Synergistic Effect of Combinations Containing EDTA and the Antimicrobial Peptide AA230, an Arenicin-3 Derivative, on Gram-Negative Bacteria


Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Strains
2.3. Preparation of Inoculum
2.4. MIC and MBC
2.5. Checkerboard Titration
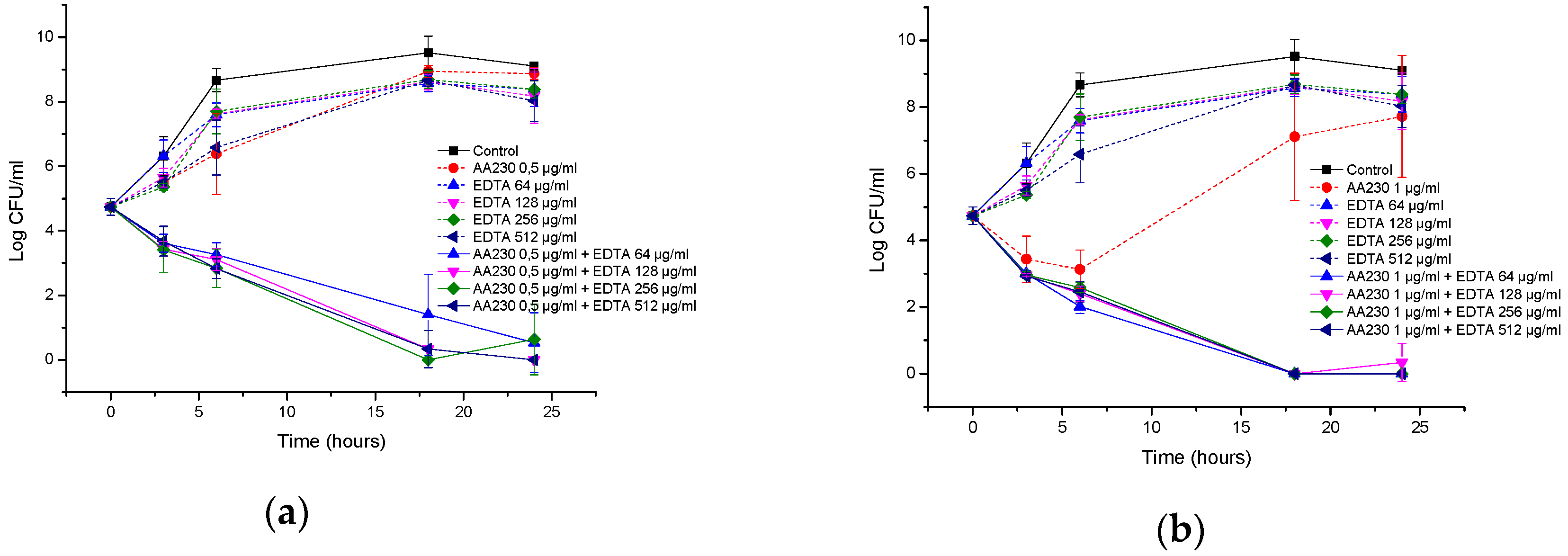
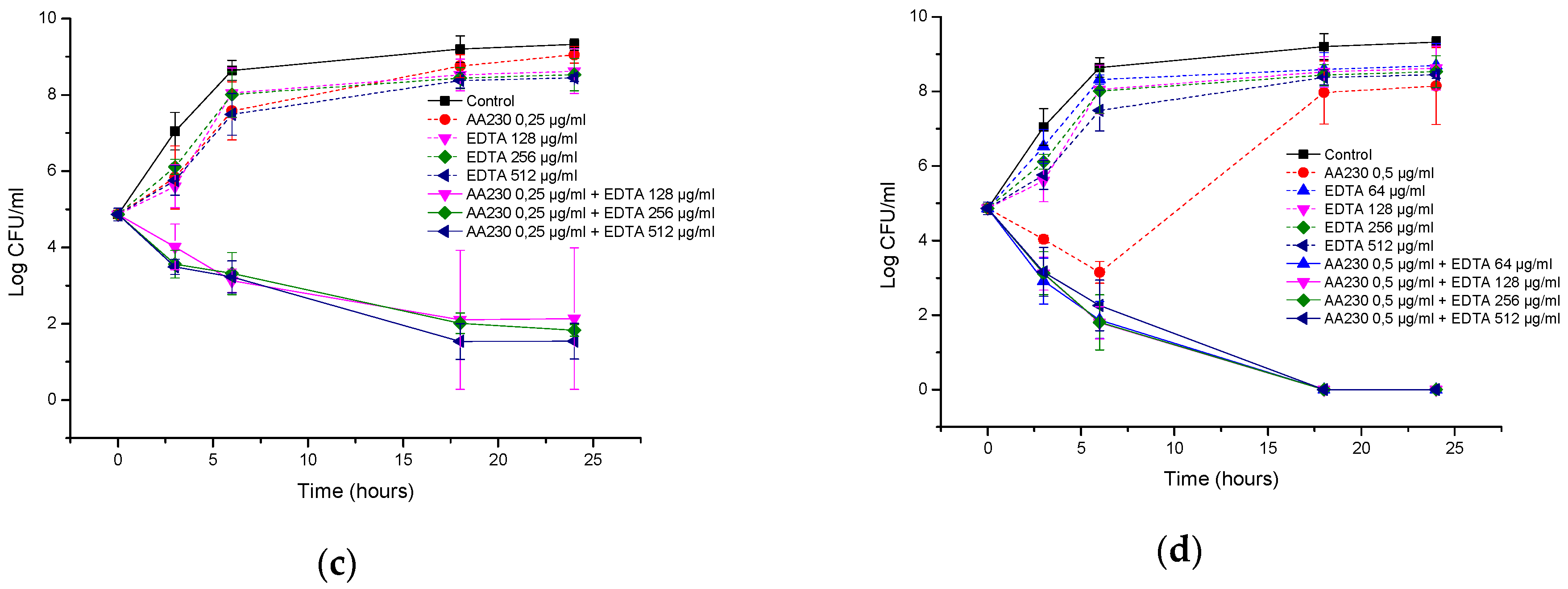
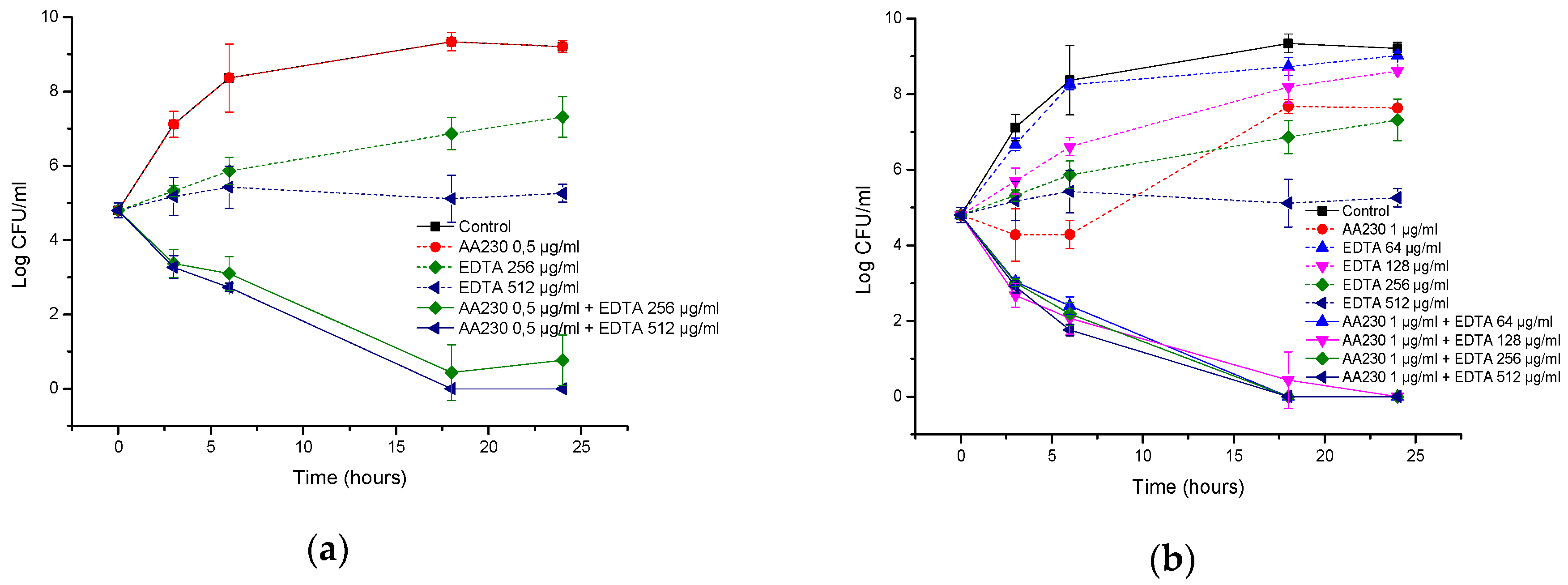
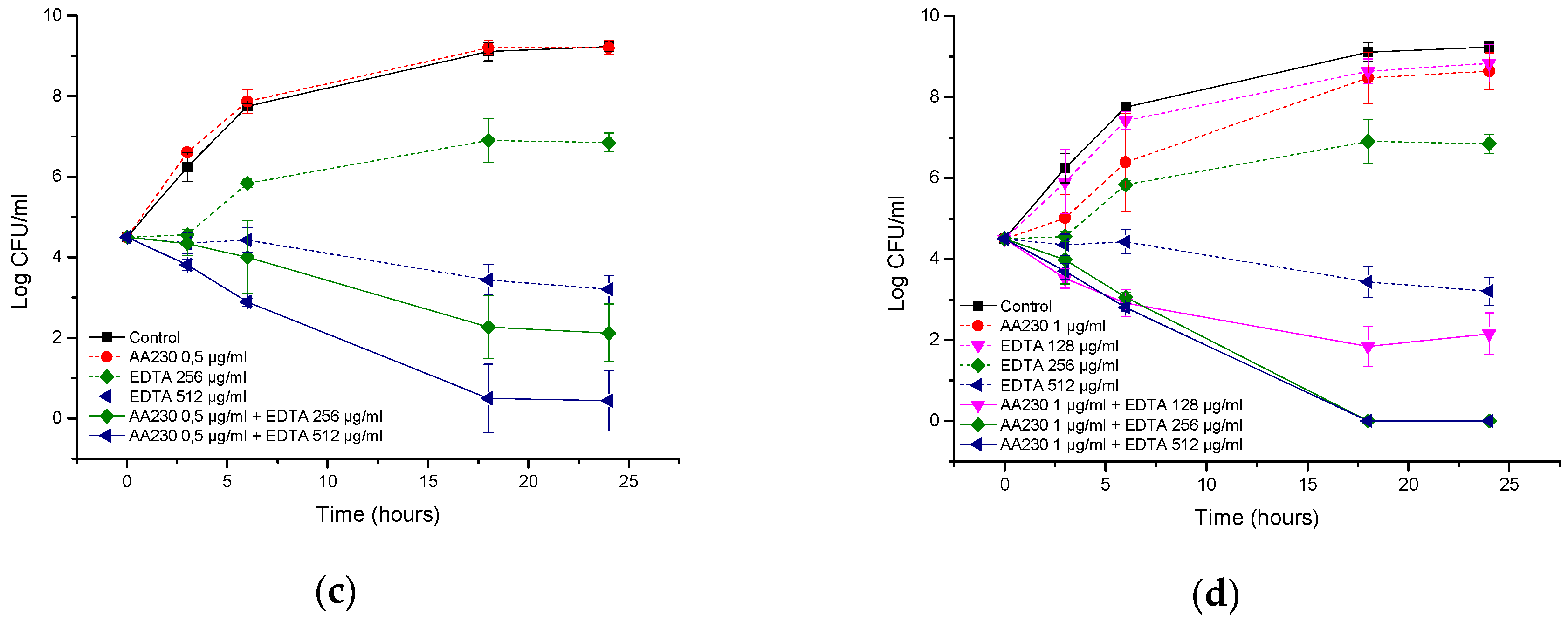
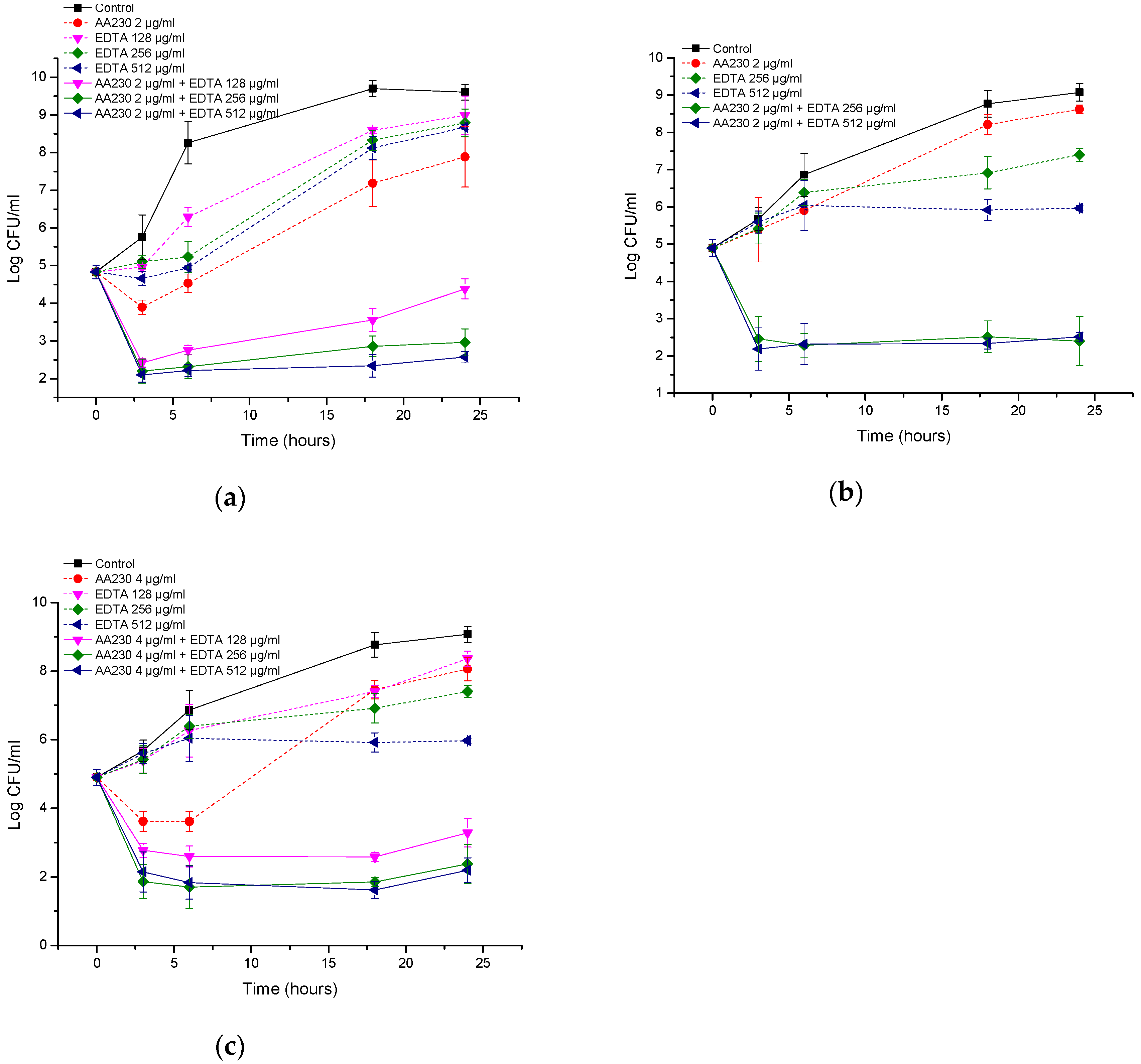
2.6. Time-Kill Studies
3. Results
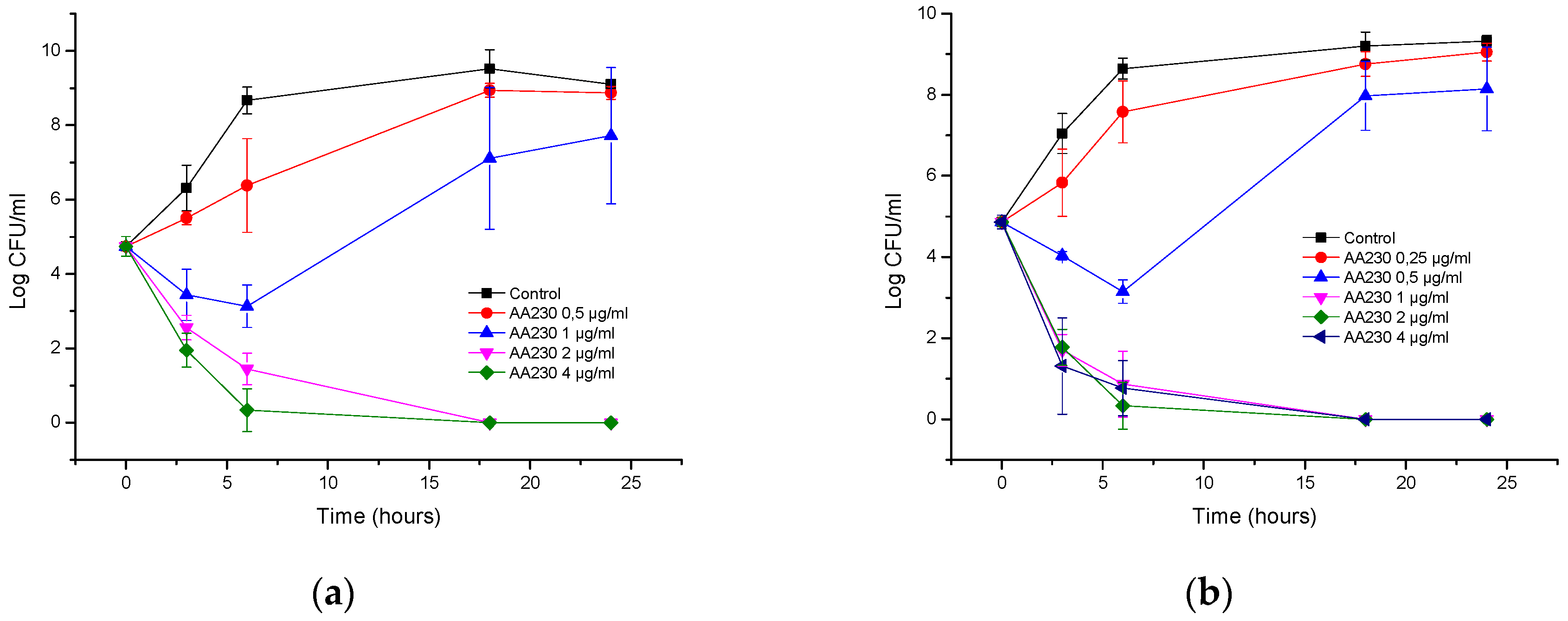
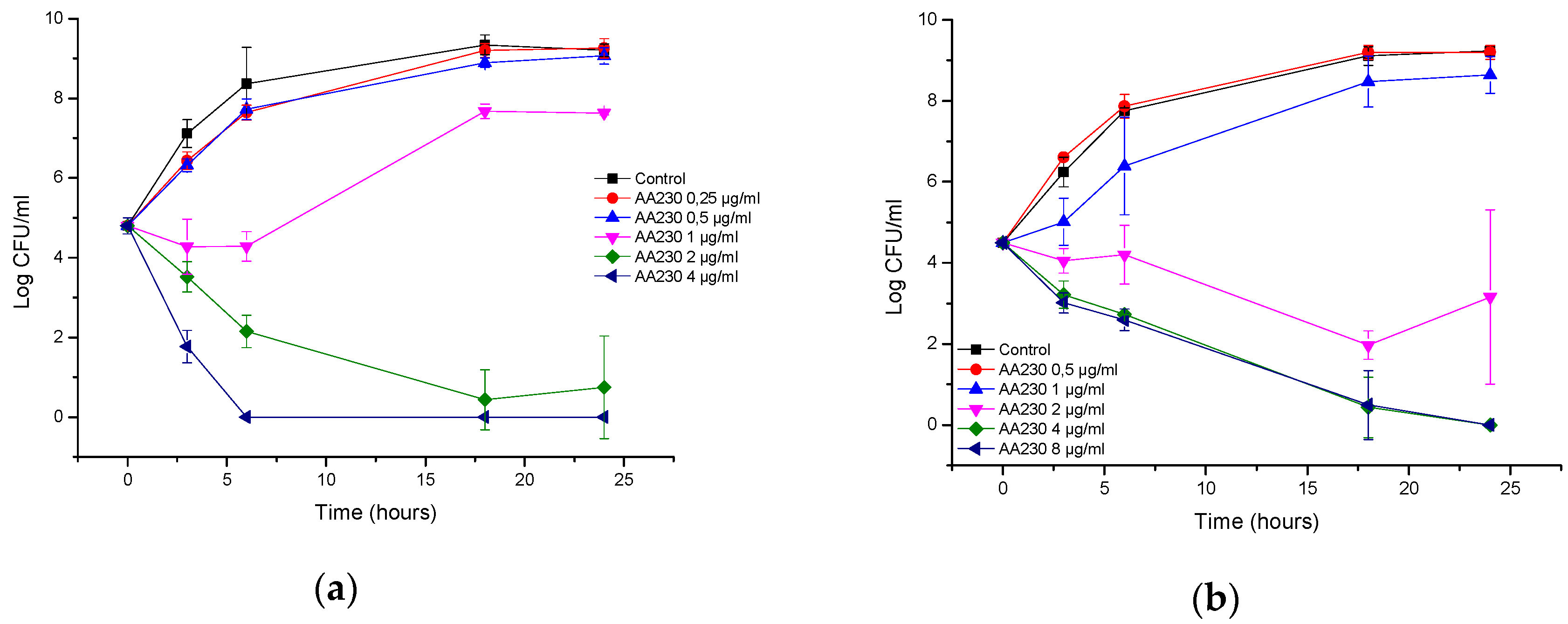
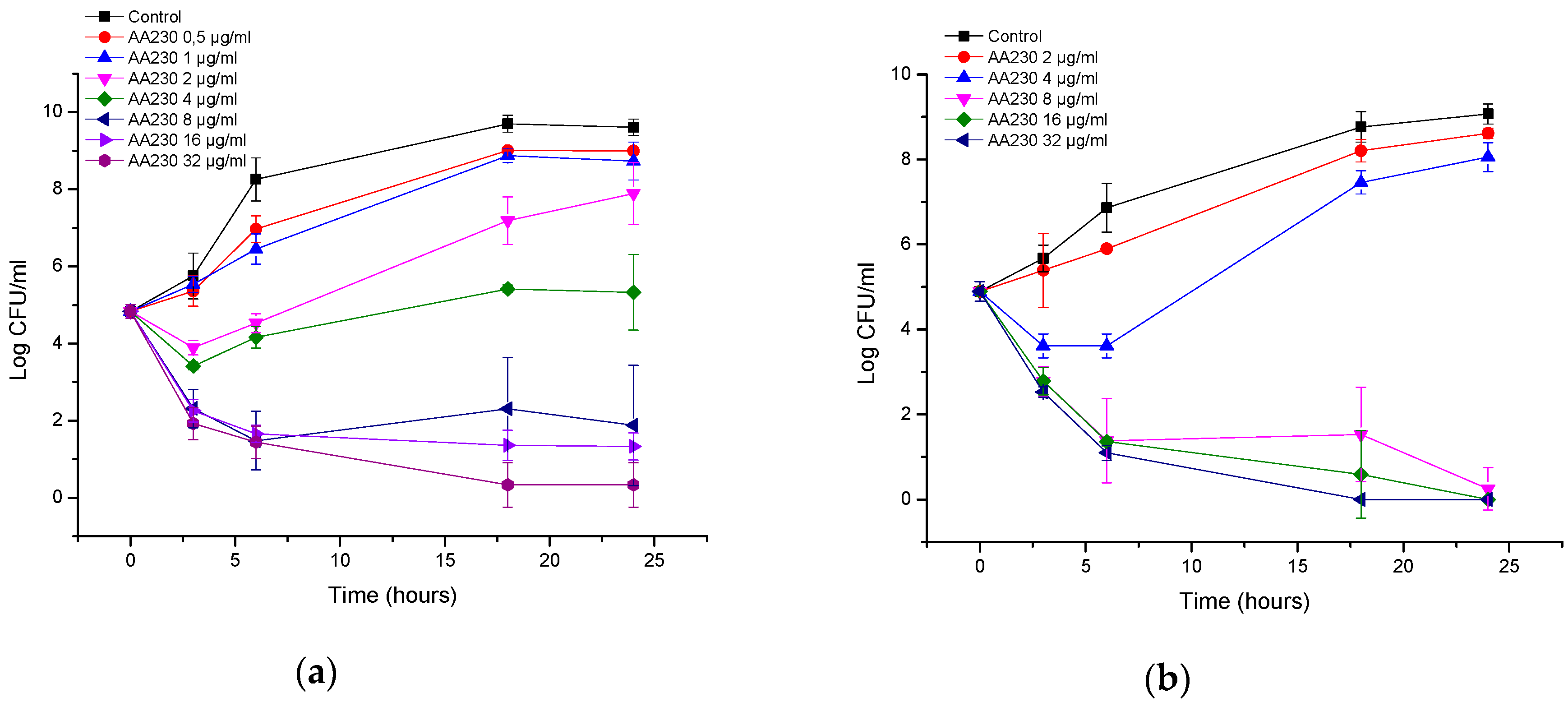
3.1. Antibacterial Activity of AA230
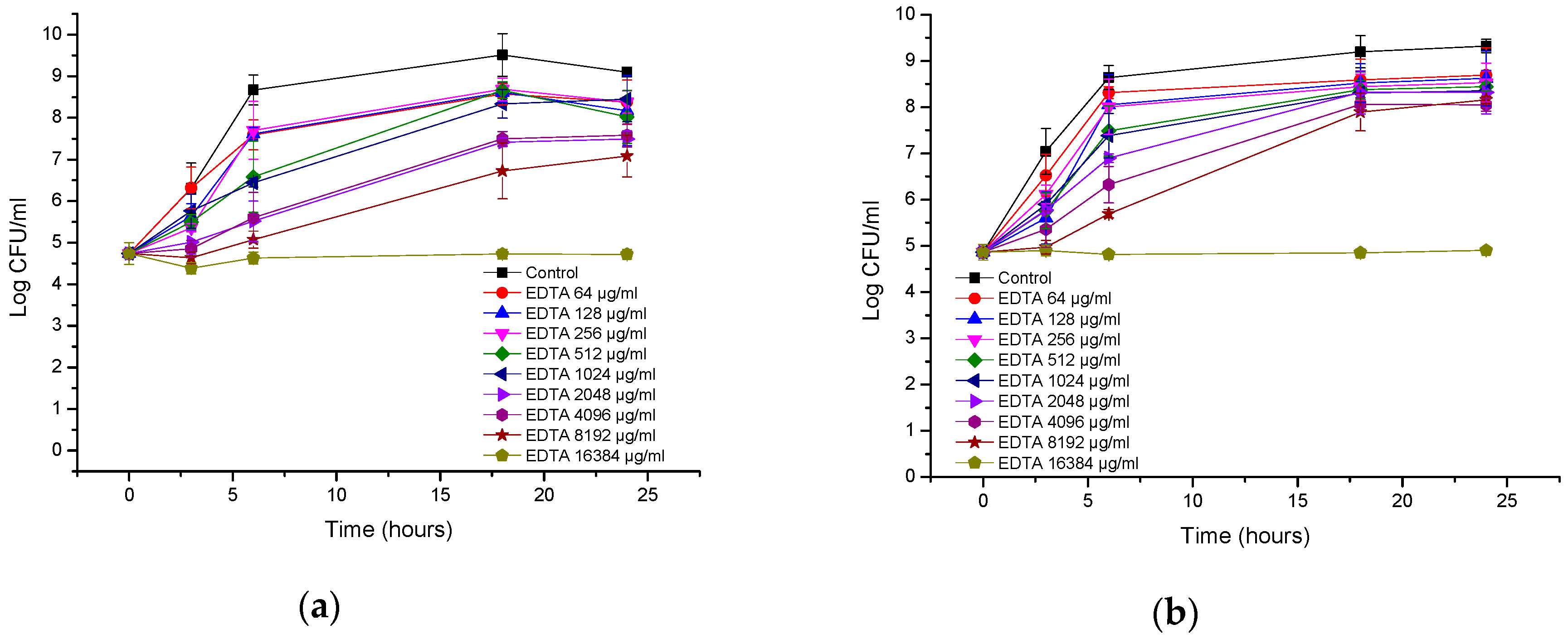
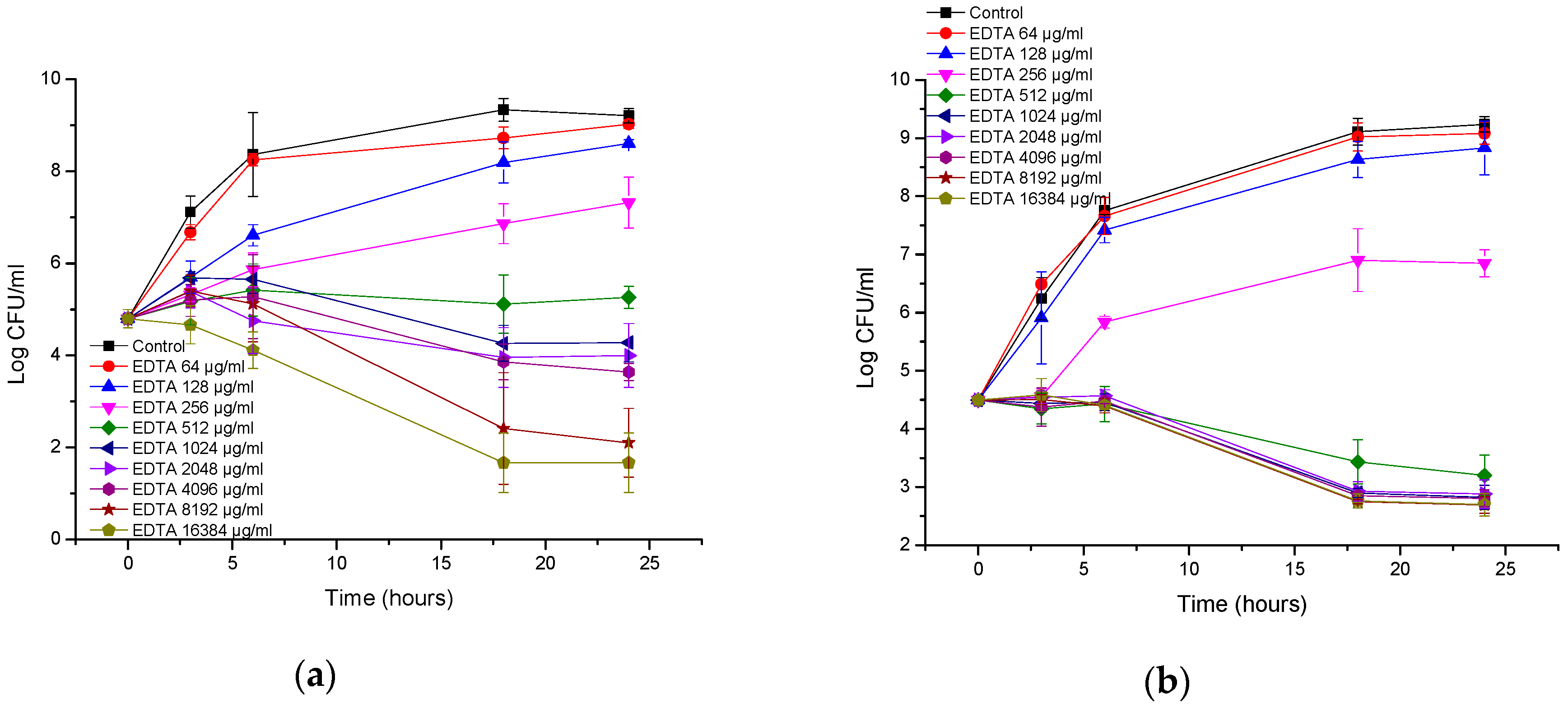
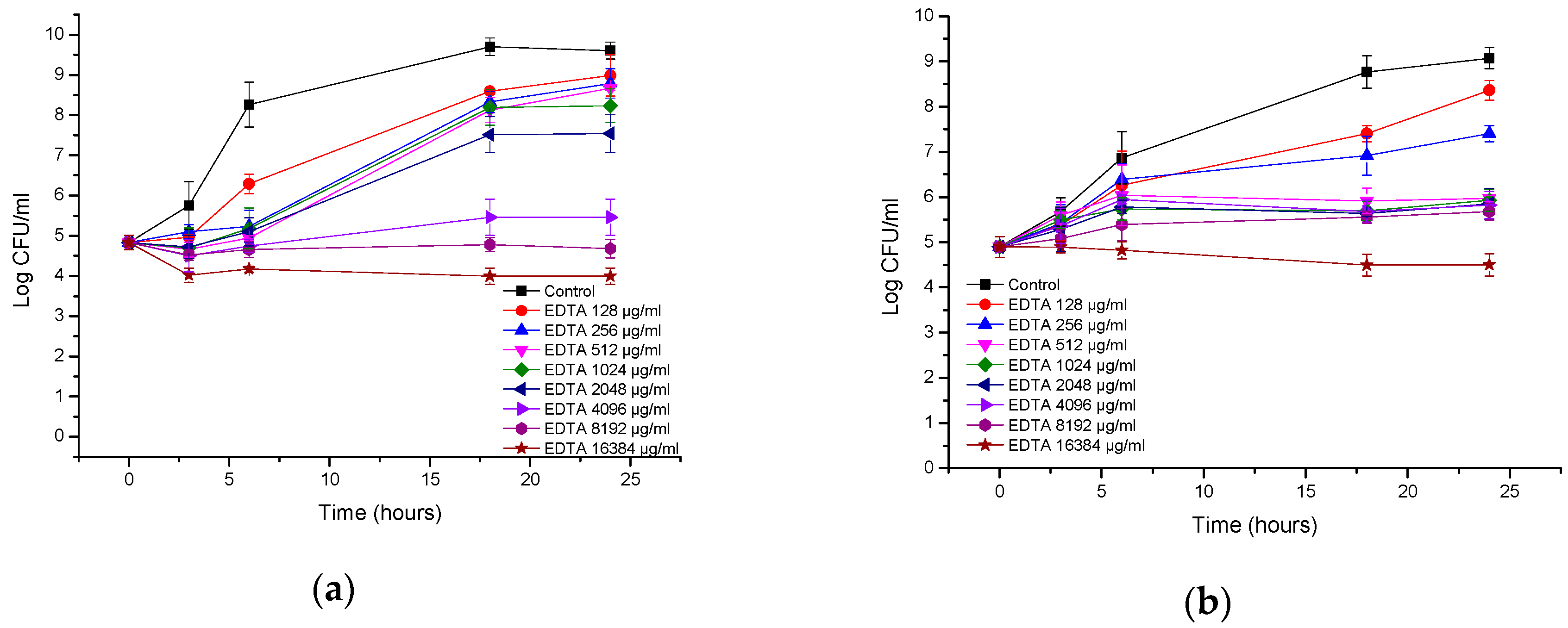
3.2. Antibacterial Activity of EDTA
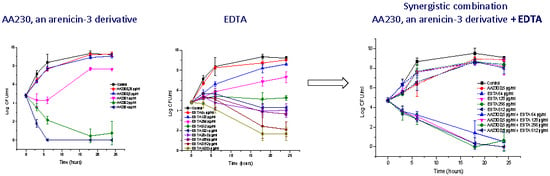
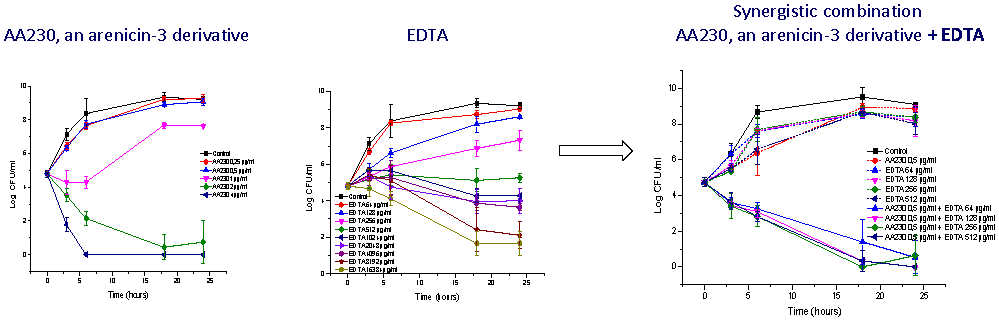
3.3. Antibacterial Activity of AA230/EDTA Combinations
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Panteleev, P.V.; Bolosov, I.A.; Balandin, S.V.; Ovchinnikova, T.V. Design of antimicrobial peptide arenicin analogs with improved therapeutic indices. J. Pept. Sci. 2015, 21, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Arias, C.A.; Murray, B.E. Antibiotic-resistant bugs in the 21st century-a clinical super-challenge. N. Engl. J. Med. 2009, 360, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Walkenhorst, W.F.; Sundrud, J.N.; Laviolette, J.M. Additivity and synergy between an antimicrobial peptide and inhibitory ions. Biochim. Biophys. Acta 2014, 1838, 2234–2242. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Umerska, A.; Cassisa, V.; Bastiat, G.; Matougui, N.; Nehme, H.; Manero, F.; Eveillard, M.; Saulnier, P. Synergistic interactions between antimicrobial peptides derived from plectasin and lipid nanocapsules containing monolaurin as a cosurfactant against Staphylococcus aureus. Int. J. Nanomed. 2017, 12, 5687–5699. [Google Scholar] [CrossRef] [PubMed]
- Boge, L.; Umerska, A.; Matougui, N.; Bysell, H.; Ringstad, L.; Davoudi, M.; Eriksson, J.; Edwards, K.; Andersson, M. Cubosomes post-loaded with antimicrobial peptides: Characterization, bactericidal effect and proteolytic stability. Int. J. Pharm. 2017, 526, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Nordström, R.; Nyström, L.; Andrén, O.C.J.; Malkoch, M.; Umerska, A.; Davoudi, M.; Schmidtchen, A.; Malmsten, M. Membrane interactions of microgels as carriers of antimicrobial peptides. J. Colloid Interface Sci. 2018, 513, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.; Piga, K.B.; Hyndman, E.; Vogel, H.J. Improving the activity of Trp-rich antimicrobial peptides by Arg/Lys substitutions and changing the length of cationic residues. Biomolecules 2018, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G. Peptide antibiotics and their role in innate immunity. Annu. Rev. Immunol. 1995, 13, 61–92. [Google Scholar] [CrossRef] [PubMed]
- Sitaram, N.; Nagaraj, R. Host-defense antimicrobial peptides: Importance of structure for activity. Curr. Pharm. Des. 2002, 8, 727–742. [Google Scholar] [CrossRef] [PubMed]
- Papagianni, M. Ribosomally synthesized peptides with antimicrobial properties: Biosynthesis, structure, function, and applications. Biotechnol. Adv. 2003, 21, 465–499. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef] [PubMed]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolym. Pept. Sci. Sect. 2002, 66, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Marquette, A.; Bechinger, B. Biophysical investigations elucidating the mechanisms of action of antimicrobial peptides and their synergism. Biomolecules 2018, 8, 18. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide antimicrobial agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.X.; Neve, S.; Elliott, A.G.; Cain, A.K.; Boinett, C.J.; Zuegg, J.; Steen, J.; Ramu, S.; Kavanagh, A.M.; Pelingon, R.; et al. Novel Arenicin-3 Peptide Antibiotics with Broad-Spectrum Activity Against MDR Gram-Negative Bacterial Act via Dual Mode of Actions. Available online: http://adeniumbiotech.com/wp-content/uploads/2011/10/Variants-characterization.pdf (accessed on 4 October 2018).
- Wang, X.; Wang, X.; Teng, D.; Zhang, Y.; Mao, R.; Xi, D.; Wang, J. Candidacidal mechanism of the arenicin-3-derived peptide NZ17074 from Arenicola marina. Appl. Microbiol. Biotechnol. 2014, 98, 7387–7398. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Lee, D.G. Synergistic effect of antimicrobial peptide arenicin-1 in combination with antibiotics against pathogenic bacteria. Res. Microbiol. 2012, 163, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Finnegan, S.; Percival, S.L. EDTA: An antimicrobial and antibiofilm agent for use in wound care. Adv. Wound Care 2015, 4, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.W.; Richards, R.M.E. Effect of ethylenediamine tetraacetate on the resistance of Pseudomonas aeruginosa to antibacterial agents. Nature 1965, 207, 1391–1393. [Google Scholar] [CrossRef] [PubMed]
- Lambert, R.J.W.; Hanlon, G.W.; Denyer, S.P. The synergistic effect of EDTA/antimicrobial combinations on Pseudomonas aeruginosa. J. Appl. Microbiol. 2004, 96, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Umerska, A.; Cassisa, V.; Matougui, N.; Joly-Guillou, M.-L.; Eveillard, M.; Saulnier, P. Antibacterial action of lipid nanocapsules containing fatty-acids or monoglycerides as co-surfactants. Eur. J. Pharm. Biopharm. 2016, 108, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Nehme, H.; Saulnier, P.; Ramadan, A.A.; Cassisa, V.; Guillet, C.; Eveillard, M.; Umerska, A. Antibacterial activity of antipsychotic agents, their association with lipid nanocapsules and its impact on the properties of the nanocarriers and on antibacterial activity. PLoS ONE 2018, 13, e0189950. [Google Scholar] [CrossRef] [PubMed]
- White, R.L.; Burgess, D.S.; Manduru, M.; Bosso, J.A. Comparison of three different in vitro methods of detecting synergy: Time-kill, checkerboard, and E test. Antimicrob. Agents Chemother. 1996, 40, 1914–1918. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C.; Weigel, L.M.; Appelbaum, P.C.; McDougal, L.K.; Chaitram, J.; McAllister, S.; Clark, N.; Killgore, G.; O’Hara, C.M.; Jevitt, L.; et al. Vancomycin-resistant Staphylococcus aureus isolate from a patient in Pennsylvania. Antimicrob. Agents Chemother. 2004, 48, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Valcourt, C.; Saulnier, P.; Umerska, A.; Zanelli, M.P.; Montagu, A.; Rossines, E.; Joly-Guillou, M.L. Synergistic interactions between doxycycline and terpenic components of essential oils encapsulated within lipid nanocapsules against gram negative bacteria. Int. J. Pharm. 2016, 498, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Kapoor, I.P.S.; Pandey, S.K.; Singh, U.K.; Singh, R.K. Studies on essential oils: Part 10; antibacterial activity of volatile oils of some spices. Phytother. Res. 2002, 16, 680–682. [Google Scholar] [CrossRef] [PubMed]
- Andrä, J.; Jakovkin, I.; Grötzinger, J.; Hecht, O.; Krasnosdembskaya, A.D.; Goldmann, T.; Gutsmann, T.; Leippe, M. Structure and mode of action of the antimicrobial peptide arenicin. Biochem. J. 2008, 410, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Hamoud, R.; Reichling, J.; Wink, M. Synergistic antimicrobial activity of combinations of sanguinarine and EDTA with vancomycin against multidrug resistant bacteria. Drug Metab. Lett. 2014, 8, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Leive, L. Release of lipopolysaccharide by EDTA treatment of E. coli. Biochem. Biophys. Res. Commun. 1965, 21, 290–296. [Google Scholar] [CrossRef]
- Gray, G.W.; Wilkinson, S.G. The action of ethylenediaminetetra-acetic acid on Pseudomonas aeruginosa. J. Appl. Microbiol. 1965, 28, 153–164. [Google Scholar] [CrossRef]
- Leive, L. Actinomycin sensitivity in Escherichia coli produced by EDTA. Biochem. Biophys. Res. Commun. 1965, 18, 13–17. [Google Scholar] [CrossRef]
- Asbell, M.A.; Eagon, R.G. The role of multivalent cations in the organization and structure of bacterial cell walls. Biochem. Biophys. Res. Commun. 1966, 22, 664–671. [Google Scholar] [CrossRef]
- Roberts, N.A.; Gray, G.W.; Wilkinson, S.G. Bactericidal action of ethylenediamine tetraacetic acid on Pseudomonas aeruginosa. Microbiology 1970, 2, 189–208. [Google Scholar]
- Rogers, S.W.; Gilleland, H.E., Jr.; Eagon, R.G. Characterization of a protein-lipopolysaccharide complex released from cell walls of Pseudomonas aeruginosa by ethylenediaminetetraacetic acid. Can. J. Microbiol. 1969, 15, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Richards, R.M.E.; Cavill, R.H. Electron microscope study of effect of benzalkonium chloride and edetate sodium on cell envelope of Pseudomonas aeruginosa. J. Pharm. Sci. 1976, 65, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Weiser, R.; Asscher, A.W.; Wimpenny, J. In vitro reversal of antibiotic resistance by ethylenediamine tetraacetic acid. Nature 1968, 219, 1365–1366. [Google Scholar] [CrossRef] [PubMed]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. Biophys. Acta 2009, 1794, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H.; Vaara, M. Molecular basis of bacterial outer membrane permeability. Microbiol. Rev. 1985, 49, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Lounatmaa, K.; Mäkelä, P.H.; Sarvas, M. The effect of polymyxin on the outer membrane of Salmonella: Ultrastructure of wild-type and polymyxin-resistant strains. J. Bacteriol. 1976, 127, 1900–1907. [Google Scholar]
- Teuber, M.; Bader, J. Action of polymyxin on bacterial membranes. Binding capacicites for polymyxin B of inner and outer membranes isolated from Salmonella typhimurium G30. Arch. Microbiol. 1976, 109, 51–58. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | MIC of AA230 (µg/mL) | MBC of AA230 (µg/mL) | MIC of EDTA (µg/mL) | MBC of EDTA (µg/mL) |
|---|---|---|---|---|
| A. baumannii AYE | 2–4 | 4 | 1024 | ≥16,384 |
| A. baumannii RCH | 2–4 | 4–8 | 512 | ≥16,384 |
| E. coli ATCC | 2–4 | 2–4 | ≥16,384 | ≥16,384 |
| ESBL E. coli | 1–2 | 1–2 | ≥16,384 | ≥16,384 |
| P. aeruginosa ATCC | 4 | 8–16 | 8192 | ≥16,384 |
| P. aeruginosa clinial | 4–8 | 8 | 8192 | ≥16,384 |
| MIC AA230 in Combination (µg/mL) | MIC EDTA in Combination (µg/mL) | FIC Index | Interpretation |
|---|---|---|---|
| E. coli ATCC | |||
| 0.5 | 64 | 0.129–0.254 | S |
| 1 | 64 | 0.254–0.504 | S |
| ESBL E. coli | |||
| 0.25 | 128 | 0.133–0.258 | S |
| 0.5 | 64 | 0.254–0.504 | S |
| A. baumannii ATCC | |||
| 0.5 | 256 | 0.375–0.500 | S |
| 1 | 64 | 0.313–0.563 | S/A |
| A. baumannii RCH | |||
| 0.5 | 256 | 0.625–0.750 | A |
| 1 | 128 | 0.500–0.750 | S/A |
| P. aeruginosa ATCC | |||
| 2 | 128 | 0.516 | A |
| P. aeruginosa clinical | |||
| 2 | 256 | 0.281–0.531 | S/A |
| 4 | 128 | 0.516–1.016 | A |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umerska, A.; Strandh, M.; Cassisa, V.; Matougui, N.; Eveillard, M.; Saulnier, P. Synergistic Effect of Combinations Containing EDTA and the Antimicrobial Peptide AA230, an Arenicin-3 Derivative, on Gram-Negative Bacteria. Biomolecules 2018, 8, 122. https://doi.org/10.3390/biom8040122
Umerska A, Strandh M, Cassisa V, Matougui N, Eveillard M, Saulnier P. Synergistic Effect of Combinations Containing EDTA and the Antimicrobial Peptide AA230, an Arenicin-3 Derivative, on Gram-Negative Bacteria. Biomolecules. 2018; 8(4):122. https://doi.org/10.3390/biom8040122
Chicago/Turabian StyleUmerska, Anita, Magnus Strandh, Viviane Cassisa, Nada Matougui, Matthieu Eveillard, and Patrick Saulnier. 2018. "Synergistic Effect of Combinations Containing EDTA and the Antimicrobial Peptide AA230, an Arenicin-3 Derivative, on Gram-Negative Bacteria" Biomolecules 8, no. 4: 122. https://doi.org/10.3390/biom8040122
APA StyleUmerska, A., Strandh, M., Cassisa, V., Matougui, N., Eveillard, M., & Saulnier, P. (2018). Synergistic Effect of Combinations Containing EDTA and the Antimicrobial Peptide AA230, an Arenicin-3 Derivative, on Gram-Negative Bacteria. Biomolecules, 8(4), 122. https://doi.org/10.3390/biom8040122