Ethanol Exposure Causes Muscle Degeneration in Zebrafish



Abstract
:1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry, Transgenic Lines, and Mutant Lines
2.2. Ethanol (EtOH) and Trichostatin A (TSA) Treatment
2.3. Heat Shock Treatment
2.4. Evans Blue Dye (EBD) Injection
2.5. Immunohistochemistry
2.6. Imaging
2.7. Analysis and Statistics
3. Results
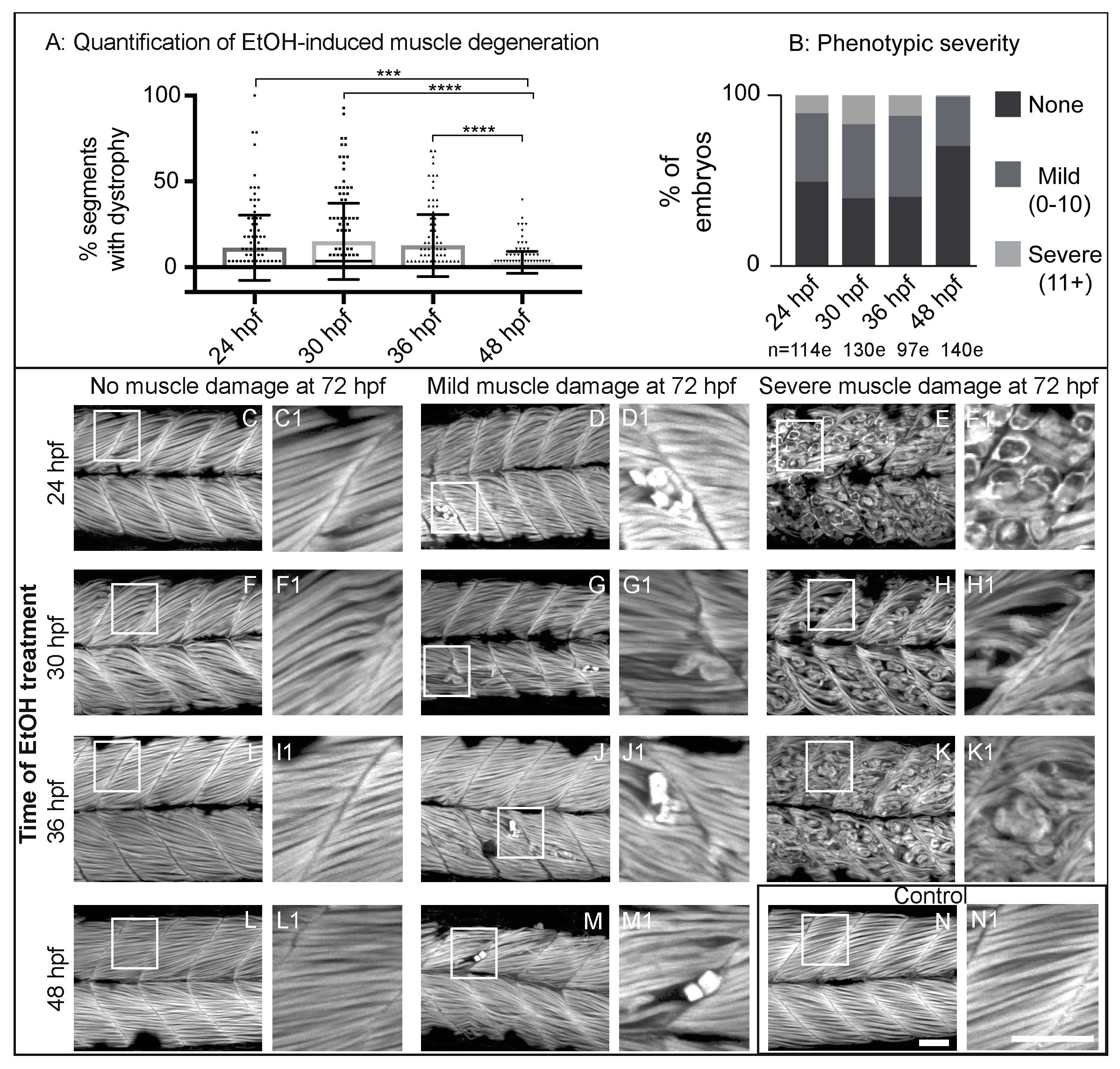
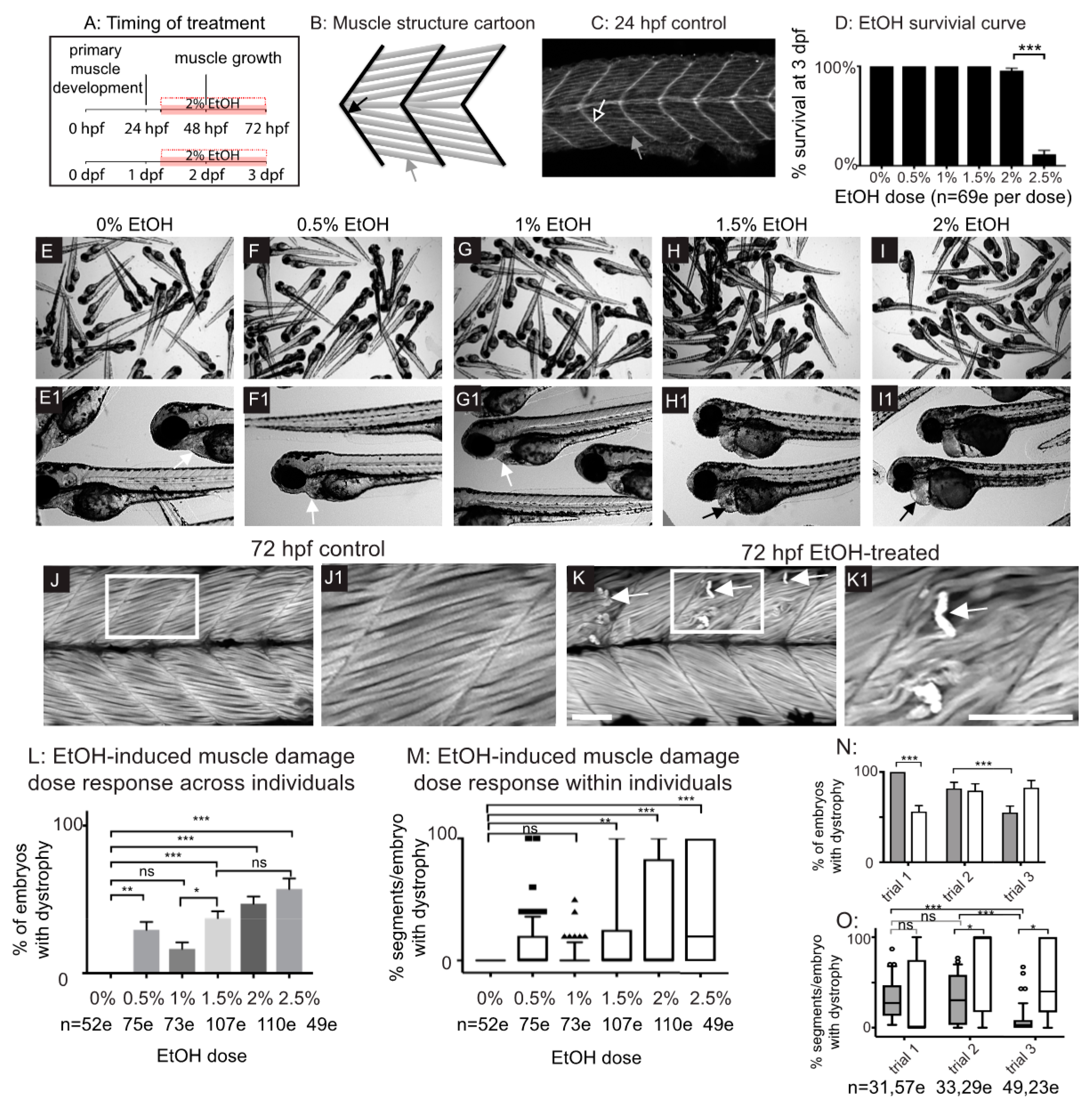
3.1. Late Embryonic Exposure to EtOH Causes Muscle Damage in Zebrafish
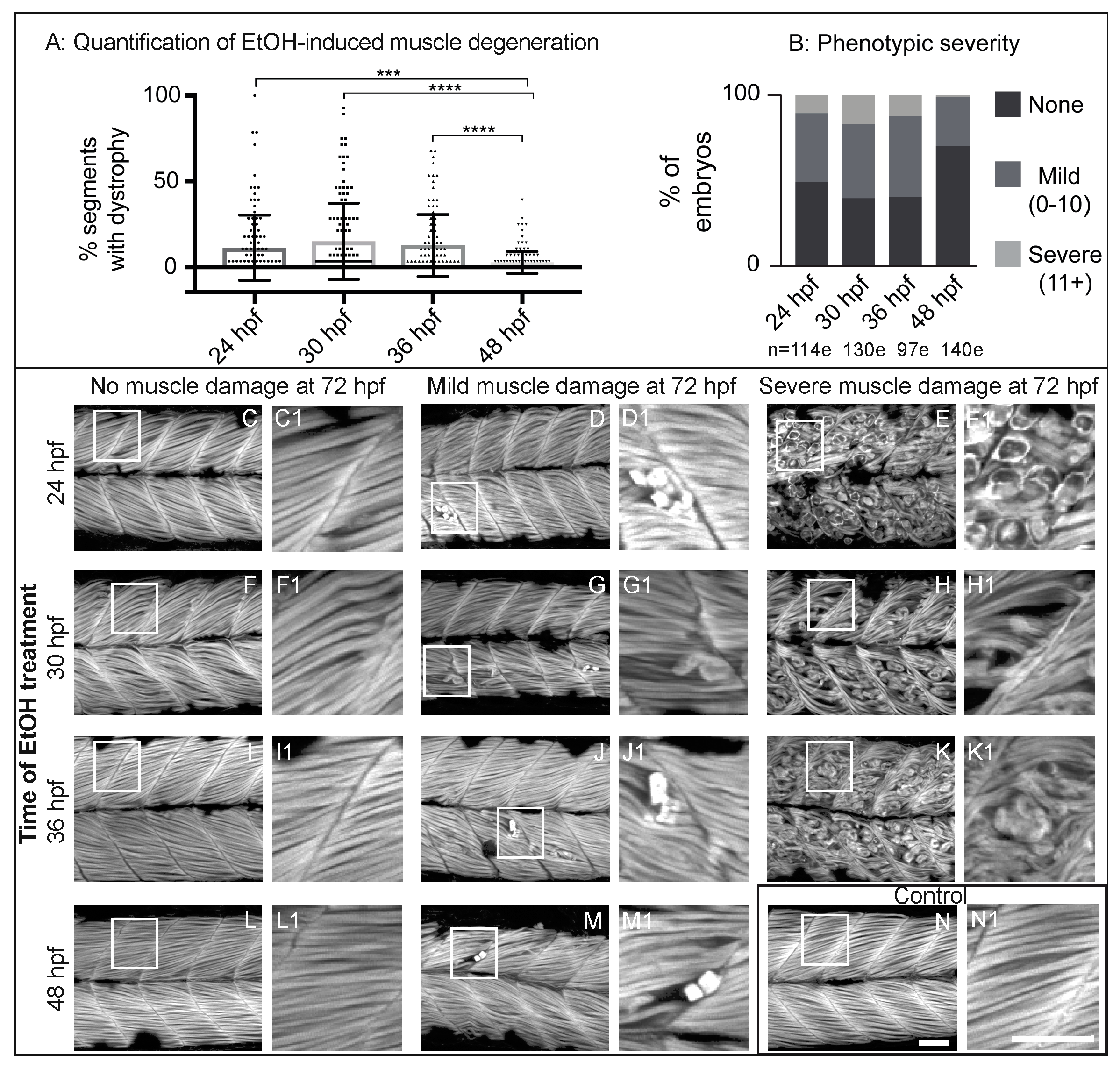
3.2. Zebrafish Muscle Is Less Sensitive to EtOH That Is Administered Later in Development
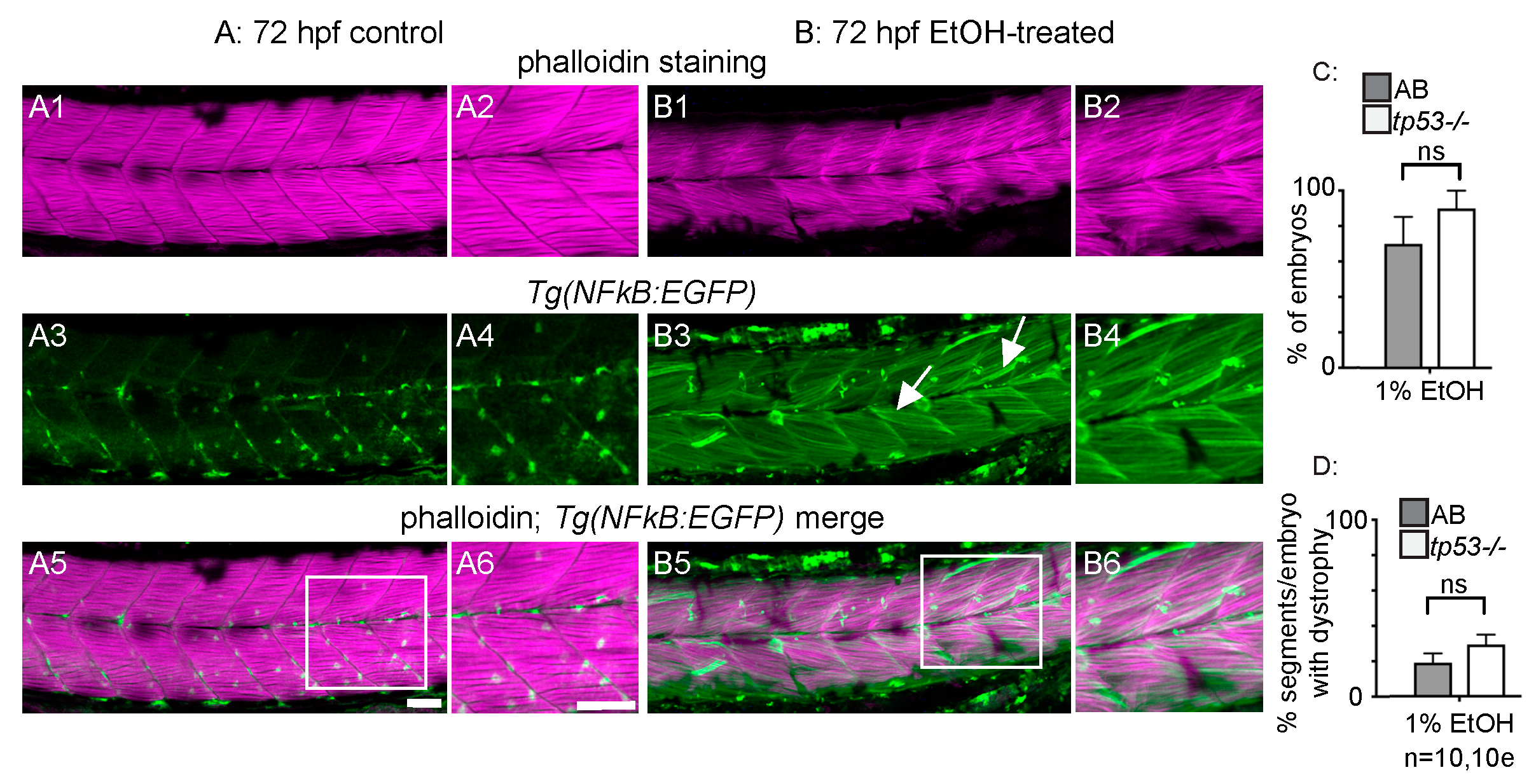
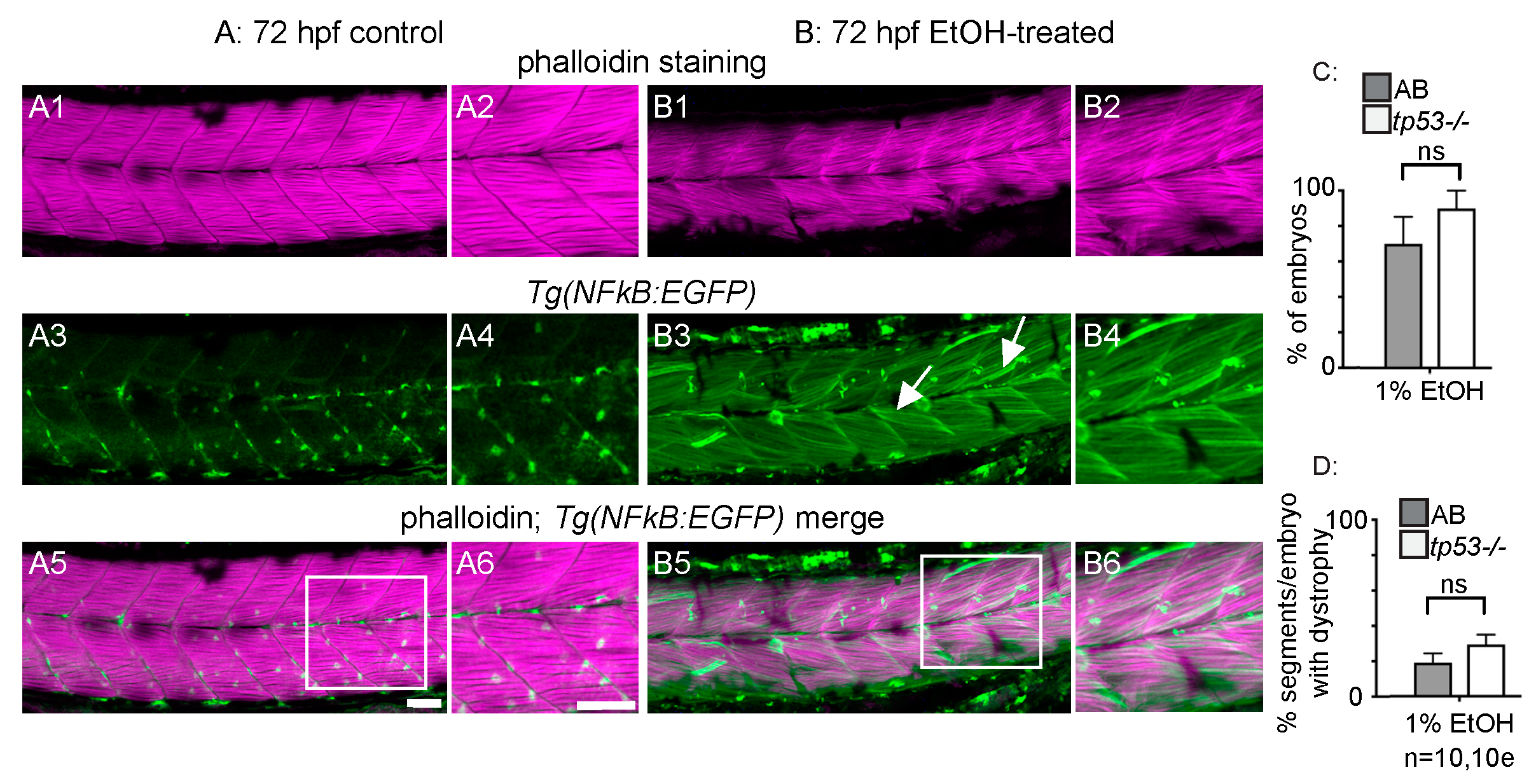
3.3. p53-Dependent Apoptosis Is Not Required for EtOH-Induced Fiber Detachments
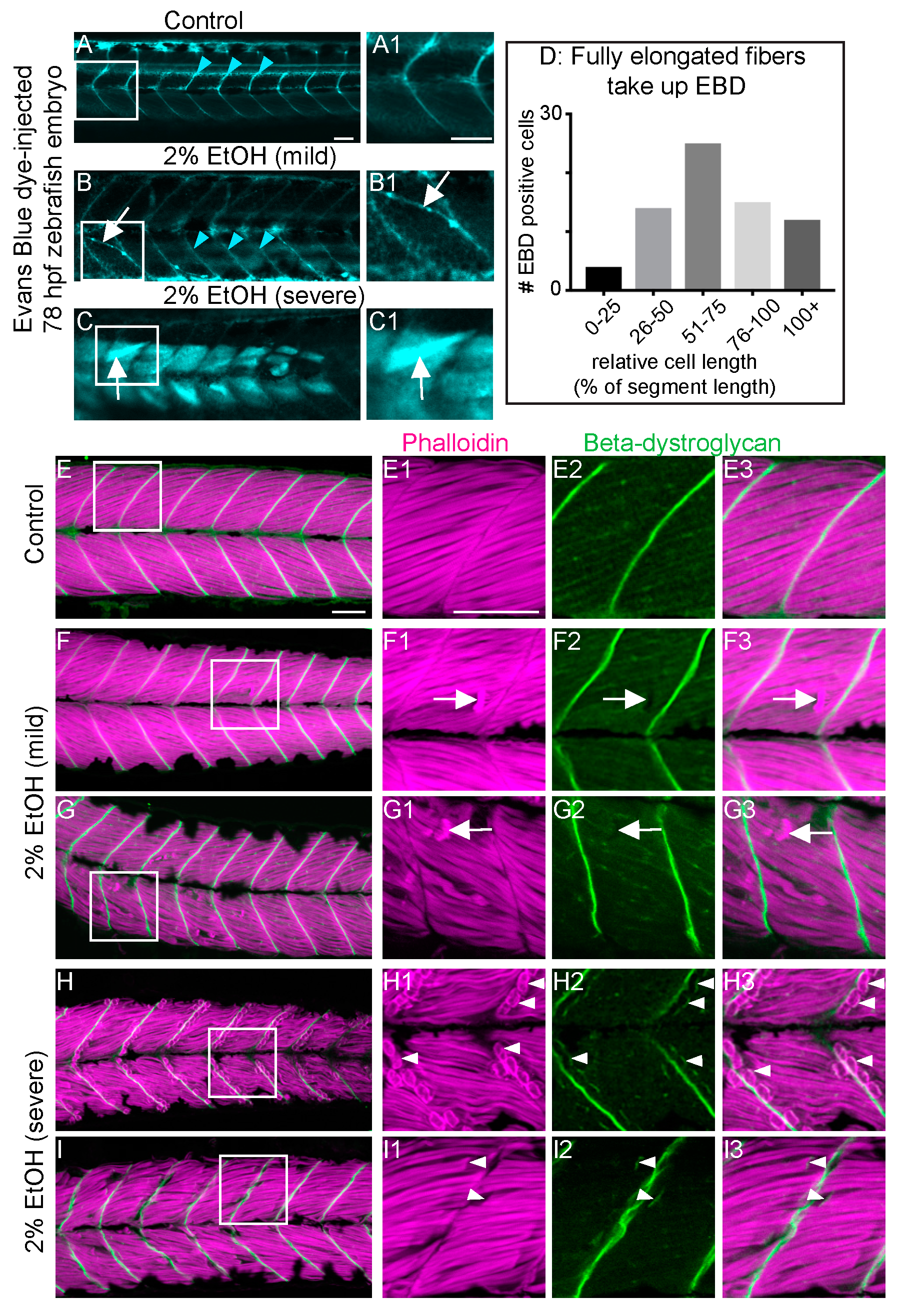
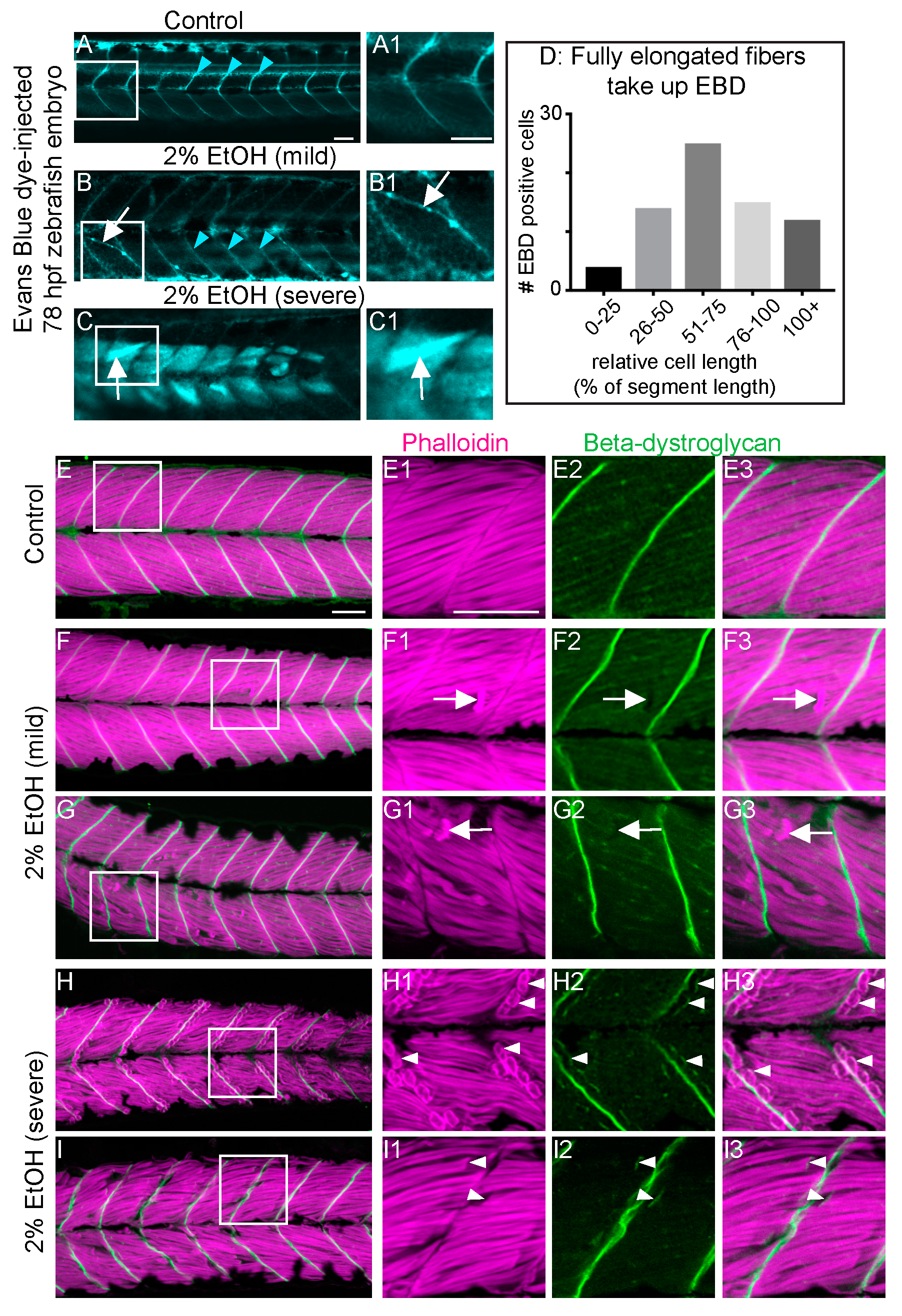
3.4. EtOH-Induced Fiber Detachments Result from Sarcolemmal Instability and Failure of Muscle End Attachments
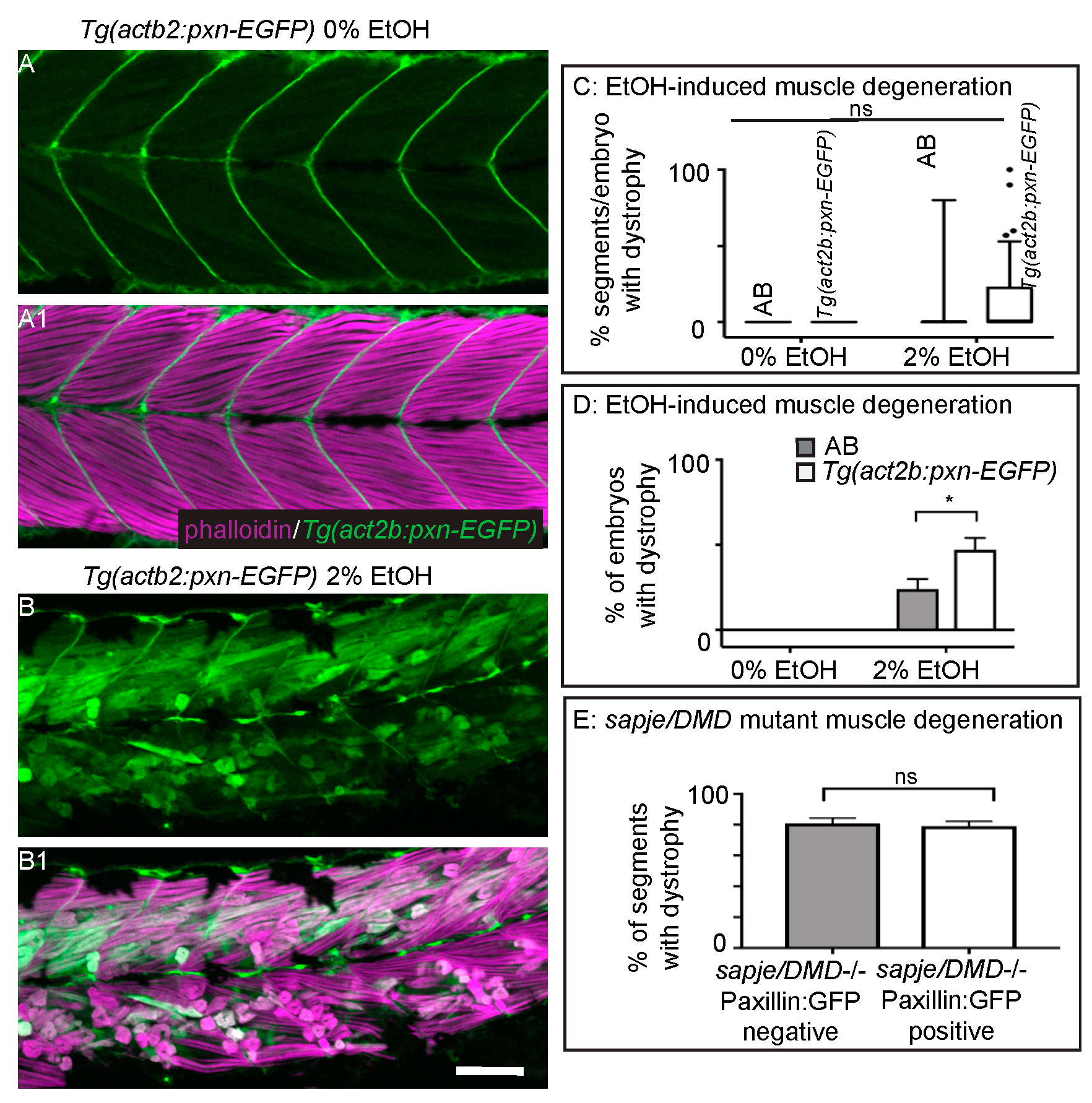
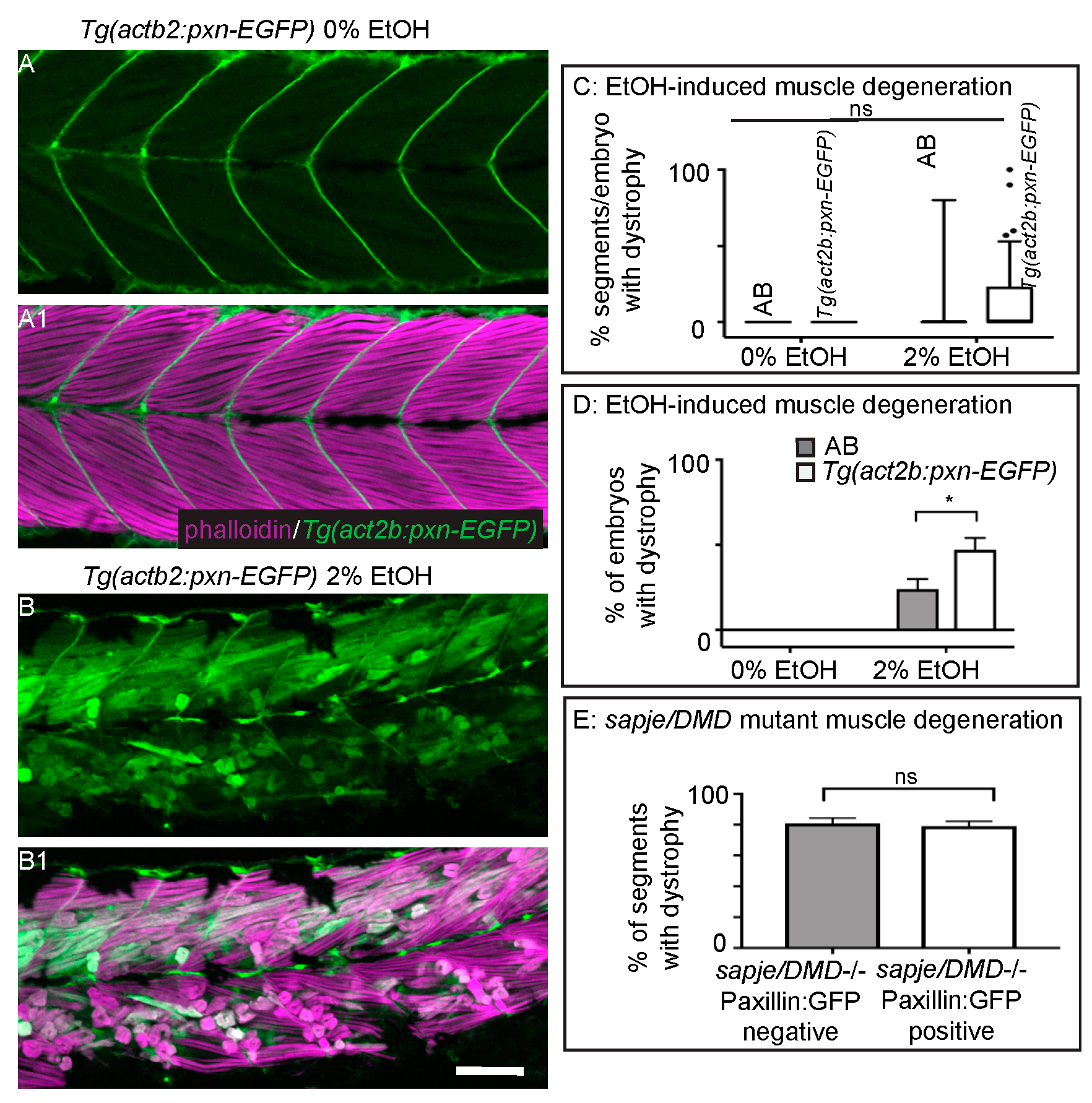
3.5. Overexpression of Paxillin Does Not Rescue Fiber Detachments Caused by Loss of Sarcolemmal Integrity
3.6. Combining the HDAC Inhibitor TSA with EtOH Is Lethal for Zebrafish Embryos
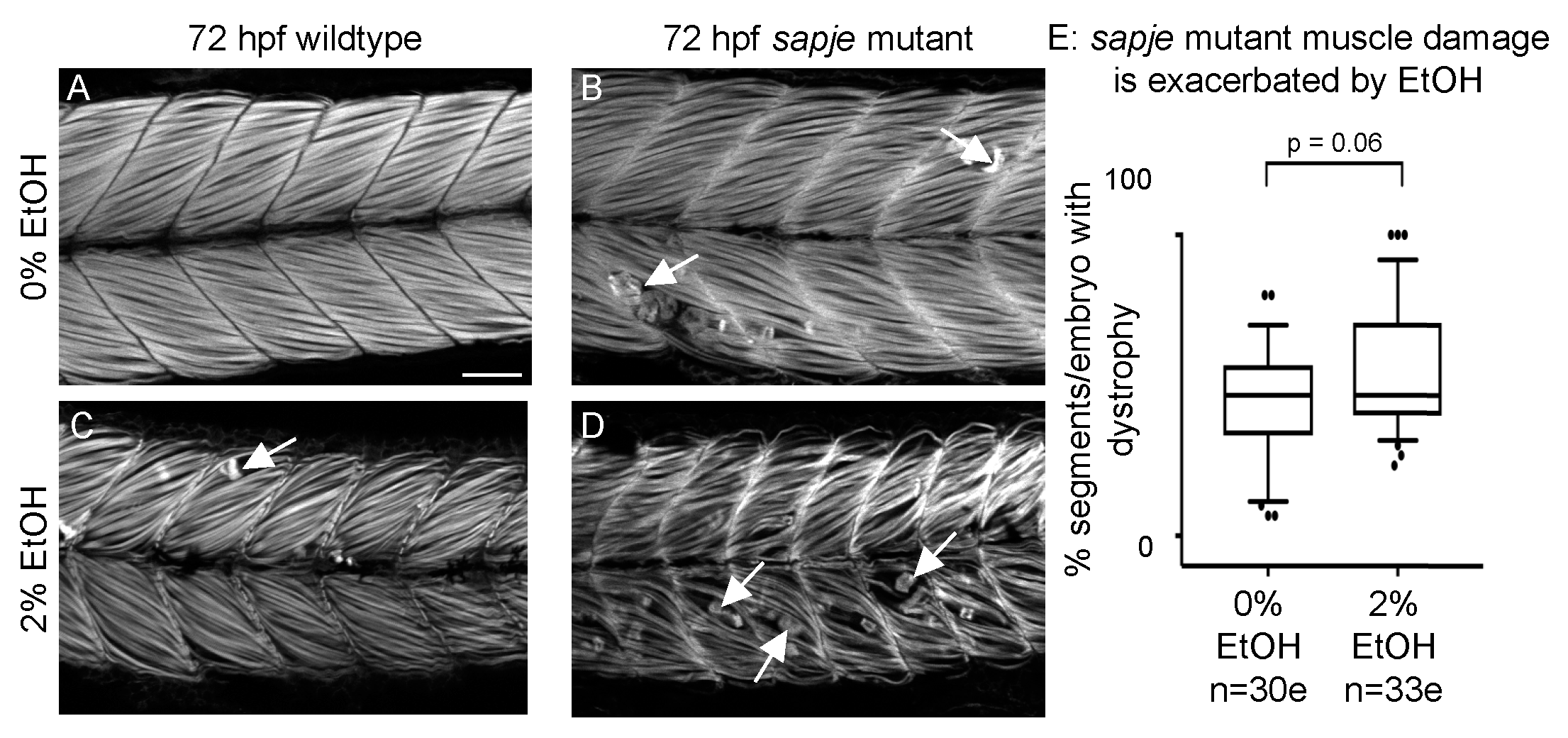
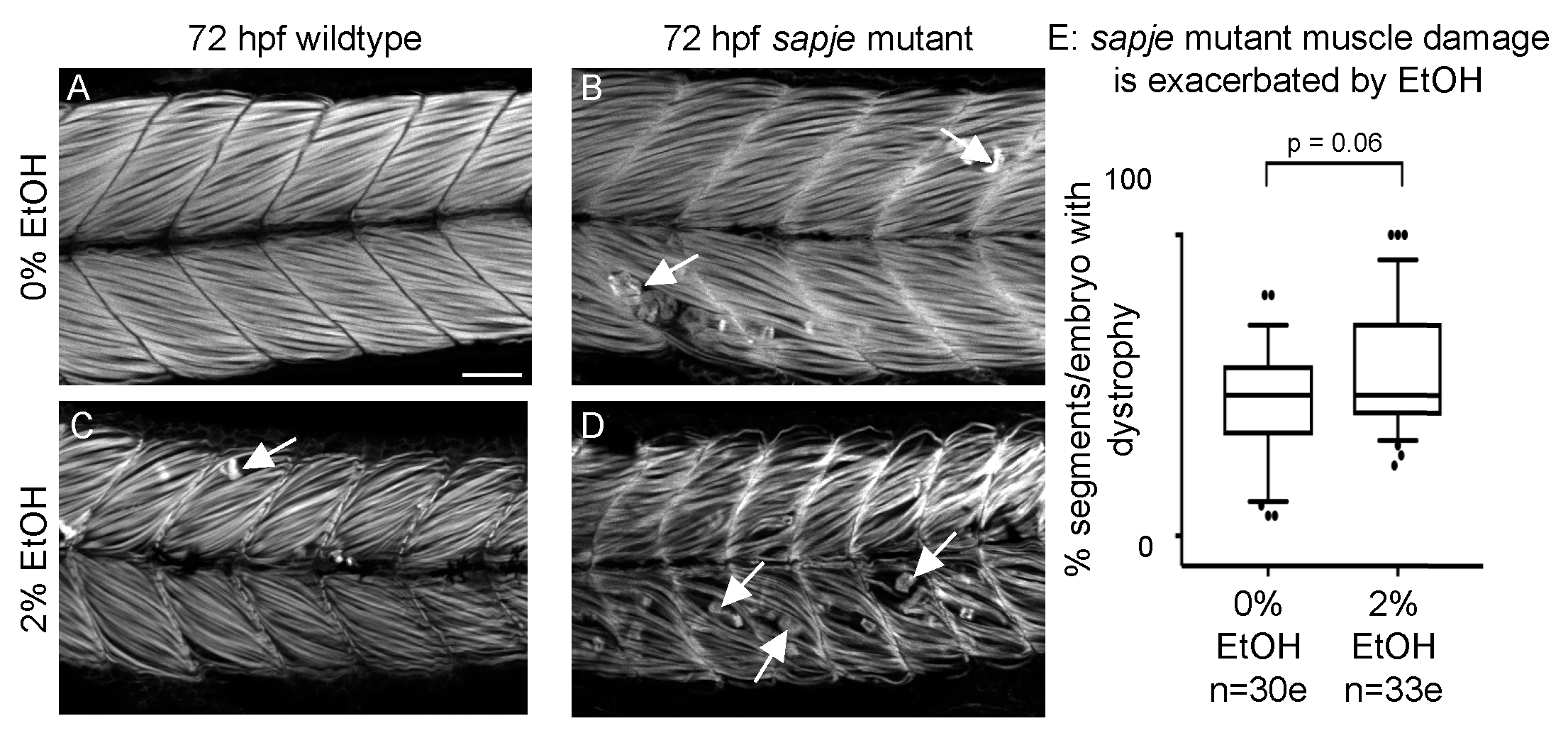
3.7. EtOH Exposure Exacerbates Muscle Degeneration in Sapje Mutant Zebrafish
4. Discussion
4.1. Variability in the Penetrance and Severity of Muscle Damage Caused by EtOH
4.2. EtOH Treatment Damages the Muscle Sarcolemma and Disrupts Muscle Fiber-MTJ Adhesion
4.3. Interaction of Environment and Muscle Disease
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Haberstick, B.C.; Young, S.E.; Zeiger, J.S.; Lessem, J.M.; Hewitt, J.K.; Hopfer, C.J. Prevalence and correlates of alcohol and cannabis use disorders in the United States: Results from the national longitudinal study of adolescent health. Drug Alcohol. Depend. 2014, 136, 158–161. [Google Scholar] [CrossRef] [PubMed]
- Rehm, J. The Risks Associated With Alcohol Use and Alcoholism. Alcohol. Res. Health 2011, 34, 135–143. [Google Scholar] [PubMed]
- Preedy, V.R.; Adachi, J.; Ueno, Y.; Ahmed, S.; Mantle, D.; Mullatti, N.; Rajendram, R.; Peters, T.J. Alcoholic skeletal muscle myopathy: Definitions, features, contribution of neuropathy, impact and diagnosis. Eur. J. Neurol. 2001, 8, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Memo, L.; Gnoato, E.; Caminiti, S.; Pichini, S.; Tarani, L. Fetal alcohol spectrum disorders and fetal alcohol syndrome: The state of the art and new diagnostic tools. Early Hum. Dev. 2013, 89 (Suppl. S1), S40–S43. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC). Fetal alcohol syndrome—Alaska, Arizona, Colorado, and New York, 1995–1997. Morb. Mortal. Wkly. Rep. 2002, 51, 433–435. [Google Scholar]
- Chaudhuri, J.D. Effect of a single dose of ethanol on developing skeletal muscle of chick embryos. Alcohol 2004, 34, 279–283. [Google Scholar] [CrossRef] [PubMed]
- David, P.; Subramaniam, K. Prenatal alcohol exposure and early postnatal changes in the developing nerve-muscle system. Birth Defects Res. A Clin. Mol. Teratol. 2005, 73, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Sylvain, N.J.; Brewster, D.L.; Ali, D.W. Zebrafish embryos exposed to alcohol undergo abnormal development of motor neurons and muscle fibers. Neurotoxicol. Teratol. 2010, 32, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Dlugos, C.A.; Rabin, R.A. Ethanol effects on three strains of zebrafish: Model system for genetic investigations. Pharmacol. Biochem. Behav. 2003, 74, 471–480. [Google Scholar] [CrossRef]
- Khayrullin, A.; Smith, L.; Mistry, D.; Dukes, A.; Pan, Y.A.; Hamrick, M.W. Chronic alcohol exposure induces muscle atrophy (myopathy) in zebrafish and alters the expression of microRNAs targeting the Notch pathway in skeletal muscle. Biochem. Biophys. Res. Commun. 2016, 479, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Gillies, A.R.; Lieber, R.L. Structure and Function of the Skeletal Muscle Extracellular Matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef] [PubMed]
- Bajanca, F.; Luz, M.; Duxson, M.J.; Thorsteinsdóttir, S. Integrins in the mouse myotome: Developmental changes and differences between the epaxial and hypaxial lineage. Dev. Dyn. 2004, 231, 402–415. [Google Scholar] [CrossRef] [PubMed]
- Bentzinger, C.F.; Wang, Y.X.; von Maltzahn, J.; Soleimani, V.D.; Yin, H.; Rudnicki, M.A. Fibronectin regulates Wnt7a signaling and satellite cell expansion. Cell Stem Cell 2013, 12, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.L.; Escaleira, R.C.; Jazenko, F.; Mermelstein, C.S. Cell adhesion in zebrafish myogenesis: Distribution of intermediate filaments, microfilaments, intracellular adhesion structures and extracellular matrix. Cell Motil. Cytoskeleton 2008, 65, 801–815. [Google Scholar] [CrossRef] [PubMed]
- Deries, M.; Gonçalves, A.B.; Vaz, R.; Martins, G.G.; Rodrigues, G.; Thorsteinsdóttir, S. Extracellular matrix remodeling accompanies axial muscle development and morphogenesis in the mouse. Dev. Dyn. 2012, 241, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Marques, L.; Thorsteinsdóttir, S. Dynamics of Akt activation during mouse embryo development: Distinct subcellular patterns distinguish proliferating versus differentiating cells. Differ. Res. Biol. Divers. 2013, 86, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Snow, C.J.; Henry, C.A. Dynamic formation of microenvironments at the myotendinous junction correlates with muscle fiber morphogenesis in zebrafish. Gene Expr. Patterns 2009, 9, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Gemballa, S.; Vogel, F. Spatial arrangement of white muscle fibers and myoseptal tendons in fishes. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2002, 133, 1013–1037. [Google Scholar] [CrossRef]
- Han, R.; Kanagawa, M.; Yoshida-Moriguchi, T.; Rader, E.P.; Ng, R.A.; Michele, D.E.; Muirhead, D.E.; Kunz, S.; Moore, S.A.; Iannaccone, S.T.; et al. Basal lamina strengthens cell membrane integrity via the laminin G domain-binding motif of alpha-dystroglycan. Proc. Natl. Acad. Sci. USA. 2009, 106, 12573–12579. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.P.; Brown, R.H.; Kunkel, L.M. Dystrophin: The protein product of the Duchenne muscular dystrophy locus. Cell 1987, 51, 919–928. [Google Scholar] [CrossRef]
- Tomé, F.M.; Evangelista, T.; Leclerc, A.; Sunada, Y.; Manole, E.; Estournet, B.; Barois, A.; Campbell, K.P.; Fardeau, M. Congenital muscular dystrophy with merosin deficiency. C. R. Acad. Sci. III 1994, 317, 351–357. [Google Scholar] [PubMed]
- Matsuda, R.; Nishikawa, A.; Tanaka, H. Visualization of dystrophic muscle fibers in mdx mouse by vital staining with Evans blue: Evidence of apoptosis in dystrophin-deficient muscle. J. Biochem. (Tokyo) 1995, 118, 959–964. [Google Scholar] [CrossRef] [PubMed]
- Straub, V.; Rafael, J.A.; Chamberlain, J.S.; Campbell, K.P. Animal models for muscular dystrophy show different patterns of sarcolemmal disruption. J. Cell Biol. 1997, 139, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.E.; Bryson-Richardson, R.J.; Berger, S.; Jacoby, A.S.; Cole, N.J.; Hollway, G.E.; Berger, J.; Currie, P.D. The zebrafish candyfloss mutant implicates extracellular matrix adhesion failure in laminin alpha2-deficient congenital muscular dystrophy. Proc. Natl. Acad. Sci. USA 2007, 104, 7092–7097. [Google Scholar] [CrossRef] [PubMed]
- Goody, M.; Jurczyszak, D.; Kim, C.; Henry, C. Influenza A Virus Infection Damages Zebrafish Skeletal Muscle and Exacerbates Disease in Zebrafish Modeling Duchenne Muscular Dystrophy. PLoS Curr. 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Goody, M.F.; Kelly, M.W.; Reynolds, C.J.; Khalil, A.; Crawford, B.D.; Henry, C.A. NAD+ biosynthesis ameliorates a zebrafish model of muscular dystrophy. PLoS Biol. 2012, 10, e1001409. [Google Scholar] [CrossRef] [PubMed]
- Kanther, M.; Sun, X.; Mühlbauer, M.; Mackey, L.C.; Flynn, E.J.; Bagnat, M.; Jobin, C.; Rawls, J.F. Microbial colonization induces dynamic temporal and spatial patterns of NF-κB activation in the zebrafish digestive tract. Gastroenterology 2011, 141, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Berghmans, S.; Murphey, R.D.; Wienholds, E.; Neuberg, D.; Kutok, J.L.; Fletcher, C.D.M.; Morris, J.P.; Liu, T.X.; Schulte-Merker, S.; Kanki, J.P.; et al. tp53 mutant zebrafish develop malignant peripheral nerve sheath tumors. Proc. Natl. Acad. Sci. USA. 2005, 102, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.; Currie, P.D. Identification of a zebrafish model of muscular dystrophy. Clin. Exp. Pharmacol. Physiol. 2004, 31, 537–540. [Google Scholar] [CrossRef] [PubMed]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef] [PubMed]
- Johnson, N.M.; Farr, G.H.; Maves, L. The HDAC Inhibitor TSA Ameliorates a Zebrafish Model of Duchenne Muscular Dystrophy. PLoS Curr. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Goody, M.F.; Kelly, M.W.; Lessard, K.N.; Khalil, A.; Henry, C.A. Nrk2b-mediated NAD+ production regulates cell adhesion and is required for muscle morphogenesis in vivo: Nrk2b and NAD+ in muscle morphogenesis. Dev. Biol. 2010, 344, 809–826. [Google Scholar] [CrossRef] [PubMed]
- Snow, C.J.; Goody, M.; Kelly, M.W.; Oster, E.C.; Jones, R.; Khalil, A.; Henry, C.A. Time-lapse analysis and mathematical characterization elucidate novel mechanisms underlying muscle morphogenesis. PLoS Genet. 2008, 4, e1000219. [Google Scholar] [CrossRef] [PubMed]
- Hollway, G.E.; Bryson-Richardson, R.J.; Berger, S.; Cole, N.J.; Hall, T.E.; Currie, P.D. Whole-somite rotation generates muscle progenitor cell compartments in the developing zebrafish embryo. Dev. Cell 2007, 12, 207–219. [Google Scholar] [CrossRef] [PubMed]
- Stellabotte, F.; Dobbs-McAuliffe, B.; Fernández, D.A.; Feng, X.; Devoto, S.H. Dynamic somite cell rearrangements lead to distinct waves of myotome growth. Dev. Camb. Engl. 2007, 134, 1253–1257. [Google Scholar] [CrossRef] [PubMed]
- Goody, M.F.; Sher, R.B.; Henry, C.A. Hanging on for the ride: Adhesion to the extracellular matrix mediates cellular responses in skeletal muscle morphogenesis and disease. Dev. Biol. 2015, 401, 75–91. [Google Scholar] [CrossRef] [PubMed]
- Baldelli, S.; Ciriolo, M.R. Altered S-nitrosylation of p53 is responsible for impaired antioxidant response in skeletal muscle during aging. Aging 2016, 8, 3450–3467. [Google Scholar] [CrossRef] [PubMed]
- Schwarzkopf, M.; Coletti, D.; Sassoon, D.; Marazzi, G. Muscle cachexia is regulated by a p53-PW1/Peg3-dependent pathway. Genes Dev. 2006, 20, 3440–3452. [Google Scholar] [CrossRef] [PubMed]
- Bassett, D.I.; Bryson-Richardson, R.J.; Daggett, D.F.; Gautier, P.; Keenan, D.G.; Currie, P.D. Dystrophin is required for the formation of stable muscle attachments in the zebrafish embryo. Dev. Camb. Engl. 2003, 130, 5851–5860. [Google Scholar] [CrossRef] [PubMed]
- Roostalu, U.; Strähle, U. In vivo imaging of molecular interactions at damaged sarcolemma. Dev. Cell 2012, 22, 515–529. [Google Scholar] [CrossRef] [PubMed]
- Doney, R.; Lucas, B.R.; Watkins, R.E.; Tsang, T.W.; Sauer, K.; Howat, P.; Latimer, J.; Fitzpatrick, J.P.; Oscar, J.; Carter, M.; et al. Visual-motor integration, visual perception, and fine motor coordination in a population of children with high levels of Fetal Alcohol Spectrum Disorder. Res. Dev. Disabil. 2016, 55, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Lovely, C.B.; Nobles, R.D.; Eberhart, J.K. Developmental age strengthens barriers to ethanol accumulation in zebrafish. Alcohol. Fayettev. N 2014, 48, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Reimers, M.J.; Flockton, A.R.; Tanguay, R.L. Ethanol- and acetaldehyde-mediated developmental toxicity in zebrafish. Neurotoxicol. Teratol. 2004, 26, 769–781. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Total Embryos | Embryos Alive at 72 hpf | %Survival |
|---|---|---|---|
| 0.66% DMSO | 48 | 48 | 100 |
| TSA (200 nM) with 0.66% DMSO | 52 | 52 | 100 |
| EtOH (2%) | 50 | 36 | 72 |
| EtOH (2%) with 0.66% DMSO | 27 | 20 | 74 |
| EtOH (2%) + TSA (200 nM) with 0.66% DMSO | 54 | 2 | 3.7 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coffey, E.C.; Pasquarella, M.E.; Goody, M.F.; Henry, C.A. Ethanol Exposure Causes Muscle Degeneration in Zebrafish. J. Dev. Biol. 2018, 6, 7. https://doi.org/10.3390/jdb6010007
Coffey EC, Pasquarella ME, Goody MF, Henry CA. Ethanol Exposure Causes Muscle Degeneration in Zebrafish. Journal of Developmental Biology. 2018; 6(1):7. https://doi.org/10.3390/jdb6010007
Chicago/Turabian StyleCoffey, Elizabeth C., Maggie E. Pasquarella, Michelle F. Goody, and Clarissa A. Henry. 2018. "Ethanol Exposure Causes Muscle Degeneration in Zebrafish" Journal of Developmental Biology 6, no. 1: 7. https://doi.org/10.3390/jdb6010007
APA StyleCoffey, E. C., Pasquarella, M. E., Goody, M. F., & Henry, C. A. (2018). Ethanol Exposure Causes Muscle Degeneration in Zebrafish. Journal of Developmental Biology, 6(1), 7. https://doi.org/10.3390/jdb6010007