The influence of nitrogen fertilisation on quantities and proportions of different protein types in wheat flour. J. Sci. Food Agric. 1998, 76, 49–55. [Google Scholar] [CrossRef]
- Payne, P.I.; Holt, L.M.; Jackson, E.A.; Law, C.N.; Damania, A.B.; Trans, P.; Lond, R.S. Wheat storage proteins: Their genetics and their potential for manipulation by plant breeding. Philos. Trans. R. Soc. Lond. B Biol. Sci. 1984, 304, 359–371. [Google Scholar]
- Daniel, C.; Triboi, E. Effects of temperature and nitrogen nutrition on the grain composition of winter wheat: Effects on gliadin content and composition. J. Cereal Sci. 2000, 32, 45–56. [Google Scholar] [CrossRef]
- Dier, M.; Hüther, L.; Schulze, W.X.; Erbs, M.; Köhler, P.; Weigel, H.J.; Manderscheid, R.; Zörb, C. Elevated Atmospheric CO2 Concentration Has Limited Effect on Wheat Grain Quality Regardless of Nitrogen Supply. J. Agric. Food Chem. 2020, 68, 3711–3721. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, A.K.; Hafskjold, R.; Kalhovd, A.H.; Sahlström, S.; Longva, Å.; Magnus, E.M. Effects of cultivar and temperature during grain filling on wheat protein content, composition, and dough mixing properties. Cereal Chem. 1998, 75, 460–465. [Google Scholar] [CrossRef]
- Kunkulberga, D.; Linina, A.; Ruza, A. Effect of nitrogen fertilization on protein content and rheological properties of winter wheat wholemeal. Foodbalt 2019, 2019, 88–92. [Google Scholar]
- Li, Y.; Wu, Y.; Hernandez-espinosa, N.; Peña, R.J. Heat and drought stress on durum wheat: Responses of genotypes, yield, and quality parameters. J. Cereal Sci. 2013, 57, 398–404. [Google Scholar] [CrossRef]
- Guzmán, C.; Autrique, J.E.; Mondal, S.; Singh, R.P.; Govindan, V.; Morales-dorantes, A.; Posadas-Romano, G.; Crossa, J.; Ammar, K.; Javier, R.; et al. Response to drought and heat stress on wheat quality, with special emphasis on bread-making quality, in durum wheat. Field Crop. Res. 2016, 186, 157–165. [Google Scholar] [CrossRef]
- Joshi, A.K.; Crossa, J.; Arun, B.; Chand, R.; Trethowan, R.; Vargas, M.; Ortiz-monasterio, I. Genotype x environment interaction for zinc and iron concentration of wheat grain in eastern Gangetic plains of India. Field Crop. Res. 2010, 116, 268–277. [Google Scholar] [CrossRef]
- Long, S.P.; Ainsworth, E.A.; Rogers, A.; Ort, D.R. Rising Atmospheric Carbon Dioxide: Plants FACE the future. Annu. Rev. Plant Biol. 2004, 55, 591–628. [Google Scholar] [CrossRef]
- Broberg, M.C.; Högy, P.; Feng, Z.; Pleijel, H. Effects of elevated CO2 on wheat yield: Non-linear response and relation to site productivity. Agronomy 2019, 9, 243. [Google Scholar] [CrossRef]
- Wardlaw, I.F.; Porter, H.K. The redistribution of stem sugars in wheat during grain development. Aust. J. Biol. Sci. 1967, 20, 309–318. [Google Scholar] [CrossRef]
- Evans, L.T.; Rawson, H.M. Photosynthesis and respiration by the flag leaf and components of the ear during grain development in wheat. Aust. J. Biol. Sci. 1970, 23, 245–254. [Google Scholar] [CrossRef]
- Ainsworth, E.A. Rice production in a changing climate: A meta-analysis of responses to elevated carbon dioxide and elevated ozone concentration. Glob. Chang. Biol. 2008, 14, 1642–1650. [Google Scholar] [CrossRef]
- Wang, J.; Wang, C.; Chen, N.; Xiong, Z.; Wolfe, D.; Zou, J. Response of rice production to elevated [CO2] and its interaction with rising temperature or nitrogen supply: A meta-analysis. Clim. Chang. 2015, 130, 529–543. [Google Scholar] [CrossRef]
- Lv, C.; Huang, Y.; Sun, W.; Yu, L.; Zhu, J. Response of rice yield and yield components to elevated [CO2]: A synthesis of updated data from FACE experiments. Eur. J. Agron. 2020, 112, 125961. [Google Scholar] [CrossRef]
- Barnes, J.D.; Ollerenshaw, J.H.; Whitfield, C.P. Effects of elevated CO2 and/or O3 on growth, development and physiology of wheat (Triticum aestivum L.). Glob. Chang. Biol. 1995, 1, 129–142. [Google Scholar] [CrossRef]
- Kimball, B.A. Carbon Dioxide and Agricultural Yield: An Assemblage and Analysis of 430 Prior Observations 1. Agron. J. 1983, 75, 779–788. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Li, X.; Tang, Y.; Zhang, P.; Duan, Z. Sustainable vegetable production under changing climate: The impact of elevated CO2 on yield of vegetables and the interactions with environments-A review. J. Clean. Prod. 2020, 253, 119920. [Google Scholar] [CrossRef]
- Fierro, A.; Tremblay, N.; Gosselin, A. Supplemental carbon dioxide and light improved tomato and pepper seedling growth and yield. HortScience 1994, 29, 152–154. [Google Scholar] [CrossRef]
- Demmers-Derks, H.; Mitchell, R.A.C.; Mitchell, V.J.; Lawlor, D.W. Response of sugar beet (Beta vulgaris L.) yield and biochemical composition to elevated CO2 and temperature at two nitrogen applications. Plant Cell Environ. 1998, 21, 829–836. [Google Scholar] [CrossRef]
- Chumley, H.; Hewlings, S. The effects of elevated atmospheric carbon dioxide [CO2] on micronutrient concentration, specifically iron (Fe) and zinc (Zn) in rice; a systematic review. J. Plant Nutr. 2020, 43, 1571–1578. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Lam, S.K.; Li, X.; Duan, Z. Effects of elevated CO2 on nutritional quality of vegetables: A review. Front. Plant Sci. 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Taub, D.R.; Miller, B.; Allen, H. Effects of elevated CO2 on the protein concentration of food crops: A meta-analysis. Glob. Chang. Biol. 2008, 14, 565–575. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Dong, G.; Gu, H.; Huang, J.; Zhu, J.; Yang, H.; Liu, G.; Han, Y. The impact of free-air CO2 enrichment (FACE) and nitrogen supply on grain quality of rice. Field Crop. Res. 2007, 102, 128–140. [Google Scholar] [CrossRef]
- Li, X.; Jiang, D.; Liu, F. Dynamics of amino acid carbon and nitrogen and relationship with grain protein in wheat under elevated CO2 and soil warming. Environ. Exp. Bot. 2016, 132, 121–129. [Google Scholar] [CrossRef]
- Blandino, M.; Badeck, F.W.; Giordano, D.; Marti, A.; Rizza, F.; Scarpino, V.; Vaccino, P. Elevated CO2 Impact on Common Wheat (Triticum aestivum L.) Yield, Wholemeal Quality, and Sanitary Risk. J. Agric. Food Chem. 2020, 68, 10574–10585. [Google Scholar] [CrossRef]
- Panozzo, J.F.; Walker, C.K.; Partington, D.L.; Neumann, N.C.; Tausz, M.; Seneweera, S.; Fitzgerald, G.J. Elevated carbon dioxide changes grain protein concentration and composition and compromises baking quality. A FACE study. J. Cereal Sci. 2014, 60, 461–470. [Google Scholar] [CrossRef]
- Erbs, M.; Manderscheid, R.; Jansen, G.; Seddig, S.; Pacholski, A.; Weigel, H.J. Effects of free-air CO2 enrichment and nitrogen supply on grain quality parameters and elemental composition of wheat and barley grown in a crop rotation. Agric. Ecosyst. Environ. 2010, 136, 59–68. [Google Scholar] [CrossRef]
- Fernando, N.; Panozzo, J.; Tausz, M.; Norton, R.; Fitzgerald, G.; Seneweera, S. Rising atmospheric CO2 concentration affects mineral nutrient and protein concentration of wheat grain. Food Chem. 2012, 133, 1307–1311. [Google Scholar] [CrossRef]
- Taub, D.R.; Wang, X. Why are nitrogen concentrations in plant tissues lower under elevated CO2? A critical examination of the hypotheses. J. Integr. Plant Biol. 2008, 50, 1365–1374. [Google Scholar] [CrossRef]
- Kuehny, J.S.; Peet, M.M.; Nelson, P.V.; Willits, D.H. Nutrient dilution by starch in CO2-enriched chrysanthemum. J. Exp. Bot. 1991, 42, 711–716. [Google Scholar] [CrossRef]
- Pandey, V.; Sharma, M.; Deeba, F.; Maurya, V.K.; Gupta, S.K.; Singh, S.P.; Mishra, A.; Nautiyal, C.S. Impact of Elevated CO2 on Wheat Growth and Yield under Free Air CO2 Enrichment Impact of Elevated CO2 on Wheat Growth and Yield under Free Air CO2 Enrichment. Am. J. Clim. Chang. 2017, 6, 573–596. [Google Scholar] [CrossRef]
- Gifford, R.M.; Barrett, D.J.; Lutze, J.L. The effects of elevated [CO2] on the C:N and C:P mass ratios of plant tissues. Plant Soil 2000, 224, 1–14. [Google Scholar] [CrossRef]
- Henning, F.P.; Wood, C.W.; Rogers, H.H.; Runion, G.B.; Prior, S.A. Composition and decomposition of soybean and sorghum tissues grown under elevated atmospheric carbon dioxide. J. Environ. Qual. 1996, 25, 822–827. [Google Scholar] [CrossRef]
- Chu, C.C.; Field, C.B.; Mooney, H.A.; Chu, C.C.; Field, C.B.; Mooney, H.A. Effects of CO2 and nutrient enrichment on tissue quality of two California annuals. Oecologia 1996, 107, 433–440. [Google Scholar] [CrossRef]
- Hattenschwiler, S.; Schweingruber, F.H.; Korner, C. Tree ring responses to elevated CO2 and increased N deposition in Picea abies. Plant Cell Environ. 1996, 19, 1369–1378. [Google Scholar] [CrossRef]
- Davidson, R.L. Effect of root/leaf temperature differentials on root/shoot ratios in some pasture grasses and clover. Ann. Bot. 1969, 33, 561–569. [Google Scholar] [CrossRef]
- BassiriRad, H.; Gutschick, V.P.; Lussenhop, J. Root system adjustments: Regulation of plant nutrient uptake and growth responses to elevated CO2. Oecologia 2001, 126, 305–320. [Google Scholar] [CrossRef]
- Ellsworth, D.S.; Reich, P.B.; Naumburg, E.S.; Koch, G.W.; Kubiske, M.E.; Smith, S.D. Photosynthesis, carboxylation and leaf nitrogen responses of 16 species to elevated pCO2 across four free-air CO2 enrichment experiments in forest, grassland and desert. Glob. Chang. Biol. 2004, 10, 2121–2138. [Google Scholar] [CrossRef]
- Runion, G.B.; Entry, J.A.; Prior, S.A.; Mitchell, R.J.; Rogers, H.H. Tissue chemistry and carbon allocation in seedlings of Pinus palustris subjected to elevated atmospheric CO2 and water stress. Tree Physiol. 1999, 19, 329–335. [Google Scholar] [CrossRef][Green Version]
- Li, X.; Jiang, D.; Liu, F. Soil warming enhances the hidden shift of elemental stoichiometry by elevated CO2 in wheat. Sci. Rep. 2016, 6, 23313. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Funct. Plant Biol. 2000, 27, 1191. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Ecological Society of America effects of c02 and temperature on growth and. Ecology 2015, 73, 1244–1259. [Google Scholar]
- Israel, D.W.; Rufty, T.W., Jr.; Cure, J.D. Nitrogen and phosphorus nutritional interactions in a CO2 enriched environment. J. Plant Nutr. 1990, 13, 1419–1433. [Google Scholar] [CrossRef]
- Zerihun, A.; Gutschick, V.P.; Bassirirad, H. Compensatory roles of nitrogen uptake and photosynthetic N-use efficiency in determining plant growth response to elevated CO2: Evaluation using a functional balance model. Ann. Bot. 2000, 86, 723–730. [Google Scholar] [CrossRef]
- Chu, C.C.; Coleman, J.S.; Mooney, H.A.; Chu, C.C.; Coleman, J.S.; Mooney, H.A. Controls of biomass partitioning between roots and shoots: Atmospheric CO2 enrichment and the acquisition and allocation of carbon and nitrogen in wild radish. Oecologia 1992, 89, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Gloser, V.; Frehner, M.; Luscher, A.; Nosberger, J.; Hartwig, U.A. Does the response of perennial ryegrass to elevated CO2 concentration depend on the from of the supplied nitrogen? Biol. Plant. 2002, 45, 51–58. [Google Scholar] [CrossRef]
- Locke, A.M.; Ort, D.R. Diurnal depression in leaf hydraulic conductance at ambient and elevated [CO2] reveals anisohydric water management in fi eld-grown soybean and possible involvement of aquaporins. Environ. Exp. Bot. 2015, 116, 39–46. [Google Scholar] [CrossRef]
- Mcgrath, J.M.; Lobell, D.B. Reduction of transpiration and altered nutrient allocation contribute to nutrient decline of crops grown in elevated CO2 concentrations. Plant Cell Environ. 2013, 36, 697–705. [Google Scholar] [CrossRef]
- Chavan, S.G.; Duursma, R.A.; Tausz, M.; Ghannoum, O. Elevated CO2 alleviates the negative impact of heat stress on wheat physiology but not on grain yield. J. Exp. Bot. 2019, 70, 6447–6459. [Google Scholar] [CrossRef]
- Yang, J.; Sears, R.G.; Gill, B.S.; Paulsen, G.M. Genotypic differences in utilization of assimilate sources during maturation of wheat under chronic and heat shock stresses: Utilization of assimilate sources by wheat under heat stresses. Euphytica 2002, 125, 179–188. [Google Scholar] [CrossRef]
- Wrigley, C.; Blumenthal, C.; Gras, P.; Barlow, E. Temperature Variation during Grain Filling and Changes in Wheat-Grain Quality. Funct. Plant Biol. 1994, 21, 875. [Google Scholar] [CrossRef]
- Castro, M.; Peterson, C.J.; Rizza, M.D.; Dellavalle, P.D.í.; Vázquez, D.; IbáÑez, V.; Ross, A. Influence of Heat Stress on Wheat Grain Characteristics and Protein Molecular Weight Distribution. Mol. Breed. Forage Turf 2007, 12, 365–371. [Google Scholar]
- Spiertz, J.H.J.; Hamer, R.J.; Xu, H.; Primo-Martin, C.; Don, C.; van der Putten, P.E.L. Heat stress in wheat (Triticum aestivum L.): Effects on grain growth and quality traits. Eur. J. Agron. 2006, 25, 89–95. [Google Scholar] [CrossRef]
- Sofield, I.; Evans, L.; Cook, M.; Wardlaw, I. Factors Influencing the Rate and Duration of Grain Filling in Wheat. Funct. Plant Biol. 1977, 4, 785. [Google Scholar] [CrossRef]
- Corbellini, M.; Canevar, M.G.; Mazza, L.; Ciaffi, M.; Lafiandra, D.; Borghi, B. Effect of the duration and intensity of heat shock during grain filling on dry matter and protein accumulation, technological quality and protein composition in bread wheat and durum wheat. Funct. Plant Biol. 1997, 24, 245–260. [Google Scholar] [CrossRef]
- Bhullar, S.; Jenner, C. Differential Responses to High Temperatures of Starch and Nitrogen Accumulation in the Grain of Four Cultivars of Wheat. Funct. Plant Biol. 1985, 12, 363. [Google Scholar] [CrossRef]
- Randall, P.J.; Moss, H.J. Some effects of temperature regime during grain filling on wheat quality. Aust. J. Agric. Res. 1990, 41, 603–617. [Google Scholar] [CrossRef]
- Ferris, R.; Ellis, R.H.; Wheeler, T.R.; Hadley, P. Effect of High Temperature Stress at Anthesis on Grain Yield and Biomass of Field-grown Crops of Wheat. Ann. Bot. 1998, 82, 631–639. [Google Scholar] [CrossRef]
- Stone, P.J.; Nicolas, M.E. The effect of duration of heat stress during grain filling on two wheat varieties differing in heat tolerance: Grain growth and fractional protein accumulation. Aust. J. Plant Physiol. 1998, 25, 13–20. [Google Scholar] [CrossRef]
- Tahir, I.S.A.; Nakata, N. Remobilization of nitrogen and carbohydrate from stems of bread wheat in response to heat stress during grain filling. J. Agron. Crop Sci. 2005, 191, 106–115. [Google Scholar] [CrossRef]
- Viswanathan, C.; Khanna-Chopra, R. Effect of heat stress on grain growth, starch synthesis and protein synthesis in grains of wheat (Triticum aestivum L.) varieties differing in grain weight stability. J. Agron. Crop Sci. 2001, 186, 1–7. [Google Scholar] [CrossRef]
- Wardlaw, F.; Sofieldb, I.; Cartwrightc, P.M. Factors Limiting the Rate of Dry Matter Accumulation in the Grain of Wheat Grown at High Temperature. Funct. Plant Biol. 1980, 7, 387–400. [Google Scholar] [CrossRef]
- Bechtel, D.B.; Zayas, I.; Kaleikau, L.; Pomeranz, Y. Size-Distribution of Wheat Starch Granules during Endosperm Development.pdf. Cereal Chem. 1990, 67, 59–63. [Google Scholar]
- Hurkman, W.J.; McCue, K.F.; Altenbach, S.B.; Korn, A.; Tanaka, C.K.; Kothari, K.M.; Johnson, E.L.; Bechtel, D.B.; Wilson, J.D.; Anderson, O.D.; et al. Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm. Plant Sci. 2003, 164, 873–881. [Google Scholar] [CrossRef]
- Ball, S.G.; Van De Wal, M.H.; Visser, R.G.F. Progress in understanding the biosynthesis of amylose. Trends Plant Sci. 1998, 3, 462–467. [Google Scholar] [CrossRef]
- Myers, A.M.; Morell, M.K.; James, M.G.; Ball, S.G. Update on Biochemistry Recent Progress toward Understanding Biosynthesis of the Amylopectin Crystal 1. Plant Physiol. 2000, 122, 989–997. [Google Scholar] [CrossRef]
- Nuttall, J.G.; O’Leary, G.J.; Panozzo, J.F.; Walker, C.K.; Barlow, K.M.; Fitzgerald, G.J. Models of grain quality in wheat—A review. Field Crop. Res. 2017, 202, 136–145. [Google Scholar] [CrossRef]
- Keeling, P.L.; Bacon, P.J.; Holt, D.C. Elevated temperature reduces starch deposition in wheat endosperm by reducing the activity of soluble starch synthase. Planta 1993, 191, 342–348. [Google Scholar] [CrossRef]
- Rijven, A.H.G.C. Heat Inactivation of Starch Synthase in Wheat Endosperm Tissue. Plant Physiol. 1986, 81, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Blumenthal, C.; Stone, P.J.; Gras, P.W.; Bekes, F.; Clarke, B.; Barlow, E.W.R.; Appels, R.; Wrigley, C.W. Heat-shock protein 70 and dough-quality changes resulting from heat stress during grain filling in wheat. Cereal Chem. 1998, 75, 43–50. [Google Scholar] [CrossRef]
- Majoul, T.; Bancel, E.; Triboi, E.; Ben Hamida, J.; Branlard, G. Proteomic analysis of the effect of heat stress on hexaploid wheat grain: Characterization of heat-responsive proteins from total endosperm. Proteomics 2003, 3, 175–183. [Google Scholar] [CrossRef]
- Blumenthal, C.S.; Batey, I.L.; Bekes, F.; Wrigley, C.W.; Barlow, E.W.R. Gliadin genes contain heat-shock elements: Possible relevance to heat-induced changes in grain quality. J. Cereal Sci. 1990, 11, 185–188. [Google Scholar] [CrossRef]
- Blumenthal, C.; Batey, I.; Wrigley, C.; Barlow, E. Involvement of a Novel Peptide in the Heat Shock Response of Australian Wheats. Funct. Plant Biol. 1990, 17, 441. [Google Scholar] [CrossRef]
- Ciaffi, M.; Colapricoz, B.M.G.; De Stefanis, E. Effect of high temperature during grain filling on the amount of insoluble proteins in durum wheat. J. Genet. Breed. 1995, 49, 285–296. [Google Scholar]
- Kumar, S.; Kumar, R.; Kumar, P.; Singh, S.K. Comparative study of Fe and Mn micronutrient accumulation in flag leaf and spike of wheat (Triticum aestivum L.) grown under heat stress. J. Pharmacogn. Phytochem. 2018, 7, 2979–2982. [Google Scholar]
- Dias, A.S.; Lidon, F.C.; Ramalho, J.C. IV. Heat stress in triticum: Kinetics of Fe and Mn accumulation. Braz. J. Plant Physiol. 2009, 21, 153–164. [Google Scholar] [CrossRef]
- Narendra, M.C.; Roy, C.; Kumar, S.; Virk, P.; De, N. Effect of terminal heat stress on physiological traits, grain zinc and iron content in wheat (Triticum aestivum L.). Czech J. Genet. Plant Breed. 2021, 57, 43–50. [Google Scholar]
- Peck, A.W.; Mcdonald, G.K.Ã.; Graham, R.D. Zinc nutrition influences the protein composition of flour in bread wheat (Triticum aestivum L.). J. Cereal Sci. 2008, 47, 266–274. [Google Scholar] [CrossRef]
- Kadam, N.N.; Xiao, G.; Melgar, R.J.; Bahuguna, R.N.; Quinones, C.; Tamilselvan, A.; Prasad, P.V.V.; Jagadish, K.S.V. Agronomic and physiological responses to high temperature, drought, and elevated CO2 interactions in cereals. Adv. Agron. 2014, 127, 111–156. [Google Scholar]
- LONG, S.P. Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentrations: Has its importance been underestimated? Plant. Cell Environ. 1991, 14, 729–739. [Google Scholar] [CrossRef]
- Abdelhakim, L.O.A.; Palma, C.F.F.; Zhou, R.; Wollenweber, B.; Ottosen, C.O.; Rosenqvist, E. The effect of individual and combined drought and heat stress under elevated CO2 on physiological responses in spring wheat genotypes. Plant Physiol. Biochem. 2021, 162, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Macabuhay, A.; Houshmandfar, A.; Nuttall, J.; Fitzgerald, G.J.; Tausz, M.; Tausz-Posch, S. Can elevated CO2 buffer the effects of heat waves on wheat in a dryland cropping system? Environ. Exp. Bot. 2018, 155, 578–588. [Google Scholar] [CrossRef]
- Kannojiya, S.; Singh, S.D.; Prasad, S.; Kumar, S.; Malav, L.C.; Kumar, V. Effect of elevated temperature and carbon dioxide on wheat (Triticum aestivum) productivity with and without weed interaction. Indian J. Agric. Sci. 2019, 89, 751–756. [Google Scholar]
- Wieser, H.; Manderscheid, R.; Erbs, M.; Weigel, H.-J. Effects of Elevated Atmospheric CO2 Concentrations on the Quantitative Protein Composition of Wheat Grain. Agric. Food Chem. 2008, 56, 6531–6535. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, A.K.; Bahuguna, R.N.; Pal, M.; Shah, D.; Maurya, S.; Jagadish, K.S.V. Elevated CO2 and heat stress interactions affect grain yield, quality and mineral nutrient composition in rice under field conditions. Field Crop. Res. 2017, 206, 149–157. [Google Scholar] [CrossRef]


{kind=link}
| Traits | Impacts | References |
|---|---|---|
| Gliadin/glutenin ratio | +/−/ns | [17,18] |
| Grain protein content (GPC) | + | [21,35] |
| Starch content | − | [86,87,88] |
| Dough strength (W) | +/−/ns | [21,37,38] |
| Gluten extensibility (L) | + | [19,22] |
| Dough tenacity (P) | − | [22] |
| Bread loaf volume (LV) | + | [22,38] |
| Micronutrient concentration (Zn, Fe) | +/− | [38,39] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Liu, F. Effects of Elevated CO2 and Heat on Wheat Grain Quality. Plants 2021, 10, 1027. https://doi.org/10.3390/plants10051027
Wang X, Liu F. Effects of Elevated CO2 and Heat on Wheat Grain Quality. Plants. 2021; 10(5):1027. https://doi.org/10.3390/plants10051027
Chicago/Turabian StyleWang, Xizi, and Fulai Liu. 2021. "Effects of Elevated CO2 and Heat on Wheat Grain Quality" Plants 10, no. 5: 1027. https://doi.org/10.3390/plants10051027
APA StyleWang, X., & Liu, F. (2021). Effects of Elevated CO2 and Heat on Wheat Grain Quality. Plants, 10(5), 1027. https://doi.org/10.3390/plants10051027