

Macrophage Reprogramming: Emerging Molecular Therapeutic Strategies for Nephrolithiasis

 ,
,
Abstract
1. Introduction
2. Macrophage Polarization: A Key Mechanism in the Nephrolithiasis
2.1. Phenotypic Characteristics of Macrophages
2.2. Macrophage Activation and Polarization in Nephrolithiasis
2.3. The Balance of Pro-/Anti-Inflammatory Polarization of Macrophages Plays a Key Role in Nephrolithiasis
3. Molecular Therapeutic Translation Potential for Macrophage Reprogramming in Nephrolithiasis
3.1. Direct Regulatory Strategies Targeting Macrophage Polarization
3.1.1. Direct Regulation of Targeted Chemokine Network and Cytokines
3.1.2. Targeting Key Signaling Pathways
3.1.3. Epigenetic Reprogramming
3.2. Anti-Inflammatory and Antioxidant Treatments Indirectly Regulate Macrophage Polarization
3.2.1. Anti-Inflammatory Drugs Derived from Natural Products
3.2.2. Novel Biomaterials and Cell Therapy Strategies
4. Future Directions of Nephrolithiasis Research
4.1. Novel Technologies and Multi-Omics Integration
4.1.1. Single-Cell and Spatial Omics for Deciphering Macrophage Heterogeneity
4.1.2. Integrative Metabolomics–Immune Microenvironment Profiling
4.2. Precision Therapeutic Targeting of the Immune Microenvironment
4.2.1. Macrophage-Targeted Nanodelivery Systems
4.2.2. Epigenetic Modulation of Macrophage Reprogramming
4.2.3. Genetically Engineered Macrophage Reprogramming via Precision Editing
5. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CaOx | Calcium Oxalate |
| TLR4-NF-κB | Toll-like Receptor 4-Nuclear Factor Kappa B |
| NLRP3 | NOD-like Receptor Family Pyrin Domain-containing Protein 3 |
| PPAR-γ | Peroxisome Proliferator-Activated Receptor Gamma |
| CCR2 | C-C Chemokine Receptor Type 2 |
| CSF-1 | Colony-Stimulating Factor 1 |
| STAT6 | Signal Transducer and Activator of Transcription 6 |
| HDAC | Histone Deacetylase |
| NF-κB | Nuclear Factor Kappa B |
| miR | MicroRNA |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| CAR-M | Chimeric Antigen Receptor Macrophage |
| scRNA-seq | Single-Cell RNA Sequencing |
| AKI | Acute Kidney Injury |
| IRG1 | Immune-Responsive Gene 1 |
| TGFβ | Transforming Growth Factor Beta |
| CCL2 | Chemokine (C-C Motif) Ligand 2 |
| MCP-1 | Monocyte Chemoattractant Protein-1 |
| IL | Interleukin |
| TNF-α | Tumor Necrosis Factor Alpha |
| ROS | Reactive Oxygen Species |
| IFN-γ | Interferon Gamma |
| LPS | Lipopolysaccharide |
| IRF1 | Interferon Regulatory Factor 1 |
| PKNOX1 | PKNOX Family Member 1 |
| CAT | Catalase |
| SOD | Superoxide Dismutase |
References
- Zeng, J.; Wang, S.; Zhong, L.; Huang, Z.; Zeng, Y.; Zheng, D.; Zou, W.; Lai, H. A Retrospective Study of Kidney Stone Recurrence in Adults. J. Clin. Med. Res. 2019, 11, 208–212. [Google Scholar] [CrossRef]
- Li, J.K.; Teoh, J.Y.; Ng, C.F. Updates in Endourological Management of Urolithiasis. Int. J. Urol. 2019, 26, 172–183. [Google Scholar] [CrossRef]
- Monga, M.; Murphy, M.; Paranjpe, R.; Cutone, B.; Eisner, B. Prevalence of Stone Disease and Procedure Trends in the United States. Urology 2023, 176, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.; Mai, Z.; Xia, S.; Wang, Z.; Zhang, K.; Wang, L.; Long, Y.; Ma, J.; Li, Y.; Wan, S.P.; et al. Prevalence of Kidney Stones in China: An Ultrasonography Based Cross-Sectional Study. BJU Int. 2017, 120, 109–116. [Google Scholar] [CrossRef]
- Fontenelle, L.F.; Sarti, T.D. Kidney Stones: Treatment and Prevention. Am. Fam. Physician 2019, 99, 490–496. [Google Scholar]
- Peerapen, P.; Thongboonkerd, V. Kidney Stone Prevention. Adv. Nutr. 2023, 14, 555–569. [Google Scholar] [CrossRef]
- Thongboonkerd, V.; Yasui, T.; Khan, S.R. Editorial: Immunity and Inflammatory Response in Kidney Stone Disease. Front. Immunol. 2021, 12, 795559. [Google Scholar] [CrossRef]
- Di, X.; Liu, S.; Xiang, L.; Jin, X. Association between the Systemic Immune-Inflammation Index and Kidney Stone: A Cross-Sectional Study of NHANES 2007–2018. Front. Immunol. 2023, 14, 1116224. [Google Scholar] [CrossRef] [PubMed]
- Canela, V.H.; Bowen, W.S.; Ferreira, R.M.; Syed, F.; Lingeman, J.E.; Sabo, A.R.; Barwinska, D.; Winfree, S.; Lake, B.B.; Cheng, Y.H.; et al. A Spatially Anchored Transcriptomic Atlas of the Human Kidney Papilla Identifies Significant Immune Injury in Patients with Stone Disease. Nat. Commun. 2023, 14, 4140. [Google Scholar] [CrossRef]
- Liu, H.; Ye, T.; Yang, X.; Liu, J.; Jiang, K.; Lu, H.; Xia, D.; Peng, E.; Chen, Z.; Sun, F.; et al. H19 Promote Calcium Oxalate Nephrocalcinosis-Induced Renal Tubular Epithelial Cell Injury via a ceRNA Pathway. EBioMedicine 2019, 50, 366–378. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Wang, W.; Peck, A.B.; Khan, S.R. Activation of the NLRP3 Inflammasome in Association with Calcium Oxalate Crystal Induced Reactive Oxygen Species in Kidneys. J. Urol. 2015, 193, 1684–1691. [Google Scholar] [CrossRef]
- Khan, S.R.; Canales, B.K.; Dominguez-Gutierrez, P.R. Randall’s Plaque and Calcium Oxalate Stone Formation: Role for Immunity and Inflammation. Nat. Rev. Nephrol. 2021, 17, 417–433. [Google Scholar] [CrossRef]
- Taguchi, K.; Okada, A.; Unno, R.; Hamamoto, S.; Yasui, T. Macrophage Function in Calcium Oxalate Kidney Stone Formation: A Systematic Review of Literature. Front. Immunol. 2021, 12, 673690. [Google Scholar] [CrossRef]
- Patel, M.; Yarlagadda, V.; Adedoyin, O.; Saini, V.; Assimos, D.G.; Holmes, R.P.; Mitchell, T. Oxalate Induces Mitochondrial Dysfunction and Disrupts Redox Homeostasis in a Human Monocyte Derived Cell Line. Redox Biol. 2018, 15, 207–215. [Google Scholar] [CrossRef]
- Hamamoto, S.; Taguchi, K.; Fujii, Y. [Molecular mechanism of renal stone formation]. Clin. Calcium 2011, 21, 1481–1487. [Google Scholar] [PubMed]
- Zhang, P.; Amarasinghe, H.E.; Whalley, J.P.; Tay, C.; Fang, H.; Migliorini, G.; Brown, A.C.; Allcock, A.; Scozzafava, G.; Rath, P.; et al. Epigenomic Analysis Reveals a Dynamic and Context-Specific Macrophage Enhancer Landscape Associated with Innate Immune Activation and Tolerance. Genome Biol. 2022, 23, 136. [Google Scholar] [CrossRef]
- Li, J.; Srivastava, R.M.; Ettyreddy, A.; Ferris, R.L. Cetuximab Ameliorates Suppressive Phenotypes of Myeloid Antigen Presenting Cells in Head and Neck Cancer Patients. J. Immunother. Cancer 2015, 3, 54. [Google Scholar] [CrossRef] [PubMed]
- Van den Bossche, J.; Baardman, J.; Otto, N.A.; van der Velden, S.; Neele, A.E.; van den Berg, S.M.; Luque-Martin, R.; Chen, H.-J.; Boshuizen, M.C.S.; Ahmed, M.; et al. Mitochondrial Dysfunction Prevents Repolarization of Inflammatory Macrophages. Cell Rep. 2016, 17, 684–696. [Google Scholar] [CrossRef]
- Kang, H.; Yang, B.; Zhang, K.; Pan, Q.; Yuan, W.; Li, G.; Bian, L. Immunoregulation of Macrophages by Dynamic Ligand Presentation via Ligand–Cation Coordination. Nat. Commun. 2019, 10, 1696. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Li, C.; Wu, Y.; Shen, L.; Ma, J.; Qian, J.; Ge, J. Role of KCa3.1 Channels in Macrophage Polarization and Its Relevance in Atherosclerotic Plaque Instability. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 226–236. [Google Scholar] [CrossRef]
- Taguchi, K.; Okada, A.; Hamamoto, S.; Unno, R.; Moritoki, Y.; Ando, R.; Mizuno, K.; Tozawa, K.; Kohri, K.; Yasui, T. M1/M2-Macrophage Phenotypes Regulate Renal Calcium Oxalate Crystal Development. Sci. Rep. 2016, 6, 35167. [Google Scholar] [CrossRef]
- Tian, F.; Chen, H.; Zhang, J.; He, W. Reprogramming Metabolism of Macrophages as a Target for Kidney Dysfunction Treatment in Autoimmune Diseases. Int. J. Mol. Sci. 2022, 23, 8024. [Google Scholar] [CrossRef]
- Chen, H.; Liu, N.; Zhuang, S. Macrophages in Renal Injury, Repair, Fibrosis Following Acute Kidney Injury and Targeted Therapy. Front. Immunol. 2022, 13, 934299. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Tian, J.; Tang, X.; Ma, J.; Wang, S. Long Non-Coding RNAs in the Regulation of Myeloid Cells. J. Hematol. Oncol. 2016, 9, 99. [Google Scholar] [CrossRef]
- Alexander, K.A.; Flynn, R.; Lineburg, K.E.; Kuns, R.D.; Teal, B.E.; Olver, S.D.; Lor, M.; Raffelt, N.C.; Koyama, M.; Leveque, L.; et al. CSF-1–Dependant Donor-Derived Macrophages Mediate Chronic Graft-versus-Host Disease. J. Clin. Investig. 2014, 124, 4266–4280. [Google Scholar] [CrossRef]
- Park, M.D.; Silvin, A.; Ginhoux, F.; Merad, M. Macrophages in Health and Disease. Cell 2022, 185, 4259–4279. [Google Scholar] [CrossRef]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Cao, Y.; Zhu, Q.; Wang, X.; Cheng, G.; Wang, Q.; He, R.; Lu, H.; Weng, Y.; Mao, G.; et al. Renal Macrophages Monitor and Remove Particles from Urine to Prevent Tubule Obstruction. Immunity 2024, 57, 106–123.e7. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.R.; Pearle, M.S.; Robertson, W.G.; Gambaro, G.; Canales, B.K.; Doizi, S.; Traxer, O.; Tiselius, H.-G. Kidney Stones. Nat. Rev. Dis. Primers 2016, 2, 16008. [Google Scholar] [CrossRef]
- Perazella, M.A.; Herlitz, L.C. The Crystalline Nephropathies. Kidney Int. Rep. 2021, 6, 2942–2957. [Google Scholar] [CrossRef]
- Khan, S.R. Reactive Oxygen Species as the Molecular Modulators of Calcium Oxalate Kidney Stone Formation: Evidence from Clinical and Experimental Investigations. J. Urol. 2013, 189, 803–811. [Google Scholar] [CrossRef]
- Kumar, P.; Yang, Z.; Lever, J.M.; Chávez, M.D.; Fatima, H.; Crossman, D.K.; Maynard, C.L.; George, J.F.; Mitchell, T. Hydroxyproline Stimulates Inflammation and Reprograms Macrophage Signaling in a Rat Kidney Stone Model. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166442. [Google Scholar] [CrossRef]
- Kusmartsev, S.; Dominguez-Gutierrez, P.R.; Canales, B.K.; Bird, V.G.; Vieweg, J.; Khan, S.R. Calcium Oxalate Stone Fragment and Crystal Phagocytosis by Human Macrophages. J. Urol. 2016, 195, 1143–1151. [Google Scholar] [CrossRef]
- Dejban, P.; Wilson, E.M.; Jayachandran, M.; Herrera Hernandez, L.P.; Haskic, Z.; Wellik, L.E.; Sinha, S.; Rule, A.D.; Denic, A.; Koo, K.; et al. Inflammatory Cells in Nephrectomy Tissue from Patients without and with a History of Urinary Stone Disease. CJASN 2022, 17, 414–422. [Google Scholar] [CrossRef]
- Capolongo, G.; Ferraro, P.M.; Unwin, R. Inflammation and Kidney Stones: Cause and Effect? Curr. Opin. Urol. 2023, 33, 129. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Liu, H.; Ye, T.; Duan, C.; Lv, P.; Wu, X.; Liu, J.; Jiang, K.; Lu, H.; Yang, H.; et al. AhR Activation Attenuates Calcium Oxalate Nephrocalcinosis by Diminishing M1 Macrophage Polarization and Promoting M2 Macrophage Polarization. Theranostics 2020, 10, 12011–12025. [Google Scholar] [CrossRef]
- Zhu, W.; Zhao, Z.; Chou, F.; Zuo, L.; Liu, T.; Yeh, S.; Bushinsky, D.; Zeng, G.; Chang, C. Loss of the Androgen Receptor Suppresses Intrarenal Calcium Oxalate Crystals Deposition via Altering Macrophage Recruitment/M2 Polarization with Change of the miR-185-5p/CSF-1 Signals. Cell Death Dis. 2019, 10, 275. [Google Scholar] [CrossRef] [PubMed]
- Wigner, P.; Grębowski, R.; Bijak, M.; Szemraj, J.; Saluk-Bijak, J. The Molecular Aspect of Nephrolithiasis Development. Cells 2021, 10, 1926. [Google Scholar] [CrossRef]
- Zhu, W.; Qiong, D.; Changzhi, X.; Meiyu, J.; Hui, L. Macrophage Polarization Regulation Shed Lights on Immunotherapy for CaOx Kidney Stone Disease. Biomed. Pharmacother. 2024, 179, 117336. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shi, M.; He, X.; Cao, Y.; Liu, P.; Li, F.; Zou, S.; Wen, C.; Zhan, Q.; Xu, Z.; et al. LncRNA-PACERR Induces pro-Tumour Macrophages via Interacting with miR-671-3p and m6A-Reader IGF2BP2 in Pancreatic Ductal Adenocarcinoma. J. Hematol. Oncol. 2022, 15, 52. [Google Scholar] [CrossRef]
- Sun, J.; Park, C.; Guenthner, N.; Gurley, S.; Zhang, L.; Lubben, B.; Adebayo, O.; Bash, H.; Chen, Y.; Maksimos, M.; et al. Tumor-Associated Macrophages in Multiple Myeloma: Advances in Biology and Therapy. J. Immunother. Cancer 2022, 10, e003975. [Google Scholar] [CrossRef]
- Wang, X.; Xie, L.; Liu, C. CCR2 Antagonist Attenuates Calcium Oxalate-Induced Kidney Oxidative Stress and Inflammation by Regulating Macrophage Activation. Exp. Anim. 2024, 73, 211–222. [Google Scholar] [CrossRef]
- Taguchi, K.; Okada, A.; Kitamura, H.; Yasui, T.; Naiki, T.; Hamamoto, S.; Ando, R.; Mizuno, K.; Kawai, N.; Tozawa, K.; et al. Colony-Stimulating Factor-1 Signaling Suppresses Renal Crystal Formation. J. Am. Soc. Nephrol. 2014, 25, 1680–1697. [Google Scholar] [CrossRef]
- Kumar, P.; Laurence, E.; Crossman, D.K.; Assimos, D.G.; Murphy, M.P.; Mitchell, T. Oxalate Disrupts Monocyte and Macrophage Cellular Function via Interleukin-10 and Mitochondrial Reactive Oxygen Species (ROS) Signaling. Redox Biol. 2023, 67, 102919. [Google Scholar] [CrossRef]
- Chen, Z.; Yuan, P.; Sun, X.; Tang, K.; Liu, H.; Han, S.; Ye, T.; Liu, X.; Yang, X.; Zeng, J.; et al. Pioglitazone Decreased Renal Calcium Oxalate Crystal Formation by Suppressing M1 Macrophage Polarization via the PPAR-γ-miR-23 Axis. Am. J. Physiol.-Ren. Physiol. 2019, 317, F137–F151. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, Y.; Kang, J.; He, Z.; Liu, Q.; Wu, J.; Li, D.; Wang, X.; Tao, Z.; Guan, X.; et al. Role of ROS-Induced NLRP3 Inflammasome Activation in the Formation of Calcium Oxalate Nephrolithiasis. Front. Immunol. 2022, 13, 818625. [Google Scholar] [CrossRef]
- Anders, H.-J.; Suarez-Alvarez, B.; Grigorescu, M.; Foresto-Neto, O.; Steiger, S.; Desai, J.; Marschner, J.A.; Honarpisheh, M.; Shi, C.; Jordan, J.; et al. The Macrophage Phenotype and Inflammasome Component NLRP3 Contributes to Nephrocalcinosis-Related Chronic Kidney Disease Independent from IL-1-Mediated Tissue Injury. Kidney Int. 2018, 93, 656–669. [Google Scholar] [CrossRef]
- Duan, C.; Liu, H.; Yang, X.; Liu, J.; Deng, Y.; Wang, T.; Xing, J.; Hu, Z.; Xu, H. Sirtuin1 Inhibits Calcium Oxalate Crystal-Induced Kidney Injury by Regulating TLR4 Signaling and Macrophage-Mediated Inflammatory Activation. Cell Signal 2023, 112, 110887. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yao, Z. Immune Cell Trafficking: A Novel Perspective on the Gut-Skin Axis. Inflamm. Regen. 2024, 44, 21. [Google Scholar] [CrossRef] [PubMed]
- Devarajan, A. Cross-Talk between Renal Lithogenesis and Atherosclerosis: An Unveiled Link between Kidney Stone Formation and Cardiovascular Diseases. Clin. Sci. 2018, 132, 615–626. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Yang, J.; Wu, W.; Amier, Y.; Li, X.; Wan, W.; Huang, Y.; Li, J.; Yu, X. Inflammatory Cytokines and Their Potential Role in Kidney Stone Disease: A Mendelian Randomization Study. Int. Urol. Nephrol. 2024, 56, 3249–3257. [Google Scholar] [CrossRef]
- Kusumi, K.; Ketz, J.; Saxena, V.; Spencer, J.D.; Safadi, F.; Schwaderer, A. Adolescents with Urinary Stones Have Elevated Urine Levels of Inflammatory Mediators. Urolithiasis 2019, 47, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Van Coillie, E.; Van Damme, J.; Opdenakker, G. The MCP/Eotaxin Subfamily of CC Chemokines. Cytokine Growth Factor. Rev. 1999, 10, 61–86. [Google Scholar] [CrossRef]
- Charo, I.F.; Ransohoff, R.M. The Many Roles of Chemokines and Chemokine Receptors in Inflammation. N. Engl. J. Med. 2006, 354, 610–621. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, Y.; Xia, R.; Wei, Y.; Wei, X. Role of the CCL2-CCR2 Signalling Axis in Cancer: Mechanisms and Therapeutic Targeting. Cell Prolif. 2021, 54, e13115. [Google Scholar] [CrossRef]
- Sehgal, A.; Irvine, K.M.; Hume, D.A. Functions of Macrophage Colony-Stimulating Factor (CSF1) in Development, Homeostasis, and Tissue Repair. Semin. Immunol. 2021, 54, 101509. [Google Scholar] [CrossRef]
- Poto, R.; Gambardella, A.R.; Marone, G.; Schroeder, J.T.; Mattei, F.; Schiavoni, G.; Varricchi, G. Basophils from Allergy to Cancer. Front. Immunol. 2022, 13, 1056838. [Google Scholar] [CrossRef]
- Cai, H.; Zhang, Y.; Wang, J.; Gu, J. Defects in Macrophage Reprogramming in Cancer Therapy: The Negative Impact of PD-L1/PD-1. Front. Immunol. 2021, 12, 690869. [Google Scholar] [CrossRef]
- Staples, K.J.; Smallie, T.; Williams, L.M.; Foey, A.; Burke, B.; Foxwell, B.M.J.; Ziegler-Heitbrock, L. IL-10 Induces IL-10 in Primary Human Monocyte-Derived Macrophages via the Transcription Factor Stat3. J. Immunol. 2007, 178, 4779–4785. [Google Scholar] [CrossRef]
- Okada, A.; Ando, R.; Taguchi, K.; Hamamoto, S.; Unno, R.; Sugino, T.; Tanaka, Y.; Mizuno, K.; Tozawa, K.; Kohri, K.; et al. Identification of New Urinary Risk Markers for Urinary Stones Using a Logistic Model and Multinomial Logit Model. Clin. Exp. Nephrol. 2019, 23, 710–716. [Google Scholar] [CrossRef]
- Li, Y.; Pan, Y.; Zhao, X.; Wu, S.; Li, F.; Wang, Y.; Liu, B.; Zhang, Y.; Gao, X.; Wang, Y.; et al. Peroxisome Proliferator-Activated Receptors: A Key Link between Lipid Metabolism and Cancer Progression. Clin. Nutr. 2024, 43, 332–345. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, J.; Luo, S.; Zhan, Y.; Lu, Q. The Roles of PPARγ and Its Agonists in Autoimmune Diseases: A Comprehensive Review. J. Autoimmun. 2020, 113, 102510. [Google Scholar] [CrossRef]
- Yuan, T.; Xia, Y.; Pan, S.; Li, B.; Ye, Z.; Yan, X.; Hu, W.; Li, L.; Song, B.; Yu, W.; et al. STAT6 Promoting Oxalate Crystal Deposition-Induced Renal Fibrosis by Mediating Macrophage-to-Myofibroblast Transition via Inhibiting Fatty Acid Oxidation. Inflamm. Res. 2023, 72, 2111–2126. [Google Scholar] [CrossRef]
- Song, Q.; Song, C.; Chen, X.; Xiong, Y.; Li, L.; Liao, W.; Xue, L.; Yang, S. FKBP5 Deficiency Attenuates Calcium Oxalate Kidney Stone Formation by Suppressing Cell-Crystal Adhesion, Apoptosis and Macrophage M1 Polarization via Inhibition of NF-κB Signaling. Cell Mol. Life Sci. 2023, 80, 301. [Google Scholar] [CrossRef]
- Abuduhalike, R.; Abudouwayiti, A.; Juan, S.; MaheMuti, A. Study on the Mechanism of NLRP3/IL-1/ NF-κB Signaling Pathway and Macrophage Polarization in the Occurrence and Development of VTE. Ann. Vasc. Surg. 2023, 89, 280–292. [Google Scholar] [CrossRef]
- Li, X.; Yan, X.; Wang, Y.; Kaur, B.; Han, H.; Yu, J. The Notch Signaling Pathway: A Potential Target for Cancer Immunotherapy. J. Hematol. Oncol. 2023, 16, 45. [Google Scholar] [CrossRef]
- Huang, F.; Zhao, J.-L.; Wang, L.; Gao, C.-C.; Liang, S.-Q.; An, D.-J.; Bai, J.; Chen, Y.; Han, H.; Qin, H.-Y. miR-148a-3p Mediates Notch Signaling to Promote the Differentiation and M1 Activation of Macrophages. Front. Immunol. 2017, 8, 1327. [Google Scholar] [CrossRef]
- Zhu, W.; Liu, Y.; Lan, Y.; Li, X.; Luo, L.; Duan, X.; Lei, M.; Liu, G.; Yang, Z.; Mai, X.; et al. Dietary Vinegar Prevents Kidney Stone Recurrence via Epigenetic Regulations. EBioMedicine 2019, 45, 231–250. [Google Scholar] [CrossRef]
- Ye, Z.; Sun, Y.; Yang, S.; Li, L.; Li, B.; Xia, Y.; Yuan, T.; Yu, W.; Chen, L.; Zhou, X.; et al. Lgals3 Promotes Calcium Oxalate Crystal Formation and Kidney Injury Through Histone Lactylation-Mediated FGFR4 Activation. Adv. Sci. 2025, 12, e2413937. [Google Scholar] [CrossRef]
- Zhu, W.; Liu, L.; Wu, J.; Gao, R.; Fu, L.; Yang, X.; Zou, Y.; Zhang, S.; Luo, D. SMYD3 Activates the TCA Cycle to Promote M1-M2 Conversion in Macrophages. Int. Immunopharmacol. 2024, 127, 111329. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Yang, J.; Wei, Y.; Wei, X. Epigenetic Regulation of Macrophages: From Homeostasis Maintenance to Host Defense. Cell Mol. Immunol. 2020, 17, 36–49. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, H.; Huang, S.; Yin, L.; Wang, F.; Luo, P.; Huang, H. Epigenetic Regulation in Cardiovascular Disease: Mechanisms and Advances in Clinical Trials. Signal Transduct. Target. Ther. 2022, 7, 200. [Google Scholar] [CrossRef]
- Kimball, A.S.; Davis, F.M.; denDekker, A.; Joshi, A.D.; Schaller, M.A.; Bermick, J.; Xing, X.; Burant, C.F.; Obi, A.T.; Nysz, D.; et al. The Histone Methyltransferase Setdb2 Modulates Macrophage Phenotype and Uric Acid Production in Diabetic Wound Repair. Immunity 2019, 51, 258–271.e5. [Google Scholar] [CrossRef] [PubMed]
- Willemsen, L.; Prange, K.H.M.; Neele, A.E.; van Roomen, C.P.A.A.; Gijbels, M.; Griffith, G.R.; den Toom, M.; Beckers, L.; Siebeler, R.; Spann, N.J.; et al. DOT1L Regulates Lipid Biosynthesis and Inflammatory Responses in Macrophages and Promotes Atherosclerotic Plaque Stability. Cell Rep. 2022, 41, 111703. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Karnam, K.; Mahale, A.; Sedmaki, K.; Krishna Venuganti, V.; Kulkarni, O.P. HDAC5 RNA Interference Ameliorates Acute Renal Injury by Upregulating KLF2 and Inhibiting NALP3 Expression in a Mouse Model of Oxalate Nephropathy. Int. Immunopharmacol. 2022, 112, 109264. [Google Scholar] [CrossRef]
- Xi, J.; Chen, Y.; Jing, J.; Zhang, Y.; Liang, C.; Hao, Z.; Zhang, L. Sirtuin 3 Suppresses the Formation of Renal Calcium Oxalate Crystals through Promoting M2 Polarization of Macrophages. J. Cell Physiol. 2019, 234, 11463–11473. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Xiang, T.; Guo, J.; Guo, F.; Wu, Y.; Feng, H.; Liu, J.; Tao, S.; Fu, P.; Ma, L. Inhibition of ACSS2-Mediated Histone Crotonylation Alleviates Kidney Fibrosis via IL-1β-Dependent Macrophage Activation and Tubular Cell Senescence. Nat. Commun. 2024, 15, 3200. [Google Scholar] [CrossRef]
- Li, Y.; Lu, X.; Yu, Z.; Wang, H.; Gao, B. Meta-Data Analysis of Kidney Stone Disease Highlights ATP1A1 Involvement in Renal Crystal Formation. Redox Biol. 2023, 61, 102648. [Google Scholar] [CrossRef]
- Daskalaki, M.G.; Tsatsanis, C.; Kampranis, S.C. Histone Methylation and Acetylation in Macrophages as a Mechanism for Regulation of Inflammatory Responses. J. Cell. Physiol. 2018, 233, 6495–6507. [Google Scholar] [CrossRef]
- Saradna, A.; Do, D.C.; Kumar, S.; Fu, Q.-L.; Gao, P. Macrophage Polarization and Allergic Asthma. Transl. Res. 2018, 191, 1–14. [Google Scholar] [CrossRef]
- Song, H.; Shen, R.; Liu, X.; Yang, X.; Xie, K.; Guo, Z.; Wang, D. Histone Post-Translational Modification and the DNA Damage Response. Genes Dis. 2023, 10, 1429–1444. [Google Scholar] [CrossRef]
- Liu, X.-Y.; Guo, C.-H.; Xi, Z.-Y.; Xu, X.-Q.; Zhao, Q.-Y.; Li, L.-S.; Wang, Y. Histone Methylation in Pancreatic Cancer and Its Clinical Implications. World J. Gastroenterol. 2021, 27, 6004–6024. [Google Scholar] [CrossRef]
- Wang, H.; Fan, Z.; Shliaha, P.V.; Miele, M.; Hendrickson, R.C.; Jiang, X.; Helin, K. H3K4me3 Regulates RNA Polymerase II Promoter-Proximal Pause-Release. Nature 2023, 615, 339–348. [Google Scholar] [CrossRef]
- Ming-Chin Lee, K.; Achuthan, A.A.; De Souza, D.P.; Lupancu, T.J.; Binger, K.J.; Lee, M.K.S.; Xu, Y.; McConville, M.J.; de Weerd, N.A.; Dragoljevic, D.; et al. Type I Interferon Antagonism of the JMJD3-IRF4 Pathway Modulates Macrophage Activation and Polarization. Cell Rep. 2022, 39, 110719. [Google Scholar] [CrossRef]
- Guo, S.; Zhao, J.; Zhang, Y.; Qin, Y.; Yuan, J.; Yu, Z.; Xing, Y.; Zhang, Y.; Hui, Y.; Wang, A.; et al. Histone Deacetylases: Potential Therapeutic Targets in Cisplatin-Induced Acute Kidney Injury. Ann. Med. 2024, 56, 2418958. [Google Scholar] [CrossRef]
- Yao, W.; Hu, X.; Wang, X. Crossing Epigenetic Frontiers: The Intersection of Novel Histone Modifications and Diseases. Signal Transduct. Target. Ther. 2024, 9, 232. [Google Scholar] [CrossRef] [PubMed]
- Cui, S.-N.; Chen, Z.-Y.; Yang, X.-B.; Chen, L.; Yang, Y.-Y.; Pan, S.-W.; Wang, Y.-X.; Xu, J.-Q.; Zhou, T.; Xiao, H.-R.; et al. Trichostatin A Modulates the Macrophage Phenotype by Enhancing Autophagy to Reduce Inflammation during Polymicrobial Sepsis. Int. Immunopharmacol. 2019, 77, 105973. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.-J.; Zhang, T.-N.; Chen, H.-H.; Yu, X.-F.; Lv, J.-L.; Liu, Y.-Y.; Liu, Y.-S.; Zheng, G.; Zhao, J.-Q.; Wei, Y.-F.; et al. The Sirtuin Family in Health and Disease. Signal Transduct. Target. Ther. 2022, 7, 402. [Google Scholar] [CrossRef]
- Zhang, K.; Jagannath, C. Crosstalk between Metabolism and Epigenetics during Macrophage Polarization. Epigenetics Chromatin 2025, 18, 16. [Google Scholar] [CrossRef]
- Song, B.-F.; Li, B.-J.; Ning, J.-Z.; Xia, Y.-Q.; Ye, Z.-H.; Yuan, T.-H.; Yan, X.-Z.; Li, L.; Zhou, X.-J.; Rao, T.; et al. Overexpression of Sirtuin 1 Attenuates Calcium Oxalate-Induced Kidney Injury by Promoting Macrophage Polarization. Int. Immunopharmacol. 2023, 121, 110398. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Kurosawa, M.; Kamiya, M.; Urano, Y.; Suzuki, A.; Yamamoto, K.; Hase, K.; Homma, K.; Sasaki, J.; Miyauchi, H.; et al. Macrophage Extracellular Trap Formation Promoted by Platelet Activation Is a Key Mediator of Rhabdomyolysis-Induced Acute Kidney Injury. Nat. Med. 2018, 24, 232–238. [Google Scholar] [CrossRef]
- Zhang, D.; Tang, Z.; Huang, H.; Zhou, G.; Cui, C.; Weng, Y.; Liu, W.; Kim, S.; Lee, S.; Perez-Neut, M.; et al. Metabolic Regulation of Gene Expression by Histone Lactylation. Nature 2019, 574, 575–580. [Google Scholar] [CrossRef]
- Mattei, A.L.; Bailly, N.; Meissner, A. DNA Methylation: A Historical Perspective. Trends Genet. 2022, 38, 676–707. [Google Scholar] [CrossRef]
- Miranda, T.B.; Jones, P.A. DNA Methylation: The Nuts and Bolts of Repression. J. Cell Physiol. 2007, 213, 384–390. [Google Scholar] [CrossRef]
- Yang, X.; Wang, X.; Liu, D.; Yu, L.; Xue, B.; Shi, H. Epigenetic Regulation of Macrophage Polarization by DNA Methyltransferase 3b. Mol. Endocrinol. 2014, 28, 565–574. [Google Scholar] [CrossRef] [PubMed]
- Qin, W.; Spek, C.A.; Scicluna, B.P.; van der Poll, T.; Duitman, J. Myeloid DNA Methyltransferase3b Deficiency Aggravates Pulmonary Fibrosis by Enhancing Profibrotic Macrophage Activation. Respir. Res. 2022, 23, 162. [Google Scholar] [CrossRef] [PubMed]
- Pan‡, W.; Yun, T.; Ouyang, X.; Ruan, Z.; Zhang, T.; An, Y.; Wang, R.; Zhu, P. A Blood-Based Multi-Omic Landscape for the Molecular Characterization of Kidney Stone Disease. Mol. Omics 2024, 20, 322–332. [Google Scholar] [CrossRef]
- Liu, H.; Yang, X.; Tang, K.; Ye, T.; Duan, C.; Lv, P.; Yan, L.; Wu, X.; Chen, Z.; Liu, J.; et al. Sulforaphane Elicts Dual Therapeutic Effects on Renal Inflammatory Injury and Crystal Deposition in Calcium Oxalate Nephrocalcinosis. Theranostics 2020, 10, 7319–7334. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Chen, D.; Yan, P.; Zheng, F.; Zhu, H.; Yuan, Z.; Yang, X.; Zuo, Y.; Chen, C.; Lu, H.; et al. Puerarin Suppresses Macrophage M1 Polarization to Alleviate Renal Inflammatory Injury through Antagonizing TLR4/MyD88-Mediated NF-κB P65 and JNK/FoxO1 Activation. Phytomedicine 2024, 132, 155813. [Google Scholar] [CrossRef]
- Deng, J.; Yu, B.; Chang, Z.; Wu, S.; Li, G.; Chen, W.; Li, S.; Duan, X.; Wu, W.; Sun, X.; et al. Cerium Oxide-Based Nanozyme Suppresses Kidney Calcium Oxalate Crystal Depositions via Reversing Hyperoxaluria-Induced Oxidative Stress Damage. J. Nanobiotechnol. 2022, 20, 516. [Google Scholar] [CrossRef]
- Tian, Y.; Ye, Z.; Wang, X.; Guan, H.; Liu, W.; Duan, X.; Liu, Y.; Zeng, G.; Liu, H. MOF-818 Nanozyme Suppresses Calcium Oxalate Kidney Stones by Alleviating Oxidative Stress and Inflammatory Injury. Adv. Healthcare Mater. 2024, 14, 2401574. [Google Scholar] [CrossRef]
- Ba, X.; Ye, T.; He, Y.; Tong, Y.; Shang, H.; Wu, J.; Deng, W.; Zhong, Z.; Yang, X.; Wang, K.; et al. Engineered Macrophage Membrane-Coated Nanoparticles Attenuate Calcium Oxalate Nephrocalcinosis-Induced Kidney Injury by Reducing Oxidative Stress and Pyroptosis. Acta Biomater. 2025, 195, 479–495. [Google Scholar] [CrossRef]
- Reiss, K.A.; Angelos, M.G.; Dees, E.C.; Yuan, Y.; Ueno, N.T.; Pohlmann, P.R.; Johnson, M.L.; Chao, J.; Shestova, O.; Serody, J.S.; et al. CAR-Macrophage Therapy for HER2-Overexpressing Advanced Solid Tumors: A Phase 1 Trial. Nat. Med. 2025, 31, 1171–1182. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Lee, M.; Kwon, S.; Kim, J.; Kwon, Y. Systematic and Mechanistic Analysis of AuNP-Induced Nanotoxicity for Risk Assessment of Nanomedicine. Nano Converg. 2022, 9, 27. [Google Scholar] [CrossRef]
- Ren, D.; Liu, M.; Cao, M.; Wang, S.; Fan, S.; Liu, M.; Zhang, J.; Sun, H. HPB@LA@PDA Nanoplatform Ameliorates Osteoarthritis by Scavenging Reactive Oxygen Species and Remodelling the Inflammatory Microenvironment: An In Vitro and In Vivo Study. Chem. Eng. J. 2025, 507, 160592. [Google Scholar] [CrossRef]
- Nguyen, V.K.; Nguyen, N.; Li, Z.; Cheng, C.; Wang, J.; Chiang, Y.; Song, H.; Lo, S.; Mac, C.; Chang, Y.; et al. Inflammation-activated Endogenous Macrophage-mediated Prodrug Delivery System Overcoming Biological Barriers for Enhanced Oral Meningitis Therapy. Adv. Funct. Mater. 2024, 34, 2401570. [Google Scholar] [CrossRef]
- Zhu, X.; Guo, Q.; Zou, J.; Wang, B.; Zhang, Z.; Wei, R.; Zhao, L.; Zhang, Y.; Chu, C.; Fu, X.; et al. MiR-19a-3p Suppresses M1 Macrophage Polarization by Inhibiting STAT1/IRF1 Pathway. Front. Pharmacol. 2021, 12, 614044. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhang, C.; Yang, S.; Yang, L.; Luo, W.; Zhang, W.; Zhang, X.; Chao, J. Macrophage-Derived MMP12 Promotes Fibrosis through Sustained Damage to Endothelial Cells. J. Hazard. Mater. 2024, 461, 132733. [Google Scholar] [CrossRef] [PubMed]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef]
- Grayson, P.C.; Eddy, S.; Taroni, J.N.; Lightfoot, Y.L.; Mariani, L.; Parikh, H.; Lindenmeyer, M.T.; Ju, W.; Greene, C.S.; Godfrey, B.; et al. Metabolic Pathways and Immunometabolism in Rare Kidney Diseases. Ann. Rheum. Dis. 2018, 77, 1226–1233. [Google Scholar] [CrossRef]
- Chen, Y.-J.; Li, G.-N.; Li, X.-J.; Wei, L.-X.; Fu, M.-J.; Cheng, Z.-L.; Yang, Z.; Zhu, G.-Q.; Wang, X.-D.; Zhang, C.; et al. Targeting IRG1 Reverses the Immunosuppressive Function of Tumor-Associated Macrophages and Enhances Cancer Immunotherapy. Sci. Adv. 2023, 9, eadg0654. [Google Scholar] [CrossRef]
- Jin, R.; Neufeld, L.; McGaha, T.L. Linking Macrophage Metabolism to Function in the Tumor Microenvironment. Nat. Cancer 2025, 6, 239–252. [Google Scholar] [CrossRef]
- Su, W.-P.; Chang, L.-C.; Song, W.-H.; Yang, L.-X.; Wang, L.-C.; Chia, Z.-C.; Chin, Y.-C.; Shan, Y.-S.; Huang, C.-C.; Yeh, C.-S. Polyaniline-Based Glyco-Condensation on Au Nanoparticles Enhances Immunotherapy in Lung Cancer. ACS Appl. Mater. Interfaces 2022, 14, 24144–24159. [Google Scholar] [CrossRef]
- Chen, C.; Liu, T.; Tang, Y.; Luo, G.; Liang, G.; He, W. Epigenetic Regulation of Macrophage Polarization in Wound Healing. Burn. Trauma 2023, 11, tkac057. [Google Scholar] [CrossRef]
- Placek, K.; Schultze, J.L.; Aschenbrenner, A.C. Epigenetic Reprogramming of Immune Cells in Injury, Repair, and Resolution. J. Clin. Investig. 2019, 129, 2994–3005. [Google Scholar] [CrossRef]
- Watson, N.; Kuppuswamy, S.; Ledford, W.L.; Sukumari-Ramesh, S. The Role of HDAC3 in Inflammation: Mechanisms and Therapeutic Implications. Front. Immunol. 2024, 15, 1419685. [Google Scholar] [CrossRef]
- Maalej, K.M.; Merhi, M.; Inchakalody, V.P.; Mestiri, S.; Alam, M.; Maccalli, C.; Cherif, H.; Uddin, S.; Steinhoff, M.; Marincola, F.M.; et al. CAR-Cell Therapy in the Era of Solid Tumor Treatment: Current Challenges and Emerging Therapeutic Advances. Mol. Cancer 2023, 22, 20. [Google Scholar] [CrossRef]
- Klichinsky, M.; Ruella, M.; Shestova, O.; Lu, X.M.; Best, A.; Zeeman, M.; Schmierer, M.; Gabrusiewicz, K.; Anderson, N.R.; Petty, N.E.; et al. Human Chimeric Antigen Receptor Macrophages for Cancer Immunotherapy. Nat. Biotechnol. 2020, 38, 947–953. [Google Scholar] [CrossRef]

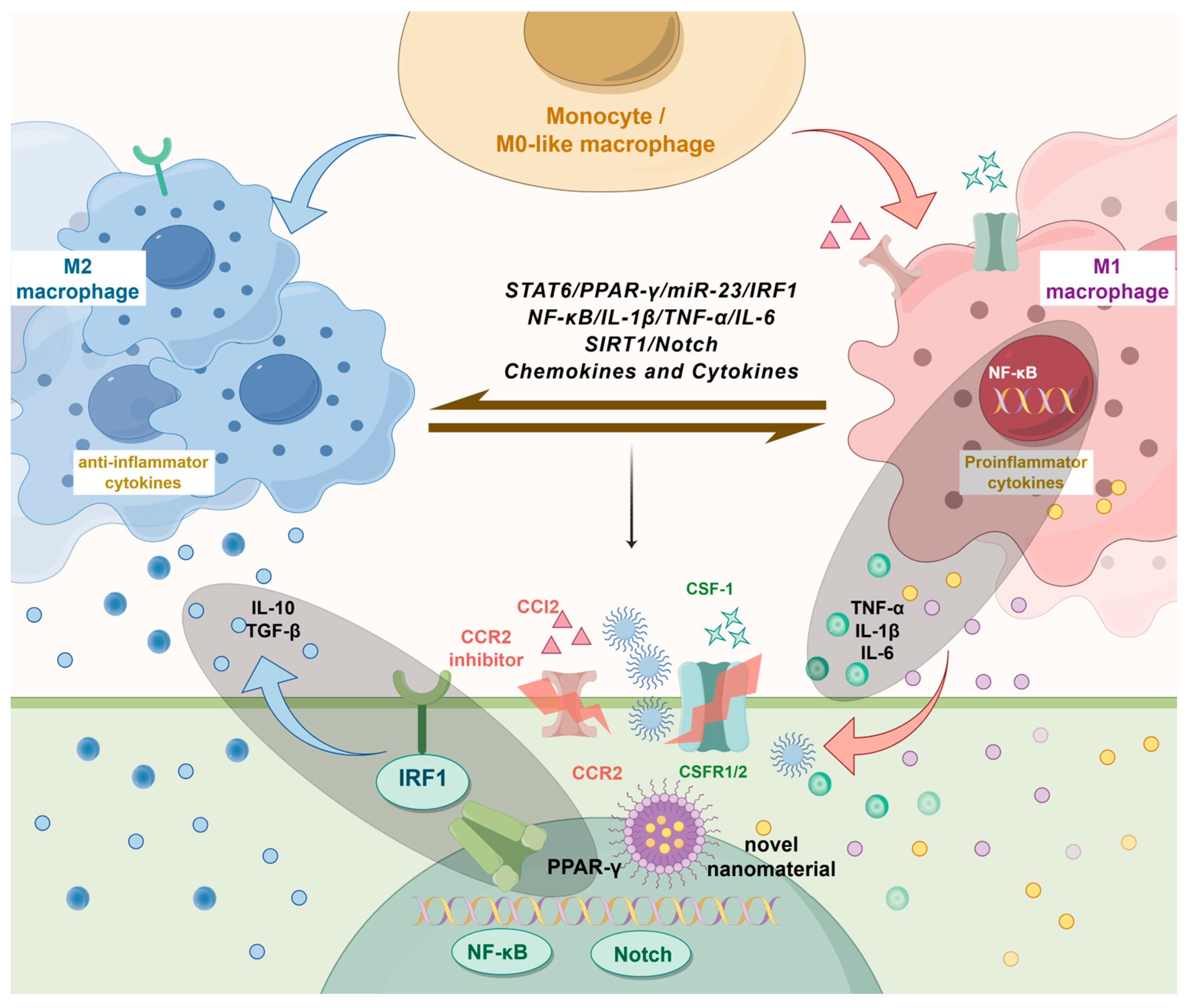

{kind=link}
{kind=link}
| Therapeutic Strategy | Specific Approach | Mechanism of Action | Key Molecules/Targets | Experimental Models | Primary Outcomes | References |
|---|---|---|---|---|---|---|
| Chemokine/Cytokine Regulation | CCR2 Antagonists | Inhibit CCL2/CCR2 axis, reduce M1 polarization, attenuate oxidative stress and inflammation | CCL2, CCR2, TNF-α, IL-6 | Glyoxylic acid mouse model/THP-1 cells, HK-2 cells | Reduced M1 polarization, tubular injury, and crystal deposition | [42] |
| Recombinant Human CSF-1 Therapy | Enhance M2 polarization, improve phagocytic function | CSF-1/CSF1R signaling pathway | Hyperoxaluric mouse model/BMDMs, RTCs | Increased M2 markers, decreased renal crystals | [43] | |
| IL-10 Treatment | Activate STAT3 signaling to promote M2 polarization | IL-10, STAT3 | High-oxalate-diet human/THP-1 cells | Reversed M1 polarization, reduced crystal deposition and inflammation | [44] | |
| Key Signaling Pathway Targeting | PPAR-γ Agonists (e.g., Pioglitazone) | Bind PPAR-γ to upregulate miR-23, suppress IRF1/Pknox1 and shift M1→M2 polarization | PPAR-γ, miR-23, IRF1, Pknox1 | Glyoxylic acid mouse model/BMDMs | Reduced M1-dominant inflammation, decreased crystal formation | [45] |
| NF-κB–NLRP3–IL-1β Feed-Forward Cascade Inhibitors (e.g., FKBP5 Deficiency, NLRP3 Deficiency) | Suppress NF-κB signaling to inhibit M1 polarization, reduce TNF-α/IL-1β secretion | NF-κB, FKBP5, NLRP3, TNF-α, IL-1β | Glyoxylic acid mouse model/BMDMs | Attenuated M1 activation, crystal aggregation, and tubular injury | [46,47] | |
| Notch Signaling Inhibitors (Sirt1 Overexpression) | Inhibit Notch pathway to promote M2 polarization, reduce renal epithelial cell apoptosis | Notch, Sirt1, TLR4 | Glyoxylic acid mouse model/BMDMs, TECs | Enhanced M2 phenotype, reduced crystal deposition and renal fibrosis | [48] |
| Therapeutic Strategy | Specific Approach | Mechanism of Action | Key Molecules/Targets | Experimental Models | Primary Outcomes | References |
|---|---|---|---|---|---|---|
| Epigenetic Histone Reprogramming | Histone methyltransferases | Suppress inflammatory cytokine gene transcription or control lipid gene synthesis through histone methylation modifications (e.g., Setdb2, DOTL1) | Setdb2, H3K9me3, NF-κB, DOT1L, H3K79me, Srebf1/2 | Diabetic (db/db) mouse model/BMDMs | Shifted macrophages to a reparative phenotype, reduced inflammation | [73,74] |
| HDACi, SIRTs | Enhance histone acetylation, promote M2 polarization (e.g., TSA SIRT1/2/3/5/6) | HDAC5, SIRT1/3, NOTCH, FOXO1 | Sodium oxalate mouse model/TCMK-1 cells | Reduced M1 polarization, improved M2 reparative phenotype, decreased fibrosis | [75,76] | |
| ACSS2 inhibitor | Lower the level of histone H3 lysine 9 crotonylation (H3K9cr) to reduce IL-1β-dependent macrophage activation | ACSS2, H3K9cr, IL-1β | UUO mouse model/RAW264.7 cells, TCMK-1 cells | Reduced M1 polarization, delayed tubular cell senescence and the progression of fibrosis | [77] | |
| CpG Methylation Intervention | DNA methyltransferases | Inhibit DNMT3b, promote M2 polarization via PPARγ1 methylation | DNMT3b, PPARγ1 | Hydroxyproline rats model/HK-2 cells | Alleviated oxidative stress, crystal adhesion, and stone formation | [78] |
| Therapeutic Strategy | Specific Approach | Mechanism of Action | Key Molecules/Targets | Experimental Models | Primary Outcomes | References |
|---|---|---|---|---|---|---|
| Anti-Inflammatory and Antioxidant Therapies | Natural Products (e.g., SFN, Puerarin) | Suppress TLR4/NF-κB pathway, reduce M1 polarization, enhance Nrf2-mediated antioxidant activity | TLR4, NF-κB, Nrf2, miR-93-5p | Glyoxylic acid mouse model/BMDMs, TECs | Reduced M1 activation, oxidative stress, and crystal adhesion | [98,99] |
| Nanotechnology and Biomaterials | ROS-Responsive Nanoparticles (e.g., CNPs, MOF-818) | Scavenge ROS, promote M1-to-M2 polarization, modulate inflammatory microenvironment | ROS, CAT, SOD, NLRP3 inflammasome | Ethylene glycol rat model, nude mice/HK-2 cells, NRK-52E cells, NRK-49F cells, MDCK cells | Reduced ROS, improved renal function, decreased crystal deposition | [100,101] |
| Biomimetic Nanoparticles (KM@M@M) | Target injured tubules, suppress NLRP3 inflammasome and ROS production | Kim-1 peptide, NLRP3, MnO2 | Glyoxylic acid mouse model/RAW 264.7 cells, HK-2 cells | Reduced pyroptosis, crystal burden, and tubular injury | [102] | |
| Gene Editing and Cellular Therapy | CRISPR-Cas9 | Target genes involved in M1/M2 polarization (e.g., CSF-1, FKBP5) | / | Glyoxylic acid mouse model | Precisely regulated macrophage phenotype, potential for anti-inflammatory shift | [43,46] |
| CAR-M Therapy (Conceptual Adaptation for Nephrolithiasis) | Engineer macrophages to express antigen receptors, enhance M1 polarization or phagocytosis | HER2 | Phase I clinical trial for solid tumors | Migrated to lesions in solid tumors and activated coordinated CD8+ T-cell-mediated immune surveillance | [103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, M.; Jia, Y.; Zhang, S.; Zou, B.; Ying, Z.; Gao, X.; Fang, Z.; Gao, X. Macrophage Reprogramming: Emerging Molecular Therapeutic Strategies for Nephrolithiasis. Biomolecules 2025, 15, 1090. https://doi.org/10.3390/biom15081090
Shu M, Jia Y, Zhang S, Zou B, Ying Z, Gao X, Fang Z, Gao X. Macrophage Reprogramming: Emerging Molecular Therapeutic Strategies for Nephrolithiasis. Biomolecules. 2025; 15(8):1090. https://doi.org/10.3390/biom15081090
Chicago/Turabian StyleShu, Meng, Yiying Jia, Shuwei Zhang, Bangyu Zou, Zhaoxin Ying, Xu Gao, Ziyu Fang, and Xiaofeng Gao. 2025. "Macrophage Reprogramming: Emerging Molecular Therapeutic Strategies for Nephrolithiasis" Biomolecules 15, no. 8: 1090. https://doi.org/10.3390/biom15081090
APA StyleShu, M., Jia, Y., Zhang, S., Zou, B., Ying, Z., Gao, X., Fang, Z., & Gao, X. (2025). Macrophage Reprogramming: Emerging Molecular Therapeutic Strategies for Nephrolithiasis. Biomolecules, 15(8), 1090. https://doi.org/10.3390/biom15081090