

Zinc-Induced Folding and Solution Structure of the Eponymous Novel Zinc Finger from the ZC4H2 Protein



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
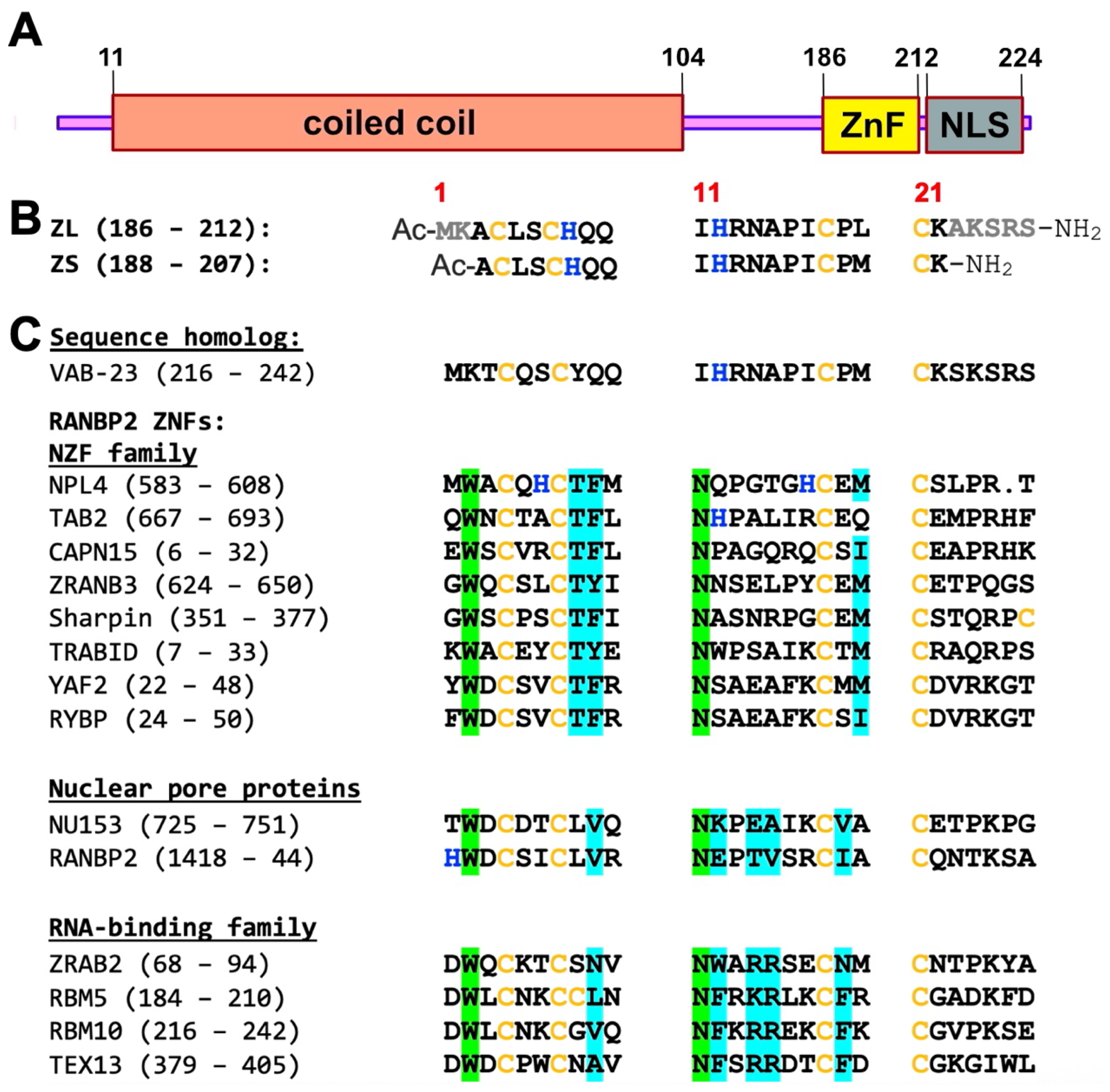
2. Materials and Methods
2.1. Samples
2.2. NMR Spectroscopy
2.3. CD Spectroscopy
2.4. UV–Visible Spectroscopy of Co2+ Complexes
2.5. Alpha Fold 3 Simulations of Random Sequences
2.6. NMR Structure Calculations
3. Results
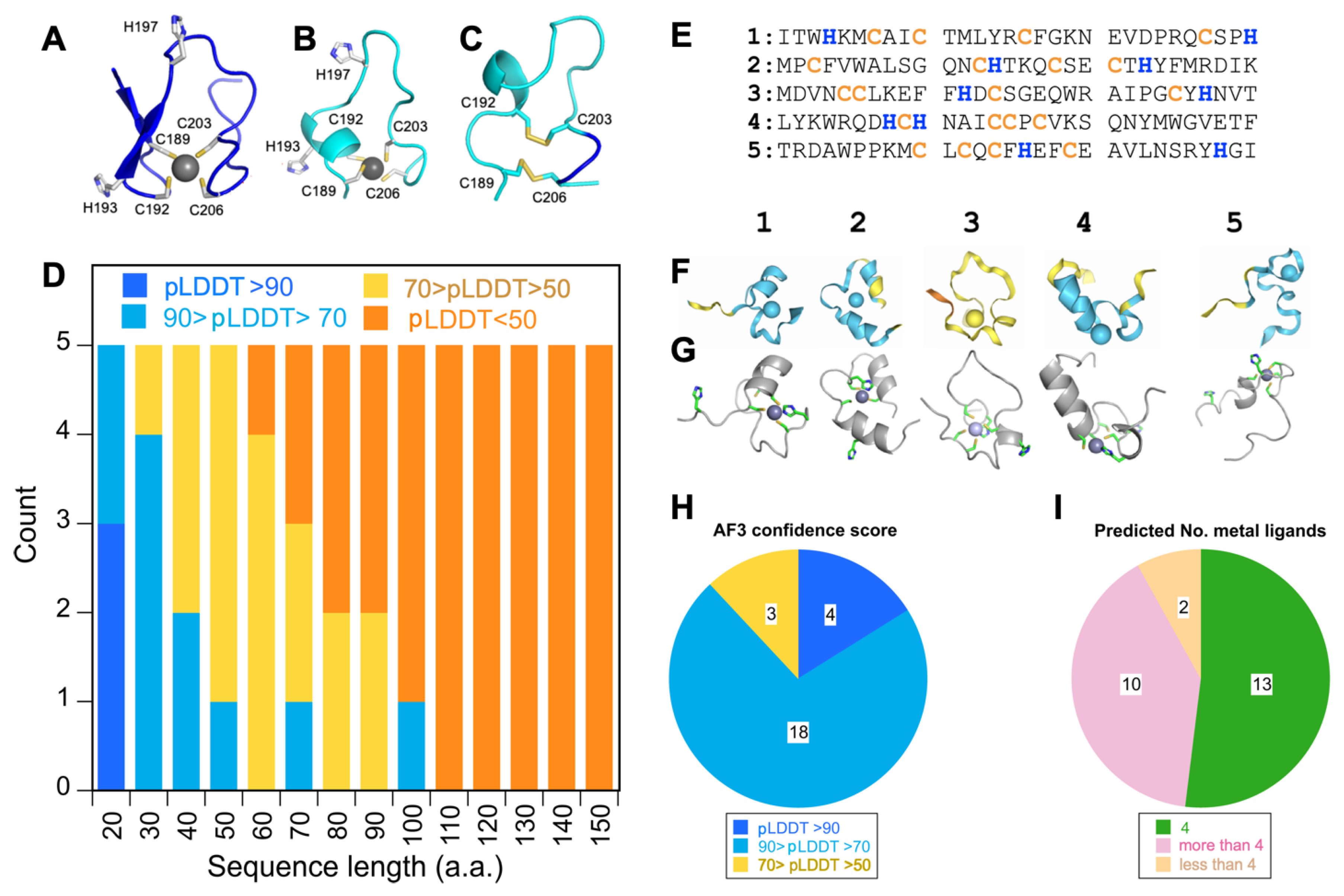
3.1. ZC4H2 Has a Genuine ZNF with Domain Boundaries Larger than Those Specified by UniProt
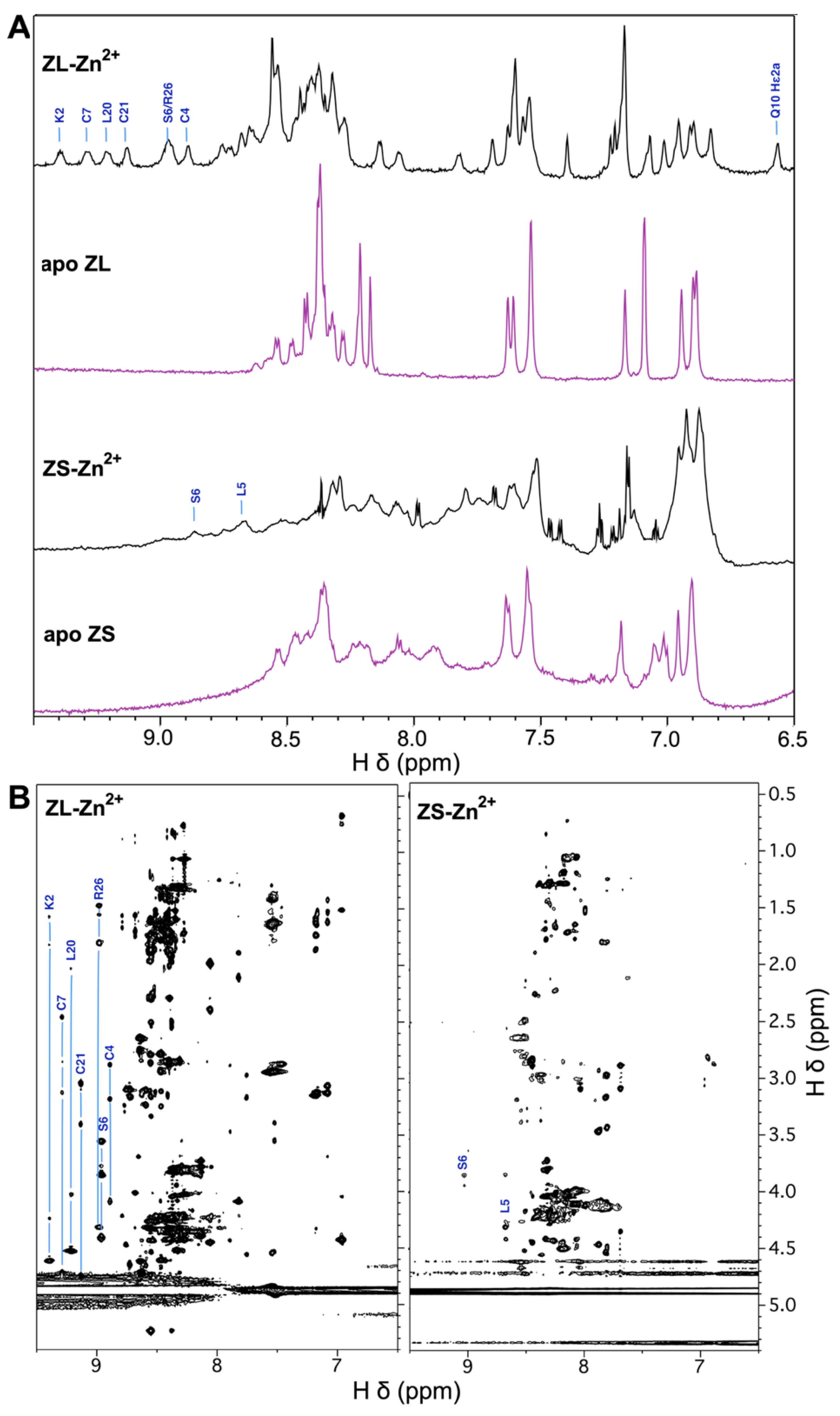
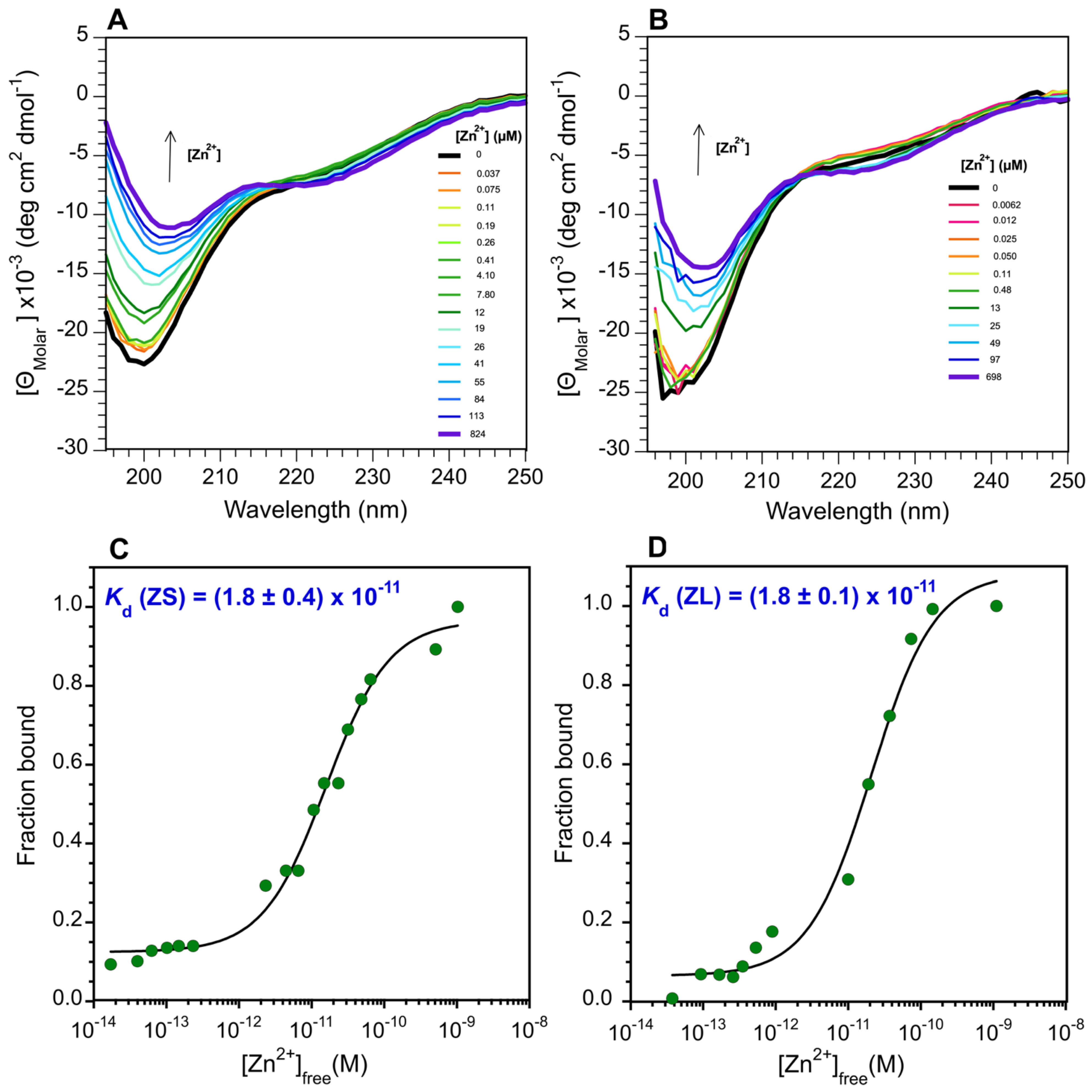
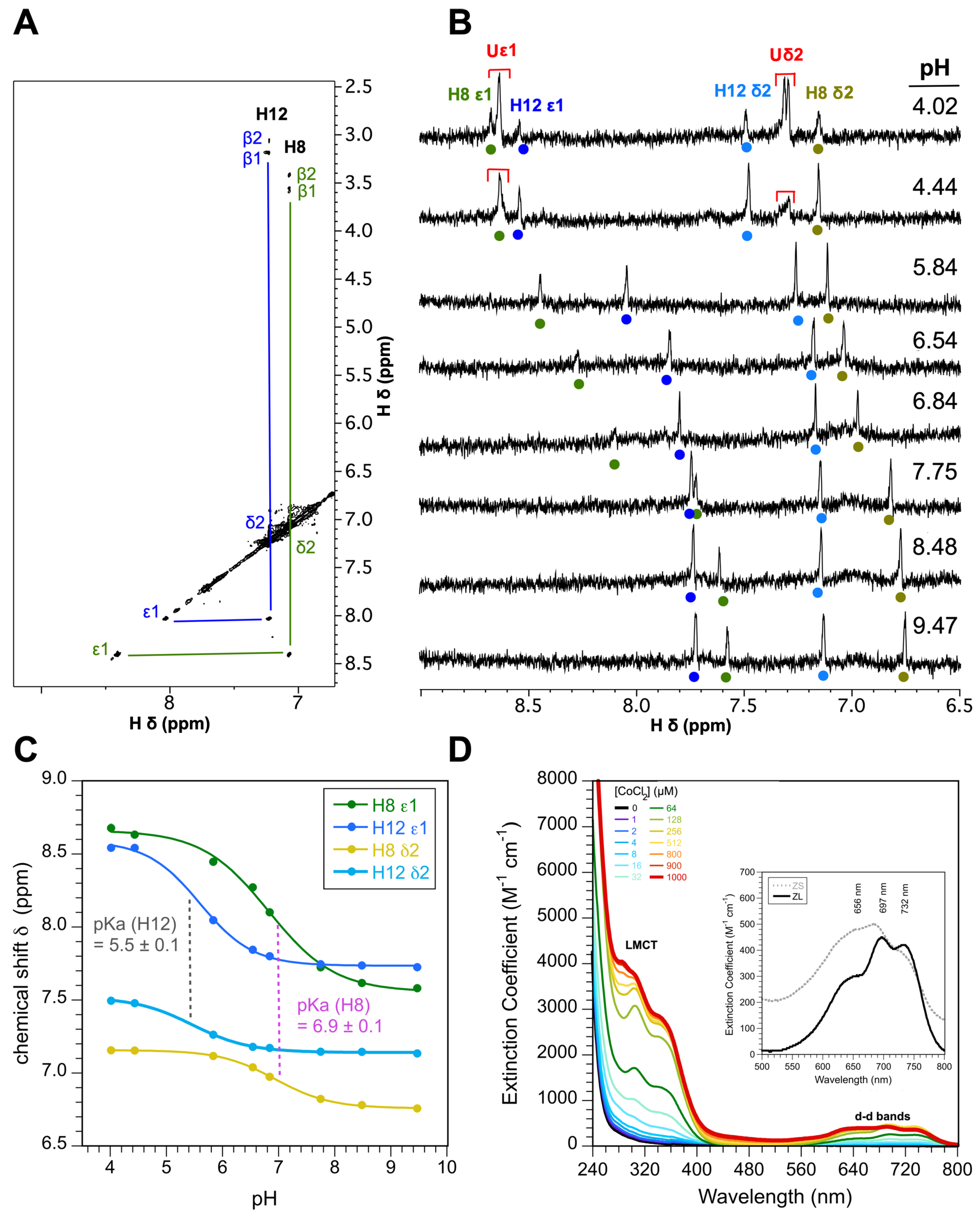
3.2. The ZnF of ZC4H2 Has a CCCC Metal Coordination Sphere
3.3. Alpha Fold 3 Predicts the ZnF Structure with Confidence but Also Similarly Sized Random Sequences
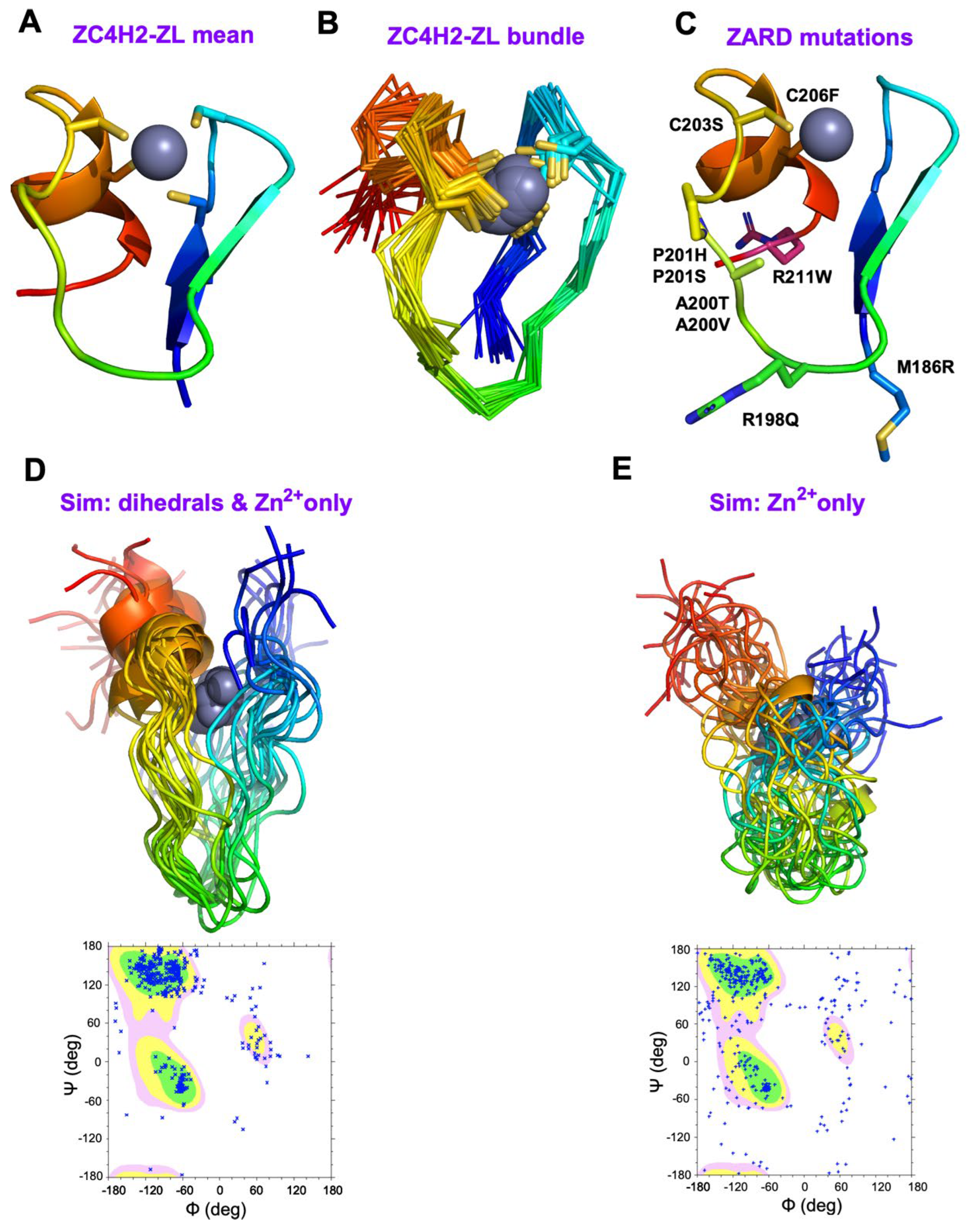
3.4. The NMR Structure of ZC4H2 Is a Variant of the RANBP2 ZnF Fold
4. Discussion
4.1. Structural Mapping of the Eight ZARD Mutations Within the ZnF
4.2. Hierarchical Folding of the ZnF Domain in ZC4H2
4.3. The ZnF of ZC4H2 Represents a Unique Sequence Family Amongst Structural Homologs
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AF3 | Alpha Fold 3 |
| CD | circular dichroism |
| DOSY | diffusion ordered spectroscopy |
| DSS | 3-(Trimethylsilyl)propane-1-sulfonic acid |
| EGTA | ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid |
| HSQC | heteronuclear single quantum correlation |
| LMCT | ligand-to-metal charge transfer |
| NLS | nuclear localization signal |
| NMR | nuclear magnetic resonance |
| NOE | nuclear Overhauser effect |
| NOESY | nuclear Overhauser effect spectroscopy |
| PDB | protein data bank |
| pLDDT | predicted local distance difference test |
| RING | really interesting new gene |
| RMSD | root mean square deviation |
| SD | standard deviation |
| Shh | Sonic Hedgehog |
| TCEP | tris(2-carboxyethyl)phosphine |
| TOCSY | total correlation spectroscopy |
| UV–Vis | ultraviolet-visible spectrophotometry |
| WRWF | Wieacker–Wolff syndrome |
| ZARD | ZC4H2-associated rare disorders |
| ZC4H2-ZL | long fragment comprising residues 186-212 of the ZC4H2 protein |
| ZC4H2-ZS | short fragment comprising residues 188-207 of the ZC4H2 protein |
| ZnF | zinc finger |
References
- Piccolo, G.; d’Annunzio, G.; Amadori, E.; Riva, A.; Borgia, P.; Tortora, D.; Maghnie, M.; Minetti, C.; Gitto, E.; Iacomino, M.; et al. Neuromuscular and Neuroendocrinological Features Associated With ZC4H2-Related Arthrogryposis Multiplex Congenita in a Sicilian Family: A Case Report. Front. Neurol. 2021, 12, 704747. [Google Scholar] [CrossRef]
- Peters, S.; Sportiello, K.; Mandalapu, S.; Nguyen, A.; Carrier, R.; Dickinson, C.; Paciorkowski, A.; Bearden, D. Genotype-Phenotype Correlations and Sex Differences in ZC4H2-Associated Rare Disorder. Pediatr. Neurol. 2024, 158, 100–112. [Google Scholar] [CrossRef]
- Frints, S.G.M.; Hennig, F.; Colombo, R.; Jacquemont, S.; Terhal, P.; Zimmerman, H.H.; Hunt, D.; Mendelsohn, B.A.; Kordass, U.; Webster, R.; et al. Deleterious de novo variants of X-linked ZC4H2 in females cause a variable phenotype with neurogenic arthrogryposis multiplex congenita. Hum. Mutat. 2019, 40, 2270–2285. [Google Scholar] [CrossRef]
- Hirata, H.; Nanda, I.; van Riesen, A.; McMichael, G.; Hu, H.; Hambrock, M.; Papon, M.A.; Fischer, U.; Marouillat, S.; Ding, C.; et al. ZC4H2 mutations are associated with arthrogryposis multiplex congenita and intellectual disability through impairment of central and peripheral synaptic plasticity. Am. J. Hum. Genet. 2013, 92, 681–695. [Google Scholar] [CrossRef]
- Wieacker, P.; Wolff, G.; Wienker, T.F.; Sauer, M. A new X-linked syndrome with muscle atrophy, congenital contractures, and oculomotor apraxia. Am. J. Med. Genet. 1985, 20, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Ibarra-Ramirez, M.; Fernandez-de-Luna, M.L.; Campos-Acevedo, L.D.; Arenas-Estala, J.; Martinez-de-Villarreal, L.E.; Rodriguez-Garza, C.; DeLagarza-Pineda, O.; Mohamed-Noriega, J. Optic nerve abnormalities in female-restricted Wieacker-Wolff syndrome by a novel variant in the ZC4H2 gene. Ophthalmic Genet. 2023, 44, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, T.; Mizukami, M.; Terada, K.; Ishikawa, A.; Hinotsu, S.; Kobayashi, M.; Kato, K.; Ogi, T.; Tsugawa, T.; Sakurai, A. A novel ZC4H2 variant in a female with severe respiratory complications. Brain Dev. 2022, 44, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Del Toro, N.; Shrivastava, A.; Ragueneau, E.; Meldal, B.; Combe, C.; Barrera, E.; Perfetto, L.; How, K.; Ratan, P.; Shirodkar, G.; et al. The IntAct database: Efficient access to fine-grained molecular interaction data. Nucleic Acids Res. 2022, 50, D648–D653. [Google Scholar] [CrossRef]
- Ma, P.; Ren, B.; Yang, X.; Sun, B.; Liu, X.; Kong, Q.; Li, C.; Mao, B. ZC4H2 stabilizes Smads to enhance BMP signalling, which is involved in neural development in Xenopus. Open Biol. 2017, 7, 170122. [Google Scholar] [CrossRef]
- Kobayashi, S.; Sato, A.; Chiba, Y.; Adachi, N.; Kakimoto, Y.; Suzuki, H.; Yamada, M.; Kosaki, K.; Tanaka, H. Wieacker-Wolff syndrome with hyperinsulinemic hypoglycemia successfully treated using diazoxide: A case report. Clin. Pediatr. Endocrinol. 2025, 34, 70–76. [Google Scholar] [CrossRef]
- Vangeel, L.; Janssens, A.; Lemmens, I.; Lievens, S.; Tavernier, J.; Voets, T. The Zinc-Finger Domain Containing Protein ZC4H2 Interacts with TRPV4, Enhancing Channel Activity and Turnover at the Plasma Membrane. Int. J. Mol. Sci. 2020, 21, 3556. [Google Scholar] [CrossRef]
- Cho, T.J.; Matsumoto, K.; Fano, V.; Dai, J.; Kim, O.H.; Chae, J.H.; Yoo, W.J.; Tanaka, Y.; Matsui, Y.; Takigami, I.; et al. TRPV4-pathy manifesting both skeletal dysplasia and peripheral neuropathy: A report of three patients. Am. J. Med. Genet. A 2012, 158A, 795–802. [Google Scholar] [CrossRef]
- Briscoe, J.; Therond, P.P. The mechanisms of Hedgehog signalling and its roles in development and disease. Nat. Rev. Mol. Cell Biol. 2013, 14, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Choi, T.I.; Park, S.; Kim, M.H.; Kim, C.H.; Lee, S. Rnf220 cooperates with Zc4h2 to specify spinal progenitor domains. Development 2018, 145, dev165340. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Mao, B. The many faces of the E3 ubiquitin ligase, RNF220, in neural development and beyond. Dev. Growth Differ. 2022, 64, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Song, N.N.; Cheng, X.; Zhu, L.; Zhang, Q.; Zhang, L.L.; Yang, X.; Wang, H.; Kong, Q.; Shi, D.; et al. ZC4H2 stabilizes RNF220 to pattern ventral spinal cord through modulating Shh/Gli signaling. J. Mol. Cell Biol. 2020, 12, 337–344. [Google Scholar] [CrossRef]
- Li, Y.; Yang, C.; Wang, H.; Zhao, L.; Kong, Q.; Cang, Y.; Zhao, S.; Lv, L.; Li, Y.; Mao, B.; et al. Sequential stabilization of RNF220 by RLIM and ZC4H2 during cerebellum development and Shh-group medulloblastoma progression. J. Mol. Cell Biol. 2022, 14, mjab082. [Google Scholar] [CrossRef]
- Wang, D.; Hu, D.; Guo, Z.; Hu, R.; Wang, Q.; Liu, Y.; Liu, M.; Meng, Z.; Yang, H.; Zhang, Y.; et al. A novel de novo nonsense mutation in ZC4H2 causes Wieacker-Wolff Syndrome. Mol. Genet. Genom. Med. 2020, 8, e1100. [Google Scholar] [CrossRef]
- Hirosawa, M.; Nagase, T.; Ishikawa, K.; Kikuno, R.; Nomura, N.; Ohara, O. Characterization of cDNA clones selected by the GeneMark analysis from size-fractionated cDNA libraries from human brain. DNA Res. 1999, 6, 329–336. [Google Scholar] [CrossRef]
- Wang, Y.; Han, K.J.; Pang, X.W.; Vaughan, H.A.; Qu, W.; Dong, X.Y.; Peng, J.R.; Zhao, H.T.; Rui, J.A.; Leng, X.S.; et al. Large scale identification of human hepatocellular carcinoma-associated antigens by autoantibodies. J. Immunol. 2002, 169, 1102–1109. [Google Scholar] [CrossRef]
- Pellegrino, M.W.; Gasser, R.B.; Sprenger, F.; Stetak, A.; Hajnal, A. The conserved zinc finger protein VAB-23 is an essential regulator of epidermal morphogenesis in Caenorhabditis elegans. Dev. Biol. 2009, 336, 84–93. [Google Scholar] [CrossRef]
- Berg, J.M. Proposed structure for the zinc-binding domains from transcription factor IIIA and related proteins. Proc. Natl. Acad. Sci. USA 1988, 85, 99–102. [Google Scholar] [CrossRef]
- Klug, A. The discovery of zinc fingers and their applications in gene regulation and genome manipulation. Annu. Rev. Biochem. 2010, 79, 213–231. [Google Scholar] [CrossRef]
- Kluska, K.; Adamczyk, J.; Krezel, A. Metal binding properties, stability and reactivity of zinc fingers. Coord. Chem. Rev. 2018, 367, 18–64. [Google Scholar] [CrossRef]
- Lee, M.S.; Gippert, G.P.; Soman, K.V.; Case, D.A.; Wright, P.E. Three-dimensional solution structure of a single zinc finger DNA-binding domain. Science 1989, 245, 635–637. [Google Scholar] [CrossRef]
- Michalek, J.L.; Besold, A.N.; Michel, S.L. Cysteine and histidine shuffling: Mixing and matching cysteine and histidine residues in zinc finger proteins to afford different folds and function. Dalton Trans. 2011, 40, 12619–12632. [Google Scholar] [CrossRef]
- Neuhaus, D. Zinc finger structure determination by NMR: Why zinc fingers can be a handful. Prog. Nucl. Magn. Reson. Spectrosc. 2022, 130–131, 62–105. [Google Scholar] [CrossRef]
- Abbehausen, C. Zinc finger domains as therapeutic targets for metal-based compounds—An update. Metallomics 2019, 11, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Padjasek, M.; Kocyla, A.; Kluska, K.; Kerber, O.; Tran, J.B.; Krezel, A. Structural zinc binding sites shaped for greater works: Structure-function relations in classical zinc finger, hook and clasp domains. J. Inorg. Biochem. 2020, 204, 110955. [Google Scholar] [CrossRef] [PubMed]
- Krishna, S.S.; Majumdar, I.; Grishin, N.V. Structural classification of zinc fingers: Survey and summary. Nucleic Acids Res. 2003, 31, 532–550. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef]
- Kluska, K.; Adamczyk, J.; Krezel, A. Metal binding properties of zinc fingers with a naturally altered metal binding site. Metallomics 2018, 10, 248–263. [Google Scholar] [CrossRef]
- Michel, M.A.; Scutts, S.; Komander, D. Secondary interactions in ubiquitin-binding domains achieve linkage or substrate specificity. Cell Rep. 2024, 43, 114545. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Alam, S.L.; Meyer, H.H.; Payne, M.; Stemmler, T.L.; Davis, D.R.; Sundquist, W.I. Structure and ubiquitin interactions of the conserved zinc finger domain of Npl4. J. Biol. Chem. 2003, 278, 20225–20234. [Google Scholar] [CrossRef]
- Singh, B.B.; Patel, H.H.; Roepman, R.; Schick, D.; Ferreira, P.A. The zinc finger cluster domain of RanBP2 is a specific docking site for the nuclear export factor, exportin-1. J. Biol. Chem. 1999, 274, 37370–37378. [Google Scholar] [CrossRef]
- Yokoyama, N.; Hayashi, N.; Seki, T.; Pante, N.; Ohba, T.; Nishii, K.; Kuma, K.; Hayashida, T.; Miyata, T.; Aebi, U.; et al. A giant nucleopore protein that binds Ran/TC4. Nature 1995, 376, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.D.; Mansfield, R.E.; Leung, W.; Vaz, P.M.; Loughlin, F.E.; Grant, R.P.; Mackay, J.P. Characterization of a family of RanBP2-type zinc fingers that can recognize single-stranded RNA. J. Mol. Biol. 2011, 407, 273–283. [Google Scholar] [CrossRef]
- Alam, S.L.; Sun, J.; Payne, M.; Welch, B.D.; Blake, B.K.; Davis, D.R.; Meyer, H.H.; Emr, S.D.; Sundquist, W.I. Ubiquitin interactions of NZF zinc fingers. EMBO J. 2004, 23, 1411–1421. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E. Revolutionary cryo-EM is taking over structural biology. Nature 2020, 578, 201. [Google Scholar] [CrossRef]
- Harris, R.E.; Whitehead, R.D., 3rd; Alexandrescu, A.T. Solution structure of the Z0 domain from transcription repressor BCL11A sheds light on the sequence properties of protein-binding zinc fingers. Protein Sci. 2025, 34, e70097. [Google Scholar] [CrossRef]
- Matousek, W.M.; Alexandrescu, A.T. NMR structure of the C-terminal domain of SecA in the free state. Biochim. Biophys. Acta 2004, 1702, 163–171. [Google Scholar] [CrossRef]
- Rua, A.J.; Alexandrescu, A.T. Formerly degenerate seventh zinc finger domain from transcription factor ZNF711 rehabilitated by experimental NMR structure. Protein Sci. 2024, 33, e5149. [Google Scholar] [CrossRef]
- Rua, A.J.; Whitehead, R.D., 3rd; Alexandrescu, A.T. NMR structure verifies the eponymous zinc finger domain of transcription factor ZNF750. J. Struct. Biol. X 2023, 8, 100093. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.M. The Bicinchoninic Acid (BCA) Assay for Protein Quantitation. In The Protein Protocols Handbook, 2nd ed.; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2002; pp. 11–15. [Google Scholar]
- Wishart, D.S.; Bigam, C.G.; Yao, J.; Abildgaard, F.; Dyson, H.J.; Oldfield, E.; Markley, J.L.; Sykes, B.D. 1H, 13C and 15N chemical shift referencing in biomolecular NMR. J. Biomol. NMR 1995, 6, 135–140. [Google Scholar] [CrossRef]
- Harprecht, C.; Okifo, O.; Robbins, K.J.; Motwani, T.; Alexandrescu, A.T.; Teschke, C.M. Contextual Role of a Salt Bridge in the Phage P22 Coat Protein I-Domain. J. Biol. Chem. 2016, 291, 11359–11372. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, R.D., 3rd; Teschke, C.M.; Alexandrescu, A.T. Pulse-field gradient nuclear magnetic resonance of protein translational diffusion from native to non-native states. Protein Sci. 2022, 31, e4321. [Google Scholar] [CrossRef]
- Vranken, W.F.; Boucher, W.; Stevens, T.J.; Fogh, R.H.; Pajon, A.; Llinas, M.; Ulrich, E.L.; Markley, J.L.; Ionides, J.; Laue, E.D. The CCPN data model for NMR spectroscopy: Development of a software pipeline. Proteins 2005, 59, 687–696. [Google Scholar] [CrossRef]
- Ivanova, E.; Ball, M.; Lu, H. Zinc binding of Tim10: Evidence for existence of an unstructured binding intermediate for a zinc finger protein. Proteins Struct. Funct. Bioinform. 2008, 71, 467–475. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Bax, A. Protein backbone and sidechain torsion angles predicted from NMR chemical shifts using artificial neural networks. J. Biomol. NMR 2013, 56, 227–241. [Google Scholar] [CrossRef]
- Wuthrich, K.; Billeter, M.; Braun, W. Pseudo-structures for the 20 common amino acids for use in studies of protein conformations by measurements of intramolecular proton-proton distance constraints with nuclear magnetic resonance. J. Mol. Biol. 1983, 169, 949–961. [Google Scholar] [CrossRef]
- Ramboarina, S.; Druillennec, S.; Morellet, N.; Bouaziz, S.; Roques, B.P. Target specificity of human immunodeficiency virus type 1 NCp7 requires an intact conformation of its CCHC N-terminal zinc finger. J. Virol. 2004, 78, 6682–6687. [Google Scholar] [CrossRef]
- Tejero, R.; Snyder, D.; Mao, B.; Aramini, J.M.; Montelione, G.T. PDBStat: A universal restraint converter and restraint analysis software package for protein NMR. J. Biomol. NMR 2013, 56, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Maciejewski, M.W.; Schuyler, A.D.; Gryk, M.R.; Moraru, I.I.; Romero, P.R.; Ulrich, E.L.; Eghbalnia, H.R.; Livny, M.; Delaglio, F.; Hoch, J.C. NMRbox: A Resource for Biomolecular NMR Computation. Biophys. J. 2017, 112, 1529–1534. [Google Scholar] [CrossRef] [PubMed]
- Bermejo, G.A.; Tjandra, N.; Clore, G.M.; Schwieters, C.D. Xplor-NIH: Better parameters and protocols for NMR protein structure determination. Protein Sci. 2024, 33, e4922. [Google Scholar] [CrossRef]
- Rieping, W.; Habeck, M.; Bardiaux, B.; Bernard, A.; Malliavin, T.E.; Nilges, M. ARIA2: Automated NOE assignment and data integration in NMR structure calculation. Bioinformatics 2007, 23, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, R.D., 3rd; Teschke, C.M.; Alexandrescu, A.T. NMR Mapping of Disordered Segments from a Viral Scaffolding Protein Enclosed in a 23 MDa Procapsid. Biophys. J. 2019, 117, 1387–1392. [Google Scholar] [CrossRef]
- Alexandrescu, A.T.; Evans, P.A.; Pitkeathly, M.; Baum, J.; Dobson, C.M. Structure and dynamics of the acid-denatured molten globule state of alpha-lactalbumin: A two-dimensional NMR study. Biochemistry 1993, 32, 1707–1718. [Google Scholar] [CrossRef]
- Nomura, A.; Sugiura, Y. Contribution of individual zinc ligands to metal binding and peptide folding of zinc finger peptides. Inorg. Chem. 2002, 41, 3693–3698. [Google Scholar] [CrossRef]
- Curtis, D.; Treiber, D.K.; Tao, F.; Zamore, P.D.; Williamson, J.R.; Lehmann, R. A CCHC metal-binding domain in Nanos is essential for translational regulation. EMBO J. 1997, 16, 834–843. [Google Scholar] [CrossRef]
- Croke, R.L.; Patil, S.M.; Quevreaux, J.; Kendall, D.A.; Alexandrescu, A.T. NMR determination of pKa values in alpha-synuclein. Protein Sci. 2011, 20, 256–269. [Google Scholar] [CrossRef]
- Jha, S.; Snell, J.M.; Sheftic, S.R.; Patil, S.M.; Daniels, S.B.; Kolling, F.W.; Alexandrescu, A.T. pH dependence of amylin fibrillization. Biochemistry 2014, 53, 300–310. [Google Scholar] [CrossRef]
- Alexandrescu, A.T.; Mills, D.A.; Ulrich, E.L.; Chinami, M.; Markley, J.L. NMR assignments of the four histidines of staphylococcal nuclease in native and denatured states. Biochemistry 1988, 27, 2158–2165. [Google Scholar] [CrossRef]
- Berg, J.M.; Merkle, D.L. On the Metal Ion Specificity of “Zinc Finger” Proteins. J. Am. Chem. Soc. 1989, 111, 3759–3761. [Google Scholar] [CrossRef]
- Krizek, B.A.; Merkle, D.L.; Berg, J.M. Ligand Variation and Metal Ion Binding Specificity in Zinc Finger Peptides. Inorg. Chem. 1993, 32, 937–940. [Google Scholar] [CrossRef]
- Sivo, V.; D’Abrosca, G.; Russo, L.; Iacovino, R.; Pedone, P.V.; Fattorusso, R.; Isernia, C.; Malgieri, G. Co(II) Coordination in Prokaryotic Zinc Finger Domains as Revealed by UV-Vis Spectroscopy. Bioinorg. Chem. Appl. 2017, 2017, 1527247. [Google Scholar] [CrossRef] [PubMed]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the Expasy Server. In The Proteomics Protocols Handbook; Walker, J.M., Ed.; Humana Press: Totowa, NJ, USA, 2005. [Google Scholar]
- Orengo, C.A.; Michie, A.D.; Jones, S.; Jones, D.T.; Swindells, M.B.; Thornton, J.M. CATH--a hierarchic classification of protein domain structures. Structure 1997, 5, 1093–1108. [Google Scholar] [CrossRef] [PubMed]
- Hassel, K.R.; Brito-Estrada, O.; Makarewich, C.A. Microproteins: Overlooked regulators of physiology and disease. iScience 2023, 26, 106781. [Google Scholar] [CrossRef]
- Pennisi, E. ’Dark proteome’ survey reveals thousands of new human genes. Science 2024, 386, 951–952. [Google Scholar] [CrossRef]
- Li, Y.; Okatsu, K.; Fukai, S.; Sato, Y. Structural basis for specific recognition of K6-linked polyubiquitin chains by the TAB2 NZF domain. Biophys. J. 2021, 120, 3355–3362. [Google Scholar] [CrossRef]
- Loughlin, F.E.; Lukavsky, P.J.; Kazeeva, T.; Reber, S.; Hock, E.M.; Colombo, M.; Von Schroetter, C.; Pauli, P.; Clery, A.; Muhlemann, O.; et al. The Solution Structure of FUS Bound to RNA Reveals a Bipartite Mode of RNA Recognition with Both Sequence and Shape Specificity. Mol. Cell 2019, 73, 490–504.e496. [Google Scholar] [CrossRef]
- Baker, D.; Sali, A. Protein structure prediction and structural genomics. Science 2001, 294, 93–96. [Google Scholar] [CrossRef]
- Miura, T.; Satoh, T.; Takeuchi, H. Role of metal-ligand coordination in the folding pathway of zinc finger peptides. Biochim. Biophys. Acta 1998, 1384, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhang, J.; Wang, J.; Wang, W. Metal-coupled folding of Cys2His2 zinc-finger. J. Am. Chem. Soc. 2008, 130, 892–900. [Google Scholar] [CrossRef]
- Cordier, F.; Vinolo, E.; Veron, M.; Delepierre, M.; Agou, F. Solution structure of NEMO zinc finger and impact of an anhidrotic ectodermal dysplasia with immunodeficiency-related point mutation. J. Mol. Biol. 2008, 377, 1419–1432. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.J.; Cram, E.D.; Czolij, R.; Matthews, J.M.; Crossley, M.; Mackay, J.P. CCHX zinc finger derivatives retain the ability to bind Zn(II) and mediate protein-DNA interactions. J. Biol. Chem. 2003, 278, 28011–28018. [Google Scholar] [CrossRef]
- Shortle, D. Propensities, probabilities, and the Boltzmann hypothesis. Protein Sci. 2003, 12, 1298–1302. [Google Scholar] [CrossRef]
- Dolgikh, D.A.; Gilmanshin, R.I.; Brazhnikov, E.V.; Bychkova, V.E.; Semisotnov, G.V.; Venyaminov, S.; Ptitsyn, O.B. Alpha-Lactalbumin: Compact state with fluctuating tertiary structure? FEBS Lett. 1981, 136, 311–315. [Google Scholar] [CrossRef]
- Petsko, G.A. An idea whose time has gone. Genome Biol. 2007, 8, 107. [Google Scholar] [CrossRef] [PubMed]
- Yaseen, N.R.; Blobel, G. Two distinct classes of Ran-binding sites on the nucleoporin Nup-358. Proc. Natl. Acad. Sci. USA 1999, 96, 5516–5521. [Google Scholar] [CrossRef]
- Higa, M.M.; Alam, S.L.; Sundquist, W.I.; Ullman, K.S. Molecular characterization of the Ran-binding zinc finger domain of Nup153. J. Biol. Chem. 2007, 282, 17090–17100. [Google Scholar] [CrossRef] [PubMed]
- Sukegawa, J.; Blobel, G. A nuclear pore complex protein that contains zinc finger motifs, binds DNA, and faces the nucleoplasm. Cell 1993, 72, 29–38. [Google Scholar] [CrossRef]
- Garcia-Barcena, C.; Osinalde, N.; Ramirez, J.; Mayor, U. How to Inactivate Human Ubiquitin E3 Ligases by Mutation. Front. Cell Dev. Biol. 2020, 8, 39. [Google Scholar] [CrossRef]
- Persikov, A.V.; Singh, M. De novo prediction of DNA-binding specificities for Cys2His2 zinc finger proteins. Nucleic Acids Res. 2014, 42, 97–108. [Google Scholar] [CrossRef]
- Schwabe, J.W.; Rhodes, D. Beyond zinc fingers: Steroid hormone receptors have a novel structural motif for DNA recognition. Trends Biochem. Sci. 1991, 16, 291–296. [Google Scholar] [CrossRef]
- Ghosh, S.; Chatterji, D. Two zinc finger proteins from Mycobacterium smegmatis: DNA binding and activation of transcription. Genes Cells 2017, 22, 699–714. [Google Scholar] [CrossRef] [PubMed]
- Loughlin, F.E.; Mansfield, R.E.; Vaz, P.M.; McGrath, A.P.; Setiyaputra, S.; Gamsjaeger, R.; Chen, E.S.; Morris, B.J.; Guss, J.M.; Mackay, J.P. The zinc fingers of the SR-like protein ZRANB2 are single-stranded RNA-binding domains that recognize 5’ splice site-like sequences. Proc. Natl. Acad. Sci. USA 2009, 106, 5581–5586. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harris, R.E.; Rua, A.J.; Alexandrescu, A.T. Zinc-Induced Folding and Solution Structure of the Eponymous Novel Zinc Finger from the ZC4H2 Protein. Biomolecules 2025, 15, 1091. https://doi.org/10.3390/biom15081091
Harris RE, Rua AJ, Alexandrescu AT. Zinc-Induced Folding and Solution Structure of the Eponymous Novel Zinc Finger from the ZC4H2 Protein. Biomolecules. 2025; 15(8):1091. https://doi.org/10.3390/biom15081091
Chicago/Turabian StyleHarris, Rilee E., Antonio J. Rua, and Andrei T. Alexandrescu. 2025. "Zinc-Induced Folding and Solution Structure of the Eponymous Novel Zinc Finger from the ZC4H2 Protein" Biomolecules 15, no. 8: 1091. https://doi.org/10.3390/biom15081091
APA StyleHarris, R. E., Rua, A. J., & Alexandrescu, A. T. (2025). Zinc-Induced Folding and Solution Structure of the Eponymous Novel Zinc Finger from the ZC4H2 Protein. Biomolecules, 15(8), 1091. https://doi.org/10.3390/biom15081091