Molecular Characterization and Expression of Four Aquaporin Genes in Impatiens walleriana during Drought Stress and Recovery

 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
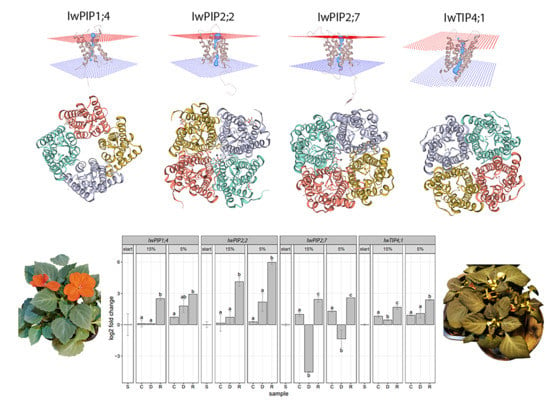
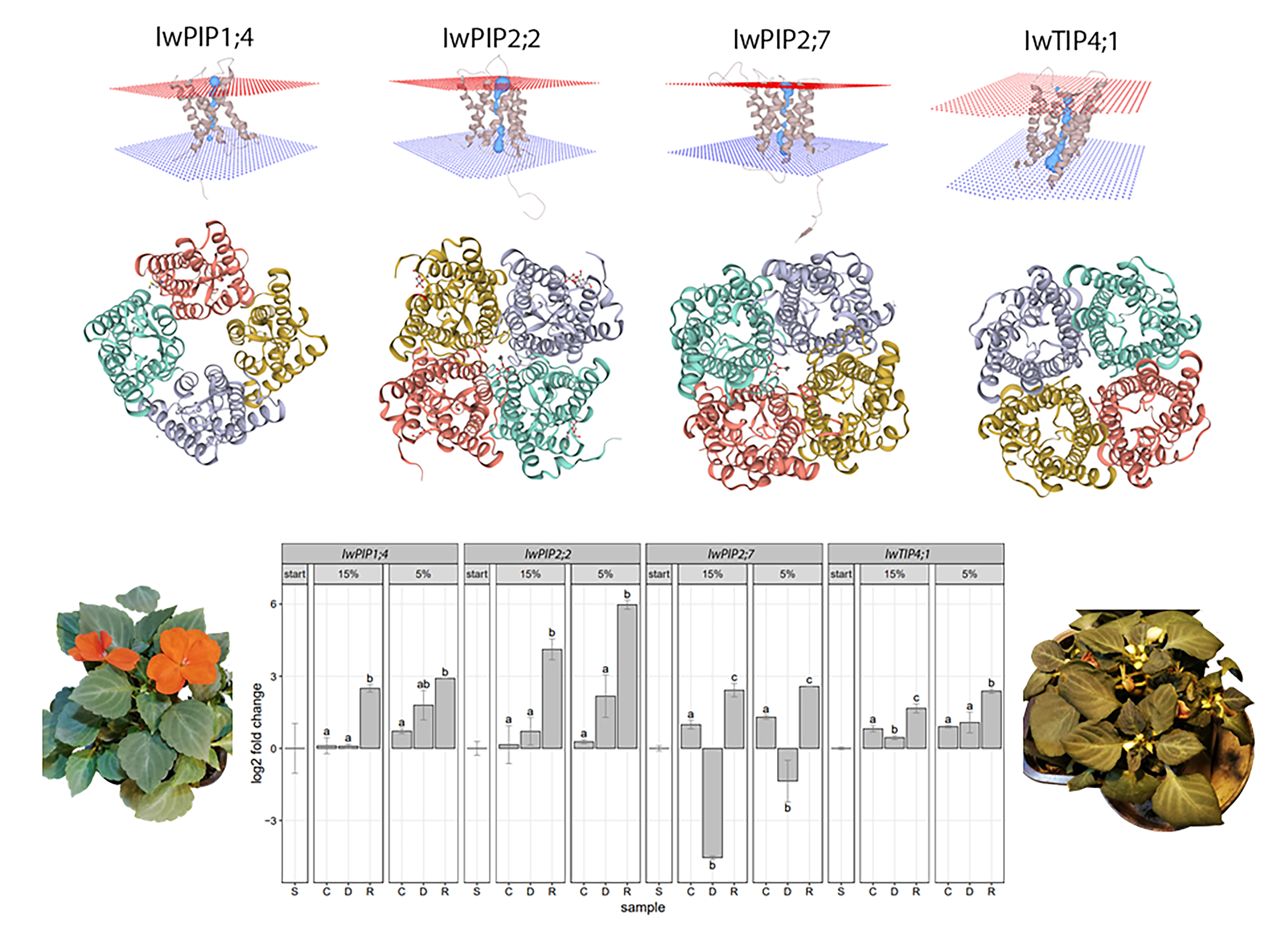
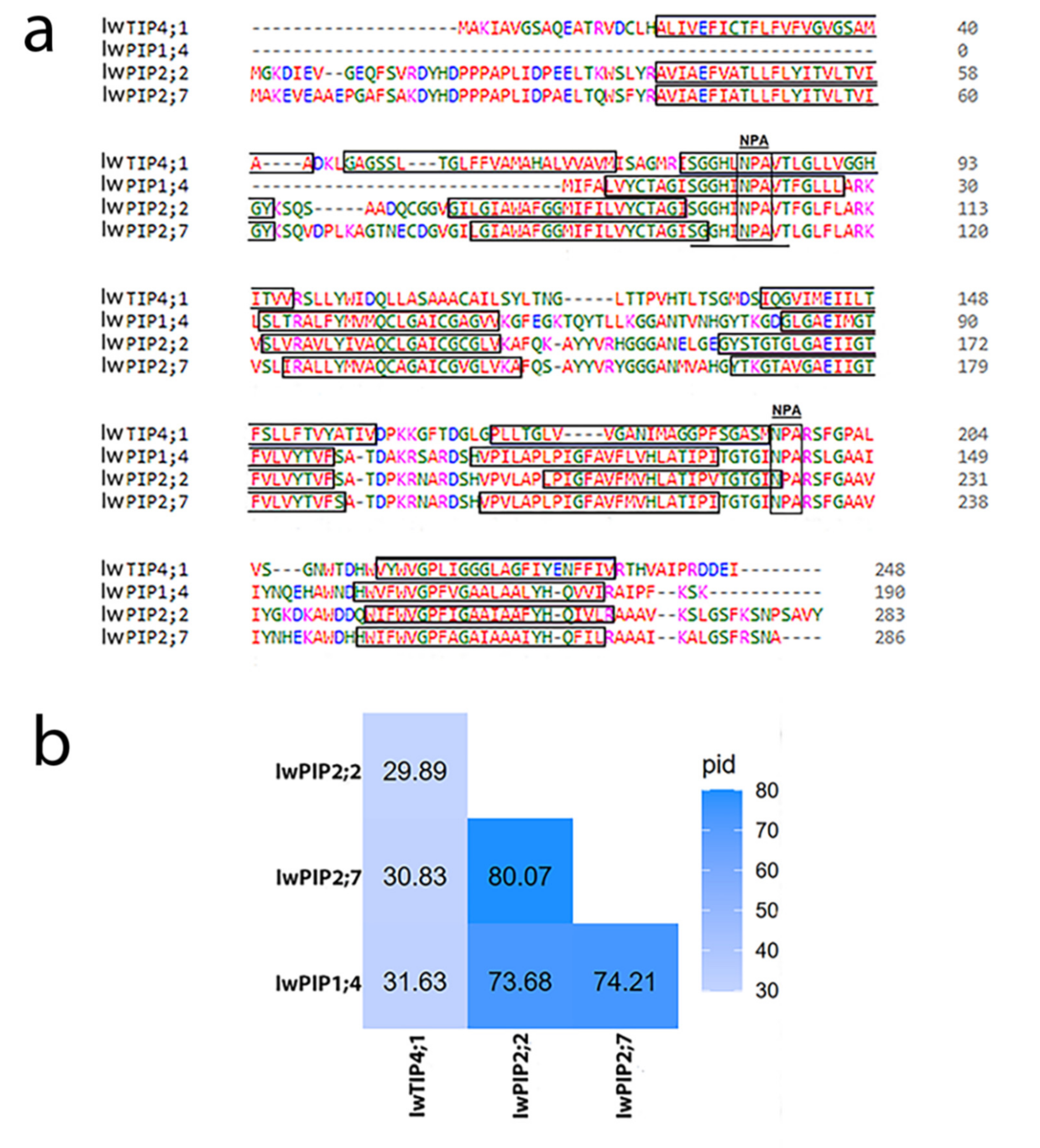
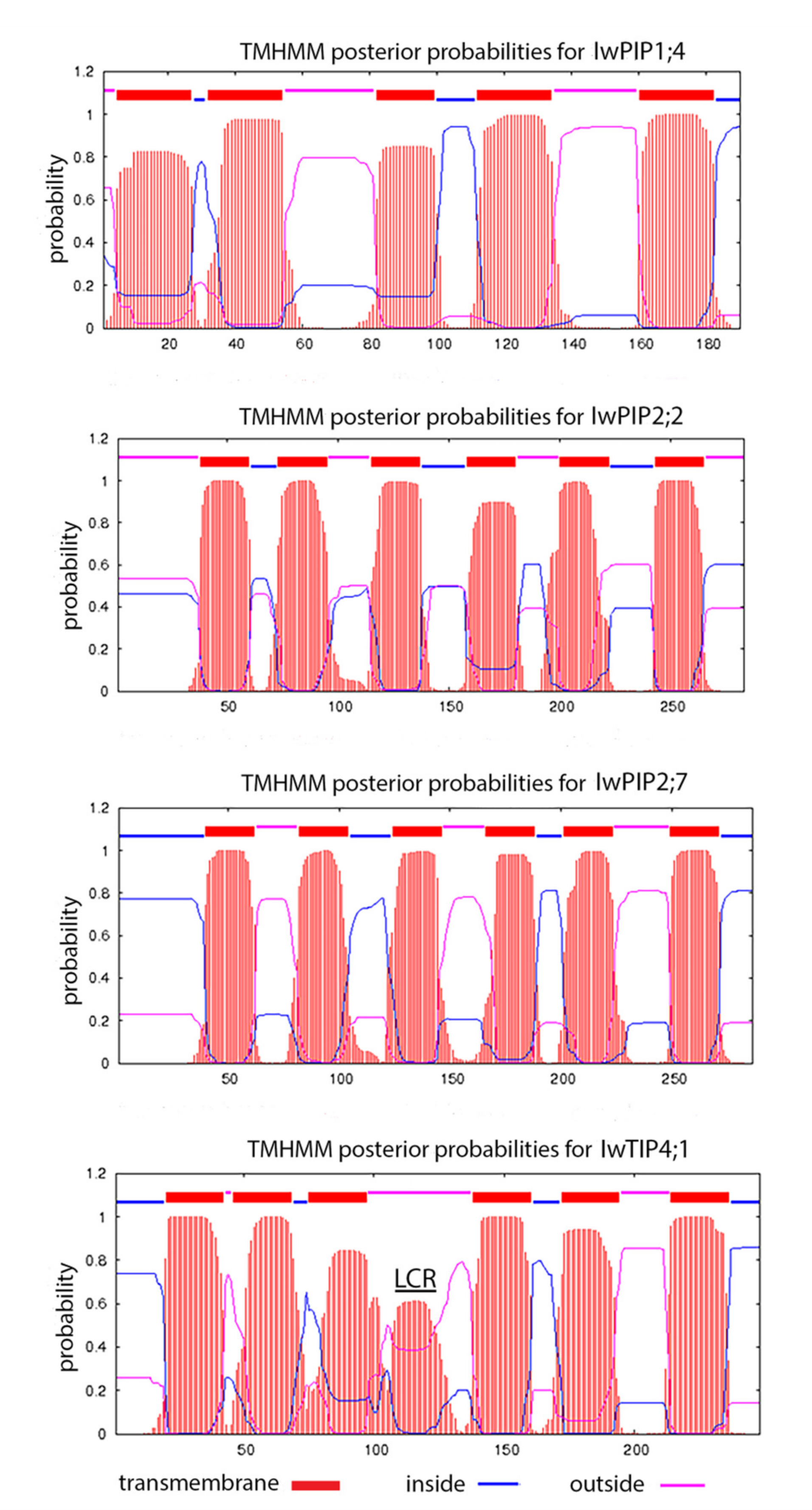
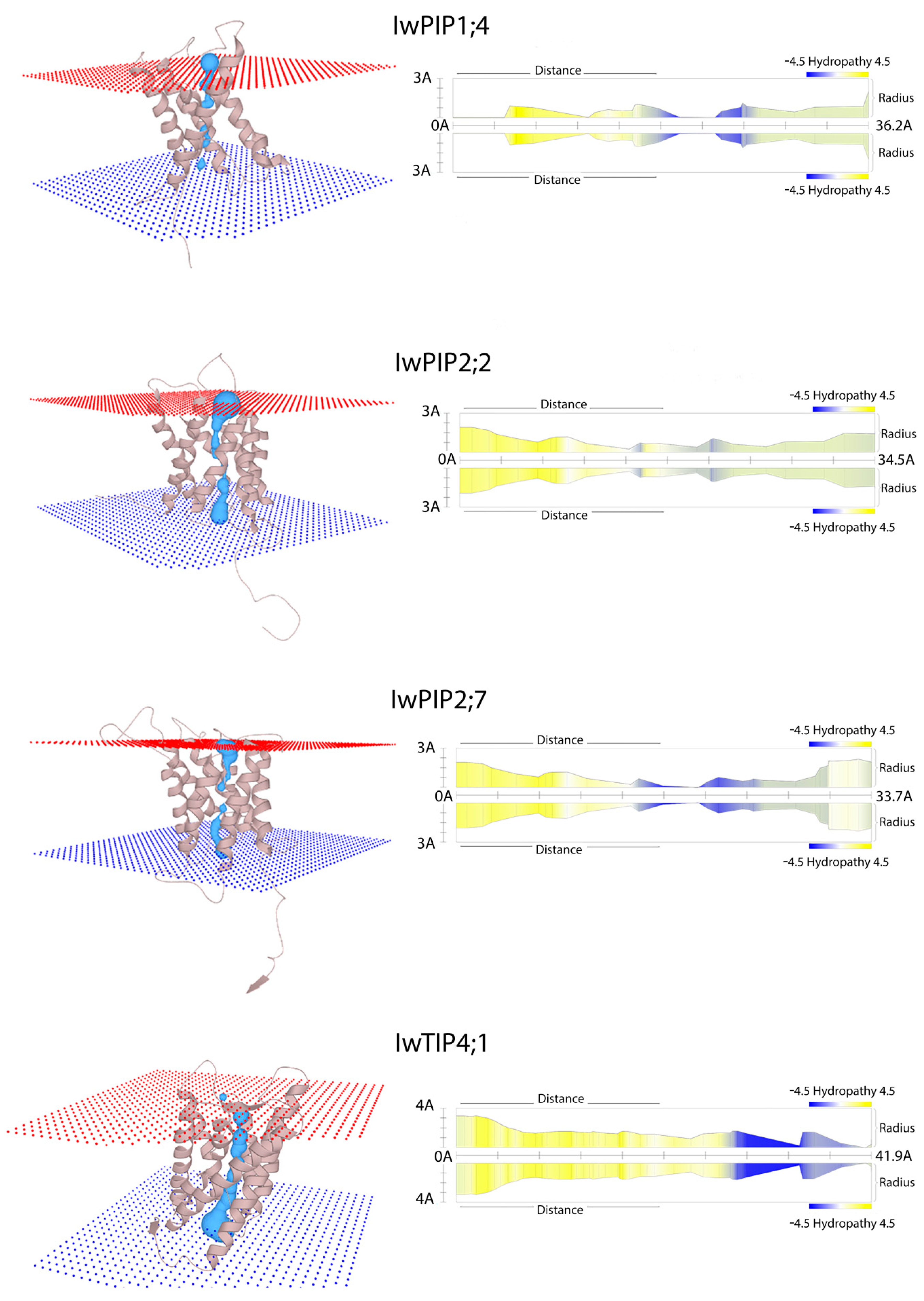
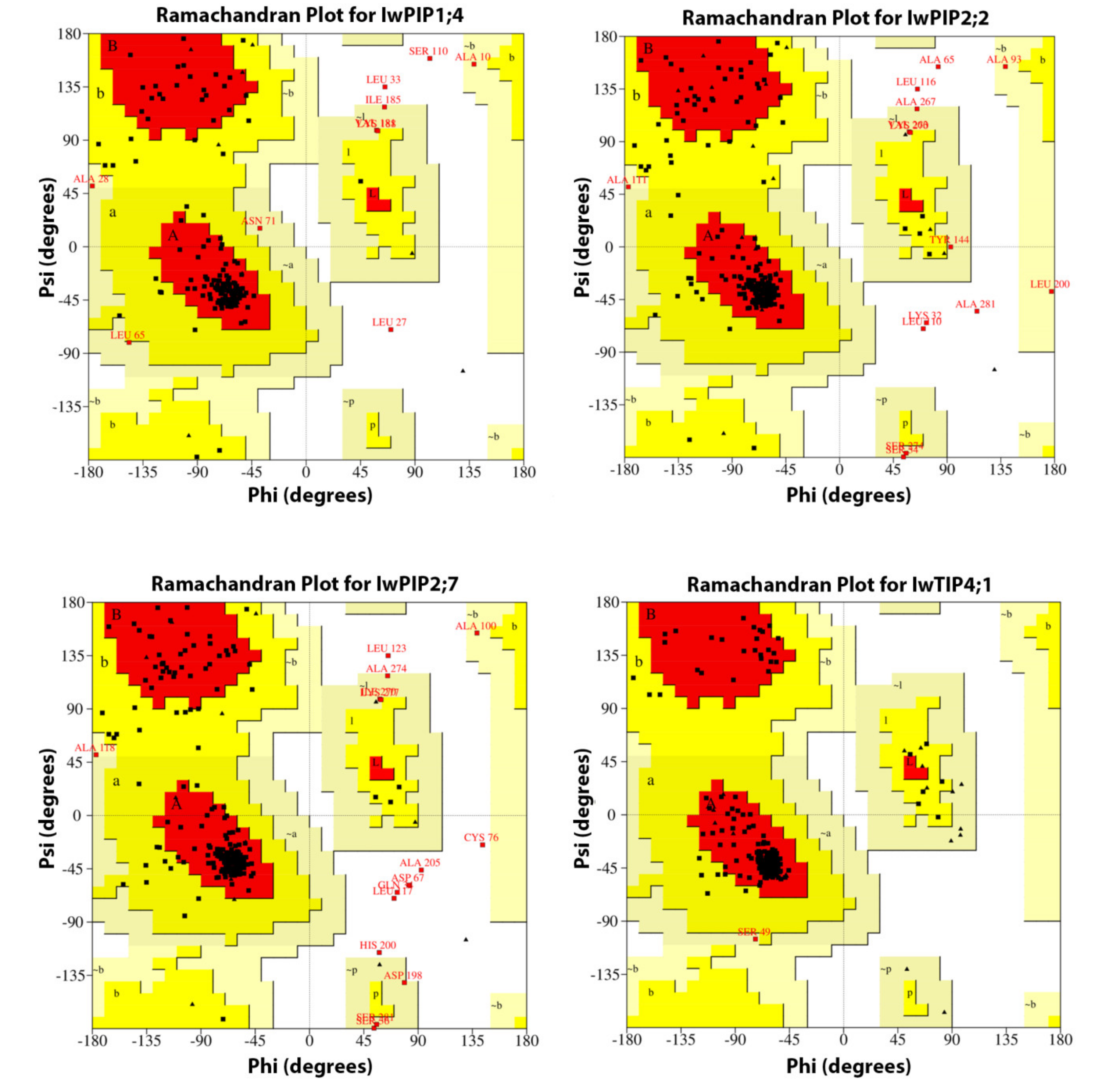
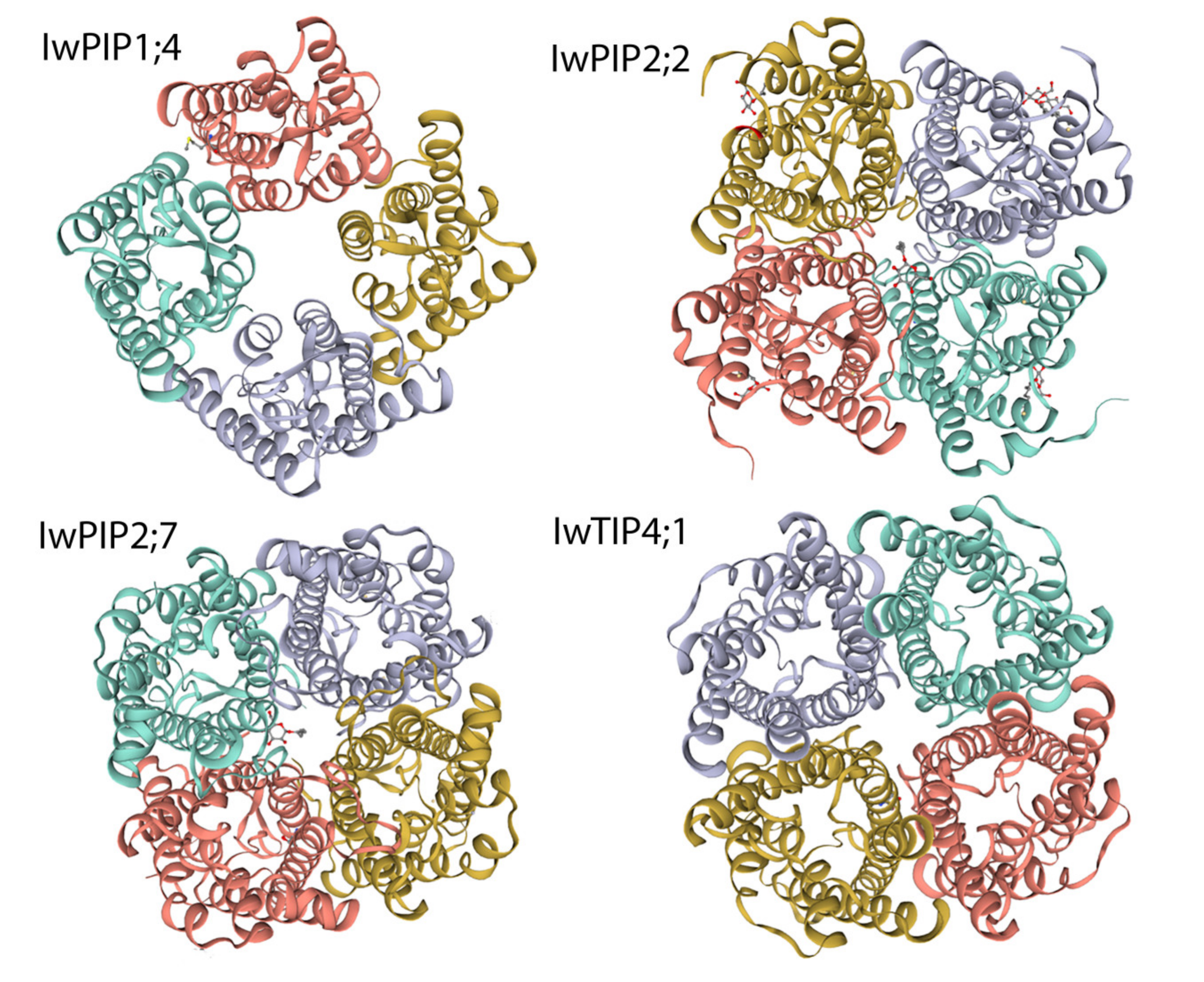
2.1. Characteristic of I. walleriana Aquaporin Proteins and Their Predicted Structural Models
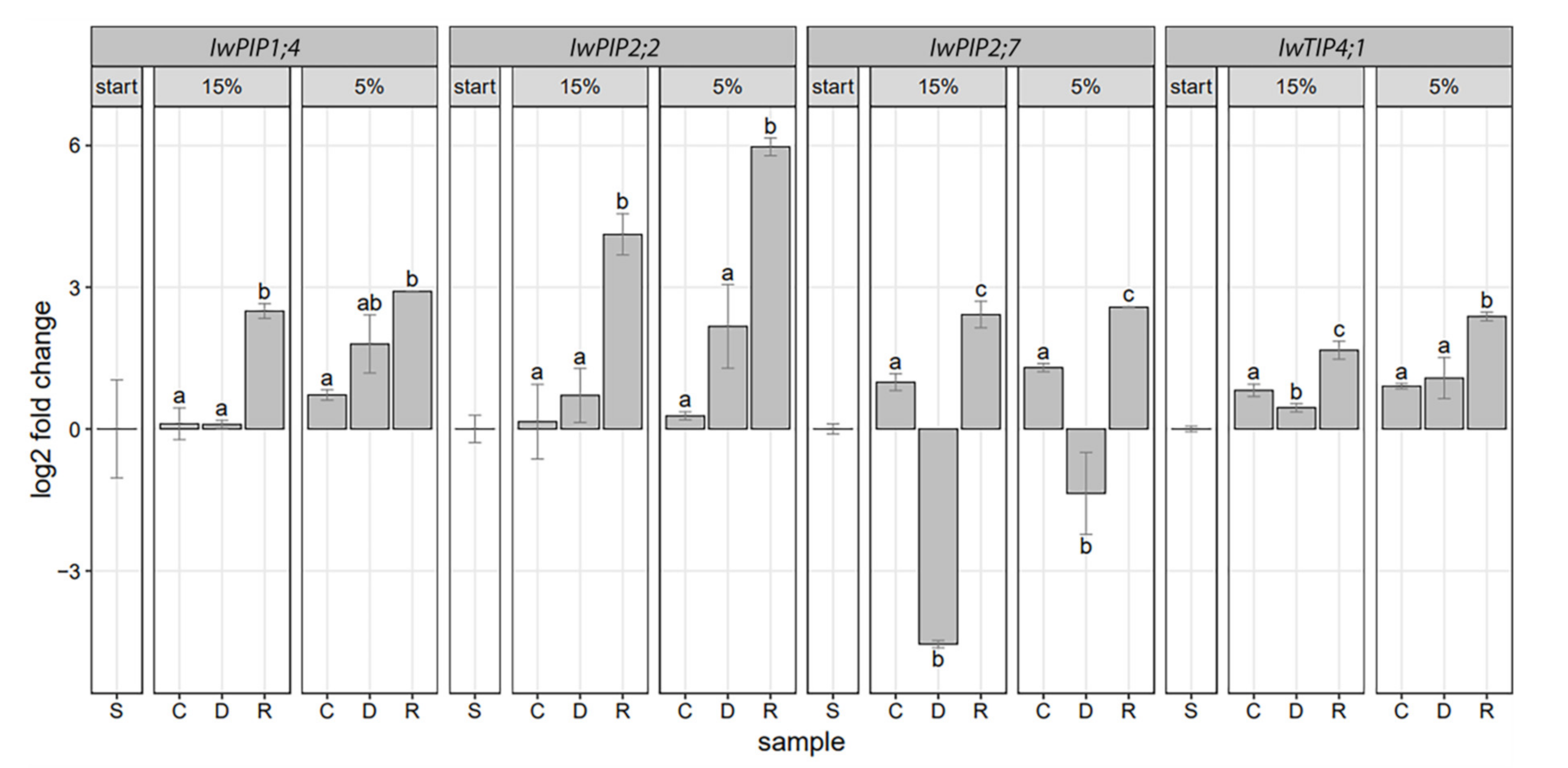
2.2. Aquaporins Expression in Drought-Stressed and Recovered I. walleriana
3. Discussion
4. Materials and Methods
4.1. Aquaporins Sequences Analysis
4.2. Experiment Design, RNA Isolation and Reverse Transcription PCR (RT-PCR)
4.3. Quantitative Real-Time PCR (qRT-PCR)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants; Springer: Cham, Switzerland, 2016; Volume 1, pp. 1–16. [Google Scholar]
- Waseem, M.; Ali, A.; Tahir, M.; Nadeem, M.A.; Ayub, M.; Tanveer, A.; Ahmad, R.; Hussain, M. Mechanism of drought tolerance in plant and its management through different methods. Cont. J. Agric. Sci. 2011, 5, 10–25. [Google Scholar]
- Ðurić, M.; Subotić, A.; Prokić, L.; Trifunović-Momčilov, M.; Cingel, A.; Vujičić, M.; Milošević, S. Morpho-Physiological and Molecular Evaluation of Drought and Recovery in Impatiens walleriana Grown Ex Vitro. Plants 2020, 9, 1559. [Google Scholar] [CrossRef]
- Afzal, Z.; Howton, T.; Sun, Y.; Mukhtar, M. The roles of aquaporins in plant stress responses. J. Dev. Biol. 2016, 4, 9. [Google Scholar] [CrossRef]
- Abascal, F.; Irisarri, I.; Zardoya, R. Diversity and evolution of membrane intrinsic proteins. BBA Gen. Subj. 2014, 1840, 1468–1481. [Google Scholar] [CrossRef]
- Maurel, C.; Boursiac, Y.; Luu, D.T.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Kapilan, R.; Vaziri, M.; Zwiazek, J.J. Regulation of aquaporins in plants under stress. Biol. Res. 2018, 51, 4. [Google Scholar]
- Pawłowicz, I.; Masajada, K. Aquaporins as a link between water relations and photosynthetic pathway in abiotic stress tolerance in plants. Gene 2019, 687, 166–172. [Google Scholar] [CrossRef]
- Hove, R.M.; Bhave, M. Plant aquaporins with non-aqua functions: Deciphering the signature sequences. Plant Mol. Biol. 2011, 75, 413–430. [Google Scholar]
- Yaneff, A.; Vitali, V.; Amodeo, G. PIP1 aquaporins: Intrinsic water channels or PIP2 aquaporin modulators? FEBS Lett. 2015, 589, 3508–3515. [Google Scholar] [CrossRef]
- Fitzpatrick, K.L.; Reid, R.J. The involvement of aquaglyceroporins in transport of boron in barley roots. Plant Cell Environ. 2009, 32, 1357–1365. [Google Scholar]
- Vajpai, M.; Mukherjee, M.; Sankararamakrishnan, R. Cooperativity in plant plasma membrane intrinsic proteins (PIPs): Mechanism of increased water transport in maize PIP1 channels in hetero-tetramers. Sci. Rep. 2018, 8, 1–17. [Google Scholar]
- Bienert, M.D.; Diehn, T.A.; Richet, N.; Chaumont, F.; Bienert, G.P. Heterotetramerization of plant PIP1 and PIP2 aquaporins is an evolutionary ancient feature to guide PIP1 plasma membrane localization and function. Front. Plant Sci. 2018, 9, 382. [Google Scholar] [CrossRef]
- Maurel, C. Plant aquaporins: Novel functions and regulation properties. FEBS Lett. 2007, 581, 2227–2236. [Google Scholar] [CrossRef]
- Kurowska, M.M. TIP Aquaporins in Plants: Role in Abiotic Stress Tolerance. In Abiotic Stress in Plants; IntechOpen: London, UK, 2020. [Google Scholar]
- Gerbeau, P.; Güçlü, J.; Ripoche, P.; Maurel, C. Aquaporin Nt-TIPa can account for the high permeability of tobacco cell vacuolar membrane to small neutral solutes. Plant J. 1999, 18, 577–587. [Google Scholar] [CrossRef]
- Jahn, T.P.; Møller, A.L.; Zeuthen, T.; Holm, L.M.; Klærke, D.A.; Mohsin, B.; Kühlbrandt, W.; Schjoerring, J.K. Aquaporin homologues in plants and mammals transport ammonia. FEBS Lett. 2004, 574, 31–36. [Google Scholar]
- Srivastava, A.K.; Penna, S.; Nguyen, D.V.; Tran, L.S.P. Multifaceted roles of aquaporins as molecular conduits in plant responses to abiotic stresses. Crit. Rev. Biotechnol. 2016, 36, 389–398. [Google Scholar] [CrossRef]
- Zargar, S.M.; Nagar, P.; Deshmukh, R.; Nazir, M.; Wani, A.A.; Masoodi, K.Z.; Agrawal, G.K.; Rakwal, R. Aquaporins as potential drought tolerance inducing proteins: Towards instigating stress tolerance. J. Proteom. 2017, 169, 233–238. [Google Scholar] [CrossRef]
- Jia, J.; Liang, Y.; Gou, T.; Hu, Y.; Zhu, Y.; Huo, H.; Guo, J.; Gong, H. The expression response of plasma membrane aquaporins to salt stress in tomato plants. Environ. Exp. Bot. 2020, 178, 104190. [Google Scholar] [CrossRef]
- Janssens, S.B.; Knox, E.B.; Huysmans, S.; Smets, E.F.; Merckx, V.S. Rapid radiation of Impatiens (Balsaminaceae) during Pliocene and Pleistocene: Result of a global climate change. Mol. Phylogenet. Evol. 2009, 52, 806–824. [Google Scholar]
- Fischer, E. Balsaminaceae. In Flowering Plants Dicotyledons; Springer: Berlin/Heidelberg, Germany, 2004; pp. 20–25. [Google Scholar]
- Grey-Wilson, C. Hydrocera triflora, Its Floral Morphology and Relationship with Impatiens: Studies in Balsaminaceae: V. Kew Bull. 1980, 35, 213–219. [Google Scholar]
- Chaumont, F.; Barrieu, F.; Jung, R.; Chrispeels, M.J. Plasma membrane intrinsic proteins from maize cluster in two sequence subgroups with differential aquaporin activity. Plant Physiol. 2000, 122, 1025–1034. [Google Scholar] [CrossRef]
- Dong, Y.; Yang, C.; Zhang, D.; Wang, Y. Cloning and sequence analysis of gene encoding plasma aquaporin of Tamarix albiflonum. Front. For. China 2007, 2, 217–221. [Google Scholar]
- Yue, C.; Cao, H.; Wang, L.; Zhou, Y.; Hao, X.; Zeng, J.; Wang, X.; Yang, Y. Molecular cloning and expression analysis of tea plant aquaporin (AQUAPORIN) gene family. Plant Physiol. Biochem. 2014, 83, 65–76. [Google Scholar] [CrossRef]
- Sun, X.; Deng, Y.; Liang, L.; Jia, X.; Xiao, Z.; Su, J. Overexpression of a PIP1 gene from Salicornia bigelovii in tobacco plants improves their drought tolerance. J. Am. Soc. Hortic. Sci. 2017, 142, 235–245. [Google Scholar] [CrossRef]
- Kumar, N.; Kumawat, S.; Khatri, P.; Singla, P.; Tandon, G.; Bhatt, V.; Shinde, S.; Patil, G.B.; Sonah, H.; Deshmukh, R. Understanding aquaporin transport system in highly stress-tolerant and medicinal plant species Jujube (Ziziphus jujuba Mill.). J. Biotechnol. 2020, 324, 103–111. [Google Scholar] [CrossRef]
- Kumar, M.; Kumari, B.; Kumar, R. Structural Analysis of Low Complexity Regions of Proteins. Can J. Biotech. 2017, 1, 219. [Google Scholar]
- Toll-Riera, M.; Radó-Trilla, N.; Martys, F.; Alba, M.M. Role of low-complexity sequences in the formation of novel protein coding sequences. Mol. Biol. Evol. 2012, 29, 883–886. [Google Scholar] [CrossRef]
- Kumari, B.; Kumar, R.; Chauhan, V.; Kumar, M. Comparative functional analysis of proteins containing low-complexity predicted amyloid regions. PeerJ 2018, 6, e5823. [Google Scholar]
- Michelitsch, M.D.; Weissman, J.S. A census of glutamine/asparagine-rich regions: Implications for their conserved function and the prediction of novel prions. Proc. Natl. Acad. Sci. USA 2000, 97, 11910–11915. [Google Scholar] [CrossRef]
- Gunawardena, S.; Goldstein, L.S. Polyglutamine diseases and transport problems: Deadly traffic jams on neuronal highways. Arch. Neurol. 2005, 62, 46–51. [Google Scholar]
- Hollingsworth, S.A.; Karplus, P.A. A fresh look at the Ramachandran plot and the occurrence of standard structures in proteins. Biomol. Concepts 2010, 1, 271–283. [Google Scholar] [CrossRef]
- Carugo, O.; Djinović-Carugo, K. A proteomic Ramachandran plot (PRplot). Amino Acids 2013, 44, 781–790. [Google Scholar]
- Laskowski, R.A.; Furnham, N.; Thornton, J.M. The Ramachandran plot and protein structure validation. In Biomolecular Forms and Functions: A Celebration of 50 Years of the Ramachandran Map; World Scientific: Singapore, 2013; pp. 62–75. [Google Scholar] [CrossRef]
- Srivastava, M.; Gupta, S.K.; Abhilash, P.C.; Singh, N. Structure prediction and binding sites analysis of curcin protein of Jatropha curcas using computational approaches. J. Mol. Model. 2012, 18, 2971–2979. [Google Scholar]
- Donde, R.; Gupta, M.K.; Gouda, G.; Kumar, J.; Vadde, R.; Sahoo, K.K.; Dash, S.K.; Behera, L. Computational characterization of structural and functional roles of DREB1A, DREB1B and DREB1C in enhancing cold tolerance in rice plant. Amino Acids 2019, 51, 839–853. [Google Scholar]
- Preeti, A.; Shaifali, S.; Kumar, P.G.; Mani, P.D. Computational characterization of lipoxygenase and hydroperoxide lyase enzymes and Real Time PCR-based expression analysis of their encoding genes in peanut under heat and drought stress. Res. J. Biotechnol. Vol. 2019, 14, 12. [Google Scholar]
- Mubassir, M.H.M.; Naser, M.A.; Abdul-Wahab, M.F.; Jawad, T.; Alvy, R.I.; Hamdan, S. Comprehensive in silico modeling of the rice plant PRR Xa21 and its interaction with RaxX21-sY and OsSERK2. RSC Adv. 2020, 10, 15800–15814. [Google Scholar]
- Gouda, G.; Gupta, M.K.; Donde, R.; Kumar, J.; Vadde, R.; Mohapatra, T.; Behera, L. Computational approach towards understanding structural and functional role of cytokinin oxidase/dehydrogenase 2 (CKX2) in enhancing grain yield in rice plant. J. Biomol. Struct. Dyn. 2020, 38, 1158–1167. [Google Scholar]
- Kumar, A.; Kumar, S.; Kumar, A.; Sharma, N.; Sharma, M.; Singh, K.P.; Rathore, M.; Gajula, M.P. Homology modeling, molecular docking and molecular dynamics based functional insights into rice urease bound to urea. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1539–1548. [Google Scholar]
- Dubey, K.; Goswami, S.; Kumar, N.; Kumar, R.R.; Niraj, R.R.K. Cloning and in silico characterization of Heat shock factor (Hsf2) from Wheat (Triticum aestivum L.). bioRxiv 2020. Available online: https://www.biorxiv.org/content/10.1101/2020.04.17.046094v1.full (accessed on 3 December 2020). [CrossRef]
- Kumar, A.; Kumar, S.; Kumar, U.; Suravajhala, P.; Gajula, M.P. Functional and structural insights into novel DREB1A transcription factors in common wheat (Triticum aestivum L.): A molecular modeling approach. Comput. Biol. Chem. 2016, 64, 217–226. [Google Scholar]
- Kyte, J.; Doolittle, R.F. A simple method for displaying the hydropathic character of a protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef]
- Pou, A.; Jeanguenin, L.; Milhiet, T.; Batoko, H.; Chaumont, F.; Hachez, C. Salinity-mediated transcriptional and post-translational regulation of the Arabidopsis aquaporin PIP2; 7. Plant Mol. Biol. 2016, 92, 731–744. [Google Scholar]
- Antonić, D.; Milošević, S.; Cingel, A.; Lojić, M.; Trifunović-Momčilov, M.; Petrić, M.; Simonović, A. Effects of exogenous salicylic acid on Impatiens walleriana L. grown in vitro under polyethylene glycol-imposed drought. S. Afr. J. Bot. 2016, 105, 226–233. [Google Scholar]
- Antonić, D.D.; Subotić, A.R.; Dragićević, M.B.; Pantelić, D.; Milošević, S.M.; Simonović, A.D.; Momčilović, I. Effects of Exogenous Salicylic Acid on Drought Response and Characterization of Dehydrins in Impatiens walleriana. Plants 2020, 9, 1589. [Google Scholar]
- Shekoofa, A.; Sinclair, T.R. Aquaporin activity to improve crop drought tolerance. Cells 2018, 7, 123. [Google Scholar] [CrossRef]
- Avila, R.T.; Cardoso, A.A.; de Almeida, W.L.; Costa, L.C.; Machado, K.L.; Barbosa, M.L.; de Souza, R.P.; Oliveira, L.A.; Batista, D.S.; Martins, S.C.; et al. Coffee plants respond to drought and elevated [CO2] through changes in stomatal function, plant hydraulic conductance, and aquaporin expression. Environ. Exp. Bot. 2020, 177, 104148. [Google Scholar] [CrossRef]
- Iwuala, E.; Odjegba, V.; Sharma, V.; Alam, A. Drought stress modulates expression of aquaporin gene and photosynthetic efficiency in Pennisetum glaucum (L.) R. Br. genotypes. Curr. Plant Biol. 2020, 21, 100131. [Google Scholar] [CrossRef]
- Boursiac, Y.; Chen, S.; Luu, D.T.; Sorieul, M.; van den Dries, N.; Maurel, C. Early effects of salinity on water transport in Arabidopsis roots. Molecular and cellular features of aquaporin expression. Plant Physiol. 2005, 139, 790–805. [Google Scholar] [CrossRef]
- Alexandersson, E.; Fraysse, L.; Sjövall-Larsen, S.; Gustavsson, S.; Fellert, M.; Karlsson, M.; Johanson, U.; Kjellbom, P. Whole gene family expression and drought stress regulation of aquaporins. Plant Mol. Biol. 2005, 59, 469–484. [Google Scholar]
- Li, R.; Wang, J.; Li, S.; Zhang, L.; Qi, C.; Weeda, S.; Zhao, B.; Ren, S.; Guo, Y.D. Plasma membrane intrinsic proteins SlPIP2; 1, SlPIP2; 7 and SlPIP2; 5 conferring enhanced drought stress tolerance in tomato. Sci. Rep. 2016, 6, 31814. [Google Scholar]
- Zupin, M.; Sedlar, A.; Kidrič, M.; Meglič, V. Drought-induced expression of aquaporin genes in leaves of two common bean cultivars differing in tolerance to drought stress. J. Plant Res. 2017, 130, 735–745. [Google Scholar]
- Hu, W.; Ding, Z.; Tie, W.; Yan, Y.; Liu, Y.; Wu, C.; Liu, J.; Wang, J.; Peng, M.; Xu, B.; et al. Comparative physiological and transcriptomic analyses provide integrated insight into osmotic, cold, and salt stress tolerance mechanisms in banana. Sci. Rep. 2017, 7, 43007. [Google Scholar]
- Xu, Y.; Hu, W.; Liu, J.; Song, S.; Hou, X.; Jia, C.; Li, J.; Miao, H.; Wang, Z.; Tie, W.; et al. An aquaporin gene MaPIP2-7 is involved in tolerance to drought, cold and salt stresses in transgenic banana (Musa acuminata L.). Plant Physiol. Biochem. 2020, 147, 66–76. [Google Scholar] [CrossRef]
- Santos, A.B.D.; Mazzafera, P. Aquaporins and the control of the water status in coffee plants. Theor. Exp. Plant Physiol. 2013, 25, 79–93. [Google Scholar]
- Javot, H.; Lauvergeat, V.; Santoni, V.; Martin-Laurent, F.; Güçlü, J.; Vinh, J.; Heyes, J.; Franck, K.I.; Schäffner, A.R.; Bouchez, D.; et al. Role of a single aquaporin isoform in root water uptake. Plant Cell 2003, 15, 509–522. [Google Scholar]
- Jang, J.Y.; Kim, D.G.; Kim, Y.O.; Kim, J.S.; Kang, H. An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana. Plant Mol. Biol. 2004, 54, 713–725. [Google Scholar]
- Paudel, I.; Gerbi, H.; Zisovich, A.; Sapir, G.; Ben-Dor, S.; Brumfeld, V.; Klein, T. Drought tolerance mechanisms and aquaporin expression of wild vs. cultivated pear tree species in the field. Environ. Exp. Bot. 2019, 167, 103832. [Google Scholar]
- Kurowska, M.M.; Wiecha, K.; Gajek, K.; Szarejko, I. Drought stress and re-watering affect the abundance of TIP aquaporin transcripts in barley. PLoS ONE 2019, 14, e0226423. [Google Scholar]
- Miniussi, M.; Del Terra, L.; Savi, T.; Pallavicini, A.; Nardini, A. Aquaporins in Coffea arabica L.: Identification, expression, and impacts on plant water relations and hydraulics. Plant Physiol. Biochem. 2015, 95, 92–102. [Google Scholar]
- Vandeleur, R.K.; Mayo, G.; Shelden, M.C.; Gilliham, M.; Kaiser, B.N.; Tyerman, S.D. The role of plasma membrane intrinsic protein aquaporins in water transport through roots: Diurnal and drought stress responses reveal different strategies between isohydric and anisohydric cultivars of grapevine. Plant Physiol. 2009, 149, 445–460. [Google Scholar] [CrossRef]
- Yooyongwech, S.; Cha-um, S.; Supaibulwatana, K. Water relation and aquaporin genes (PIP1; 2 and PIP2; 1) expression at the reproductive stage of rice (Oryza sativa L. spp. indica) mutant subjected to water deficit stress. Plant Omics 2013, 6, 79. [Google Scholar]
- Hasan, S.A.; Rabei, S.H.; Nada, R.M.; Abogadallah, G.M. Water use efficiency in the drought-stressed sorghum and maize in relation to expression of aquaporin genes. Biol. Plant. 2017, 61, 127–137. [Google Scholar]
- Reddy, K.S.; Sekhar, K.M.; Reddy, A.R. Genotypic variation in tolerance to drought stress is highly coordinated with hydraulic conductivity–photosynthesis interplay and aquaporin expression in field-grown mulberry (Morus spp.). Tree Physiol. 2017, 37, 926–937. [Google Scholar] [CrossRef]
- Merlaen, B.; De Keyser, E.; Ding, L.; Leroux, O.; Chaumont, F.; Van Labeke, M.C. Physiological responses and aquaporin expression upon drought and osmotic stress in a conservative vs prodigal Fragaria x ananassa cultivar. Plant Physiol. Biochem. 2019, 145, 95–106. [Google Scholar] [CrossRef]
- Muries, B.; Mom, R.; Benoit, P.; Brunel-Michac, N.; Cochard, H.; Drevet, P.; Petel, G.; Badel, E.; Fumanal, B.; Gousset-dupont, A.; et al. Aquaporins and water control in drought-stressed poplar leaves: A glimpse into the extraxylem vascular territories. Environ. Exp. Bot. 2019, 162, 25–37. [Google Scholar] [CrossRef]
- Pagès, H.A.; Gentleman, P.; DebRoy, R.S. R package version 2.56. 0. In Biostrings: Efficient Manipulation of Biological Strings; R Package, 2020; Available online: https://bioconductor.org/packages/release/bioc/html/Biostrings.html (accessed on 3 December 2020). [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar]
- Sehnal, D.; Vařeková, R.S.; Berka, K.; Pravda, L.; Navrátilová, V.; Banáš, P.; Ionescu, C.M.; Otyepka, M.; Koča, J. MOLE 2.0: Advanced approach for analysis of biomacromolecular channels. J. Cheminform. 2013, 5, 39. [Google Scholar]
- Laskowski, R.A.; MacArthur, M.W.; Thornton, J.M. Procheck: Validation of Protein-Structure Coordinates; Wiley: Hoboken, NJ, USA, 2006. [Google Scholar]
- Bertoni, M.; Kiefer, F.; Biasini, M.; Bordoli, L.; Schwede, T. Modeling protein quaternary structure of homo-and hetero-oligomers beyond binary interactions by homology. Sci. Rep. 2017, 7, 1–15. [Google Scholar]
- Bienert, S.; Waterhouse, A.; de Beer, T.A.; Tauriello, G.; Studer, G.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository—new features and functionality. Nucl. Acids. Res. 2017, 45, D313–D319. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucl. Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Gasic, K.; Hernandez, A.; Korban, S.S. RNA extraction from different apple tissues rich in polyphenols and polysaccharides for cDNA library construction. Plant. Mol. Biol. Rep. 2004, 22, 437–438. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Welch, B.L. The generalization of student’s’ problem when several different population variances are involved. Biometrika 1947, 34, 28–35. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Series B Stat. Methodol. 1995, 57, 289–300. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Nucleotide Length (bp) | Amino Acid Number | Coding Sequence (CDS) | MW (kDa) | pI | II | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| IwPIP1;4 | 673 | 190 | partial | 20.76 | 9.51 | 22.08 | Plasma membrane |
| IwPIP2;2 | 1280 | 283 | complete | 30.05 | 8.22 | 26.50 | Plasma membrane |
| IwPIP2;7 | 1302 | 286 | complete | 30.51 | 8.61 | 26.68 | Plasma membrane |
| IwTIP4;1 | 1010 | 248 | complete | 26.39 | 5.91 | 21.26 | vacuole |
| Protein Name | A, B, L | a, b, l, p | ~a,~b,~l,~p | Disallowed Regions (%) | Energetically Allowed (%) Ʃ |
|---|---|---|---|---|---|
| IwPIP1;4 | 81.5 | 12.1 | 4.5 | 1.9 | 98.1 |
| IwPIP2;2 | 79.9 | 13.6 | 4.2 | 2.3 | 97.7 |
| IwPIP2;7 | 80.3 | 12.8 | 3.7 | 3.2 | 96.8 |
| IwTIP4;1 | 91.8 | 7.7 | 0.5 | 0 | 100 |
| Gene Nme | Accesion Number | Primer Sequence | Ta (°C) | Amplicon Length (bp) |
|---|---|---|---|---|
| IwPIP1;4 | MW316882 | FW 5′-ACACTCTTCTGAAAGGCGG-3′R 5′-AGACCCAGTGATCGTTCCAG-3′ | 60 | 300 |
| IwPIP2;2 | MW316883 | FW 5′-AGCCGTTGAAGATCATGGGTTA-3′R 5′-CAATCCCTCCAAATCAATACCCT-3′ | 60 | 136 |
| IwPIP2;7 | MW316884 | FW 5′-TGGGTTGCTCTGTTCTGTCA-3′R 5′-GTGGGTCGTGGTAGTCCTTG-3′ | 60 | 136 |
| IwTIP4;1 | MW316885 | FW 5′-GCGAGTCCACCTCCGATTAG-3′R 5′-CGATGAATCCCGCAAGGTCT-3′ | 60 | 100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Đurić, M.J.; Subotić, A.R.; Prokić, L.T.; Trifunović-Momčilov, M.M.; Cingel, A.D.; Dragićević, M.B.; Simonović, A.D.; Milošević, S.M. Molecular Characterization and Expression of Four Aquaporin Genes in Impatiens walleriana during Drought Stress and Recovery. Plants 2021, 10, 154. https://doi.org/10.3390/plants10010154
Đurić MJ, Subotić AR, Prokić LT, Trifunović-Momčilov MM, Cingel AD, Dragićević MB, Simonović AD, Milošević SM. Molecular Characterization and Expression of Four Aquaporin Genes in Impatiens walleriana during Drought Stress and Recovery. Plants. 2021; 10(1):154. https://doi.org/10.3390/plants10010154
Chicago/Turabian StyleĐurić, Marija J., Angelina R. Subotić, Ljiljana T. Prokić, Milana M. Trifunović-Momčilov, Aleksandar D. Cingel, Milan B. Dragićević, Ana D. Simonović, and Snežana M. Milošević. 2021. "Molecular Characterization and Expression of Four Aquaporin Genes in Impatiens walleriana during Drought Stress and Recovery" Plants 10, no. 1: 154. https://doi.org/10.3390/plants10010154
APA StyleĐurić, M. J., Subotić, A. R., Prokić, L. T., Trifunović-Momčilov, M. M., Cingel, A. D., Dragićević, M. B., Simonović, A. D., & Milošević, S. M. (2021). Molecular Characterization and Expression of Four Aquaporin Genes in Impatiens walleriana during Drought Stress and Recovery. Plants, 10(1), 154. https://doi.org/10.3390/plants10010154