
Predictive and Prognostic Relevance of ABC Transporters for Resistance to Anthracycline Derivatives

 , , and
, , and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Cell Lines
2.2. Drugs
2.3. Expression of ABC Transporter Genes
2.4. Molecular Docking
2.5. Hierarchical Cluster Analysis and Statistical Methods
2.6. Resazurin Cell Viability Assay
2.7. Live Cell Time-Lapse Microscopy
2.8. Kaplan–Meier Survival Statistics
3. Results
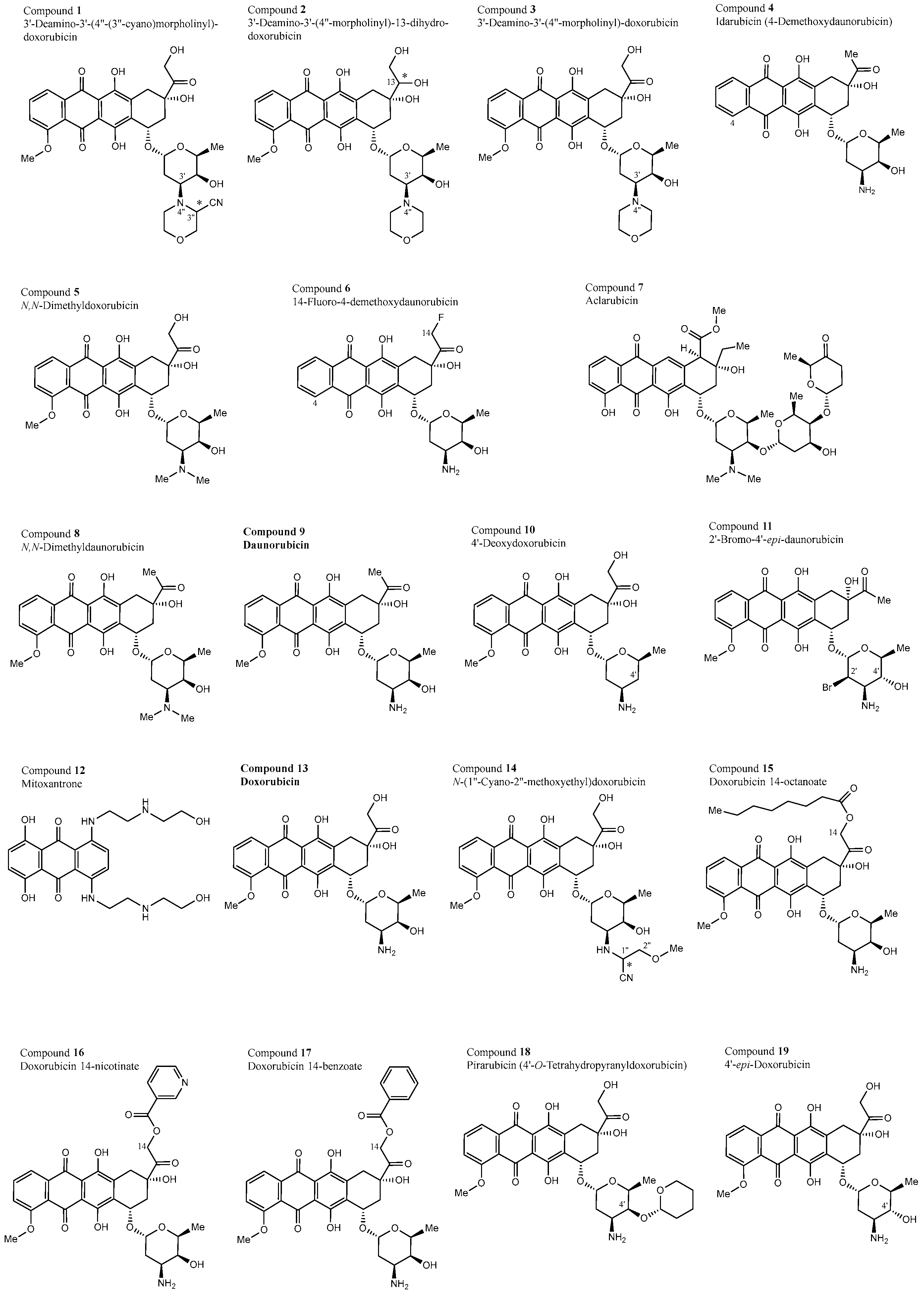
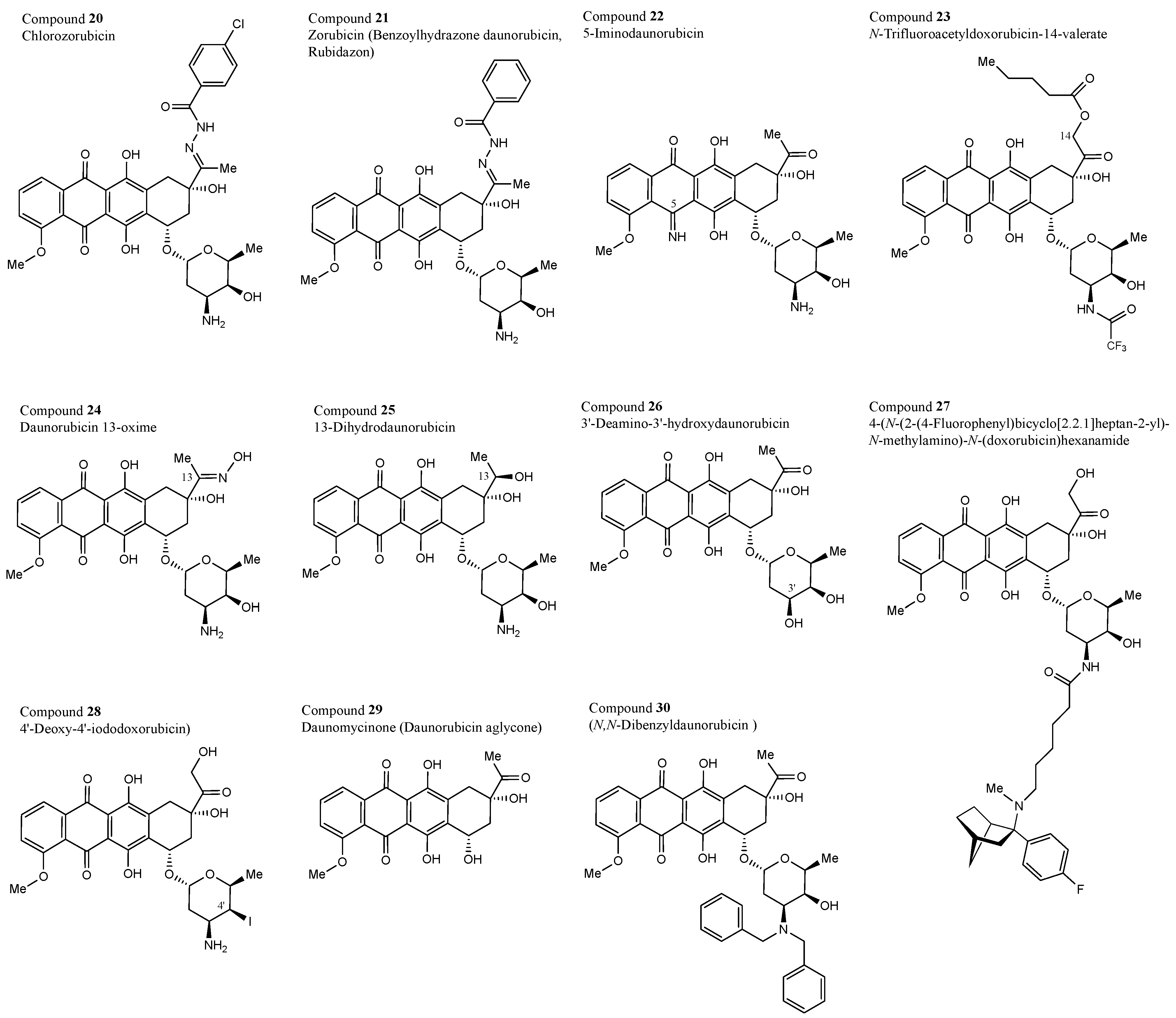
3.1. Structural Properties of the 30 Test Compounds
- •
- The presence of an amino sugar entity (even with two further yet nitrogen-free sugar units in the case of 7), but none for 12, 26, and 29 (see above)
- •
- Two phenolic OH groups in the anthraquinone part (invariably, without any exception)
- •
- In most cases two further aliphatic OH groups (three for the nine structures 1, 3, 5, 13, 14, 19, 25, 26, and 27, and even four in the case of compound 2, but in no case just one OH group or even none)
- •
- Except for compound 23, whose nitrogen is amidated (and there is another, additional amide nitrogen in 27) and compound 26, all contain basic amino functions, in most cases primary ones (-NH2); only in two cases (2, 12) is it a secondary one (-NHR); in 12, there are even four secondary amino groups; and in eight cases (1, 2, 3, 5, 7, 8, 27, and 30) they are tertiary amines (-NR2).
- •
- fluorine: in compounds 6, 23 (CF3 group), and 27
- •
- chlorine: 20
- •
- bromine: 11
- •
- iodine: 28.
3.2. Stereochemistry
3.3. Structure–Activity Relationships Regarding the Cytotoxicity of the Anthracyclines
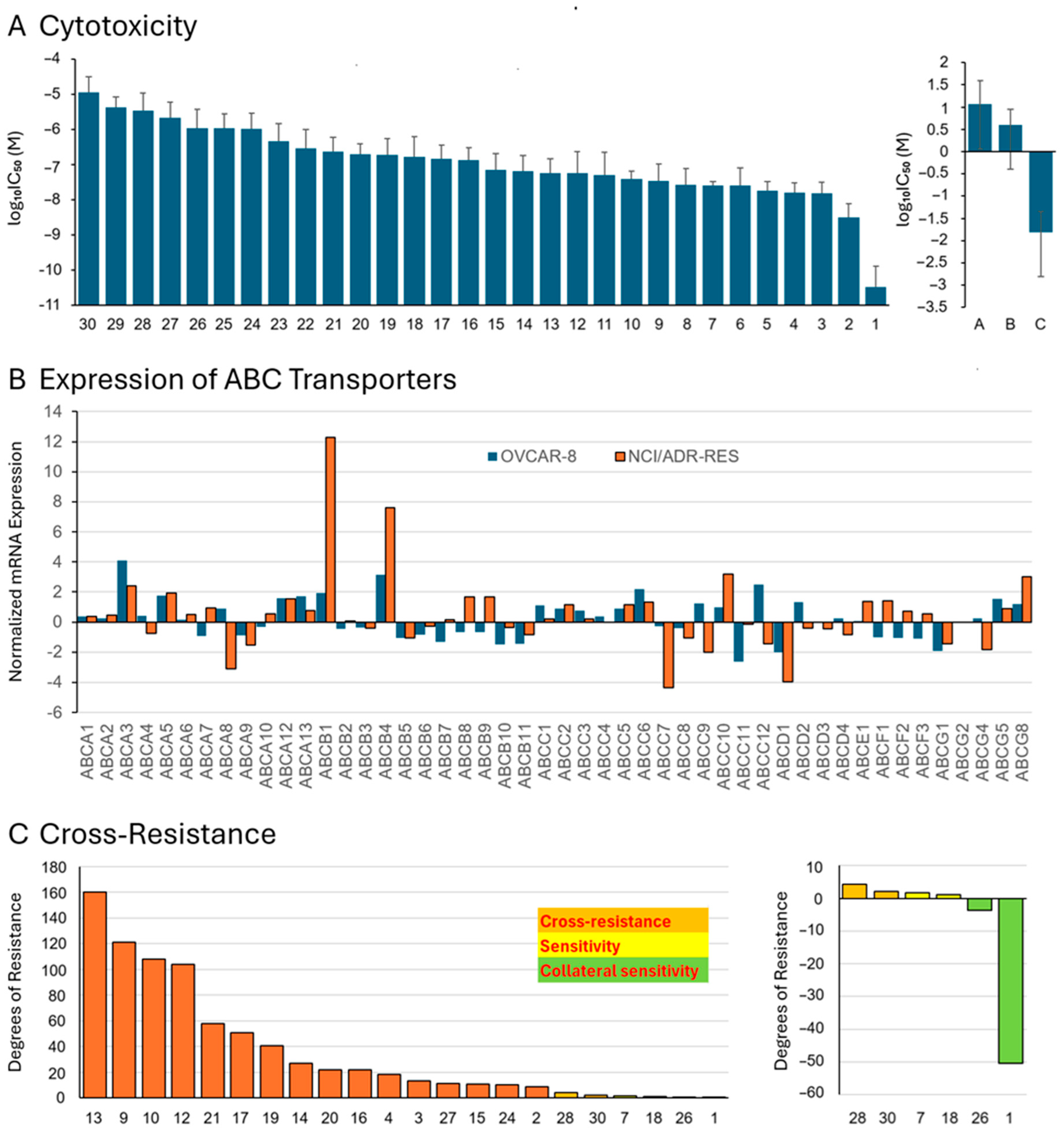
3.4. Cross-Resistance to Multidrug-Resistant Tumor Cells with Acquired Resistance
3.5. Structure–Activity Relationships Regarding the Cross-Resistance to Multidrug-Resistant Tumor Cells
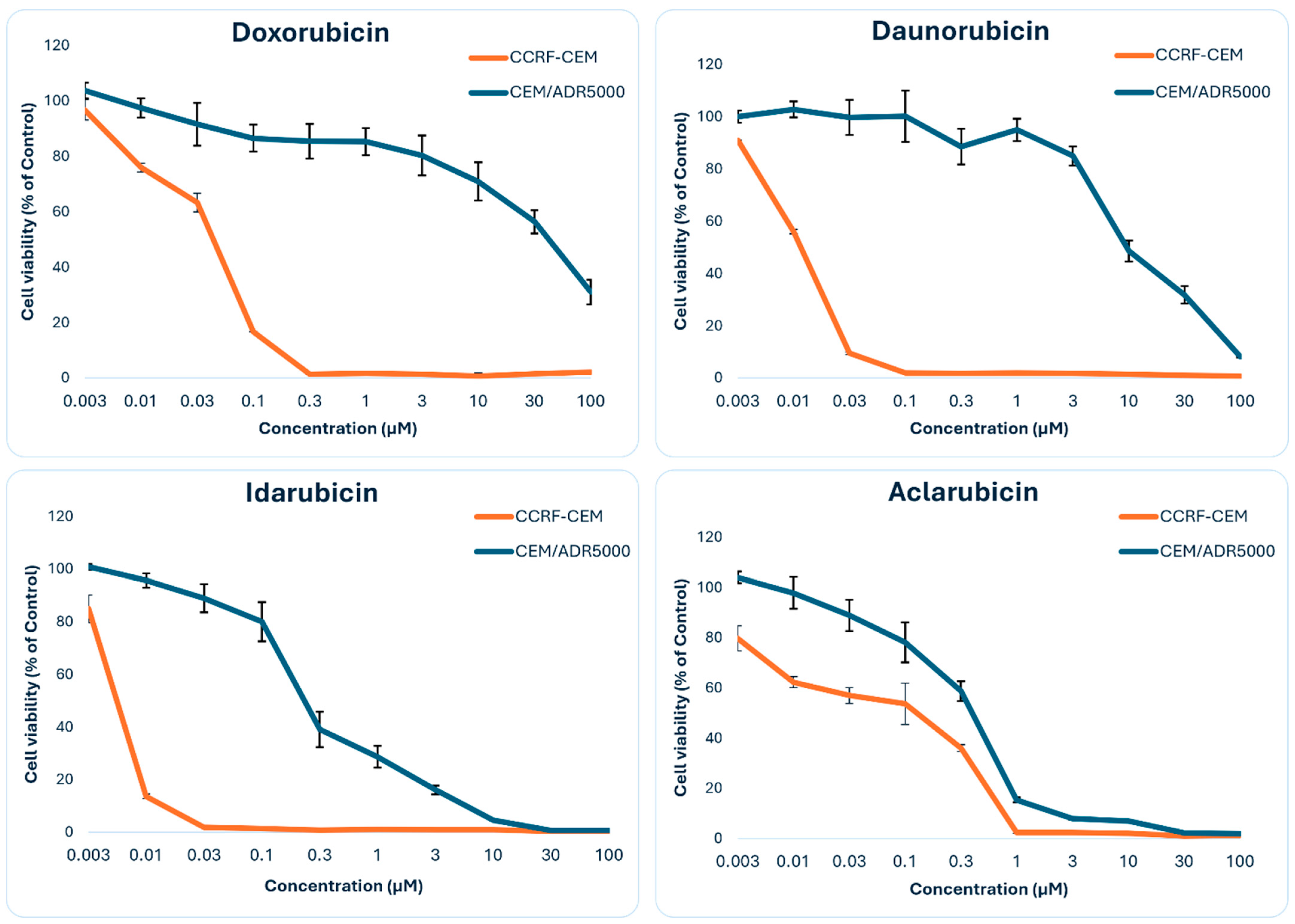
3.6. Resazurin Assay Results
3.7. Live Cell Time-Lapse Imaging
3.8. Correlation of IC50 Values and P-Glycoprotein/MDR1 Expression
3.9. Correlation of IC50 Values and All ABC Transporters
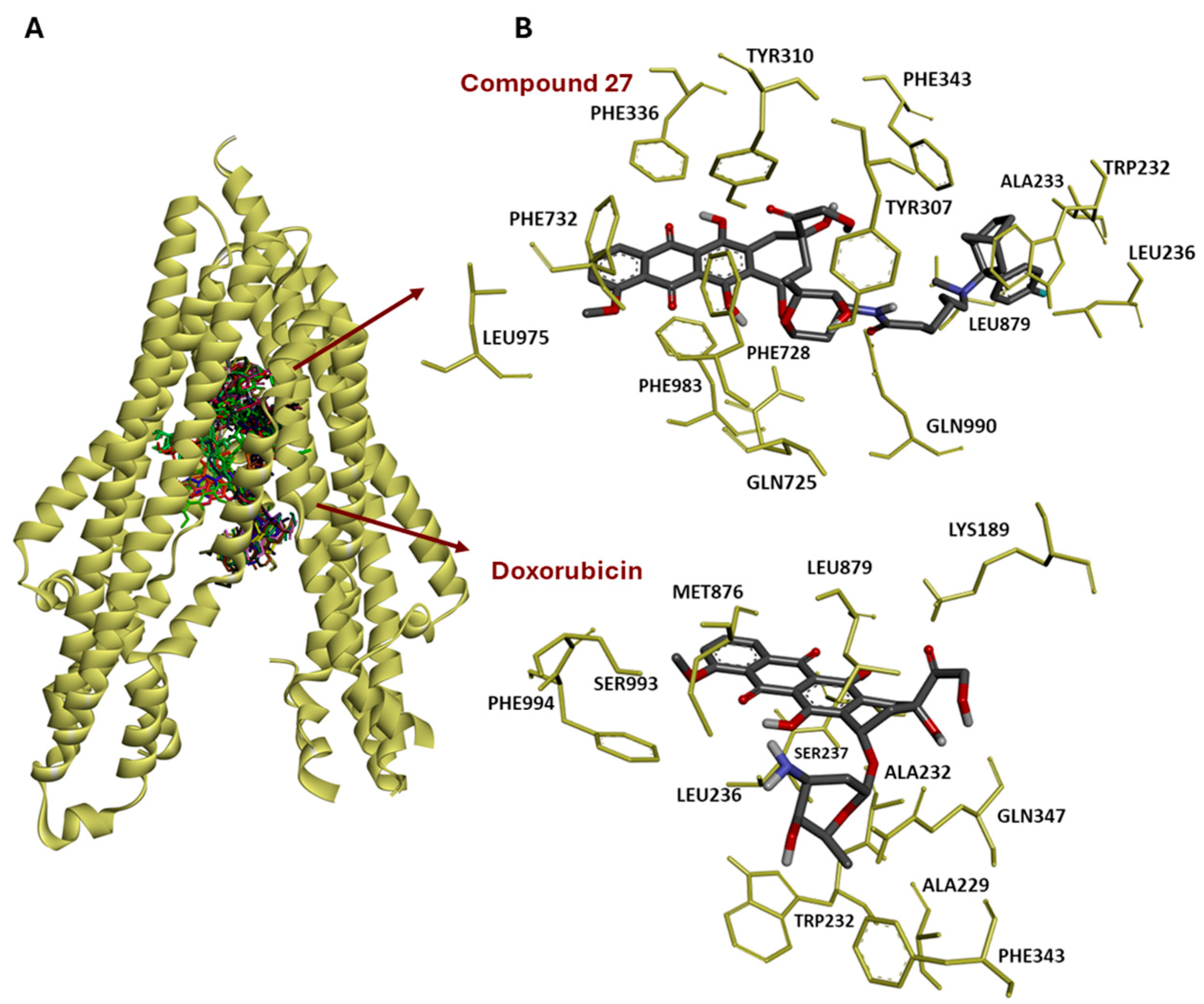
3.10. Molecular Docking of 30 Anthracyclines to P-Glycoprotein
3.11. Structure–Activity Relationships Regarding Molecular Docking of the 30 Anthracyclines to P-Glycoprotein/ABCB1
- (1)
- The 52 amino acids involved interacted in most different ways: some (less frequently occurring ones) were involved only for just one of the compounds (in the cases of Gln 195, Ile 235, Thr 240, Ala 295, Ile 306, Ser 344, Phe 372), but there was also one, Phe 983, which was interacting with nearly all compounds (21/22 compounds in the three clusters). Thus, it played a central role.
- (2)
- Likewise, most divergent were the numbers of amino acids interacting with representatives of the three different groups, extremes being some amino acids specifically did not interact with members of all three groups, but only with compounds from one or two groups, with examples being Gln 195, Ile 235, Ser 237, Phe 239, Thr 240, Ala 295, Ile 306, Leu 339, and Ile 340, Ser 344, Lys 877, Ser 880, Ser 993, which interact with none of the compounds from Group I.
- (3)
- Particularly noteworthy is the behavior of the three pairs of diastereomers of compounds 1a/b, 2a/b, and 14a/b, although they differed only by the absolute configuration at only one out of seven chiral centers, and all bound with energies that differed by only 0.3 kcal/mol between the respective diastereomers:
- •
- 1a bound to ten amino acids, of which three (Tyr 307, Ile 340, Phe 732) did not bind to 1b, while 1b bound to eleven amino acids, of which four (Tyr 310, Phe 336, Leu 339, Phe 728) did not bind to 1a. The other interactions were identical.
- •
- In a similar way, 2a bound to ten amino acids, of which two (Tyr 932, Phe 978) did not bind to 2b, while 2b bound to nine amino acids, of which one (Tyr 953) did not bind to 2a. The other interactions were identical.
- •
- In a sharp contrast, 14a/14b differed dramatically from each other: 14a bound to nine amino acids, of which none was identical to any of the eight amino acids to which its diastereomer, 14b, bound.
3.12. Kaplan–Meier Survival Analysis
4. Discussion
4.1. Structure–Activity Relationships
4.2. ABC Transporters in Acquired and Inherent Resistance
4.3. Prognostic Relevance of P-Glycoprotein/ABCB1 and Diagnostic/Therapeutic Implications
- •
- P-glycoprotein/ABCB1 could be used as a prognostic marker in these two tumor types
- •
- P-glycoprotein/ABCB1 expression may serve as a biomarker for individualized therapy with anthracyclines (and other MDR-related drugs)
- •
- P-glycoprotein/ABCB1 may serve as a target protein for inhibitors of the efflux function and the development of collaterally sensitive drugs.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| KM | Kaplan–Meier |
| miR | Micro-RNA |
| NCI | National Cancer Institute |
| TCGA | The Cancer Genome Atlas |
| qRT-PCR | Real-Time Quantitative Reverse Transcription PCR |
References
- Kessel, D. Relevance of In Vitro tests for predicting responsiveness to antitumor agents. Natl. Cancer Inst. Monogr. 1971, 34, 138–143. [Google Scholar]
- Shoemaker, R.H.; Curt, G.A.; Carney, D.N. Evidence for Multidrug-Resistant Cells in Human Tumor Cell Populations. Cancer Treat. Rep. 1983, 67, 883. [Google Scholar]
- Kartner, N.; Riordan, J.R.; Ling, V. Cell surface P-glycoprotein associated with multidrug resistance in mammalian cell lines. Science 1983, 221, 1285–1288. [Google Scholar] [CrossRef]
- Shen, D.-W.; Fojo, A.; Chin, J.; Roninson, I.; Richert, N.; Pastan, I.; Gottesman, M. Human multidrug-resistant cell lines: Increased mdr 1 expression can precede gene amplification. Science 1986, 232, 643–645. [Google Scholar] [CrossRef]
- Gottesman, M.M.; Ling, V. The molecular basis of multidrug resistance in cancer: The early years of P-glycoprotein research. FEBS Lett. 2006, 580, 998–1009. [Google Scholar] [CrossRef]
- Glisson, B.; Sullivan, D.; Gupta, R.; Ross, W. Mediation of multi-drug resistance in a Chinese hamster ovary cell line by a mutant type II topoisomerase. NCI Monogr. A Publ. Natl. Cancer Inst. 1987, 4, 89–93. [Google Scholar]
- Danks, M.K.; Schmidt, C.A.; Cirtain, M.C.; Suttle, D.P.; Beck, W.T. Altered catalytic activity of and DNA cleavage by DNA topoisomerase II from human leukemic cells selected for resistance to VM-26. Biochemistry 1988, 27, 8861–8869. [Google Scholar] [CrossRef]
- Mirski, S.E.; Gerlach, J.H.; Cole, S.P. Multidrug resistance in a human small cell lung cancer cell line selected in adriamycin. Cancer Res. 1987, 47, 2594–2598. [Google Scholar]
- Doyle, L.A.; Yang, W.; Abruzzo, L.V.; Krogmann, T.; Gao, Y.; Rishi, A.K.; Ross, D.D. A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc. Natl. Acad. Sci. USA 1998, 95, 15665–15670. [Google Scholar] [CrossRef]
- Efferth, T. The human ATP-binding cassette transporter genes from the bench to the bedside. Curr. Mol. Med. 2001, 1, 45–65. [Google Scholar] [CrossRef]
- Vasiliou, V.; Vasiliou, K.; Nebert, D.W. Human ATP-binding cassette (ABC) transporter family. Hum. Genom. 2009, 3, 281–290. [Google Scholar] [CrossRef]
- Dean, M.; Moitra, K.; Allikmets, R. The human ATP-binding cassette (ABC) transporter superfamily. Hum. Mutat. 2022, 43, 1162–1182. [Google Scholar] [CrossRef]
- Sheps, J.A.; Ralph, S.; Zhao, Z.; Baillie, D.L.; Ling, V. The ABC transporter gene family of Caenorhabditis elegans has implications for the evolutionary dynamics of multidrug resistance in eukaryotes. Genome Biol. 2004, 5, R15. [Google Scholar] [CrossRef]
- Ferreira, M.; Costa, J.; Reis-Henriques, M.A. ABC transporters in fish species: A review. Front. Physiol. 2014, 5, 266. [Google Scholar] [CrossRef]
- Theodoulou, F.L.; Kerr, I.D. ABC transporter research: Going strong 40 years on. Biochem. Soc. Trans. 2015, 43, 1033–1040. [Google Scholar] [CrossRef]
- Lefèvre, F.; Baijot, A.; Boutry, M. Plant ABC transporters: Time for biochemistry? Biochem. Soc. Trans. 2015, 43, 931–936. [Google Scholar] [CrossRef]
- Du, D.; Wang-Kan, X.; Neuberger, A.; Van Veen, H.W.; Pos, K.M.; Piddock, L.J.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Gillet, J.-P.; Efferth, T.; Remacle, J. Chemotherapy-induced resistance by ATP-binding cassette transporter genes. Biochim. et Biophys. Acta (BBA)-Rev. Cancer 2007, 1775, 237–262. [Google Scholar] [CrossRef]
- Minotti, G.; Menna, P.; Salvatorelli, E.; Cairo, G.; Gianni, L. Anthracyclines: Molecular advances and pharmacologic developments in antitumor activity and cardiotoxicity. Pharmacol. Rev. 2004, 56, 185–229. [Google Scholar] [CrossRef]
- Gewering, T.; Waghray, D.; Parey, K.; Jung, H.; Tran, N.N.; Zapata, J.; Zhao, P.; Chen, H.; Januliene, D.; Hummer, G. Tracing the substrate translocation mechanism in P-glycoprotein. eLife 2024, 12, RP90174. [Google Scholar] [CrossRef]
- Scudder, C.A.; Brown, J.M.; Sikic, B.I. DNA cross-linking and cytotoxicity of the alkylating cyanomorpholino derivative of doxorubicin in multidrug-resistant cells. J Natl Cancer Inst. 1988, 20, 1294–1298. [Google Scholar] [CrossRef]
- Shoemaker, R.H. The NCI60 human tumour cell line anticancer drug screen. Nat. Rev. Cancer 2006, 6, 813–823. [Google Scholar] [CrossRef]
- Volm, M.; Efferth, T. Role of P-Glycoprotein for Resistance of Tumors to Anticancer Drugs: From Bench to Bedside. In Resistance to Targeted ABC Transporters in Cancer; Efferth, T., Ed.; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Trock, B.J.; Leonessa, F.; Clarke, R. Multidrug resistance in breast cancer: A meta-analysis of MDR1/gp170 expression and its possible functional significance. J. Natl. Cancer Inst. 1997, 89, 917–931. [Google Scholar] [CrossRef]
- Marie, J.-P.; Legrand, O. MDR1/P-GP expression as a prognostic factor in acute leukemias. Drug Resist. Leuk. Lymphoma III 1999, 457, 1–9. [Google Scholar]
- Weinstein, J.N.; Collisson, E.A.; Mills, G.B.; Shaw, K.R.; Ozenberger, B.A.; Ellrott, K.; Shmulevich, I.; Sander, C.; Stuart, J.M. The cancer genome atlas pan-cancer analysis project. Nat. Genet. 2013, 45, 1113–1120. [Google Scholar] [CrossRef]
- Ross, D.T.; Scherf, U.; Eisen, M.B.; Perou, C.M.; Rees, C.; Spellman, P.; Iyer, V.; Jeffrey, S.S.; Van de Rijn, M.; Waltham, M.; et al. Systematic variation in gene expression patterns in human cancer cell lines. Nat. Genet. 2000, 24, 227–235. [Google Scholar] [CrossRef]
- Monga, M.; Sausville, E.A. Developmental Therapeutics Program at the NCI: Molecular target and drug discovery process. Leukemia 2002, 16, 520–526. [Google Scholar] [CrossRef]
- Reinhold, W.C.; Sunshine, M.; Liu, H.; Varma, S.; Kohn, K.W.; Morris, J.; Doroshow, J.; Pommier, Y. CellMiner: A web-based suite of genomic and pharmacologic tools to explore transcript and drug patterns in the NCI-60 cell line set. Cancer Res. 2012, 72, 3499–3511. [Google Scholar] [CrossRef]
- Liscovitch, M.; Ravid, D. A case study in misidentification of cancer cell lines: MCF-7/AdrR cells (re-designated NCI/ADR-RES) are derived from OVCAR-8 human ovarian carcinoma cells. Cancer Lett. 2007, 245, 350–352. [Google Scholar] [CrossRef]
- Brimacombe, K.R.; Hall, M.D.; Auld, D.S.; Inglese, J.; Austin, C.P.; Gottesman, M.M.; Fung, K.-L. A dual-fluorescence high-throughput cell line system for probing multidrug resistance. Assay Drug Dev. Technol. 2009, 7, 233–249. [Google Scholar] [CrossRef]
- Vert, A.; Castro, J.; Ribó, M.; Vilanova, M.; Benito, A. Transcriptional profiling of NCI/ADR-RES cells unveils a complex network of signaling pathways and molecular mechanisms of drug resistance. OncoTargets Ther. 2018, 11, 221–237. [Google Scholar] [CrossRef]
- Rubinstein, L.; Shoemaker, R.; Paull, K.; Simon, R.; Tosini, S.; Skehan, P.; Scudiero, D.; Monks, A.; Boyd, M. Comparison of In Vitro anticancer-drug-screening data generated with a tetrazolium assay versus a protein assay against a diverse panel of human tumor cell lines. JNCI J. Natl. Cancer Inst. 1990, 82, 1113–1117. [Google Scholar] [CrossRef]
- Roschke, A.V.; Lababidi, S.; Tonon, G.; Gehlhaus, K.S.; Bussey, K.; Weinstein, J.N.; Kirsch, I.R. Karyotypic “state” as a potential determinant for anticancer drug discovery. Proc. Natl. Acad. Sci. USA 2005, 102, 2964–2969. [Google Scholar] [CrossRef]
- Szakács, G.; Annereau, J.-P.; Lababidi, S.; Shankavaram, U.; Arciello, A.; Bussey, K.J.; Reinhold, W.; Guo, Y.; Kruh, G.D.; Reimers, M. Predicting drug sensitivity and resistance: Profiling ABC transporter genes in cancer cells. Cancer Cell 2004, 6, 129–137. [Google Scholar] [CrossRef]
- Hashemi, A.; Gorji-Bahri, G. MicroRNA: Promising roles in cancer therapy. Curr. Pharm. Biotechnol. 2020, 21, 1186–1203. [Google Scholar] [CrossRef]
- Katayama, K.; Noguchi, K.; Sugimoto, Y. Regulations of P-glycoprotein/ABCB1/MDR1 in human cancer cells. New J. Sci. 2014, 2014, 476974. [Google Scholar] [CrossRef]
- Liao, R.; Lin, Y.; Zhu, L. Molecular pathways involved in microRNA-mediated regulation of multidrug resistance. Mol. Biol. Rep. 2018, 45, 2913–2923. [Google Scholar] [CrossRef]
- Wang, Y.; Tu, M.-J.; Yu, A.-M. Efflux ABC transporters in drug disposition and their posttranscriptional gene regulation by microRNAs. Front. Pharmacol. 2024, 15, 1423416. [Google Scholar] [CrossRef]
- Scherf, U.; Ross, D.T.; Waltham, M.; Smith, L.H.; Lee, J.K.; Tanabe, L.; Kohn, K.W.; Reinhold, W.C.; Myers, T.G.; Andrews, D.T. A gene expression database for the molecular pharmacology of cancer. Nat. Genet. 2000, 24, 236–244. [Google Scholar] [CrossRef]
- Seo, E.-J.; Khelifi, D.; Fayez, S.; Feineis, D.; Bringmann, G.; Efferth, T.; Dawood, M. Molecular determinants of the response of cancer cells towards geldanamycin and its derivatives. Chem. Biol. Interact. 2023, 383, 110677. [Google Scholar] [CrossRef]
- Adham, A.N.; Abdelfatah, S.; Naqishbandi, A.M.; Mahmoud, N.; Efferth, T. Cytotoxicity of apigenin toward multiple myeloma cell lines and suppression of iNOS and COX-2 expression in STAT1-transfected HEK293 cells. Phytomedicine 2021, 80, 153371. [Google Scholar] [CrossRef]
- Nagy, Á.; Munkácsy, G.; Győrffy, B. Pancancer survival analysis of cancer hallmark genes. Sci. Rep. 2021, 11, 6047. [Google Scholar] [CrossRef]
- Özenver, N.; Efferth, T. Identification of Prognostic and Predictive Biomarkers and Druggable Targets among 205 Antioxidant Genes in 21 Different Tumor Types via Data-Mining. Pharmaceutics 2023, 15, 427. [Google Scholar] [CrossRef]
- Laigle, A.; Fiallo, M.M.L.; Garnier-Suillerot, A. Spectral shape modifications of anthracyclines bound to cell nuclei: A microspectrofluorometric study. Chem. Biol. Interact. 1996, 101, 49–58. [Google Scholar] [CrossRef]
- Hortobágyi, G.N. Anthracyclines in the treatment of cancer. An overview. Drugs 1997, 54 (Suppl. S4), 1–7. [Google Scholar] [CrossRef]
- Leslie, E.M.; Deeley, R.G.; Cole, S.P. Multidrug resistance proteins: Role of P-glycoprotein, MRP1, MRP2, and BCRP (ABCG2) in tissue defense. Toxicol. Appl. Pharmacol. 2005, 204, 216–237. [Google Scholar] [CrossRef]
- Bech-Hansen, N.T.; Till, J.E.; Ling, V. Pleiotropic phenotype of colchicine-resistant CHO cells: Cross-resistance and collateral sensitivity. J. Cell. Physiol. 1976, 88, 23–31. [Google Scholar] [CrossRef]
- Pluchino, K.M.; Hall, M.D.; Goldsborough, A.S.; Callaghan, R.; Gottesman, M.M. Collateral sensitivity as a strategy against cancer multidrug resistance. Drug Resist. Updat. 2012, 15, 98–105. [Google Scholar] [CrossRef]
- Efferth, T.; Saeed, M.E.M.; Kadioglu, O.; Seo, E.J.; Shirooie, S.; Mbaveng, A.T.; Nabavi, S.M.; Kuete, V. Collateral sensitivity of natural products in drug-resistant cancer cells. Biotechnol. Adv. 2020, 38, 107342. [Google Scholar] [CrossRef]
- Hall, M.D.; Marshall, T.S.; Kwit, A.D.T.; Miller Jenkins, L.M.; Dulcey, A.E.; Madigan, J.P.; Pluchino, K.M.; Goldsborough, A.S.; Brimacombe, K.R.; Griffiths, G.L.; et al. Correction: Inhibition of glutathione peroxidase mediates the collateral sensitivity of multidrug-resistant cells to tiopronin. J. Biol. Chem. 2020, 295, 4370. [Google Scholar] [CrossRef]
- Hamdoun, S.; Fleischer, E.; Klinger, A.; Efferth, T. Lawsone derivatives target the Wnt/β-catenin signaling pathway in multidrug-resistant acute lymphoblastic leukemia cells. Biochem. Pharmacol. 2017, 146, 63–73. [Google Scholar] [CrossRef]
- Gao, X.; Aguanno, D.; Board, M.; Callaghan, R. Exploiting the metabolic energy demands of drug efflux pumps provides a strategy to overcome multidrug resistance in cancer. Biochim. et Biophys. Acta (BBA)-Gen. Subj. 2021, 1865, 129915. [Google Scholar] [CrossRef]
- Stojković, S.; Podolski-Renić, A.; Dinić, J.; Stanković, T.; Banković, J.; Hadžić, S.; Paunović, V.; Isaković, A.; Tanić, N.; Pešić, M. Development of resistance to antiglioma agents in rat C6 cells caused collateral sensitivity to doxorubicin. Exp. Cell Res. 2015, 335, 248–257. [Google Scholar] [CrossRef]
- Frye, W.J.E.; Huff, L.M.; González Dalmasy, J.M.; Salazar, P.; Carter, R.M.; Gensler, R.T.; Esposito, D.; Robey, R.W.; Ambudkar, S.V.; Gottesman, M.M. The multidrug resistance transporter P-glycoprotein confers resistance to ferroptosis inducers. Cancer Drug Resist. 2023, 6, 468–480. [Google Scholar] [CrossRef]
- Dash, R.P.; Jayachandra Babu, R.; Srinivas, N.R. Therapeutic Potential and Utility of Elacridar with Respect to P-glycoprotein Inhibition: An Insight from the Published In Vitro, Preclinical and Clinical Studies. Eur. J. Drug Metab. Pharmacokinet. 2017, 42, 915–933. [Google Scholar] [CrossRef]
- Kciuk, M.; Gielecińska, A.; Mujwar, S.; Kołat, D.; Kałuzińska-Kołat, Ż.; Celik, I.; Kontek, R. Doxorubicin-An Agent with Multiple Mechanisms of Anticancer Activity. Cells 2023, 12, 659. [Google Scholar] [CrossRef]
- Theile, D.; Wizgall, P. Acquired ABC-transporter overexpression in cancer cells: Transcriptional induction or Darwinian selection? Naunyn Schmiedeberg’s Arch. Pharmacol. 2021, 394, 1621–1632. [Google Scholar] [CrossRef]
- Hientz, K.; Mohr, A.; Bhakta-Guha, D.; Efferth, T. The role of p53 in cancer drug resistance and targeted chemotherapy. Oncotarget 2017, 8, 8921–8946. [Google Scholar] [CrossRef]
- Yan, G.E.; Efferth, T. Broad-spectrum Cross-resistance to Anticancer Drugs Mediated by Epidermal Growth Factor Receptor. Anticancer Res. 2019, 39, 3585–3593. [Google Scholar] [CrossRef]
- Böhmdorfer, M.; Maier-Salamon, A.; Riha, J.; Brenner, S.; Höferl, M.; Jäger, W. Interplay of drug metabolizing enzymes with cellular transporters. Wien. Med. Wochenschr. 2014, 164, 461–471. [Google Scholar] [CrossRef]
- Iversen, D.B.; Andersen, N.E.; Dalgård Dunvald, A.C.; Pottegård, A.; Stage, T.B. Drug metabolism and drug transport of the 100 most prescribed oral drugs. Basic Clin. Pharmacol. Toxicol. 2022, 131, 311–324. [Google Scholar] [CrossRef]
- Roos, W.P.; Thomas, A.D.; Kaina, B. DNA damage and the balance between survival and death in cancer biology. Nat. Rev. Cancer 2016, 16, 20–33. [Google Scholar] [CrossRef]
- Bukowski, K.; Kciuk, M.; Kontek, R. Mechanisms of Multidrug Resistance in Cancer Chemotherapy. Int. J. Mol. Sci. 2020, 21, 3233. [Google Scholar] [CrossRef]
- Yan, G.; Elbadawi, M.; Efferth, T. Multiple cell death modalities and their key features. World Acad. Sci. J. 2020, 2, 39–48. [Google Scholar] [CrossRef]
- Gordon, R.R.; Nelson, P.S. Cellular senescence and cancer chemotherapy resistance. Drug Resist. Updat. 2012, 15, 123–131. [Google Scholar] [CrossRef]
- De Angelis, M.L.; Francescangeli, F.; La Torre, F.; Zeuner, A. Stem Cell Plasticity and Dormancy in the Development of Cancer Therapy Resistance. Front. Oncol. 2019, 9, 626. [Google Scholar] [CrossRef]
- Wigner, P.; Zielinski, K.; Labieniec-Watala, M.; Marczak, A.; Szwed, M. Doxorubicin-transferrin conjugate alters mitochondrial homeostasis and energy metabolism in human breast cancer cells. Sci. Rep. 2021, 11, 4544. [Google Scholar] [CrossRef]
- Testa, U.; Pelosi, E.; Peschle, C. The transferrin receptor. Crit. Rev. Oncog. 1993, 4, 241–276. [Google Scholar]
- Kelter, G.; Steinbach, D.; Konkimalla, V.B.; Tahara, T.; Taketani, S.; Fiebig, H.H.; Efferth, T. Role of transferrin receptor and the ABC transporters ABCB6 and ABCB7 for resistance and differentiation of tumor cells towards artesunate. PLoS ONE 2007, 2, e798. [Google Scholar] [CrossRef]
- Xu, X.; Persson, H.L.; Richardson, D.R. Molecular pharmacology of the interaction of anthracyclines with iron. Mol. Pharmacol. 2005, 68, 261–271. [Google Scholar] [CrossRef]
- Yang, R.; Li, Y.; Wang, X.; Yan, J.; Pan, D.; Xu, Y.; Wang, L.; Yang, M. Doxorubicin loaded ferritin nanoparticles for ferroptosis enhanced targeted killing of cancer cells. RSC Adv. 2019, 9, 28548–28553. [Google Scholar] [CrossRef]
- Tan, R.; Tian, D.; Liu, J.; Wang, C.; Wan, Y. Doxorubicin-Bound Hydroxyethyl Starch Conjugate Nanoparticles with pH/Redox Responsive Linkage for Enhancing Antitumor Therapy. Int. J. Nanomed. 2021, 16, 4527–4544. [Google Scholar] [CrossRef]
- Mohammad, S.N.; Choi, Y.S.; Chung, J.Y.; Cedrone, E.; Neun, B.W.; Dobrovolskaia, M.A.; Yang, X.; Guo, W.; Chew, Y.C.; Kim, J.; et al. Nanocomplexes of doxorubicin and DNA fragments for efficient and safe cancer chemotherapy. J. Control. Release 2023, 354, 91–108. [Google Scholar] [CrossRef]
- Robey, R.W.; Pluchino, K.M.; Hall, M.D.; Fojo, A.T.; Bates, S.E.; Gottesman, M.M. Revisiting the role of ABC transporters in multidrug-resistant cancer. Nat. Rev. Cancer 2018, 18, 452–464. [Google Scholar] [CrossRef]
- Cornelissen, J.J.; Sonneveld, P.; Schoester, M.; Raaijmakers, H.G.; Nieuwenhuis, H.K.; Dekker, A.W.; Lokhorst, H.M. MDR-1 expression and response to vincristine, doxorubicin, and dexamethasone chemotherapy in multiple myeloma refractory to alkylating agents. J. Clin. Oncol. 1994, 12, 115–119. [Google Scholar] [CrossRef]
- Nooter, K.; Sonneveld, P. Clinical relevance of P-glycoprotein expression in haematological malignancies. Leuk. Res. 1994, 18, 233–243. [Google Scholar] [CrossRef]
- Kato, A.; Miyazaki, M.; Ambiru, S.; Yoshitomi, H.; Ito, H.; Nakagawa, K.; Shimizu, H.; Yokosuka, O.; Nakajima, N. Multidrug resistance gene (MDR-1) expression as a useful prognostic factor in patients with human hepatocellular carcinoma after surgical resection. J. Surg. Oncol. 2001, 78, 110–115. [Google Scholar] [CrossRef]
- Benderra, Z.; Faussat, A.M.; Sayada, L.; Perrot, J.Y.; Tang, R.; Chaoui, D.; Morjani, H.; Marzac, C.; Marie, J.P.; Legrand, O. MRP3, BCRP, and P-glycoprotein activities are prognostic factors in adult acute myeloid leukemia. Clin. Cancer Res. 2005, 11, 7764–7772. [Google Scholar] [CrossRef]
- van der Kolk, D.M.; de Vries, E.G.; van Putten, W.J.; Verdonck, L.F.; Ossenkoppele, G.J.; Verhoef, G.E.; Vellenga, E. P-glycoprotein and multidrug resistance protein activities in relation to treatment outcome in acute myeloid leukemia. Clin. Cancer Res. 2000, 6, 3205–3214. [Google Scholar]
- Chakraborty, K.; Ghosh, P. P-glycoprotein Expression in Solid Tumors—An Analysis. Int. J. Pharm. Investig. 2021, 11, 345–348. [Google Scholar] [CrossRef]
- Beck, W.T.; Grogan, T.M.; Willman, C.L.; Cordon-Cardo, C.; Parham, D.M.; Kuttesch, J.F.; Andreeff, M.; Bates, S.E.; Berard, C.W.; Boyett, J.M.; et al. Methods to detect P-glycoprotein-associated multidrug resistance in patients’ tumors: Consensus recommendations. Cancer Res. 1996, 56, 3010–3020. [Google Scholar]
- Efferth, T. Testing for tumor drug resistance in the age of molecular medicine. A contribution to the Debate Round-Table on Phenotypic and Genotypic Analyses of Multidrug Resistance (MDR) in Clinical Hospital Practice. Leukemia 1999, 13, 1627–1629. [Google Scholar] [CrossRef]
- Ford, J.M.; Yang, J.M.; Hait, W.N. P-glycoprotein-mediated multidrug resistance: Experimental and clinical strategies for its reversal. Cancer Treat. Res. 1996, 87, 3–38. [Google Scholar]
- Leopoldo, M.; Nardulli, P.; Contino, M.; Leonetti, F.; Luurtsema, G.; Colabufo, N.A. An updated patent review on P-glycoprotein inhibitors (2011–2018). Expert Opin. Ther. Pat. 2019, 29, 455–461. [Google Scholar] [CrossRef]
- Engle, K.; Kumar, G. Cancer multidrug-resistance reversal by ABCB1 inhibition: A recent update. Eur. J. Med. Chem. 2022, 239, 114542. [Google Scholar] [CrossRef]
- Schäfer, J.; Klösgen, V.J.; Omer, E.A.; Kadioglu, O.; Mbaveng, A.T.; Kuete, V.; Hildebrandt, A.; Efferth, T. In Silico and In Vitro Identification of P-Glycoprotein Inhibitors from a Library of 375 Phytochemicals. Int. J. Mol. Sci. 2023, 24, 10240. [Google Scholar] [CrossRef]
- Tamaki, A.; Ierano, C.; Szakacs, G.; Robey, R.W.; Bates, S.E. The controversial role of ABC transporters in clinical oncology. Essays Biochem. 2011, 50, 209–232. [Google Scholar]
- Amiri-Kordestani, L.; Fojo, T. Why do phase III clinical trials in oncology fail so often? J. Natl. Cancer Inst. 2012, 104, 568–569. [Google Scholar] [CrossRef]
- Rugo, H.S.; Umanzor, G.A.; Barrios, F.J.; Vasallo, R.H.; Chivalan, M.A.; Bejarano, S.; Ramírez, J.R.; Fein, L.; Kowalyszyn, R.D.; Kramer, E.D.; et al. Open-Label, Randomized, Multicenter, Phase III Study Comparing Oral Paclitaxel Plus Encequidar Versus Intravenous Paclitaxel in Patients with Metastatic Breast Cancer. J. Clin. Oncol. 2023, 41, 65–74. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound No | NSC Code | Trivial Name | Chemical Name |
|---|---|---|---|
| 1a (3″S) | NSC 357704 | 3′-Deamino-3′-(4″-(3″-cyano)morpholinyl)-doxorubicin | |
| 1b (3″R) | NSC 357704 | 3′-Deamino-3′-(4″-(3″-cyano)morpholinyl)-doxorubicin | |
| 2a (13R) | NSC 639655 | 3’-Deamino-3′-(4″-morpholinyl)-13-dihydro-doxorubicin | |
| 2b (13S) | NSC 639655 | 3′-Deamino-3′-(4″-morpholinyl)-13-dihydro-doxorubicin | |
| 3 | NSC 354646 | 3′-Deamino-3′-(4″-morpholinyl)-doxorubicin | |
| 4 | NSC 256439 | Idarubicin | 4-Demethoxydaunorubicin |
| 5 | NSC 261045 | N,N-Dimethyldoxorubicin | |
| 6 | NSC 623128 | 14-Fluoro-4-demethoxydaunorubicin | |
| 7 | NSC 208734 | Aclarubicin | (1S,2S,4R)-Methyl-4-(((2S,5R,6R)-4-(dimethylamino)-5-(((1S,3R,4S)-3-hydroxy-5-methyl-4-(((2S,6R)-6-methyl-5-oxotetrahydro-2H-pyran-2-yl)oxy)cyclohexyl)oxy)-6-methyltetrahydro-2H-pyran-2-yl)oxy)-2-ethyl-2,5,7-trihydroxy-6,11-dioxo-1,2,3,4,6,11-hexahydrotetracene-1-carboxylate |
| 8 | NSC 258812 | N,N-Dimethyldaunorubicin | |
| 9 | NSC 83142 | Daunorubicin | (7S,9S)-9-acetyl-7-[(2R,4S,5S,6S)-4-amino-5-hydroxy-6-methyloxan-2-yl]oxy-6,9,11-trihydroxy-4-methoxy-8,10-dihydro-7H-tetracene-5,12-dione |
| 10 | NSC 267469 | 4′-Deoxydoxorubicin | |
| 11 | NSC 650931 | 2′-Bromo-4′-epi-daunorubicin | |
| 12 | NSC 301739 | Mitoxantrone | 1,4-dihydroxy-5,8-bis [2-(2-hydroxyethylamino)ethylamino]anthracene-9,10-dione |
| 13 | NSC 759155 | Doxorubicin | 14-Hydroxydaunorubicine |
| 14a (1″R) | NSC 639659 | N-(1″-Cyano-2″-methoxyethyl)doxorubicin | |
| 14b (1″S) | NSC 639659 | N-(1″-Cyano-2″-methoxyethyl)doxorubicin | |
| 15 | NSC 149584 | Doxorubicin 14-octanoate | |
| 16 | NSC 149583 | Doxorubicin 14-nicotinate | |
| 17 | NSC 149585 | Doxorubicin 14-benzoate | |
| 18 | NSC 333054 | Pirarubicin | 4′-O-Tetrahydropyranyldoxorubicin |
| 19 | NSC 759195 | Epirubicin | 4′-epi-Doxorubicin |
| 20 | NSC 219977 | Chlorozorubicin | Chlorobenzoylhydrazone daunorubicin |
| 21 | NSC 164011 | Zorubicin/Rubidazone | Benzoylhydrazone daunorubicin |
| 22 | NSC 254681 | 5-Iminodaunorubicin | |
| 23 | NSC 246131 | N-Trifluoroacetyldoxorubicin-14-valerate | |
| 24 | NSC 143491 | Daunorubicin 13-oxime | |
| 25 | NSC 180510 | 13-Dihydrodaunorubicin | |
| 26 | NSC 284682 | 3′-Deamino-3′-hydroxydaunorubicin | |
| 27 | NSC 788321 | 4-(N-(2-(4-Fluorophenyl)bicyclo [2.2.1]heptan-2-yl)-N-methylamino)-N-(doxorubicin)hexanamide | |
| 28 | NSC 378901 | 4′-Deoxy-4′-iododoxorubicin | |
| 29 | NSC 109351 | Daunomycinone | Daunorubicin aglycone |
| 30 | NSC 268242 | N,N-Dibenzyldaunorubicin |
| CCRF-CEM | CEM/ADR5000 | Degree of Resistance | |
|---|---|---|---|
| Doxorubicin (13) | 0.042 ± 0.003 | 41.51 ± 8.17 | 988.3 |
| Daunorubicin (9) | 0.012 ± 0.000 | 9.89 ± 1.37 | 824.2 |
| Idarubicin (4) | 0.005 ± 0.000 | 0.23 ± 0.04 | 46.0 |
| Aclarubicin (7) | 0.145 ± 0.015 | 0.38 ± 0.03 | 2.6 |
| Compound No | Binding Energy (kcal/mol) | Inhibition Constant (pKi, nM) | Hydrogen Bonds and Polar Interactions | Hydrophobic and Aromatic Interactions |
|---|---|---|---|---|
| 1a (3″S) | −11.9 ± 0.1 | 2.03 ± 0.20 | Tyr 307, Ser 979, Tyr 953 | Leu 65, Met 68, Ile 340, Phe 732, Met 949, Tyr 950, Phe 983, Met 986 |
| 1b (3″R) | −11.6 ± 0.1 | 3.35 ± 0.76 | Tyr 310, Tyr 953, Ser 979 | Leu 65, Met 68, Phe 728, Met 949, Tyr 950, Phe 983 |
| 2a (13R) | −11.3 ± 0.2 | 5.74 ± 1.38 | Gln 725, Phe 983, Gln 990 | Met 69, Phe 72, Phe 336, Leu 339, Ile 340, Phe 732, Tyr 953, Phe 978 |
| 2b (13S) | −11.0 ± 0.1 | 8.99 ± 1.74 | Gln 725, Phe 983, Gly 990 | Met 69, Phe 72, Phe 336, Leu 339, Ile 340, Tyr 953 |
| 3 | −12.4 ± 0.0 | 0.84 ± 0.05 | Tyr310, Tyr 953, Leu 975 | Leu 65, Met 68, Phe 72, Ile 340, Phe 728, Phe 732, Met 949, Tyr 950, Phe 978, Phe 983 |
| 4 | −10.4 ± 0.1 | 24.08 ± 3.16 | Lys 189, Gln 347 | Ala 229, Trp 232, Ala 233, Leu 236, Phe 343, Pro 350, Met 876 |
| 5 | −11.7 ± 0.0 | 2.90 ± 0.07 | Tyr 310, Tyr 953 | Leu 65, Met 68, Ile 340, Phe 728, Met 949, Tyr 950, Phe 983 |
| 6 | −10.4 ± 0.2 | 25.11 ± 6.45 | Tyr 310, Gln 990 | Tyr 307, Phe 336, Phe 732, Phe 978, Phe 983, Ala 987 |
| 7 | −15.1 ± 0.2 | (9.41 ± 3.34) × 10−3 | Gln 195, Ile 340, Ser 344, Gln 725, Tyr 953, Ser 979 | Leu 65, Met 69, Phe 336, Phe 983 |
| 8 | −11.0 ± 0.1 | 8.74 ± 1.87 | Ala 229, Trp 232, Gln 990 | Leu 236, Ile 299, Phe 303, Phe 770, Met 876, Leu 879, Phe 994 |
| 9 | −10.9 ± 0.1 | 9.96 ± 1.32 | Tyr 310, Leu 339, Ile 340, Gln 347, Gln 725, Glu 875, Phe 983 | Met 986, Ala 987 |
| 10 | −11.4 ± 0.4 | 5.55 ± 3.08 | Tyr 310, Tyr 953 | Leu 65, Met 68, Phe 72, Phe 336, Phe 732, Met 949, Tyr 950, Phe 978, Phe 983 |
| 11 | −11.0 ± 0.0 | 8.98 ± 0.28 | Tyr 310, Gln 725, Tyr 953, Ser 979 | Leu 65, Met 68, Ile 340, Phe 728, Met 949, Tyr 950 |
| 12 | −10.3 ± 0.2 | 29.1 ± 9.20 | Phe 983, Tyr 950, Tyr 953 | Phe 336 |
| 13 | −12.2 ± 0.0 | 1.16 ± 0.08 | Lys 189, Ser 237, Gln 347, Met 876, Ser 993 | Ala 229, Trp 232, Ala 233, Leu 236, Phe 343, Phe 994 |
| 14a (1″R) | −11.9 ± 0.1 | 1.88 ± 0.36 | Tyr 310, Gln 725, Leu 975 | Tyr 307, Phe 336, Phe 728, Phe 732, Phe 978, Phe 983 |
| 14b (1″S) | −11.6 ± 0.1 | 3.07 ± 0.53 | Trp 232, Gln 838 | Ile 235, Leu 236, Phe 239, Ala 295, Ile 299, Met 876 |
| 15 | −15.4 ± 0.4 | (6.23 ± 5.00) × 10−3 | Asn 721, Gln 725, Gln 838, Gln 990 | Trp 232, Phe 303, Phe 336, Phe 728, Phe 732, Met 876, Phe 983, Val 991, Phe 994 |
| 16 | −14.7 ± 0.1 | (17.37 ± 3.00) × 10−3 | Tyr 307, Asn 721, Gln 725, Gln 838, Ser 979 | Trp 232, Phe 303, Phe 728, Phe 983, Phe 994 |
| 17 | −14.6 ± 0.1 | (20.01 ± 4.24) × 10−3 | Tyr 310, Gln 725, Glu 875, Met 986 | Leu 65, Phe 343, Phe 728, Phe 732, Met 949, Tyr 950, Tyr 953, Phe 983 |
| 18 | −13.1 ± 0.0 | 0.25 ± 0.02 | Tyr 307, Gln 725, Tyr 953 | Met 69, Phe 72, Ile 306, Phe 336, Leu 339, Ile 340, Phe 343, Phe 983 |
| 19 | −11.8 ± 0.1 | 2.38 ± 0.43 | Lys 189, Phe 239, Thr 240, Ser 880, Ser 993 | Ala 233, Leu 236, Pro 350, Leu 879, Phe 994 |
| 20 | −15.1 ± 0.1 | (9.22 ± 1.6) × 10−3 | Tyr 310, Asn 721, Gln 990 | Ile 299, Trp 232, Phe 303, Gln 725, Phe 732, Phe 770, Met 876, Leu 975, Phe 978, Phe 983, Ala 987 |
| 21 | −14.4 ± 0.0 | (26.6 ± 1.5) × 10−3 | Trp 232, Glu 875, Gln 990 | Ala 233, Phe 303, Phe 343, Pro 350, Met 876, Leu 879, Ala 987, Val 991 |
| 22 | −10.8 ± 0.0 | 11.17 ± 0.17 | Glu 875, Gln 990 | Trp 232, Phe 303, Phe 343, Met 876, Ala 987, Val 991 |
| 23 | −16.2 ± 0.6 | (1.86 ± 1.41) × 10−3 | Tyr 307, Gln 725, Tyr 953, Gln 990 | Met 69, Phe 72, Phe 336, Leu 339, Ile 340, Phe 343, Phe 983, Ala 987 |
| 24 | −11.0 ± 0.2 | 9.59 ± 2.83 | Lys 189, Ala 233, Leu 236, Met 876, Leu 879 | Trp 232, Phe 343, Ser 880, Lys 877 |
| 25 | −11.2 ± 0.1 | 6.79 ± 1.34 | Gln 347, Met 876 | Trp 232, Ala 233, Leu 236, Phe 343, Pro 350, Lys 877, Leu 879, Ser 880 |
| 26 | −11.3 ± 0.1 | 5.02 ± 0.96 | Tyr 307, Tyr 310, Ser 979 | Met 69, Phe 72, Phe 336, Leu 339, Ile 340, Phe 983 |
| 27 | −17.5 ± 0.5 | (0.18 ± 0.13) × 10−3 | Tyr 310, Gln 725, Leu879, Leu 975, Gln 990 | Trp 232, Ala 233, Leu 236, Tyr 307, Phe 336, Phe 343, Phe 372, Phe 728, Phe 983 |
| 28 | −11.5 ± 0.0 | 3.87 ± 0.19 | Trp 232, Glu 875, Gln 990 | Phe 303, Phe 343, Met 876, Ala 987, Val 991 |
| 29 | −9.5 ± 0.0 | 103.74 ± 7.08 | Lys 189, Ser 237, Gln 347 | Ala 233, Leu 236, Met 876, Leu 879, Ser 880, Phe 994 |
| 30 | −14.9 ± 0.0 | (12.15 ± 0.9) × 10−3 | Tyr 953, Ser 979 | Leu 65, Met 68, Tyr 307, Phe 732, Met 949, Tyr 950, Phe 983, Met 986, Ala 987 |
| Elacridar | −14.3 ± 0.1 | (35.49 ± 3.5) × 10−3 | Lys 189 | Trp 232, Ala 233, Leu 236, Phe 303, Pro 350, Phe 770, Met 876, Leu 879, Val 991, Phe 994 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yücer, R.; Piccinno, R.; Ooko, E.; Dawood, M.; Bringmann, G.; Efferth, T. Predictive and Prognostic Relevance of ABC Transporters for Resistance to Anthracycline Derivatives. Biomolecules 2025, 15, 971. https://doi.org/10.3390/biom15070971
Yücer R, Piccinno R, Ooko E, Dawood M, Bringmann G, Efferth T. Predictive and Prognostic Relevance of ABC Transporters for Resistance to Anthracycline Derivatives. Biomolecules. 2025; 15(7):971. https://doi.org/10.3390/biom15070971
Chicago/Turabian StyleYücer, Rümeysa, Rossana Piccinno, Ednah Ooko, Mona Dawood, Gerhard Bringmann, and Thomas Efferth. 2025. "Predictive and Prognostic Relevance of ABC Transporters for Resistance to Anthracycline Derivatives" Biomolecules 15, no. 7: 971. https://doi.org/10.3390/biom15070971
APA StyleYücer, R., Piccinno, R., Ooko, E., Dawood, M., Bringmann, G., & Efferth, T. (2025). Predictive and Prognostic Relevance of ABC Transporters for Resistance to Anthracycline Derivatives. Biomolecules, 15(7), 971. https://doi.org/10.3390/biom15070971