Effect of Lead and Copper on Photosynthetic Apparatus in Citrus (Citrus aurantium L.) Plants. The Role of Antioxidants in Oxidative Damage as a Response to Heavy Metal Stress




Abstract
1. Introduction
2. Results
2.1. Effects of Copper and Lead on Plant Growth and Toxicity
2.2. Effect of Copper and Lead on Chlorophylls and Carotenoid Concentration
2.3. Effect of Copper and Lead on Chlorophyll Fluorescence
2.4. Effect of Copper and Lead on H2O2 and Lipid Peroxidation
2.5. Effect of Copper and Lead on Total Flavonoids and Phenolics
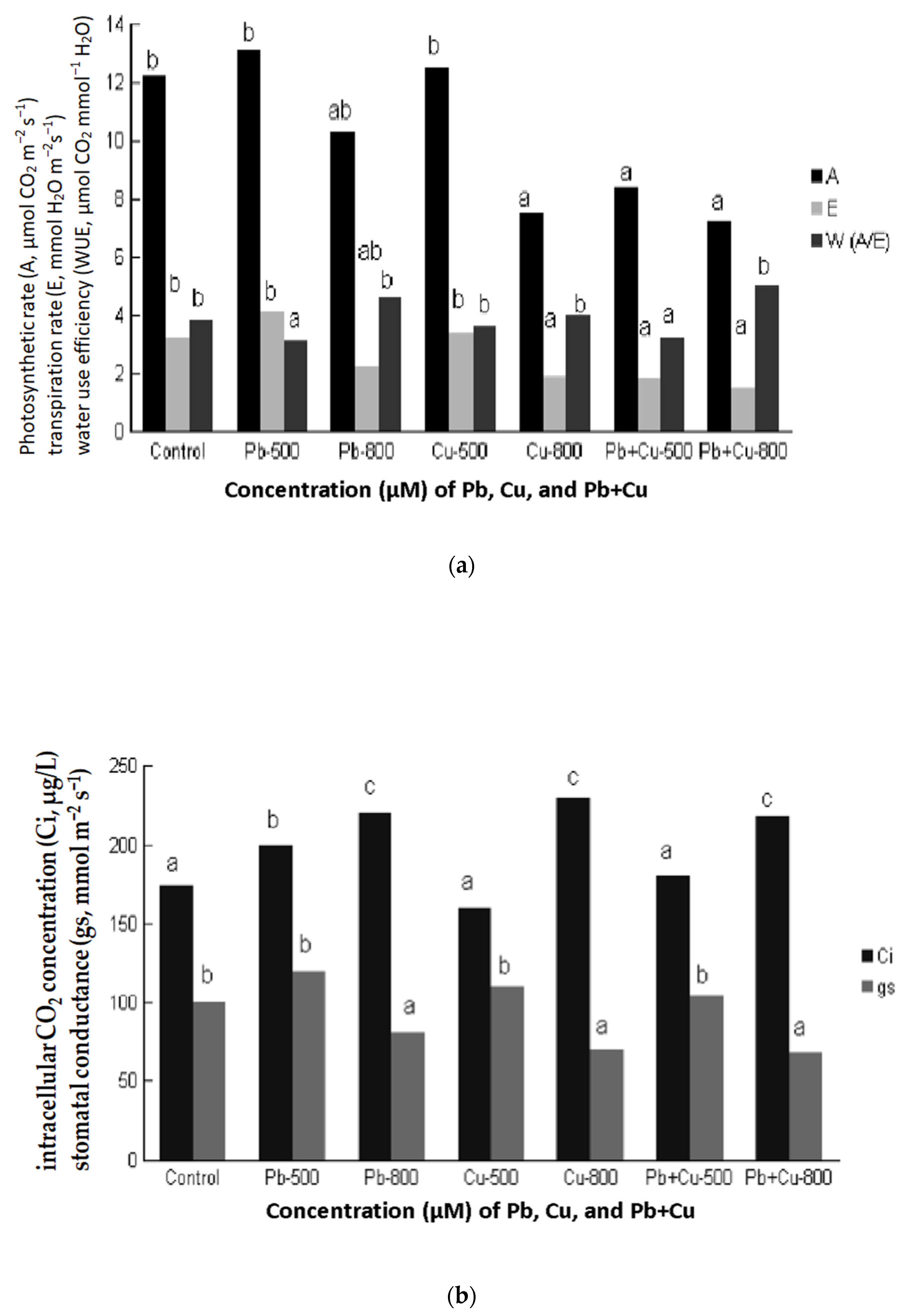
2.6. Effect of Cu + Pb on Leaf Gas Exchange
3. Discussion
4. Material and Methods
4.1. Plant Material and Culture
4.2. Chlorophyll and Carotenoid Estimation
4.3. In Vivo Chlorophyll Fluorescence Measurements
4.4. Photosynthesis Measurements
4.5. MDA Content
4.6. Determination of H2O2 Concentration
4.7. Sample Preparation for Antioxidants
4.8. Evaluation of Total Flavonoid Content
4.9. Determination of Phenolics
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Heckathorn, S.A.; Mueller, J.K.; LaGuidice, S.; Zhu, B.; Barrett, T.; Blair, B.; Dong, Y. Chloroplast small heat-shock proteins protect photosynthesis during heavy metal stress. Am. J. Bot. 2004, 91, 1312–1318. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Moustaka, J.; Ouzounidou, G.; Bayçu, G.; Moustakas, M. Aluminum resistance in wheat involves maintenance of leaf Ca2+ and Mg2+ content, decreased lipid peroxidation and Al accumulation, and low photosystem II excitation pressure. BioMetals 2016, 29, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Cortés, V.; Phelps, B.; Van Ryswyk, H.; Srebotnjak, T. Experimental Analysis of Soil and Mandarin Orange Plants Treated with Heavy Metals Found in Oilfield-Produced Wastewater. Sustainability 2018, 10, 1493. [Google Scholar] [CrossRef]
- Panou-Filotheou, H.; Bosabalidis, A.M.; Karataglis, S. Effects of Copper Toxicity on Leaves of Oregano (Origanum vulgare subsp. hirtum). Ann. Bot. 2001, 88, 207–214. [Google Scholar] [CrossRef]
- Rocchetta, I.; Küpper, H. Chromium and copper induced inhibition of photosynthesis in Euglena gracilis analysed on the single-cell level by fluorescence kinetic microscopy. New Phytol. 2009, 182, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Malecka, A.; Piechalak, A.; Tomaszewska, B. Reactive oxygen species production and anioxidative defense system in pea root tissues treated with lead ions the whole root level. Acta Physiol. Plant. 2009, 31, 1053–1063. [Google Scholar] [CrossRef]
- Petrovic, D.; Krivokapic, S. The Effect of Cu, Zn, Cd, and Pb Accumulation on Biochemical Parameters (Proline, Chlorophyll) in the Water Caltrop (Trapa natans L.), Lake Skadar, Montenegro. Plants 2020, 9, 1287. [Google Scholar] [CrossRef]
- Kumar, P.B.A.N.; Dushenkov, V.; Motto, H.; Raskin, I. Phytoextraction: The Use of Plants To Remove Heavy Metals from Soils. Environ. Sci. Technol. 1995, 29, 1232–1238. [Google Scholar] [CrossRef]
- Chaney, R.I.; Ryan, J.A. Risk Based Standars Standars for Arsenic Lead and Cadmium in Urban Soils; Oechema: Frankfurt Germany, 1994. [Google Scholar]
- Lane, S.D.; Martin, E.S. A histochemical investigation of lead uptake in raphanus sativus. New Phytol. 1977, 79, 281–286. [Google Scholar] [CrossRef]
- Zulfiqar, U.; Farooq, M.; Hussain, S.; Maqsood, M.; Hussaind, M.; Ishfaqa, M.; Ahmada, M.; Anjumf, M.Z. Lead toxicity in plants: Impacts and remediation. J. Environ. Manag. 2019, 250, 109557. [Google Scholar] [CrossRef] [PubMed]
- Mashhadi, M.; Boojar, A.; Goodarzi, F. The copper tolerance strategies and the role of antioxidative enzymes in three plant species grown on copper mine. Chemosphere 2007, 67, 2138–2147. [Google Scholar]
- Kumar, A.; Prasad, M.; Sytar, O. Lead toxicity, defense strategies and associated indicative biomarkers in Talinum triangulare grown hydroponically. Chemosphere 2012, 89, 1056–1065. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, D.; Johnston, K.M.; Dietrich, A.M. Fate and Transport of Copper-based Crop Protectants in Plasticulture Runoff and the Impact of Sedimentation as a Best Management Practice. Water Res. 2001, 35, 2984–2994. [Google Scholar] [CrossRef]
- Yusuf, M.; Fariduddin, Q.; Varshney, P.; Ahmad, A. Salicylic acid minimizes nickel and/or salinity-induced toxici-ty in Indian mustard (Brassica juncea) through an improved antioxidant system. Environ. Sci. Pollut. Res. 2012, 19, 8–18. [Google Scholar] [CrossRef]
- Polle, A.; Pfirrmann, T.; Chakrabarti, S.; Rennenberg, H. The effects of enhanced ozone and enhanced carbon dioxide concentrations on biomass, pigments and antioxidative enzymes in spruce needles (Picea abies L.). Plant Cell Environ. 1993, 16, 311–316. [Google Scholar] [CrossRef]
- Schützendübel, A.; Nikolova, P.; Rudolf, C.; Polle, A. Cadmium and H2O2-induced oxidative stress in Populus × canescens roots. Plant Physiol. Biochem. 2002, 40, 577–584. [Google Scholar] [CrossRef]
- Ames, B.N.; Shigenaga, M.K.; Hagen, T.M. Oxidants, antioxidants, and the degenerative diseases of aging. Proc. Natl. Acad. Sci. USA 1993, 90, 7915–7922. [Google Scholar] [CrossRef]
- Ahmad, M.S.A.; Ashraf, M.; Tabassam, Q.; Hussain, M.; Firdous, H. Lead (Pb)- induced regulation of growth, pho-tosynthesis and mineral nutrition in maize (Zea mays L.) plants at early growth stages. Biol. Trace Elem. Res. 2011, 44, 1229–1239. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic Compounds and Their Antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. Stud. 2006, 15, 523. [Google Scholar]
- Havsteen, B. Flavonoids, a class of natural products of high pharmacological potency. Biochem. Pharmacol. 1983, 32, 1141–1148. [Google Scholar] [CrossRef]
- Tahara, S. A Journey of Twenty-Five Years through the Ecological Biochemistry of Flavonoids. Biosci. Biotechnol. Biochem. 2007, 71, 1387–1404. [Google Scholar] [CrossRef] [PubMed]
- Posmyk, M.M.; Kontek, R.; Janas, K. Antioxidant enzymes activity and phenolic compounds content in red cabbage seedlings exposed to copper stress. Ecotoxicol. Environ. Saf. 2009, 72, 596–602. [Google Scholar] [CrossRef] [PubMed]
- Shu, X.; Yin, L.; Zhang, Q.; Wang, W. Effect of Pb toxicity on leaf growth, antioxidant enzyme activities, and photosynthesis in cuttings and seedlings of Jatropha curcas L. Environ. Sci. Pollut. Res. 2012, 19, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, M.; Ouzounidou, G.; Symeonidis, L.; Karataglis, S. Field study of the effects of excess copper on wheat photosynthesis and productivity. Soil Sci. Plant Nutr. 1997, 43, 531–539. [Google Scholar] [CrossRef]
- Küpper, H.; Šetlík, I.; Spiller, M.; Küpper, F.C.; Prášil, O. Heavy metal-induced inhibition of photosynthesis: Targets of in vivo heavy metal chlorophyll formation. J. Phycol. 2002, 38, 429–441. [Google Scholar] [CrossRef]
- Küpper, H.; Šetlík, I.; Setlikova, E.; Ferimazova, N.; Spiller, M.; Küpper, F.C. Copper-induced inhibition of photosynthesis: Limiting steps of in vivo copper chlorophyll formation in Scmedesmus quadricanda. Funct. Plant Biol. 2003, 30, 1187–1190. [Google Scholar]
- Azab, E.; Hegazy, A.K. Monitoring the Efficiency of Rhazya stricta L. Plants in Phytoremediation of Heavy Met-al-Contaminated Soil. Plants 2020, 9, 1057. [Google Scholar] [CrossRef]
- Baker, A.J.M. Accumulators and excluders -strategies in the response of plants to heavy metals. J. Plant Nutr. 1981, 3, 643–654. [Google Scholar] [CrossRef]
- Ouzounidou, G. Changes in variable chlorophyll fluorescence as a result of Cu treatment: Dose-respons relations in Silene and Thlaspi. Photosynthetica 1993, 29, 455–462. [Google Scholar]
- Ouzounidou, G.; Moustakas, M.; Strasser, R.J. Sites of Action of Copper in the Photosynthetic Apparatus of Maize Leaves: Kinetic Analysis of Chlorophyll Fluorescence, Oxygen Evolution, Absorption Changes and Thermal Dissipation as Monitored by Photoacoustic Signals. Funct. Plant Biol. 1997, 24, 81–90. [Google Scholar] [CrossRef]
- Małkowski, E.; Kita, A.; Galas, W.; Karcz, W.; Kuperberg, J.M. Lead distribution in corn seedlings (Zea mays L.) and its effect on growth and the concentrations of potassium and calcium. Plant Growth Regul. 2002, 37, 69–76. [Google Scholar] [CrossRef]
- Chen, S.; Wang, Q.; Lu, H.; Li, J.; Yang, D.; Liu, J.; Yan, C. Phenolic metabolism and related heavy metal tolerance mechanism in Kandelia Obovata under Cd and Zn stress. Ecotoxicol. Environ. Saf. 2019, 169, 134–143. [Google Scholar] [CrossRef]
- Gajewska, E.; Sklodowska, M.; Slaba, M.; Mazur, J. Effect of nickel on antioxidative enzyme activities, proline and chlorophyll contents in wheat shoots. Biol. Plant. 2006, 50, 653–659. [Google Scholar] [CrossRef]
- Vázquez, M.D.; Poschenrieder, C.; Barceló, J. Chromium VI Induced Structural and Ultrastructural Changes in Bush Bean Plants (Phaseolus vulgaris L.). Ann. Bot. 1987, 59, 427–438. [Google Scholar] [CrossRef]
- Ahmad, M.S.A.; Hussain, M.; Ijaz, M.; Alvi, A.K. Photosynthetic Performance of Two Mung Bean (Vigna radiata) Cultivars under Lead and Copper Stress. Int. J. Agri. Biol. 2008, 10, 167–172. [Google Scholar]
- Moustakas, M.; Lanaras, T.; Symeonidis, L.; Karataglis, S. Growth and some photosynthetic characteristics of field grown Avena sativa under copper and lead stress. Photosynthetica 1994, 30, 389–396. [Google Scholar]
- Burzynski, M. The influence of lead and cadmium on the absorption and distribution of potassium, calcium, magnesium and iron in cucumber seedlings. Acta Physiol. Plant. 1987, 9, 229–239. [Google Scholar]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants, 2nd ed.; CRC Press: Boca Raton, FL, USA, 1992. [Google Scholar]
- Rashid, P.; Mukherii, S. Changes in cyclase and ascorbic oxidase activities in response to lead nitrate treatments in mungbean. Ind. J. Plant Physiol. 1991, 34, 143–146. [Google Scholar]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 1677. [Google Scholar] [CrossRef]
- Kosobrukhov, A.; Knyazeva, I.; Mudrik, V. Plantago major plants responses to increase of content of lead in soil: Growth and photosynthesis. Plant Growth Regul. 2004, 42, 145–151. [Google Scholar] [CrossRef]
- Pätsikkä, E.; Kairavuo, M.; Šeršen, F.; Aro, E.M.; Tyystjärvi, E. Excess copper predisposes photosystem to photoinhi-bition in vivo by outcompeting iron and causing decrease in leaf chlorophyll. Plant Physiol. 2002, 129, 1359–1367. [Google Scholar] [CrossRef] [PubMed]
- Chaneva, G.; Parvanova, P.; Tzvetkova, N.; Uzunova, A. Photosynthetic Response of maize plants against cad-mium and paraquat impact. Water Air Soil Pollut. 2010, 208, 287–293. [Google Scholar] [CrossRef]
- Pandey, N.; Patkak, G.C.; Pandey, D.K.; Pandey, R. Heavy metals, Co, Ni, Cu, Zn and Cd, produce oxidative dam-age and evoke differential antioxidant responses in spinach. Braz. J. Plant Physiol. 2009, 21, 103–111. [Google Scholar] [CrossRef]
- Jubany-Marí, T.; Munné-Bosch, S.; Alegre, L. Redox regulation of water stress responses in field-grown plants. Role of hydrogen peroxide and ascorbate. Plant Physiol. Biochem. 2010, 48, 351–358. [Google Scholar] [CrossRef]
- SinghGill, S.; Narendra, T. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine; Oxford University Press: Oxford, UK, 1999. [Google Scholar]
- Liu, A.; Chen, S.; Chang, R.; Liu, D.; Chen, H.; Ahammed, G.J.; Lin, X.; He, C. Arbuscular mycorrhizae improve low temperature tolerance in cucumber via alterations in H2O2 accumulation and ATPase activity. J. Plant Res. 2014, 127, 775–785. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Q.; He, M.; Cao, T.; Yusuyin, Y.; Han, W. Antioxidative responses related to H2O2 depletion in Hypnum plumaeforma under the combined stress induced by Pb and Ni. Environ. Monit. Assess. 2010, 163, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Eapen, S.; D’Souza, S. Cadmium accumulation and its influence on lipid peroxidation and antioxidative system in an aquatic plant, Bacopa monnieri L. Chemosphere 2006, 62, 233–246. [Google Scholar] [CrossRef]
- Keser, G.; Saygideger, S. Effects of Lead on the Activities of Antioxidant Enzymes in Watercress, Nasturtium officinale R. Br. Biol. Trace Elem. Res. 2009, 137, 235–243. [Google Scholar] [CrossRef]
- Zancheta, A.F.; De Abreu, C.A.; Zambrosi, F.B.; Erismann, N.M.; Lagôa, M.A. Copper phytoextraction by different plant species grown in nutrient solution. Bragantia 2011, 70, 737–744. [Google Scholar] [CrossRef]
- Vidal, C.; Ruiz, A.; Ortiz, J.; Larama, G.; Perez, R.; Santander, C.; Ferreira, P.A.A.; Cornejo, P. Antioxidant Responses of Phenolic Compounds and Immobilization of Copper in Imperata Cylindrica, a Plant with Potential Use for Bioremediation of Cu Contaminated Environments. Plants 2020, 9, 1397. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, D.R.; Arnon, D.I. The Water-culture Method for Growing Plants without Soil. Californian Agricultural Experimental Station. Circular No. 347; University of California: Berkeley, CA, USA, 1950. [Google Scholar]
- Wintermans, J.F.; De Mots, A. Spectrophotometric characteristics of chlorophyll ‘a’ and ‘b’ and their pheophytins in ethanol. Biochim. Biophys. Acta 1965, 109, 448–453. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Academic Press: Orlando, FL, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Yang, C.M.; Chang, K.M.; Yin, M.H.; Hung, H.M. Methods for the determination of the chlorophylls and their deriva-tives. Taiwania 1998, 43, 116–122. [Google Scholar]
- Giannakoula, A.; Ouzounidou, G.; Ilias, I.; Bunnell, T.B. Application of plant growth regulators in lentils for salinity stress alleviation. J. Environ. Prot. Ecol. 2015, 16, 567–576. [Google Scholar]
- Von Caemmerer, S.; Farquhar, G.D. Some relationships between the biochemistry of photosynthesis and the gas exchange of leaves. Planta 1981, 153, 376–387. [Google Scholar] [CrossRef]
- Giannakoula, A.; Ilias, Ι. The effect of water stress and salinity on growth and physiology of tomato (Lycopersicon esculentum Mil.). Arch. Biol. Sci. (Belgrade) 2013, 65, 611–620. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Alexieva, V.; Sergiev, I.; Mapelli, S.; Karanov, E. The effect of drought and ultraviolet radiation on growth and stress markers in pea and wheat. Plant Cell Environ. 2001, 24, 1337–1344. [Google Scholar] [CrossRef]
- Kim, D.-O.; Jeong, S.W.; Lee, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Scalbert, A.; Monties, B.; Janin, G. Tannins in wood: Comparison of different estimation methods. J. Agric. Food Chem. 1989, 37, 1324–1329. [Google Scholar] [CrossRef]


{kind=link}
| Treatments | Concentration μM | Plant Height cm | H2O2 mol/g FW | MDA nmol/g FW | Flavonoids g CE./100 g DW |
|---|---|---|---|---|---|
| Control | 0 | 50 c | 10.31 a | 25.4 a | 128.4 c |
| Pb | 500 μM | 58.0 d | 10.23 a | 23.3 a | 151.6 d |
| Pb | 800 μM | 49.1 c | 16.93 b | 29.5 b | 115.1 b |
| Cu | 500 μM | 52.2 c | 15.41 b | 27.3 b | 178.8 e |
| Cu | 800 μM | 40.1 b | 18.23 b | 33.3 a | 101.7 a |
| Cu + Pb | 500 μM | 42.1 b | 17.92 c | 30.5 b | 165.1 e |
| Cu + Pb | 800 μM | 31.2 a | 20.41 c | 37.3 c | 98.8 a |
| Treatments | Concentration μM | Chl(a + b) mg g−1 DW | Chla mg g−1 DW | Carotenoids mg g−1 FW | Phenolics GAE mg/100 g FW |
|---|---|---|---|---|---|
| Control | 0 | 11.7 c | 7.60 c | 8.30 c | 223.4 c |
| Pb | 500 μM | 12.6 d | 8.84 d | 10.3 d | 281.2 d |
| Pb | 800 μM | 10.1 b | 6.75 b | 7.2 b | 187.1 b |
| Cu | 500 μM | 13.2 d | 9.85 d | 10.9 d | 284.5 d |
| Cu | 800 μM | 8.6 a | 6.02 a | 6.3 a | 171.2 a |
| Cu + Pb | 500 μM | 12.9 d | 8.71 d | 7.2 b | 287.1 d |
| Cu + Pb | 800 μM | 7.2 a | 5.63 a | 6.1 a | 164.5 a |
| Treatments | Concentration μM | Fo | Fm | Fv/Fm | Fv/Fo |
|---|---|---|---|---|---|
| Control | 0 | 437 c | 2200 c | 0.81 a | 4.0 a |
| Pb | 500 μM | 430 d | 2538 b | 0.83 a | 4.9 a |
| Pb | 800 μM | 481 c | 2012 c | 0.78 b | 3.6 b |
| Cu | 500 μM | 379 d | 2409 bc | 0.85 a | 4.2 a |
| Cu | 800 μM | 1006 a | 3383 a | 0.64 d | 2.4 c |
| Cu + Pb | 500 μM | 509 d | 1912 d | 0.74 c | 2.7 c |
| Cu + Pb | 800 μM | 1107 a | 3651 a | 0.61 d | 2.2 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giannakoula, A.; Therios, I.; Chatzissavvidis, C. Effect of Lead and Copper on Photosynthetic Apparatus in Citrus (Citrus aurantium L.) Plants. The Role of Antioxidants in Oxidative Damage as a Response to Heavy Metal Stress. Plants 2021, 10, 155. https://doi.org/10.3390/plants10010155
Giannakoula A, Therios I, Chatzissavvidis C. Effect of Lead and Copper on Photosynthetic Apparatus in Citrus (Citrus aurantium L.) Plants. The Role of Antioxidants in Oxidative Damage as a Response to Heavy Metal Stress. Plants. 2021; 10(1):155. https://doi.org/10.3390/plants10010155
Chicago/Turabian StyleGiannakoula, Anastasia, Ioannis Therios, and Christos Chatzissavvidis. 2021. "Effect of Lead and Copper on Photosynthetic Apparatus in Citrus (Citrus aurantium L.) Plants. The Role of Antioxidants in Oxidative Damage as a Response to Heavy Metal Stress" Plants 10, no. 1: 155. https://doi.org/10.3390/plants10010155
APA StyleGiannakoula, A., Therios, I., & Chatzissavvidis, C. (2021). Effect of Lead and Copper on Photosynthetic Apparatus in Citrus (Citrus aurantium L.) Plants. The Role of Antioxidants in Oxidative Damage as a Response to Heavy Metal Stress. Plants, 10(1), 155. https://doi.org/10.3390/plants10010155