
The Impact of Two Different Cold-Extruded Feeds and Feeding Regimens on Zebrafish Survival, Growth and Reproductive Performance


Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Strains and Embryo Generation
2.2. Experimental Dietary Groups
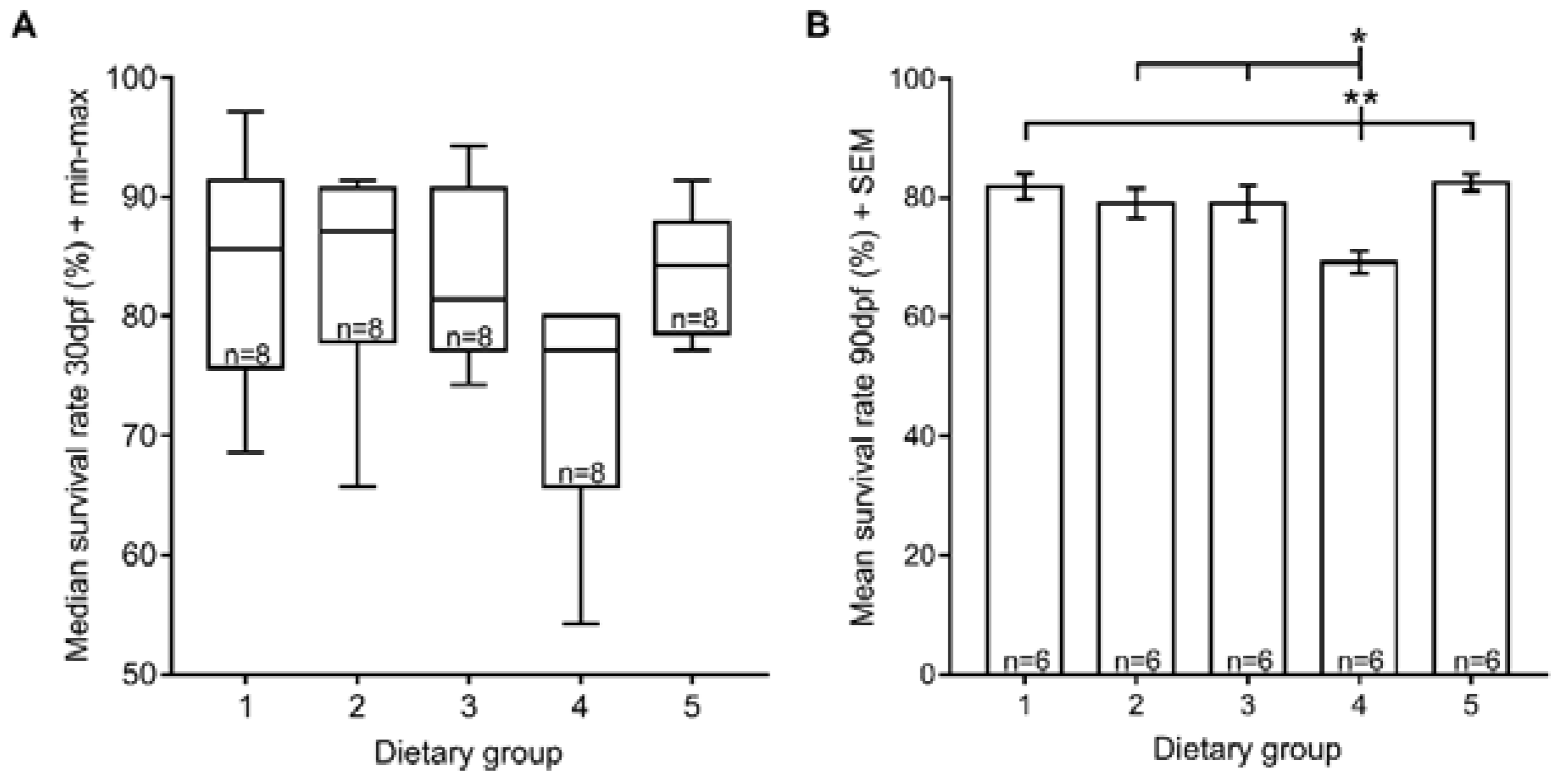
2.3. Fish Survival Rate
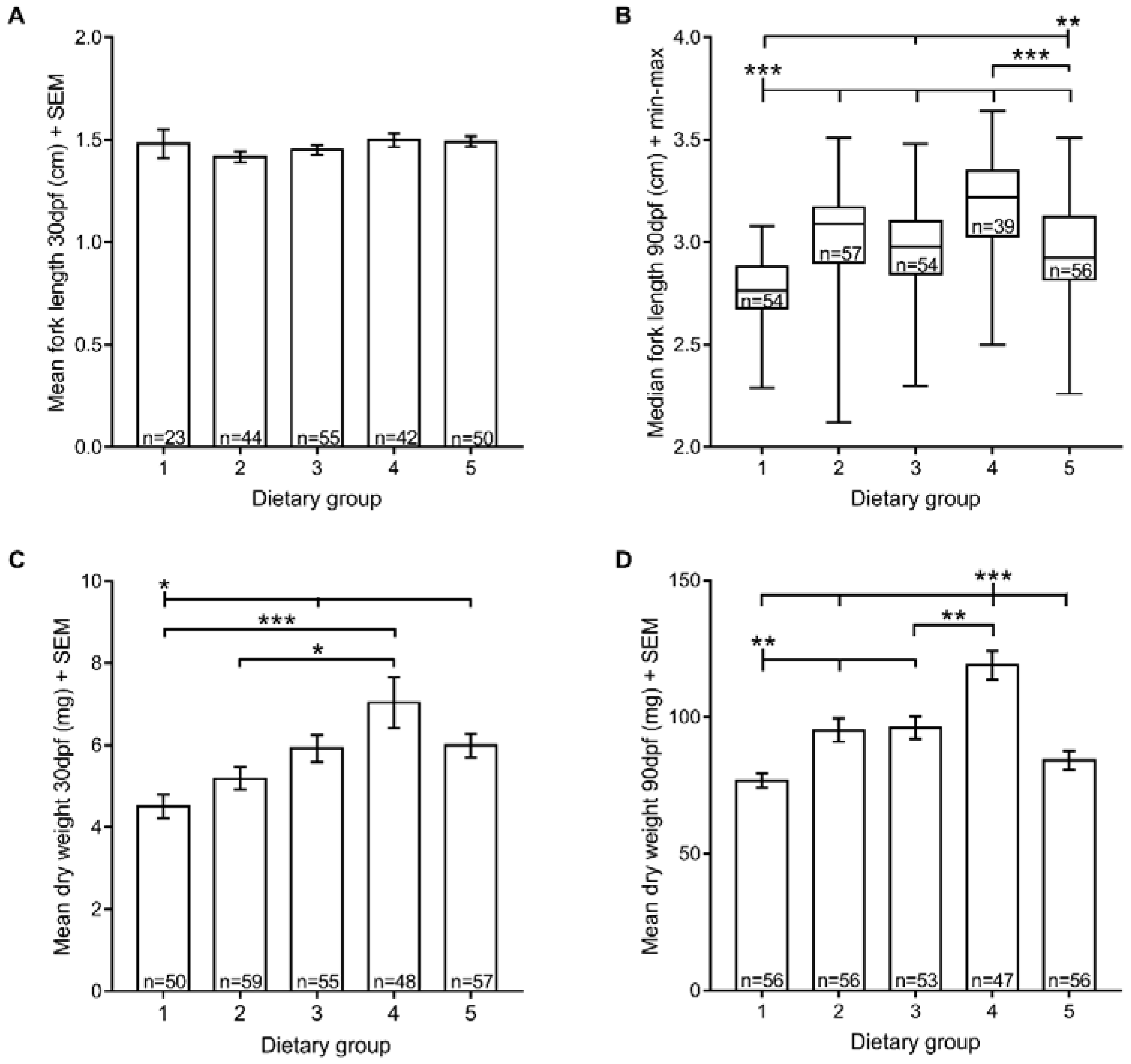
2.4. Growth
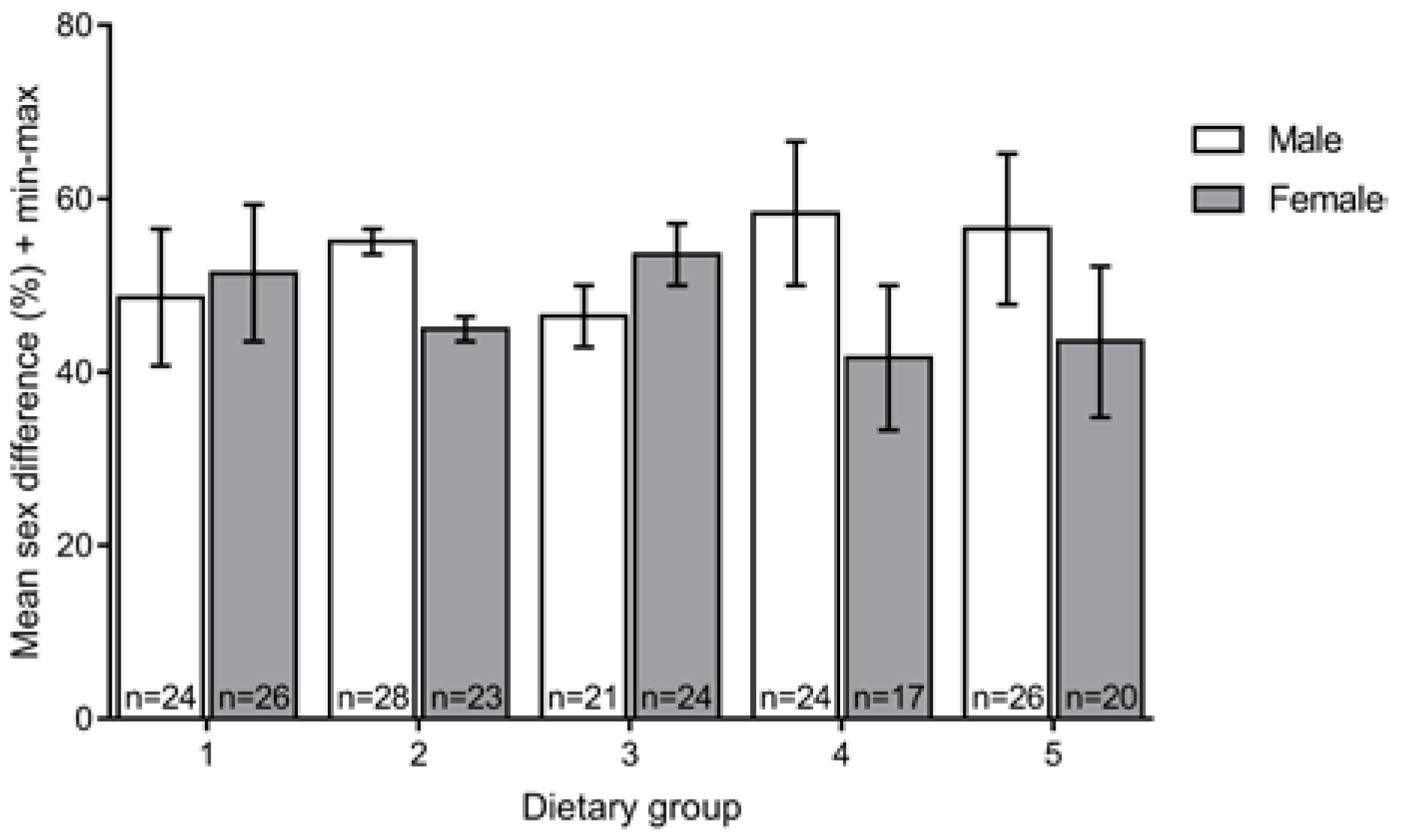
2.5. Fish Sex Ratio
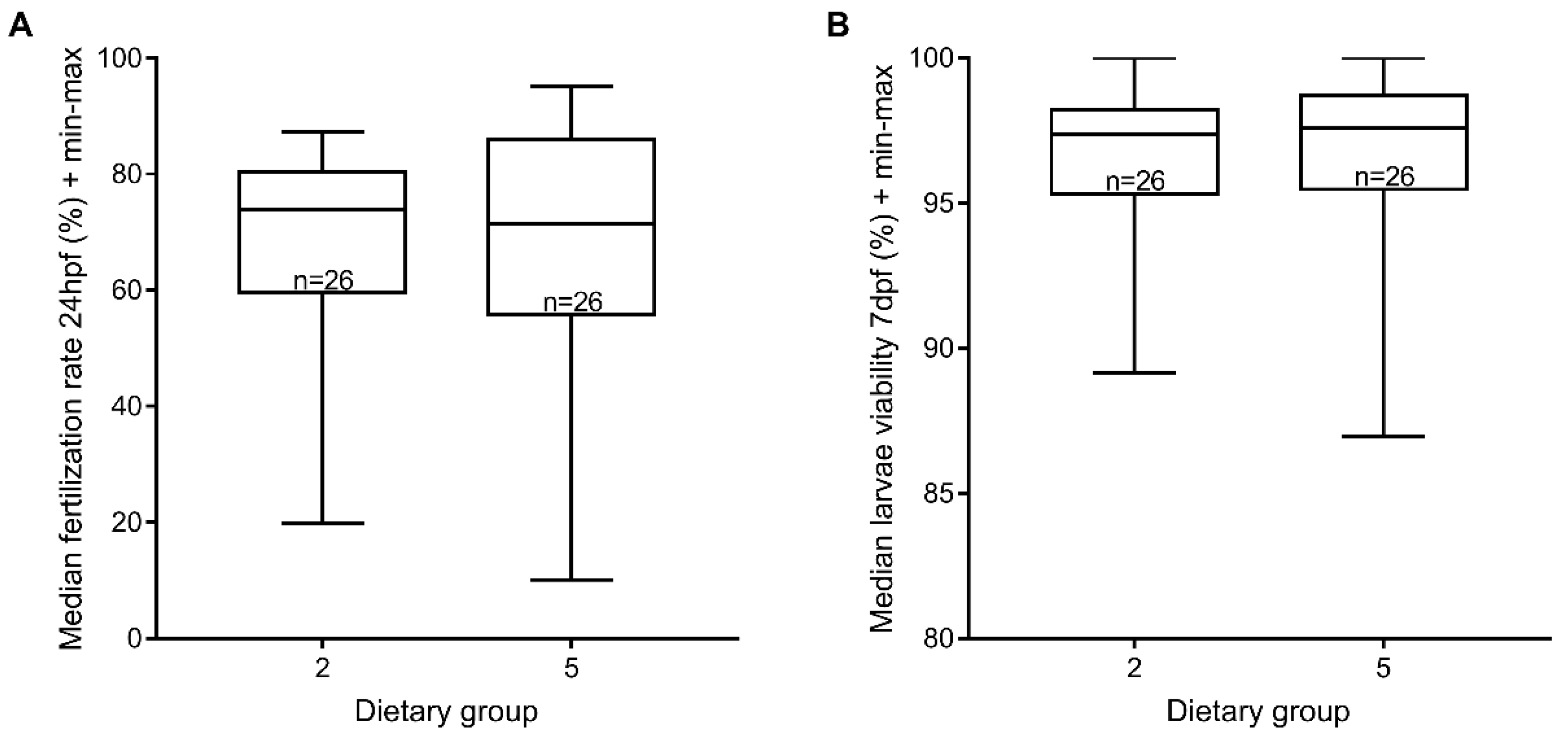
2.6. Reproductive Performance
2.7. Statistical Analysis
3. Results
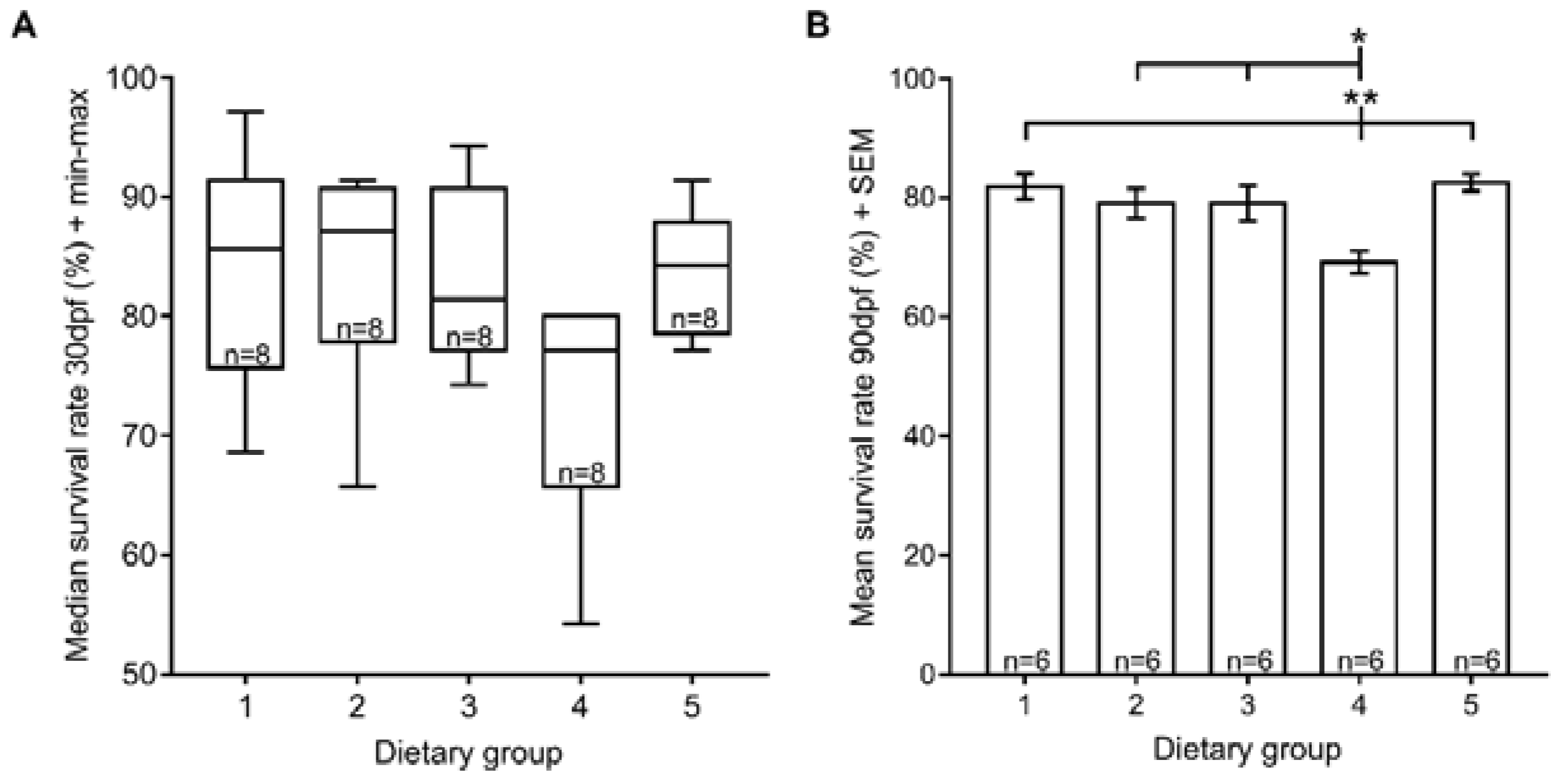
3.1. Survival Rate
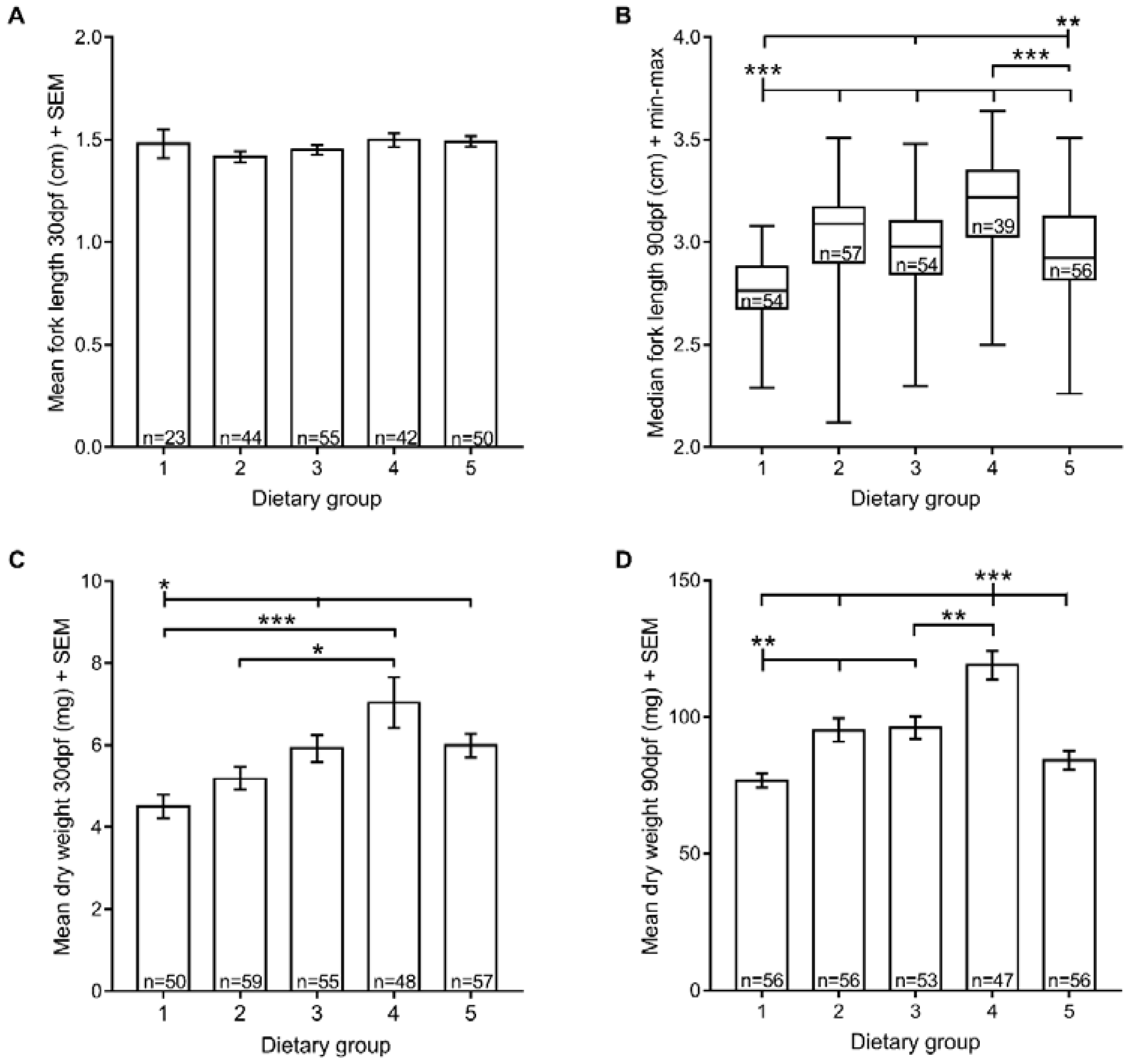
3.2. Growth
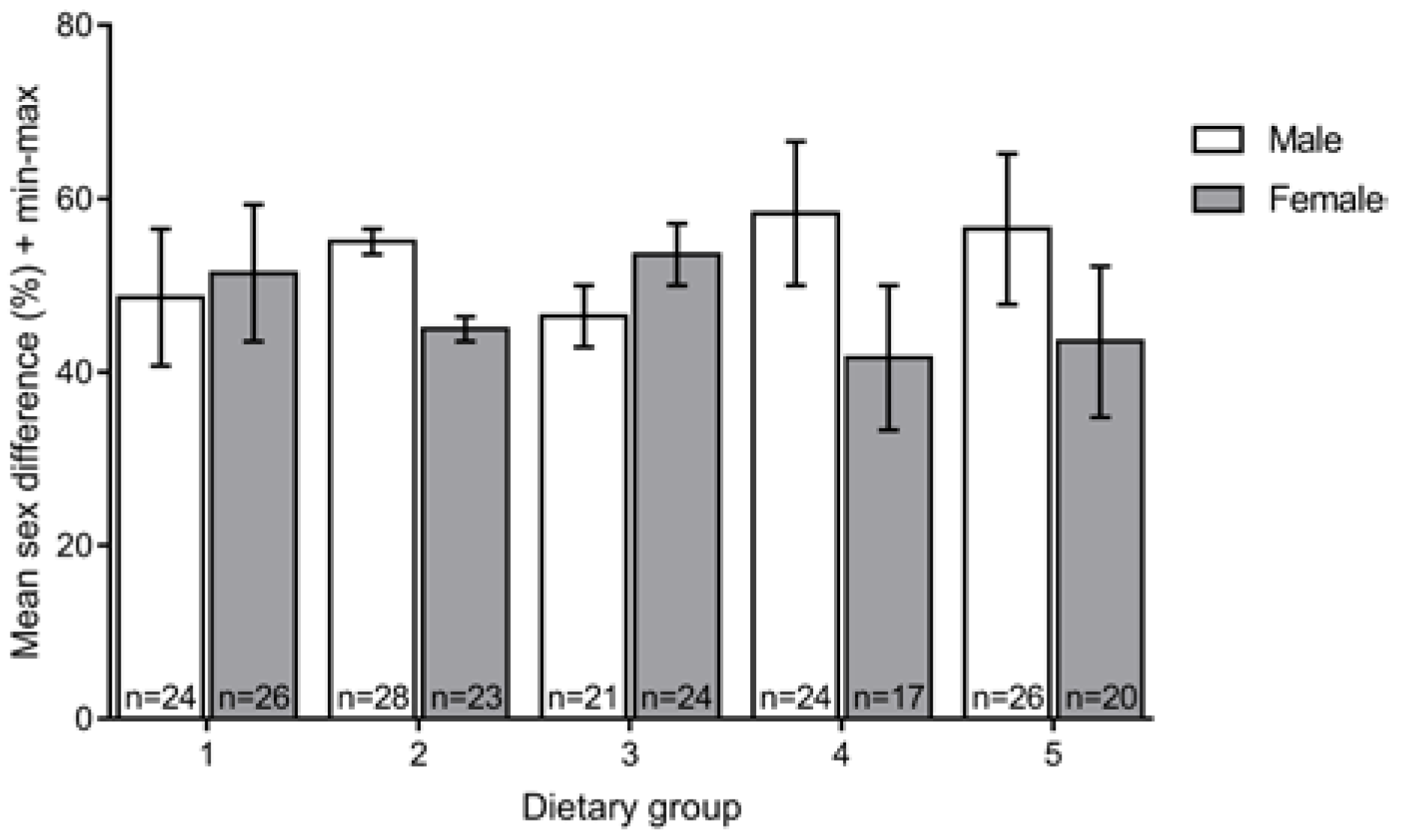
3.3. Sex Ratio
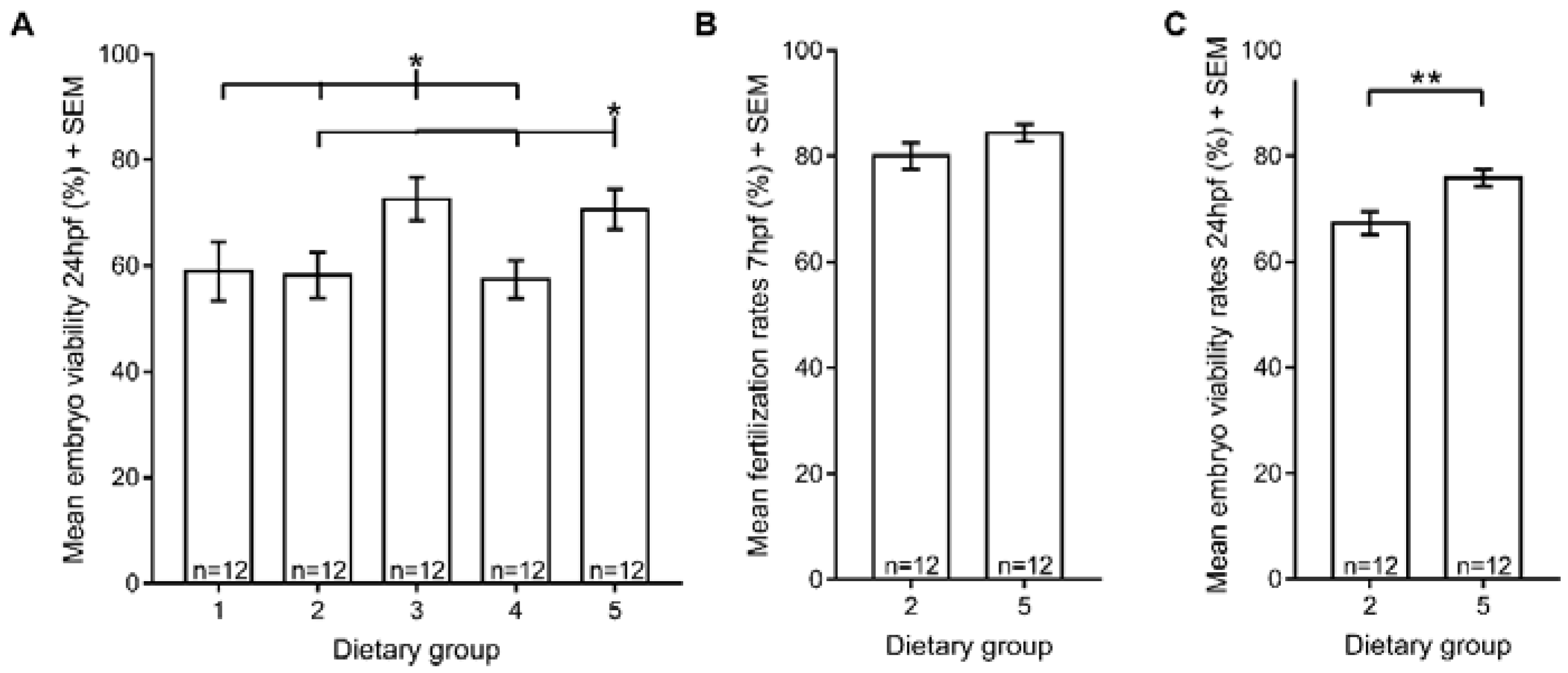
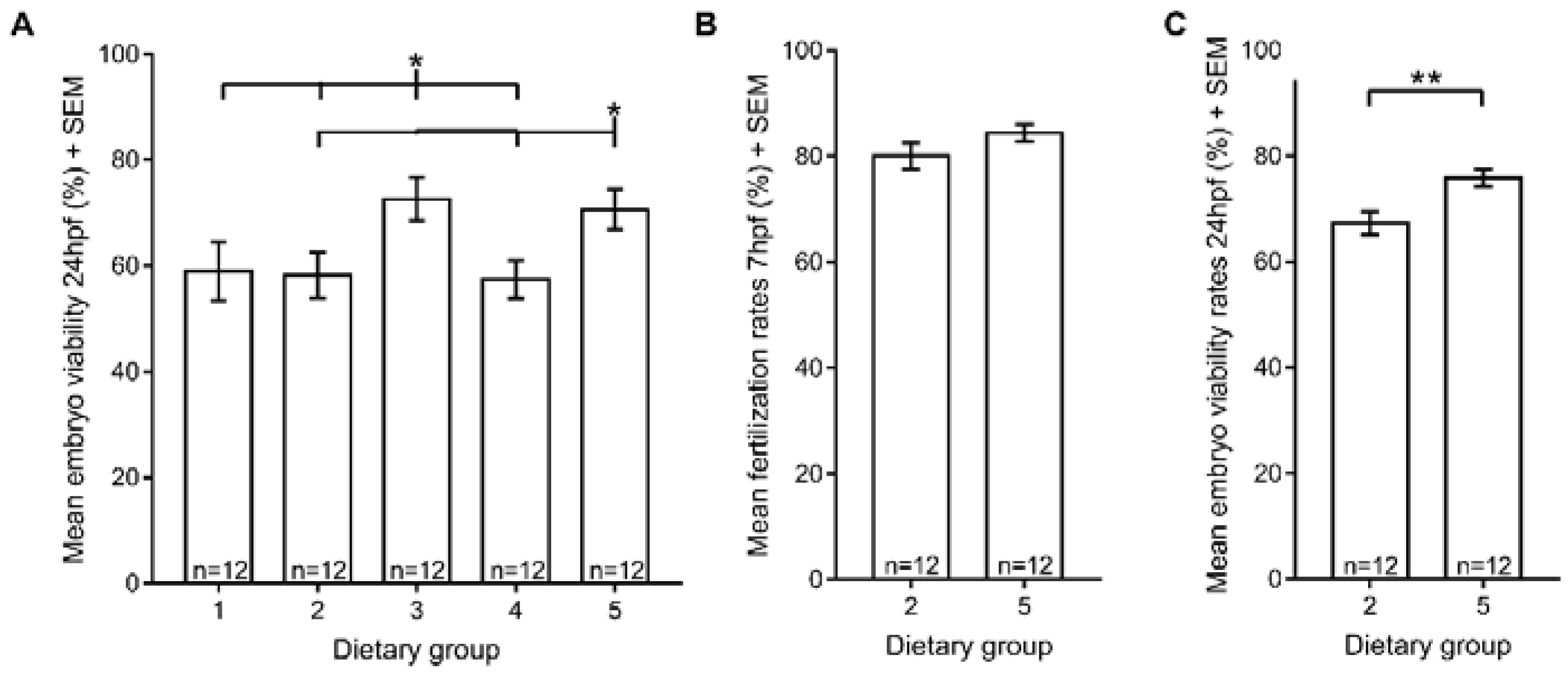
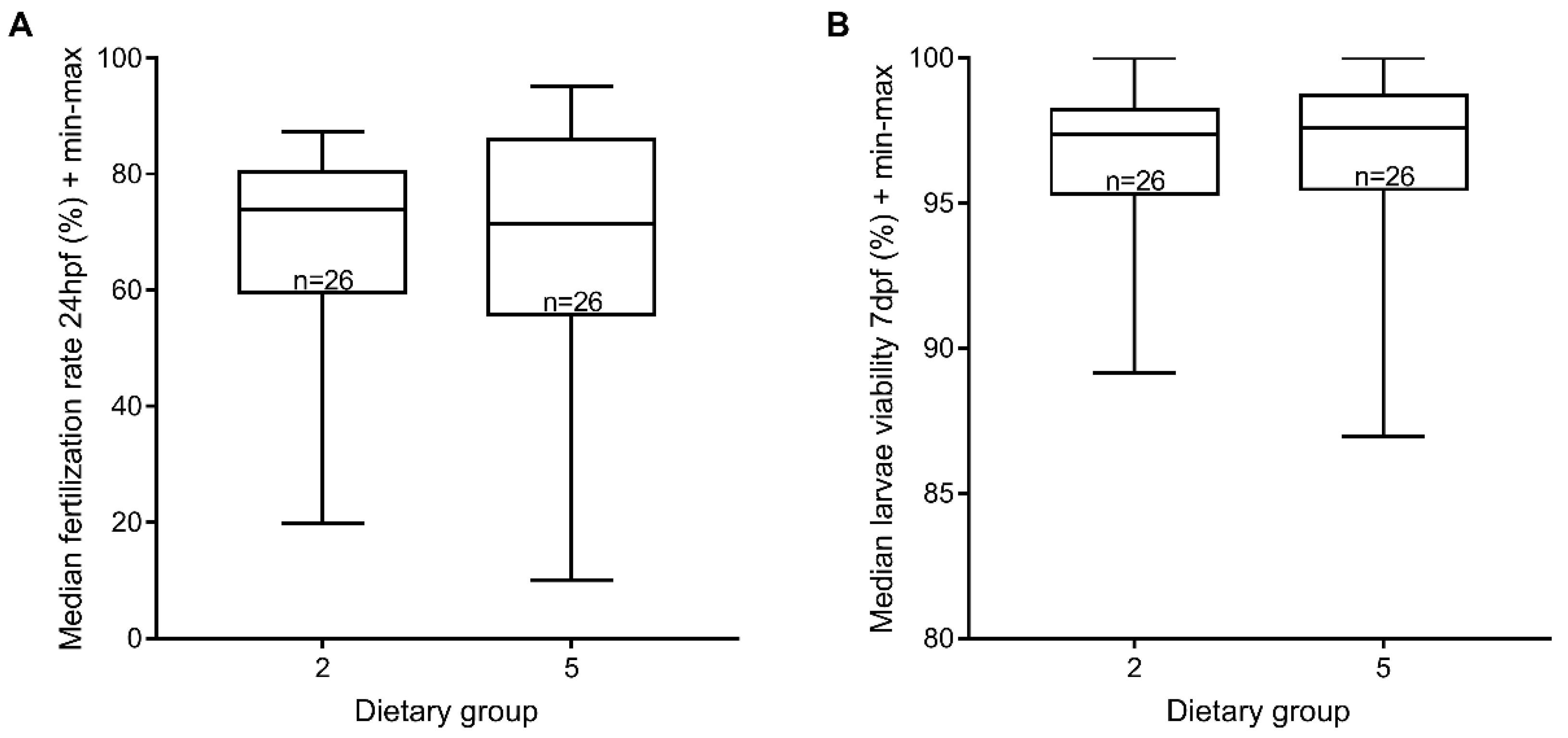
3.4. Reproductive Performance
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barnard, D.E.; Lewis, S.M.; Teter, B.B.; Thigpen, J.E. Open- and closed-formula laboratory animal diets and their importance to research. J. Am. Assoc. Lab. Anim. Sci. 2009, 48, 709–713. [Google Scholar] [PubMed]
- Harding, J.D.; Van Hoosier, J.L., Jr.; Grieder, F.B. The contribution of laboratory animals to medical progress—past, present and future. In Handbook of Laboratory Animal Science—Essential Principles and Practices, 3rd ed.; Hau, J., Schapiro, S.J., Eds.; CRC Press LLC: London, UK, 2011; Volume 1, pp. 1–20. ISBN 978-1-4200-8456-6. [Google Scholar]
- Hau, J. Animal models. In Handbook of Laboratory Animal Science—Animal Models, 2nd ed.; Hau, J., Gerald, L., Eds.; CRC Press: Boca Raton, FL, USA, 2005; Volume 2, pp. 1–11. ISBN 0-8493-1086-5. [Google Scholar]
- Watts, S.A.; Lawrence, C.; Powell, M.; D’Abramo, R.L. The Vital Relationship between Nutrition and Health in Zebrafish. Zebrafish 2016, 13, S72–S76. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C. The husbandry of zebrafish (Danio rerio): A review. Aquaculture 2007, 269, 1–20. [Google Scholar] [CrossRef]
- Kaushik, S.; Georgia, I.; Koumoundouros, G. Growth and body composition of zebrafish (Danio rerio) larvae fed a compound feed from first feeding onward: toward implications on nutrient requirements. Zebrafish 2011, 8, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Castranova, D.; Lawton, A.; Lawrence, C.; Baumann, D.; Best, J.; Coscolla, J.; Doherty, A.; Ramos, J.; Hakkesteeg, J.; Wang, C.; et al. The effect of stocking densities on reproductive performance in laboratory zebrafish (Danio rerio). Zebrafish 2011, 8, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J. Preliminary evaluations on the growth and early reproductive performance of zebrafish (Danio rerio). J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 412–417. [Google Scholar] [PubMed]
- SOFIA: The State of World Fisheries and Aquaculture; FAO Fisheries and Aquaculture Department: Roma, Italy, 2010.
- Geay, F.; Ferraresso, S.; Zambonino-Infante, J.L.; Bargelloni, L.; Quentel, C.; Vandeputte, M.; Kaushik, S.; Cahu, C.L.; Mazurais, D. Effects of the total replacement of fish based diet with plant-based diet on the hepatic transcriptome of two European sea bass (Dicentrarchus labrax) half-sibfamilies showing different growth rates with the plant-based diet. BMC Genom. 2011, 12, 522. [Google Scholar] [CrossRef] [PubMed]
- Sahlmann, C.; Sutherland, B.J.G.; Kortner, T.M.; Koop, B.F.; Krogdahl, A.; Bakke, A.M. Early response of gene expression in the distal intestine of Atlantic salmon (Salmo salar L.) during the development of soybean meal induced enteritis. Fish Shellfish Immunol. 2013, 34, 599–609. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, H.; Peres, H.; Carvalho, A.P. Dietary Protein Requirement during Juvenile Growth of Zebrafish (Danio rerio). Zebrafish 2016, 6, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C. The Zebrafish: Genetics, Genomics and Informatics. In Methods in Cell Biology, 3rd ed.; William, D.H., Ed.; Academic Press: Waltham, MA, USA, 2011; Volume 104, ISBN 9780080888576. [Google Scholar]
- Diogo, P.; Martins, G.; Gavaia, P.; Pinto, W.; Dias, J.; Cancela, L.; Martınez-Páramo, S. Assessment of nutritional supplementation in phospholipids on the reproductive performance of zebrafish, Danio rerio (Hamilton, 1822). Appl. Ichthyol. 2015, 31, 3–9. [Google Scholar] [CrossRef]
- Rainuzzo, J.R.; Reitan, K.I.; Olsen, Y. The significance of lipids at early stages of marine fish: A review. Aquaculture 1997, 155, 103–115. [Google Scholar] [CrossRef]
- Watanabe, T. Importance of the study of broodstock nutrition for further development of aquaculture. In Nutrition and Feeding in Fish; Cowley, C.B., Mackie, A.M., Bell, J.K., Eds.; Academic Press: London, UK, 1985; pp. 395–414. [Google Scholar]
- Fernandez-Palacios, H.; Izquierdo, M.; Robaina, L.; Valencia, A.; Salhi, M.; Montero, D. The effect of dietary protein and lipid from squid and fish meals on egg quality of broodstock for gilthead seabream (Sparus aurata). Aquaculture 1997, 148, 233–246. [Google Scholar] [CrossRef]
- Izquierdo, M.S.; Fernandez-Palacios, H.; Tacon, A.G.J. Effect of broodstock nutrition on reproductive performance of fish. Aquaculture 2001, 197, 25–42. [Google Scholar] [CrossRef]
- Alsop, D.; Matsumoto, J.; Brown, S.; Van Der Kraak, G. Retinoid requirements in the reproduction of zebrafish. Gen. Comp. Endocrinol. 2008, 156, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Jaya-Ram, A.; Kuah, M.K.; Lim, P.S.; Kolkovski, S.; Shu-Chien, A.C. Influence of dietary HUFA levels on reproductive performance, tissue fatty acid profile and desaturase and elongase mRNAs expression in female zebrafish Danio rerio. Aquaculture 2008, 277, 275–281. [Google Scholar] [CrossRef]
- Miller, G.W.; Labut, E.M.; Lebold, K.M.; Floeter, A.; Tanguay, R.L.; Traber, M.G. Zebrafish (Danio rerio) fed vitamin E-deficient diets produce embryos with increased morphologic abnormalities and mortality. J. Nutr. Biochem. 2012, 23, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Robinson, B.D.; Drew, R.E.; Murdoch, G.K.; Powell, M.; Rodnick, K.J.; Settles, M.; Stone, D.; Churchill, E.; Hill, R.A.; Papasani, R.A.; et al. Sexual dimorphism in hepatic gene expression and the response to dietary carbohydrate manipulation in the zebrafish (Danio rerio). Comp. Biochem. Physiol. D-Genom. Proteom. 2008, 3, 141–154. [Google Scholar] [CrossRef]
- Cabrita, E.; Robles, V.; Sarasquete, C.; Herráez, M.P. New insights on sperm quality analysis for the improvement of broodstock. In Cryopreservation of Aquatic Species, 2nd ed.; Tiersch, T., Mazik, P.M., Eds.; World Aquaculture Society: Baton Rouge, LA, USA, 2011; pp. 146–161. [Google Scholar]
- James, G.; Witten, D.; Hastie, T.; Tibshirani, R. Statistical learning. In An Introduction to Statistical Learning: with Applications in R; Springer Publishing Company: New York, NY, USA, 2013; pp. 21–24. ISBN 978-1-4614-7138-7. [Google Scholar]
- Martins, S.; Monteiro, J.F.; Vito, M.; Weintraub, D.; Almeida, J.; Certal, A.C. Toward an Integrated Zebrafish Health Management Program Supporting Cancer and Neuroscience Research. Zebrafish 2016, 13 (Suppl. 1), 47–55. [Google Scholar] [CrossRef] [PubMed]
- Westerfield, M. General Methods for zebrafish care. In The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio), 4th ed.; Univ. of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Zebrafish International Resource Center—Monitoring of Fish Morbidity and Mortality. Available online: http://zebrafish.org/documents/protocols/pdf/health_monitoring/daily_monitoring_fish_morbidity_07_2015.pdf (accessed on 12 April 2018).
- Darrow, K.O.; Harris, W. Characterization and development of courtship in Zebrafish, Danio rerio. Zebrafish 2004, 1, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Zebrafish in the Classroom. Available online: http://www.zfic.org/common%20techniques/Gender%20identification%20guide.pdf (accessed on 26 May 2018).
- Farias, M.; Certal, A.C. Different Feeds and Feeding Regimens have an Impact on Zebrafish Larval Rearing and Breeding Performance. SOJ Aquat. Res. 2016, 1, 1–8. [Google Scholar]
- Yang, H.; Tiersch, T.R. Current status of sperm cryopreservation in biomedical research fish models: Zebrafish, Medaka, and Xiphophorus. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2009, 149, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Best, J.; Adatto, I.; Cockington, J.; James, A.; Lawrence, C. A novel method for rearing first-feeding larval zebrafish: polyculture with Type L saltwater rotifers (Brachionus plicatilis). Zebrafish 2010, 7, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C.; James, A.; Mobley, S. Successful Replacement of Artemia salina nauplii with Marine Rotifers (Brachionus plicatilis) in the Diet of Preadult Zebrafish (Danio rerio). Zebrafish 2015, 12, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Boyaci, B.B.; Han, J.; Masatcioglu, M.T.; Yalcin, E.; Celik, S.; Ryu, G.; Koksel, H. Analytical Methods—Effects of cold extrusion process on thiamine and riboflavin contents of fortified corn extrudates. Food Chem. 2012, 132, 2165–2170. [Google Scholar] [CrossRef]
- Steel, C.J.; Leoro, M.G.V.; Schmiele, M.; Ferreira, R.E.; Chang, Y.K. Thermoplastic Extrusion in Food Processing. In Thermoplastic Elastomers; El-Sonbati, A., Ed.; InTech Europe; University Campus: Rijeka, Croatia, 2012; pp. 265–290. ISBN 978-953-51-0346-2. [Google Scholar]
- New! TECH Feature: Cold Extrusion—Headline News. Available online: http://www.aquafeed.com/news/headline-news-article/601/NEW-TECH-FEATURE-COLD-EXTRUSION/ (accessed on 21 March 2018).
- Watts, S.A.; Powell, M.; D’Abramo, L.R. Fundamental approaches to the study of zebrafish nutrition. ILAR J. 2012, 53, 144–160. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C. Aspects of larval rearing. ILAR J. 2012, 53, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Siccardi, A.J.; Garris, H.W.; Jones, W.T.; Moseley, D.B.; D’Abramo, L.R.; Watts, SA. Growth and survival of zebrafish (Danio rerio) fed different commercial and laboratory diets. Zebrafish 2009, 6, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.P.; Araújo, L.; Santos, M.M. Rearing zebrafish (Danio rerio) larvae without live food: Evaluation of a commercial, a practical and a purified starter diet on larval performance. Aquac. Res. 2006, 37, 1107–1111. [Google Scholar] [CrossRef]
- Goolish, E.M.; Okutake, K.; Lesure, S. Growth and Survivorship of Larval Zebrafish Danio rerio on Processed Diets. N. Am. J. Aquac. 1999, 61, 189–198. [Google Scholar] [CrossRef]
- Gonzales, J.M., Jr.; Law, S.H. Feed and feeding regime affect growth rate and gonadosomatic index of adult zebrafish (Danio rerio). Zebrafish 2013, 10, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Mishra, U.; Roy, S.D.; Chadha, N.K.; Venkateshwarlu, G. Effect of feeding enriched formulated diet and live feed on growth, survival and fatty acid profile of Deccan Mahseer, Tor khudree (Sykes) first feeding fry. J. Aquac. Res. Dev. 2012, 3. [Google Scholar] [CrossRef]
- Schramm, M.J. “Tiny Krill: Giants in Marine Food Chain”. NOAA National Marine Sanctuary Program. Available online: https://sanctuaries.noaa.gov/news/features/1007_krill.html (accessed on 1 June 2018).
- Lawrence, C.; Best, J.; James, A.; Maloney, K. The effects of feeding frequency on growth and reproduction in zebrafish (Danio rerio). Aquaculture 2012, s368–s369, 103–108. [Google Scholar] [CrossRef]
- Hopkins, K.D. Reporting Fish Growth: A Review of the Basics. J. World Aquac. Soc. 1992, 23, 173–179. [Google Scholar] [CrossRef]
- Gómez-Requeni, P.; Conceição, L.E.; Jordal, A.E.O.; Rønnestad, I. A reference growth curve for nutritional experiments in zebrafish (Danio rerio) and changes in whole body proteome during development. Fish Physiol. Biochem. 2010, 36, 1199–1215. [Google Scholar] [CrossRef] [PubMed]
- Harper, C.; Lawrence, C. Husbandry. In The Laboratory Zebrafish; Suckow, M.A., Ed.; CRC Press: Boca Raton, FL, USA, 2011; pp. 47–59. ISBN 9781439807439. [Google Scholar]
- Bobe, J.; Labbé, C. Egg and sperm quality in fish. Gen. Comp. Endocrinol. 2010, 165, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Nowosad, J.; Kucharczyk, D.; Targońska, K. Enrichment of Zebrafish Danio rerio (Hamilton, 1822) Diet with Polyunsaturated Fatty Acids Improves Fecundity and Larvae Quality. Zebrafish 2017, 14, 364–370. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G.; Atkinson, J. Vitamin E antioxidant and nothing more. Free Radic. Biol. Med. 2007, 43, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, C.; Sanders, E.; Henry, E. Methods for culturing saltwater rotifers (Brachionus licatilis) for rearing larval zebrafish. Zebrafish 2012, 9, 140–146. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Age (dpf) | Skretting®GemmaMicro | Sparos®Zebrafeed | Live Feed | |
|---|---|---|---|---|
| Experimental Dietary Group 1 | 8–15 | - | 1× 100–200 | 3× Rotifers |
| 15–30 | - | 1× 100–200 | 3× Rotifers | |
| 30–60 | - | 1× 200–400 | 3× Rotifers | |
| 60–90 | - | 3× 200–400 | - | |
| >90 | - | 2× 400–600 | - | |
| Experimental Dietary Group 2 | 8–15 | 1× 150 | - | 3× Rotifers |
| 15–30 | 1× 150 | - | 3× Rotifers | |
| 30–60 | 1× 300 | - | 3× Rotifers | |
| 60–90 | 3× 300 | - | - | |
| >90 | 2× 500 | - | - | |
| Experimental Dietary Group 3 | 8–15 | - | 1× 100–200 | 3× Rotifers |
| 15–30 | - | 4× 100–200 | - | |
| 30–60 | - | 4× 200–400 | - | |
| 60–90 | - | 3× 200–400 | - | |
| >90 | - | 2× 400–600 | - | |
| Experimental Dietary Group 4 | 8–15 | 1× 150 | 3× Rotifers | |
| 15–30 | 4× 150 | - | ||
| 30–60 | 4× 300 | - | ||
| 60–90 | 3× 300 | - | ||
| >90 | 2× 500 | - | ||
| Experimental Dietary Group 5 | 8–15 | 1× 150 | - | 3× Rotifers |
| 15–30 | 1× 150 | - | 3× Rotifers | |
| 30–60 | 1× 300 | - | 3× Rotifers | |
| 60–90 | - | 3× 200–400 | - | |
| >90 | - | 2× 400–600 | - |
| Skretting® Gemma Micro (%) | Sparos® Zebrafeed (%) | |
|---|---|---|
| Protein | 59 | 63 |
| Fat and Oils | 14 | 14 |
| Fiber | 0.2 | 1.8 |
| Ash | 14 | 12 |
| Phosphorus | 1.3 | 2.3 |
| Main ingredients | Fish meal, lecithin, wheat gluten, dried seaweed, fish oil, maize starch, vitamins and minerals | Squid meal, wheat gluten, fish meal, fish protein concentrate, krill meal, pea protein concentrate, starch, lecithin, krill oil, fish oil. |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, J.F.; Martins, S.; Farias, M.; Costa, T.; Certal, A.C. The Impact of Two Different Cold-Extruded Feeds and Feeding Regimens on Zebrafish Survival, Growth and Reproductive Performance. J. Dev. Biol. 2018, 6, 15. https://doi.org/10.3390/jdb6030015
Monteiro JF, Martins S, Farias M, Costa T, Certal AC. The Impact of Two Different Cold-Extruded Feeds and Feeding Regimens on Zebrafish Survival, Growth and Reproductive Performance. Journal of Developmental Biology. 2018; 6(3):15. https://doi.org/10.3390/jdb6030015
Chicago/Turabian StyleMonteiro, Joana F., Sandra Martins, Matheus Farias, Telma Costa, and Ana Catarina Certal. 2018. "The Impact of Two Different Cold-Extruded Feeds and Feeding Regimens on Zebrafish Survival, Growth and Reproductive Performance" Journal of Developmental Biology 6, no. 3: 15. https://doi.org/10.3390/jdb6030015
APA StyleMonteiro, J. F., Martins, S., Farias, M., Costa, T., & Certal, A. C. (2018). The Impact of Two Different Cold-Extruded Feeds and Feeding Regimens on Zebrafish Survival, Growth and Reproductive Performance. Journal of Developmental Biology, 6(3), 15. https://doi.org/10.3390/jdb6030015