Effect of Coenzyme Q10 Supplementation on Testosterone


Abstract
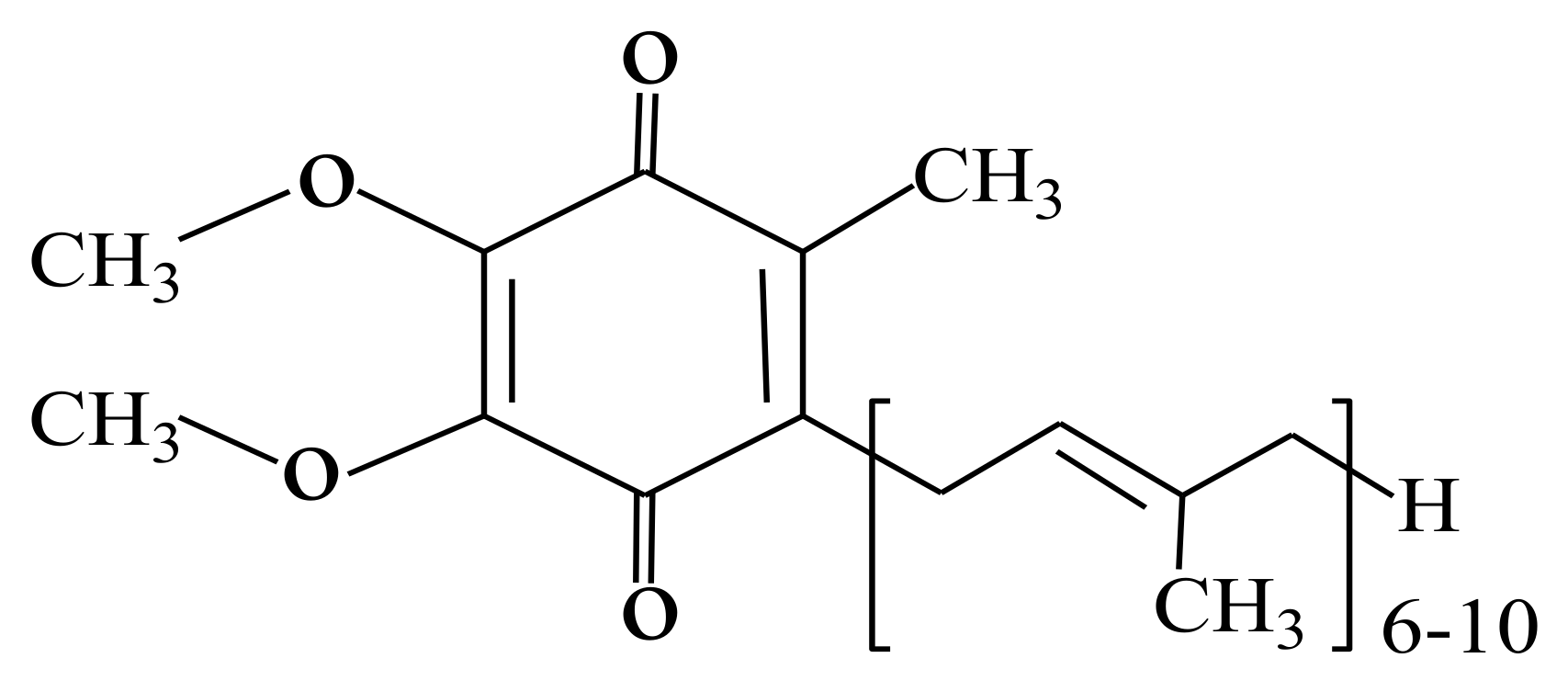
1. Introduction
2. Effect of Coenzyme Q10 Supplementation on Testosterone
3. Mechanistic Studies
4. Conclusions and Future Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- Sood, B.; Keenaghan, M. Coenzyme Q10; StatPearls Publishing: Treasure Island, FL, USA, 2018. [Google Scholar]
- Battino, M.; Ferri, E.; Gorini, A.; Villa, R.F.; Rodriguez Huertas, J.F.; Fiorella, P.; Genova, M.L.; Lenaz, G.; Marchetti, M. Natural distribution and occurrence of coenzyme Q homologues. Membr. Biochem. 1990, 9, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Cluis, C.P.; Burja, A.M.; Martin, V.J. Current prospects for the production of coenzyme Q10 in microbes. Trends Biotechnol. 2007, 25, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Varela-Lopez, A.; Giampieri, F.; Battino, M.; Quiles, J.L. Coenzyme Q and its role in the dietary therapy against aging. Molecules 2016, 21, 373. [Google Scholar] [CrossRef] [PubMed]
- Pham, T.; Loiselle, D.; Power, A.; Hickey, A.J. Mitochondrial inefficiencies and anoxic ATP hydrolysis capacities in diabetic rat heart. Am. J. Physiol. Cell Physiol. 2014, 307, C499–C507. [Google Scholar] [CrossRef] [PubMed]
- Merlo Pich, M.; Castagnoli, A.; Biondi, A.; Bernacchia, A.; Tazzari, P.L.; D’Aurelio, M.; Parenti Castelli, G.; Formiggini, G.; Conte, R.; Bovina, C.; et al. Ubiquinol and a coenzyme Q reducing system protect platelet mitochondrial function of transfusional buffy coats from oxidative stress. Free Radic. Res. 2002, 36, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Pepe, S.; Marasco, S.F.; Haas, S.J.; Sheeran, F.L.; Krum, H.; Rosenfeldt, F.L. Coenzyme Q10 in cardiovascular disease. Mitochondrion 2007, 7, S154–S167. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Fonarow, G.C.; Butler, J.; Ezekowitz, J.A.; Felker, G.M. Coenzyme Q10 and heart failure: A state-of-the-art review. Circ. Heart Fail. 2016, 9, e002639. [Google Scholar] [CrossRef]
- Zhang, S.Y.; Yang, K.L.; Zeng, L.T.; Wu, X.H.; Huang, H.Y. Effectiveness of coenzyme Q10 supplementation for type 2 diabetes mellitus: A systematic review and meta-analysis. Int. J. Endocrinol. 2018, 2018, 6484839. [Google Scholar] [CrossRef]
- Tarry-Adkins, J.L.; Fernandez-Twinn, D.S.; Madsen, R.; Chen, J.H.; Carpenter, A.; Hargreaves, I.P.; McConnell, J.M.; Ozanne, S.E. Coenzyme Q10 prevents insulin signaling dysregulation and inflammation prior to development of insulin resistance in male offspring of a rat model of poor maternal nutrition and accelerated postnatal growth. Endocrinology 2015, 156, 3528–3537. [Google Scholar] [CrossRef]
- Hernandez-Camacho, J.D.; Bernier, M.; Lopez-Lluch, G.; Navas, P. Coenzyme Q10 supplementation in aging and disease. Front. Physiol. 2018, 9, 44. [Google Scholar] [CrossRef]
- Mischley, L.K.; Lau, R.C.; Bennett, R.D. Role of diet and nutritional supplements in Parkinson’s disease progression. Oxid. Med. Cell. Longev. 2017, 2017, 6405278. [Google Scholar] [CrossRef] [PubMed]
- Yoritaka, A.; Kawajiri, S.; Yamamoto, Y.; Nakahara, T.; Ando, M.; Hashimoto, K.; Nagase, M.; Saito, Y.; Hattori, N. Randomized, double-blind, placebo-controlled pilot trial of reduced coenzyme Q10 for Parkinson’s disease. Parkinsonism Relat. Disord. 2015, 21, 911–916. [Google Scholar] [CrossRef] [PubMed]
- Shannon, K.M.; Fraint, A. Therapeutic advances in Huntington’s disease. Mov. Disord. 2015, 30, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.T.; Huang, Y.C.; Cheng, S.B.; Huang, Y.T.; Lin, P.T. Effects of coenzyme Q10 supplementation on antioxidant capacity and inflammation in hepatocellular carcinoma patients after surgery: A randomized, placebo-controlled trial. Nutr. J. 2016, 15, 85. [Google Scholar] [CrossRef]
- Liu, H.T.; Cheng, S.B.; Huang, Y.C.; Huang, Y.T.; Lin, P.T. Coenzyme Q10 and oxidative stress: Inflammation status in hepatocellular carcinoma patients after surgery. Nutrients 2017, 9, 29. [Google Scholar] [CrossRef]
- Jain, S.; Garg, T.; Kushwah, V.; Thanki, K.; Agrawal, A.K.; Dora, C.P. α-Tocopherol as functional excipient for resveratrol and coenzyme Q10-loaded SNEDDS for improved bioavailability and prophylaxis of breast cancer. J. Drug Target. 2017, 25, 554–565. [Google Scholar] [CrossRef]
- Gvozdjakova, A.; Kucharska, J.; Dubravicky, J.; Mojto, V.; Singh, R.B. Coenzyme Q10, α-tocopherol, and oxidative stress could be important metabolic biomarkers of male infertility. Dis. Markers 2015, 2015, 827941. [Google Scholar] [CrossRef]
- Hathcock, J.N.; Shao, A. Risk assessment for coenzyme Q10 (ubiquinone). Regul. Toxicol. Pharmacol. 2006, 45, 282–288. [Google Scholar] [CrossRef]
- McGarry, A.; McDermott, M.; Kieburtz, K.; de Blieck, E.A.; Beal, F.; Marder, K.; Ross, C.; Shoulson, I.; Gilbert, P.; Mallonee, W.M.; et al. A randomized, double-blind, placebo-controlled trial of coenzyme Q10 in Huntington disease. Neurology 2017, 88, 152–159. [Google Scholar] [CrossRef]
- Banihani, S.A. Ginger and testosterone. Biomolecules 2018, 8, 119. [Google Scholar] [CrossRef]
- Mooradian, A.D.; Morley, J.E.; Korenman, S.G. Biological actions of androgens. Endocr. Rev. 1987, 8, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Khaki, A.; Khaki, A.A.; Hajhosseini, L.; Golzar, F.S.; Ainehchi, N. The anti-oxidant effects of ginger and cinnamon on spermatogenesis dys-function of diabetes rats. Afr. J. Tradit. Complement. Altern. Med. 2014, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Paternostro, G. Longevity and testosterone. Nature 1994, 368, 408. [Google Scholar] [CrossRef] [PubMed]
- Audet-Walsh, E.; Yee, T.; Tam, I.S.; Giguere, V. Inverse regulation of DHT synthesis enzymes 5α-reductase types 1 and 2 by the androgen receptor in prostate cancer. Endocrinology 2017, 158, 1015–1021. [Google Scholar] [CrossRef] [PubMed]
- Dehghan, F.; Muniandy, S.; Yusof, A.; Salleh, N. Testosterone reduces knee passive range of motion and expression of relaxin receptor isoforms via 5α-dihydrotestosterone and androgen receptor binding. Int. J. Mol. Sci. 2014, 15, 4619–4634. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.M.; Akhtar, S.; Sellers, D.J.; Muraleedharan, V.; Channer, K.S.; Jones, T.H. Testosterone differentially regulates targets of lipid and glucose metabolism in liver, muscle and adipose tissues of the testicular feminised mouse. Endocrine 2016, 54, 504–515. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, A.; Kaufman, J.M.; Goemaere, S.; van Pottelberg, I. Estradiol in elderly men. Aging Male 2002, 5, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Travison, T.G.; Morley, J.E.; Araujo, A.B.; O’Donnell, A.B.; McKinlay, J.B. The relationship between libido and testosterone levels in aging men. J. Clin. Endocrinol. Metab. 2006, 91, 2509–2513. [Google Scholar] [CrossRef]
- Moffat, S.D. Effects of testosterone on cognitive and brain aging in elderly men. Ann. N. Y. Acad. Sci. 2005, 1055, 80–92. [Google Scholar] [CrossRef]
- Bassil, N.; Alkaade, S.; Morley, J.E. The benefits and risks of testosterone replacement therapy: A review. Ther. Clin. Risk Manag. 2009, 5, 427–448. [Google Scholar]
- Karakas, M.; Schafer, S.; Appelbaum, S.; Ojeda, F.; Kuulasmaa, K.; Bruckmann, B.; Berisha, F.; Schulte-Steinberg, B.; Jousilahti, P.; Blankenberg, S.; et al. Testosterone levels and type 2 diabetes—No correlation with age, differential predictive value in men and women. Biomolecules 2018, 8, 76. [Google Scholar] [CrossRef] [PubMed]
- Tuck, S.P.; Francis, R.M. Testosterone, bone and osteoporosis. Front. Horm Res. 2009, 37, 123–132. [Google Scholar] [PubMed]
- Yarrow, J.F.; Phillips, E.G.; Conover, C.F.; Bassett, T.E.; Chen, C.; Teurlings, T.; Vasconez, A.; Alerte, J.; Prock, H.; Jiron, J.M.; et al. Testosterone plus finasteride prevents bone loss without prostate growth in a rodent spinal cord injury model. J. Neurotrauma 2017, 34, 2972–2981. [Google Scholar] [CrossRef] [PubMed]
- El-Migdadi, F.; Banihani, I.; Banihani, S.A. Clinico-hormonal correlation of oligospermic patients in the below sea level environment (Jordan Valley). Neuro Endocrinol. Lett. 2005, 26, 13–18. [Google Scholar] [PubMed]
- Kelly, D.M.; Jones, T.H. Testosterone: A metabolic hormone in health and disease. J. Endocrinol. 2013, 217, R25–R45. [Google Scholar] [CrossRef] [PubMed]
- Hoenjet, K.M.; Dagnelie, P.C.; Delaere, K.P.; Wijckmans, N.E.; Zambon, J.V.; Oosterhof, G.O. Effect of a nutritional supplement containing vitamin E, selenium, vitamin C and coenzyme Q10 on serum PSA in patients with hormonally untreated carcinoma of the prostate: A randomised placebo-controlled study. Eur. Urol. 2005, 47, 433–439, discussion 439–440. [Google Scholar] [CrossRef]
- Safarinejad, M.R. Efficacy of coenzyme Q10 on semen parameters, sperm function and reproductive hormones in infertile men. J. Urol. 2009, 182, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Safarinejad, M.R. The effect of coenzyme Q10 supplementation on partner pregnancy rate in infertile men with idiopathic oligoasthenoteratozoospermia: An open-label prospective study. Int. Urol. Nephrol. 2012, 44, 689–700. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.F.; Xing, Y.; Wu, C.Y.; Liu, R.Z.; Wang, X.Y.; Xing, J.P. Tamoxifen combined with coenzyme Q10 for idiopathic oligoasthenospermia. Zhonghua Nan Ke Xue 2011, 17, 615–618. (In Chinese) [Google Scholar]
- Izadi, A.; Ebrahimi, S.; Shirzai, S.; Taghizadeh, S.; Parizad, M.; Farzadi, L.; Pourghassem Gargari, B. Hormonal and metabolic effects of coenzyme Q10 and/or vitamin E in patients with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2018. [Google Scholar] [CrossRef]
- Souri, Z.; Bidmeshki-Pour, A.; Karimi, I. Effect of coenzyme Q10 supplementation on p53 tumor suppressor gene expression in mouse model of andropause. Biharean Biol. 2017, 11, 53–56. [Google Scholar]
- Souri, Z.; Bidmeshki-Pour, A.; Karimi, I. Influence of coenzyme Q10 supplementation on skin follicle characteristics in gonadectomized mice. Biharean Biol. 2016, 10, 12–15. [Google Scholar]
- Vazir, N.; Adibmoradi, M.; Tajik, P.; Rezaian, M.; Babapour, V.; Baghcheghi, Y. A morphometrical study of testis and histometrical study of seminiferous tubules in coenzyme Q10 fed ostriches. J. Vet. Res. 2013, 68, 349–357. [Google Scholar]
- Fouad, A.A.; Al-Sultan, A.I.; Yacoubi, M.T. Coenzyme Q10 counteracts testicular injury induced by sodium arsenite in rats. Eur. J. Pharmacol. 2011, 655, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Ghanbarzadeh, S.; Garjani, A.; Ziaee, M.; Khorrami, A. Effects of l-carnitine and coenzyme Q10 on impaired spermatogenesis caused by isoproterenol in male rats. Drug Res. (Stuttg.) 2014, 64, 449–453. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, N.S.; Arafa, M.H.; Atteia, H.H. Coenzyme Q10 and fish oil synergistically alleviate aluminum chloride-induced suppression of testicular steroidogenesis and antioxidant defense. Free Radic. Res. 2015, 49, 1319–1334. [Google Scholar] [CrossRef] [PubMed]
- Roy, J.; Fournier, M.A.; Maltais, R.; Kenmogne, L.C.; Poirier, D. In vitro and in vivo evaluation of a 3β-androsterone derivative as inhibitor of 17β-hydroxysteroid dehydrogenase type 3. J. Steroid Biochem. Mol. Biol. 2014, 141, 44–51. [Google Scholar] [CrossRef]
- Griswold, M.D. The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol. 1998, 9, 411–416. [Google Scholar] [CrossRef]
- Lafuente, R.; Gonzalez-Comadran, M.; Sola, I.; Lopez, G.; Brassesco, M.; Carreras, R.; Checa, M.A. Coenzyme Q10 and male infertility: A meta-analysis. J. Assist. Reprod. Genet. 2013, 30, 1147–1156. [Google Scholar] [CrossRef]
- Al-Amin, M.M.; Reza, H.M.; Saadi, H.M.; Mahmud, W.; Ibrahim, A.A.; Alam, M.M.; Kabir, N.; Saifullah, A.R.; Tropa, S.T.; Quddus, A.H. Astaxanthin ameliorates aluminum chloride-induced spatial memory impairment and neuronal oxidative stress in mice. Eur. J. Pharmacol. 2016, 777, 60–69. [Google Scholar] [CrossRef]
- Banihani, S.A. Vitamin B12 and semen quality. Biomolecules 2017, 7, 42. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Kshetrimayum, C.; Sadhu, H.G.; Kumar, S. Arsenic-induced oxidative stress, cholinesterase activity in the brain of swiss albino mice, and its amelioration by antioxidants vitamin E and coenzyme Q10. Environ. Sci. Pollut. Res. Int. 2018, 25, 23946–23953. [Google Scholar] [CrossRef] [PubMed]
- Bergquist, E.R.; Fischer, R.J.; Sugden, K.D.; Martin, B.D. Inhibition by methylated organo-arsenicals of the respiratory 2-oxo-acid dehydrogenases. J. Organomet. Chem. 2009, 694, 973–980. [Google Scholar] [CrossRef] [PubMed]
- Pant, N.; Murthy, R.C.; Srivastava, S.P. Male reproductive toxicity of sodium arsenite in mice. Hum. Exp. Toxicol. 2004, 23, 399–403. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Banihani, S.A. Role of uric acid in semen. Biomolecules 2018, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Chiou, T.J.; Chu, S.T.; Tzeng, W.F.; Huang, Y.C.; Liao, C.J. Arsenic trioxide impairs spermatogenesis via reducing gene expression levels in testosterone synthesis pathway. Chem. Res. Toxicol. 2008, 21, 1562–1569. [Google Scholar] [CrossRef] [PubMed]
- Fetherolf, M.M.; Boyd, S.D.; Taylor, A.B.; Kim, H.J.; Wohlschlegel, J.A.; Blackburn, N.J.; Hart, P.J.; Winge, D.R.; Winkler, D.D. Copper-zinc superoxide dismutase is activated through a sulfenic acid intermediate at a copper ion entry site. J. Biol. Chem. 2017, 292, 12025–12040. [Google Scholar] [CrossRef]
- Salem, K.; McCormick, M.L.; Wendlandt, E.; Zhan, F.; Goel, A. Copper-zinc superoxide dismutase-mediated redox regulation of bortezomib resistance in multiple myeloma. Redox. Biol. 2015, 4, 23–33. [Google Scholar] [CrossRef]
- Paunovic, M.G.; Matic, M.M.; Ognjanovic, B.I.; Saicic, Z.S. Antioxidative and haematoprotective activity of coenzyme Q10 and vitamin E against cadmium-induced oxidative stress in Wistar rats. Toxicol. Ind. Health 2017, 33, 746–756. [Google Scholar] [CrossRef]
- Knott, A.; Achterberg, V.; Smuda, C.; Mielke, H.; Sperling, G.; Dunckelmann, K.; Vogelsang, A.; Kruger, A.; Schwengler, H.; Behtash, M.; et al. Topical treatment with coenzyme Q10-containing formulas improves skin’s Q10 level and provides antioxidative effects. Biofactors 2015, 41, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Hu, C.; Xue, Y. In vitro evaluation of chitosan-coated liposome containing both coenzyme Q10 and α-lipoic acid: Cytotoxicity, antioxidant activity, and antimicrobial activity. J. Cosmet. Dermatol. 2018, 17, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Du, J.; Lian, Y.; Zhang, Y.; Li, X.; Liu, Y.; Zou, L.; Wu, T. Protective effects of coenzyme Q10 against hydrogen peroxide-induced oxidative stress in PC12 cell: The role of Nrf2 and antioxidant enzymes. Cell. Mol. Neurobiol. 2016, 36, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Pala, R.; Orhan, C.; Tuzcu, M.; Sahin, N.; Ali, S.; Cinar, V.; Atalay, M.; Sahin, K. Coenzyme Q10 supplementation modulates NFκb and Nrf2 pathways in exercise training. J. Sports Sci. Med. 2016, 15, 196–203. [Google Scholar]
- Rahmani, A.; Abangah, G.; Moradkhani, A.; Hafezi Ahmadi, M.R.; Asadollahi, K. Coenzyme Q10 in combination with triple therapy regimens ameliorates oxidative stress and lipid peroxidation in chronic gastritis associated with H. pylori infection. J. Clin. Pharmacol. 2015, 55, 842–847. [Google Scholar] [CrossRef]
- Ahmadvand, H.; Ghasemi Dehnoo, M.; Dehghani, A.; Bagheri, S.; Cheraghi, R.A. Serum paraoxonase 1 status and its association with atherogenic indexes in gentamicin-induced nephrotoxicity in rats treated with coenzyme Q10. Ren. Fail. 2014, 36, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Orlando, P.; Silvestri, S.; Galeazzi, R.; Antonicelli, R.; Marcheggiani, F.; Cirilli, I.; Bacchetti, T.; Tiano, L. Effect of ubiquinol supplementation on biochemical and oxidative stress indexes after intense exercise in young athletes. Redox Rep. 2018, 23, 136–145. [Google Scholar] [CrossRef]
- Palmeira, C.M.; Santos, D.L.; Seica, R.; Moreno, A.J.; Santos, M.S. Enhanced mitochondrial testicular antioxidant capacity in Goto-Kakizaki diabetic rats: Role of coenzyme Q. Am. J. Physiol. Cell Physiol. 2001, 281, C1023–C1028. [Google Scholar] [CrossRef]


{kind=link}
| Source | Dose (Mode of Treatment) | Duration | Study Population | Effect on Testosterone | Ref. |
|---|---|---|---|---|---|
| Coenzyme Q10 (Bio-Quinon Q10) | 200 mg day−1 (orally) | 21 weeks | Patients with hormonally untreated carcinoma of the prostate | (±) | [37] |
| Coenzyme Q10 (Kaneka, Osaka, Japan) | 300 mg day−1 (orally) | 26 weeks | Infertile men | (±) | [38] |
| Coenzyme Q10 (Nutri Q10, Nutri Century, Toronto, ON, Canada) | 900 mg day−1 (orally) | 12 months | Infertile men with idiopathic oligoasthenoteratozoospermia | (±) | [39] |
| Coenzyme Q10 | Therapeutic dose (orally) | 3 and 6 months | Patients with idiopathic oligoasthenospermia | (±) | [40] |
| Coenzyme Q10 (Nature Made Pharmaceutical Company, Mission Hills, CA, USA) | 200 mg day−1 (orally) | 8 weeks | Patients with polycystic ovary syndrome | (−) | [41] |
| Coenzyme Q10 (Nutralife Co., Richmond Hill, Canada) | 125, 250, and 500 mg kg−1 day−1 (orally) | 96 days | Bilateral orchidectomized male mice | (±) | [42] |
| Coenzyme Q10 (Nutralife Co.) | 125 and 250 mg kg−1 day−1 (orally) | 96 days | Gonadectomized male mice | (−) | [43] |
| Coenzyme Q10 | 10 and 20 mg kg−1 day−1 (orally) | 2 months | Male ostriches | (±) | [44] |
| Coenzyme Q10 (Sigma Chemical Company, St. Louis, MO, USA) | 10 mg kg−1 day−1 (intraperitoneally) | 5 days | Male rats with sodium arsenite-induced reproductive toxicity | (+) | [45] |
| Coenzyme Q10 | 10 mg kg−1 day−1 (orally) | 20 days | Male rats with isoproterenol-induced reproductive toxicity | (+) | [46] |
| Coenzyme Q10 (Arab Co. for Pharmaceuticals & Medicinal Plants, Cairo, Egypt) | 10 mg kg−1 day−1 (orally) | 10 weeks | Male rats with aluminum chloride-induced reproductive toxicity | (+) | [47] |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banihani, S.A. Effect of Coenzyme Q10 Supplementation on Testosterone. Biomolecules 2018, 8, 172. https://doi.org/10.3390/biom8040172
Banihani SA. Effect of Coenzyme Q10 Supplementation on Testosterone. Biomolecules. 2018; 8(4):172. https://doi.org/10.3390/biom8040172
Chicago/Turabian StyleBanihani, Saleem Ali. 2018. "Effect of Coenzyme Q10 Supplementation on Testosterone" Biomolecules 8, no. 4: 172. https://doi.org/10.3390/biom8040172
APA StyleBanihani, S. A. (2018). Effect of Coenzyme Q10 Supplementation on Testosterone. Biomolecules, 8(4), 172. https://doi.org/10.3390/biom8040172