N-Acyl Amino Acids: Metabolism, Molecular Targets, and Role in Biological Processes




Abstract
1. Introduction
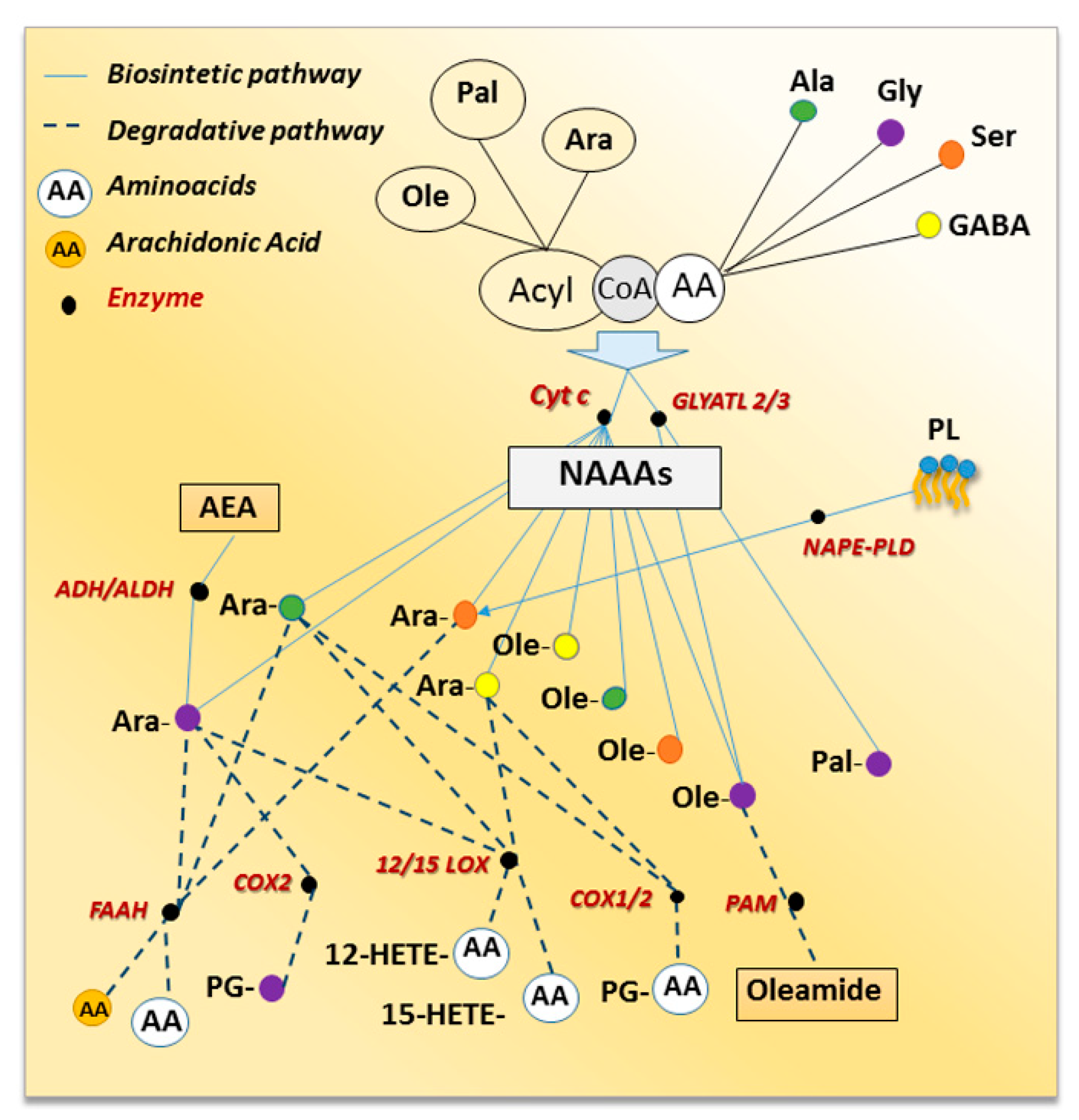
2. Metabolic Pathways for NAAAs
3. N-Acyl Glycines
4. N-Acyl-Serines
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kawai, Y.; Akagawa, K.; Yano, I. Lipoamino acids which are similar to bacterial endotoxin in both structure and biological activity related to physiological function. Adv. Exp. Med. Biol. 1990, 256, 159–162. [Google Scholar] [PubMed]
- Geiger, O.; González-Silva, N.; López-Lara, I.M.; Sohlenkamp, C. Amino acid-containing membrane lipids in bacteria. Prog. Lipid Res. 2010, 49, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Minarini, A.; Ferrari, S.; Galletti, M.; Giambalvo, N.; Perrone, D.; Rioli, G.; Galeazzi, G.M. N-acetylcysteine in the treatment of psychiatric disorders: Current status and future prospects. Expert Opin. Drug Metab. Toxicol. 2017, 13, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Erwan, E.; Chowdhury, V.S.; Ito, K.; Furuse, M. Lauroyl-L-aspartate decreased food intake and body temperature in neonatal chicks. Pharmacol. Biochem. Behav. 2013, 113, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Hershfield, J.R.; Pattabiraman, N.; Madhavarao, C.N.; Namboodiri, M.A. Mutational analysis of aspartoacylase: Implications for Canavan disease. Brain Res. 2007, 1148, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sheskin, T.; Hanus, L.; Slager, J.; Vogel, Z.; Mechoulam, R. Structural requirements for binding of anandamide-type compounds to the brain cannabinoid receptor. J. Med. Chem. 1997, 40, 659–667. [Google Scholar] [CrossRef]
- Huang, S.M.; Bisogno, T.; Petros, T.J.; Chang, S.Y.; Zavitsanos, P.A.; Zipkin, R.E.; Sivakumar, R.; Coop, A.; Maeda, D.Y.; De Petrocellis, L.; et al. Identification of a new class of molecules, the arachidonyl amino acids, and characterization of one member that inhibits pain. J. Biol. Chem. 2001, 276, 42639–42644. [Google Scholar] [CrossRef]
- Saghatelian, A.; Cravatt, B.F. Discovery metabolite profiling—Forging functional connections between the proteome and metabolome. Life Sci. 2005, 77, 1759–1766. [Google Scholar] [CrossRef]
- Tan, B.; O’Dell, D.K.; Yu, Y.W.; Monn, M.F.; Hughes, H.V.; Burstein, S.; Walker, J.M. Identification of endogenous acyl amino acids based on a targeted lipidomics approach. J. Lipid. Res. 2010, 51, 112–119. [Google Scholar] [CrossRef]
- Leishman, E.; Cornett, B.; Spork, K.; Straiker, A.; Mackie, K.; Bradshaw, H.B. Broad impact of deleting endogenous cannabinoid hydrolyzing enzymes and the CB1 cannabinoid receptor on the endogenous cannabinoid-related lipidome in eight regions of the mouse brain. Pharmacol. Res. 2016, 110, 159–172. [Google Scholar] [CrossRef]
- Tortoriello, G.; Rhodes, B.P.; Takacs, S.M.; Stuart, J.M.; Basnet, A.; Raboune, S.; Widlanski, T.S.; Doherty, P.; Harkany, T.; Bradshaw, H. Targeted lipidomics in Drosophila melanogaster identified novel 2monoacylglycerols and N-acyl amides. PLoS ONE 2013, 8, e67865. [Google Scholar] [CrossRef] [PubMed]
- Park, S.W.; Hah, J.H.; Oh, S.M.; Jeong, W.J.; Sung, M.W. 5-lipoxygenase mediates docosahexaenoyl ethanolamide and N-arachidonoyl-L-alanine-induced reactive oxygen species production and inhibition of proliferation of head and neck squamous cell carcinoma cells. BMC Cancer 2016, 16, 458–471. [Google Scholar] [CrossRef] [PubMed]
- Long, J.Z.; Svensson, K.J.; Bateman, L.A.; Lin, H.; Kamenecka, T.; Lokurkar, I.A.; Lou, J.; Rao, R.R.; Chang, M.R.; Jedrychowski, M.P.; et al. The secreted enzyme PM20D1 regulates lipidated amino acid uncouplers of mitochondria. Cell 2016, 166, 424–435. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.Y.; Lin, Q.; Niu, Y.Y.; Deng, K.M.; Zhang, J.H.; Lu, Y. Synthesis of Lipoamino acids and their activity against cerebral ischemic injury. Molecules 2009, 14, 4051–4064. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Li, J.; Zhou, Y.; Zheng, Q.; Ming, X.; Liu, S. N-Linoleyltyrosine Protects against Transient Cerebral Ischemia in Gerbil via CB2 Receptor Involvement in PI3K/Akt Signaling Pathway. Biol. Pharm. Bull. 2019, 42, 1867–1876. [Google Scholar] [CrossRef] [PubMed]
- Burstein, S.H. N-Acyl Amino Acids (Elmiric Acids): Endogenous Signaling Molecules with Therapeutic Potential. Mol. Pharmacol. 2018, 93, 228–238. [Google Scholar] [CrossRef]
- McCue, J.M.; Driscoll, W.J.; Mueller, G.P. Cytochrome c catalyzes the in vitro synthesis of arachidonoyl glycine. Biochem. Biophys. Res. Commun. 2008, 365, 322–327. [Google Scholar] [CrossRef]
- Mueller, G.P.; Driscoll, W.J. In vitro synthesis of oleoylglycine by cytochrome c points to a novel pathway for the production of lipid signaling molecules. J. Biol. Chem. 2007, 282, 22364–22369. [Google Scholar] [CrossRef]
- Waluk, D.P.; Schultz, N.; Hunt, M.C. Identification of glycine Nacyltransferase-like 2 (GLYATL2) as a transferase that produces N-acyl glycines in humans. FASEB J. 2010, 24, 2795–2803. [Google Scholar] [CrossRef]
- Jeffries, K.A.; Dempsey, D.R.; Farrell, E.K.; Anderson, R.L.; Garbade, G.J.; Gurina, T.S.; Gruhonjic, I.; Gunderson, C.A.; Merkler, D.J. Glycine N-acyltransferase-like 3 is responsible for long-chain N-acylglycine formation in N18TG2 cells. J. Lipid Res. 2016, 57, 781–790. [Google Scholar] [CrossRef]
- Burstein, S.H.; Rossetti, R.G.; Yagen, B.; Zurier, R.B. Oxidative metabolism of anandamide. Prostaglandins Other Lipid Mediat. 2000, 61, 29–41. [Google Scholar] [CrossRef]
- Bradshaw, H.B.; Rimmerman, N.; Hu, S.S.J.; Benton, V.M.; Stuart, J.M.; Masuda, K.; Cravatt, B.F.; O’Dell, D.K.; Walker, J.M. The endocannabinoid anandamide is a precursor for the signaling lipid N-arachidonoyl glycine by two distinct pathways. BMC Biochem. 2009, 10, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Aneetha, H.; O’Dell, D.K.; Tan, B.; Walker, J.M.; Hurley, T.D. Alcohol dehydrogenase-catalyzed in vitro oxidation of anandamide to N-arachidonoyl glycine, a lipid mediator: Synthesis of N-acyl glycinals. Bioorg. Med. Chem. Lett. 2009, 19, 237–241. [Google Scholar] [CrossRef] [PubMed]
- Leishman, E.; Mackie, K.; Luquet, S.; Bradshaw, H.B. Lipidomics profile of a NAPE-PLD KO mouse provides evidence of a broader role of this enzyme in lipid metabolism in the brain. Biochim. Biophys. Acta 2016, 1861, 491–500. [Google Scholar] [CrossRef]
- Cascio, M.G.; Minassi, A.; Ligresti, A.; Appendino, G.; Burstein, S.; Di Marzo, V. A structure-activity relationship study on N-arachidonoyl-amino acids as possible endogenous inhibitors of fatty acid amide hydrolase. Biochem. Biophys. Res. Commun. 2004, 314, 192–196. [Google Scholar] [CrossRef]
- Prusakiewicz, J.J.; Turman, M.V.; Vila, A.; Ball, H.L.; Al-Mestarihi, A.H.; Di Marzo, V.; Marnett, L. Oxidative metabolism of lipoamino acids and vanilloids by lipoxygenases and cyclooxygenases. Arch. Biochem. Biophys. 2007, 464, 260–268. [Google Scholar] [CrossRef]
- Prusakiewicz, J.J.; Kingsley, P.J.; Kozak, K.R.; Marnett, L.J. Selective oxygenation of N-arachidonylglycine by cyclooxygenase-2. Biochem. Biophys. Res. Commun. 2002, 296, 612–617. [Google Scholar] [CrossRef]
- Merkler, D.J.; Merkler, K.A.; Stern, W.; Fleming, F.F. Fatty acid amide biosynthesis: A possible new role for peptidylglycine α-amidating enzyme and acyl-coenzyme A: Glycine N-acyltransferase. Arch. Biochem. Biophys. 1996, 330, 430–434. [Google Scholar] [CrossRef]
- Merkler, D.J.; Chew, G.H.; Gee, A.J.; Merkler, K.A.; Sorondo, J.P.O.; Johnson, M.E. Oleic acid derived metabolites in mouse neuroblastoma cells. Biochemistry 2004, 43, 12667–12674. [Google Scholar] [CrossRef]
- Anderson, R.L.; Merkler, D.J. N-fatty acylglycines: Underappreciated endocannabinoid-like fatty acid amides? J. Biol. Nat. 2017, 8, 156–165. [Google Scholar]
- Console-Bram, L.; Brailoiu, E.; Brailoiu, G.C.; Sharir, H.; Abood, M.E. Activation of GPR18 by cannabinoid compounds: A tale of biased agonism. Br. J. Pharmacol. 2014, 171, 3908–3917. [Google Scholar] [CrossRef] [PubMed]
- Console-Bram, L.; Ciuciu, S.M.; Zhao, P.; Zipkin, R.E.; Brailoiu, E.; Abood, M.E. N-arachidonoyl glycine, another endogenous agonist of GPR55. Biochem. Biophys. Res. Commun. 2017, 490, 1389–1393. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.Y.; Yoon, J.M.; Moon, M.J.; Hwang, J.I.; Choe, H.; Lee, J.Y.; Kim, J.I.; Kim, S.; Rhim, H.; O’Dell, D.K.; et al. Identification of farnesyl pyrophosphate and N-arachidonylglycine as endogenous ligands for GPR92. J. Biol. Chem. 2008, 283, 21054–21064. [Google Scholar] [CrossRef] [PubMed]
- Takenouchi, R.; Inoue, K.; Kambe, Y.; Miyata, A. N-arachidonoyl glycine induces macrophage apoptosis via GPR18. Biochem. Biophys. Res. Commun. 2012, 418, 366–371. [Google Scholar] [CrossRef] [PubMed]
- McHugh, D.; Page, J.; Dunn, E.; Bradshaw, H.B. D(9)-Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharmacol. 2012, 165, 2414–2424. [Google Scholar] [CrossRef] [PubMed]
- Burstein, S.H.; McQuain, C.A.; Ross, A.H.; Salmonsen, R.A.; Zurier, R.E. Resolution of inflammation by N-arachidonoylglycine. J. Cell. Biochem. 2011, 112, 3227–3233. [Google Scholar] [CrossRef] [PubMed]
- Vuong, L.A.; Mitchell, V.A.; Vaughan, C.W. Actions of N-arachidonyl-glycine in a rat neuropathic pain model. Neuropharmacology 2008, 54, 189–193. [Google Scholar] [CrossRef]
- MacIntyre, J.; Dong, A.; Straiker, A.; Zhu, J.; Howlett, S.E.; Bagher, A.; Denovan-Wright, E.; Yu, D.Y.; Kelly, M.E. Cannabinoid and lipid-mediated vasorelaxation in retinal microvasculature. Eur. J. Pharmacol. 2014, 735, 105–114. [Google Scholar] [CrossRef]
- McHugh, D.; Roskowski, D.; Xie, S.; Bradshaw, H.B. Δ(9)-THC and N-arachidonoyl glycine regulate BV-2 microglial morphology and cytokine release plasticity: Implications for signaling at GPR18. Front. Pharmacol. 2014, 4, 162–171. [Google Scholar] [CrossRef]
- Grabiec, U.; Hohmann, T.; Ghadban, C.; Rothgänger, C.; Wong, D.; Antonietti, A.; Growth, T.; Mackie, K.; Dehghani, F. Protective Effect of N-Arachidonoyl Glycine-GPR18 Signaling after Excitotoxical Lesion in Murine Organotypic Hippocampal Slice Cultures. Int. J. Mol. Sci. 2019, 20, 1266. [Google Scholar] [CrossRef]
- Yin, H.; Chu, A.; Li, W.; Wang, B.; Shelton, F.; Otero, F.; Nguyen, D.G.; Caldwell, J.S.; Chen, Y.A. Lipid G protein-coupled receptor ligand identification using betaarrestin PathHunter assay. J. Biol. Chem. 2009, 284, 12328–12338. [Google Scholar] [CrossRef] [PubMed]
- Lu, V.B.; Puhl, H.L.; Ikeda, S.R. N-Arachidonyl glycine does not activate G protein-coupled receptor 18 signaling via canonical pathways. Mol. Pharmacol. 2013, 83, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Baur, R.; Gertsch, J.; Sigel, E. Do N-arachidonoyl-glycine (NA-glycine) and 2-arachidonoylglycerol (2-AG) share mode of action and the binding site on β2 subunit of GABAA receptors? Peer. J. 2013, 1, e149. [Google Scholar] [CrossRef] [PubMed]
- Yévenes, G.E.; Zeilhofer, H.U. Molecular sites for the positive allosteric modulation of glycine receptors by endocannabinoids. PLoS ONE 2011, 6, e23886. [Google Scholar]
- Wiles, A.L.; Pearlman, R.J.; Rosvall, M.; Aubrey, K.R.; Vandenberg, R.J. N-Arachidonyl-glycine inhibits the glycine transporter, GLYT2. J. Neurochem. 2006, 99, 781–786. [Google Scholar] [CrossRef]
- Burstein, S.H. The cannabinoid acids: Nonpsychoactive derivatives with therapeutic potential. Pharmacol. Ther. 1999, 82, 87–96. [Google Scholar] [CrossRef]
- Jeong, H.J.; Vandenberg, R.J.; Vaughan, C.W. N-arachidonyl-glycine modulates synaptic transmission in superficial dorsal horn. Br. J. Pharmacol. 2010, 161, 925–935. [Google Scholar] [CrossRef]
- Barbara, G.; Alou, A.; Nargeot, J.; Lory, P.; Eschalier, A.; Bourinet, E.; Chemin, J. T-type calcium channel inhibition underlies the analgesic effects of the endogenous lipoamino acids. J. Neurosci. 2009, 29, 13106–13114. [Google Scholar] [CrossRef]
- Ross, H.R.; Gilmore, A.J.; Connor, M. Inhibition of human recombinant T-type calcium channels by the endocannabinoid N-arachidonoyl dopamine. Br. J. Pharmacol. 2009, 156, 740–750. [Google Scholar] [CrossRef]
- Bradshaw, H.B.; Rimmerman, N.; Hu, S.S.J.; Burstein, S.; Walker, J.M. Novel endogenous N-acyl glycines identification and characterization. Vitam. Horm. 2009, 81, 191–205. [Google Scholar]
- Carland, J.E.; Mansfield, R.E.; Ryan, R.M.; Vandenberg, R.J. Oleoyl-L-carnitine inhibits glycine transport by GlyT2. Br. J. Pharmacol. 2013, 168, 891–902. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Xu, Q.; Shu, G.; Wang, L.; Gao, P.; Xi, Q.; Zhang, Y.; Jiang, Q.; Zhu, X. NOleoyl glycine, a lipoamino acid, stimulates adipogenesis associated with activation of CB1 receptor and Akt signaling pathway in 3T3-L1 adipocyte. Biochem. Biophys. Res. Commun. 2015, 466, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Takao, K.; Noguchi, K.; Hashimoto, Y.; Shirahata, A.; Sugita, Y. Synthesis and evaluation of fatty acid amides on the N-oleoylethanolamide-like activation of peroxisome proliferator activated receptor α. Chem. Pharm. Bull. 2015, 63, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, S.; Driscoll, W.J.; Elliot, B.M.; Faraday, M.M.; Grunberg, N.E.; Mueller, G.P. In vivo evidence that N-oleoylglycine acts independently of its conversion to oleamide. Prostaglandins. Other. Lipid. Mediat. 2006, 81, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhu, C.; Yang, L.; Wang, Z.; Wang, L.; Wang, S.; Gao, P.; Zhang, Y.; Jiang, Q.; Zhu, X.; et al. N-oleoylglycine-induced hyperphagia is associated with the activation of Agouti-Related Protein (AgRP) neuron by Cannabinoid Receptor Type 1 (CB1R). J. Agric. Food. Chem. 2017, 65, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Donvito, G.; Piscitelli, F.; Muldoon, P.; Jackson, A.; Vitale, R.M.; D’Aniello, E.; Giordano, C.; Ignatowska-Jankowska, B.M.; Mustafa, M.A.; Guida, F.; et al. N-Oleoyl-glycine reduces nicotine reward and withdrawal in mice. Neuropharmacology 2019, 148, 320–331. [Google Scholar] [CrossRef]
- Petrie, G.N.; Wills, K.L.; Piscitelli, F.; Smoum, R.; Limebeer, C.L.; Rock, E.M.; Humphrey, A.E.; Sheppard-Perkins, M.; Lichtman, A.H.; Mechoulam, R.; et al. Oleoyl glycine: Interference with the aversive effects of acute naloxone-precipitated MWD, but not morphine reward, in male Sprague-Dawley rats. Psychopharmacology (Berl) 2019, 236, 2623–2633. [Google Scholar] [CrossRef]
- Burstein, S.H.; Adams, J.K.; Bradshaw, H.B.; Fraioli, C.; Rossetti, R.G.; Salmonsen, R.A.; Shaw, J.W.; Walker, J.M.; Zipkin, R.E.; Zurier, R.B. Potential anti-inflammatory actions of the elmiric (lipoamino) acids. Bioorg. Med. Chem. 2007, 15, 3345–3355. [Google Scholar] [CrossRef]
- Milman, G.; Maor, Y.; Abu-Lafi, S.; Horowitz, M.; Gallily, R.; Batkai, S.; Mo, F.M.; Offertaler, L.; Pacher, P.; Kunos, G.; et al. N-arachidonoyl L-serine, an endocannabinoid-like brain constituent with vasodilatory properties. Proc. Natl. Acad. Sci. USA 2006, 103, 2428–2433. [Google Scholar] [CrossRef]
- Feldman, M.; Smoum, R.; Mechoulam, R.; Steinberg, D. Antimicrobial potential of endocannabinoid and endocannabinoid-like compounds against methicillin-resistant Staphylococcus aureus. Sci. Rep. 2018, 8, 17696–17705. [Google Scholar] [CrossRef]
- Cohen-Yeshurun, A.; Willner, D.; Trembovler, V.; Alexandrovich, A.; Mechoulam, R.; Shohami, E.; Leker, R.R. N-arachidonoyl-L-serine (AraS) possesses proneurogenic properties in vitro and in vivo after traumatic brain injury. J. Cereb. Blood Flow Metab. 2013, 33, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Kino, T.; Tomori, T.; Abutarboush, R.; Castri, P.; Chen, Y.; Lenz, F.A.; McCarron, R.M.; Spatz, M. Effect of N-arachidonoyl-l-serine on human cerebromicrovascular endothelium. Biochem. Biophys. Rep. 2016, 8, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Yeshurun, A.; Trembovler, V.; Alexandrovich, A.; Ryberg, E.; Greasley, P.J.; Mechoulam, R.; Shohami, E.; Leker, R.R. N-arachidonoyl-L-serine is neuroprotective after traumatic brain injury by reducing apoptosis. J. Cereb. Blood Flow Metab. 2011, 31, 1768–1777. [Google Scholar] [CrossRef] [PubMed]
- Mann, A.; Smoum, R.; Trembovler, V.; Alexandrovich, A.; Breuer, A.; Mechoulam, R.; Shohami, E. Palmitoyl Serine: An Endogenous Neuroprotective Endocannabinoid-Like Entity After Traumatic Brain Injury. J. Neuroimmune Pharmacol. 2015, 10, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Smoum, R.; Bar, A.; Tan, B.; Milman, G.; Attar-Namdar, M.; Ofek, O.; Stuart, J.M.; Bajayo, A.; Tam, J.; Kram, V.; et al. Oleoyl serine, an endogenous N-acyl amide, modulates bone remodeling and mass. Proc. Natl. Acad. Sci. USA 2010, 107, 17710–17715. [Google Scholar] [CrossRef]
- Baraghithy, S.; Smoum, R.; Drori, A.; Hadar, R.; Gammal, A.; Hirsch, S.; Attar-Namdar, M.; Nemirovski, A.; Gabet, Y.; Langer, Y.; et al. Magel2 Modulates Bone Remodeling and Mass in Prader-Willi Syndrome by Affecting Oleoyl Serine Levels and Activity. J. Bone Miner. Res. 2019, 34, 93–105. [Google Scholar] [CrossRef]
- Rimmerman, N.; Bradshaw, H.B.; Hughes, H.V.; Chen, J.S.; Hu, S.S.; McHugh, D.; Vefring, E.; Jahnsen, J.A.; Thompson, E.L.; Masuda, K.; et al. N-palmitoyl glycine, a novel endogenous lipid that acts as a modulator of calcium influx and nitric oxide production in sensory neurons. Mol. Pharmacol. 2008, 74, 213–224. [Google Scholar] [CrossRef]
- Liu, P.; Duan, J.; Wang, P.; Qian, D.; Guo, J.; Shang, E.; Su, S.; Tang, Y.; Tang, Z. Biomarkers of primary dysmenorrhea and herbal formula intervention: An exploratory metabonomics study of blood plasma and urine. Mol. Biosyst. 2013, 9, 77–87. [Google Scholar] [CrossRef]
- Jeffries, K.A.; Dempsey, D.R.; Behari, A.L.; Anderson, R.L.; Merkler, D.J. Drosophila melanogaster as a model system to study long-chain fatty acid amide metabolism. Febs. Lett. 2014, 588, 1596–1602. [Google Scholar] [CrossRef]
- Epand, R.F.; Infante, M.R.; Flanagan, T.D.; Epand, R.M. Properties of lipoamino acids incorporated into membrane bilayers. Biochim. Biophys. Acta 1998, 1373, 67–75. [Google Scholar] [CrossRef][Green Version]
- Bradshaw, H.B.; Raboune, S.; Hollis, J.L. Opportunistic activation of TRP receptors by endogenous lipids: Exploiting lipidomics to understand TRP receptor cellular communcation. Life Sci. 2013, 92, 404–409. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Baumann, J.; Kokabee, M.; Wong, J.; Balasubramaniyam, R.; Sun, Y.; Conklin, D.S. Global metabolite profiling analysis of lipotoxicity in HER2/neu-positive breast cancer cells. Oncotarget 2018, 9, 27133–27150. [Google Scholar] [CrossRef] [PubMed]
- Cazade, M.; Nuss, C.E.; Bidaud, I.; Renger, J.J.; Uebele, V.N.; Lory, P.; Chemin, J. Cross-modulation and molecular interaction at the Cav3.3 protein between the endogenous lipids and the T-type calcium channel antagonist TTA-A2. Mol. Pharmacol. 2014, 85, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Hanuš, L.; Shohami, E.; Bab, I.; Mechoulam, R. N-Acyl amino acids and their impact on biological processes. Biofactors 2014, 40, 381–388. [Google Scholar]
- Zhang, X.; Maor, Y.; Wang, J.F.; Kunos, G.; Groopman, J.E. Endocannabinoid-like N-arachidonoyl serine is a novel pro-angiogenic mediator. Br. J. Pharmacol. 2010, 160, 1583–1594. [Google Scholar] [CrossRef]
- Di Marzo, V. New approaches and challenges to targeting the endocannabinoid system. Nat. Rev. Drug Discov. 2018, 17, 623–639. [Google Scholar] [CrossRef]


{kind=link}
| Glycine | Serine | |
|---|---|---|
| N-arachidonoyl | Calcium mobilization [31] Cancer cell proliferation [58] Cell apoptosis and migration [34,35] Cell morphology [40] Inflammation [36] Intraocular pressure [38] Microglial-neuronal communication [39] Neuropathic pain [45] Thermal analgesia [46] | Angiogenesis [59] Anti-microbial effect [60] Cell proliferation [61] Cytoskeleton reorganization [62] Neuroprotection [61,63,64] |
| N-Oleoyl | Body and motor activities [54] Cancer cell proliferation [58] Energy homeostasis [52] Food intake [55] Nicotine addiction [56] | Bone metabolism [65,66] |
| N-Palmitoyl | Calcium mobilization [67] Primary dysmenorrhea [68] | Neuroprotection [64] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Battista, N.; Bari, M.; Bisogno, T. N-Acyl Amino Acids: Metabolism, Molecular Targets, and Role in Biological Processes. Biomolecules 2019, 9, 822. https://doi.org/10.3390/biom9120822
Battista N, Bari M, Bisogno T. N-Acyl Amino Acids: Metabolism, Molecular Targets, and Role in Biological Processes. Biomolecules. 2019; 9(12):822. https://doi.org/10.3390/biom9120822
Chicago/Turabian StyleBattista, Natalia, Monica Bari, and Tiziana Bisogno. 2019. "N-Acyl Amino Acids: Metabolism, Molecular Targets, and Role in Biological Processes" Biomolecules 9, no. 12: 822. https://doi.org/10.3390/biom9120822
APA StyleBattista, N., Bari, M., & Bisogno, T. (2019). N-Acyl Amino Acids: Metabolism, Molecular Targets, and Role in Biological Processes. Biomolecules, 9(12), 822. https://doi.org/10.3390/biom9120822