
Synovial Fluid Extracellular Vesicles from Patients with Severe Osteoarthritis Differentially Promote a Pro-Catabolic, Inflammatory Chondrocyte Phenotype



Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Recruitment and Sample Collection
2.2. Isolation and Culture of Primary Human OA Chondrocytes
2.3. Synovial Fluid EV Isolation and Characterisation
2.4. SFEV and Synovial Fluid Treatment of Chondrocytes
2.5. RNA Isolation and Bulk RNA Sequencing
2.6. Luminex and ELISA
2.7. Statistical Analysis
3. Results
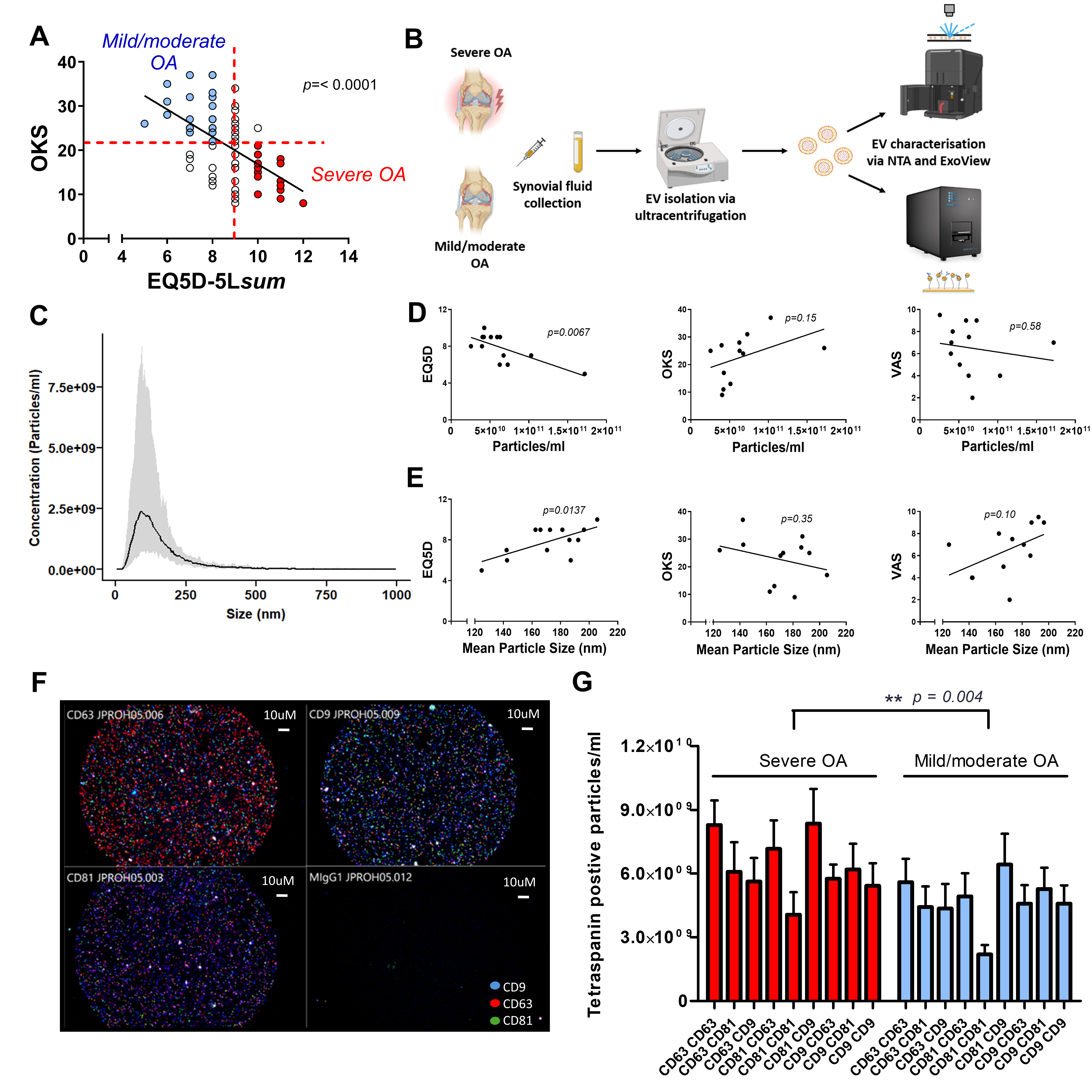
3.1. Patient Characteristics
3.2. Characterisation of Synovial Fluid Extracellular Vesicles
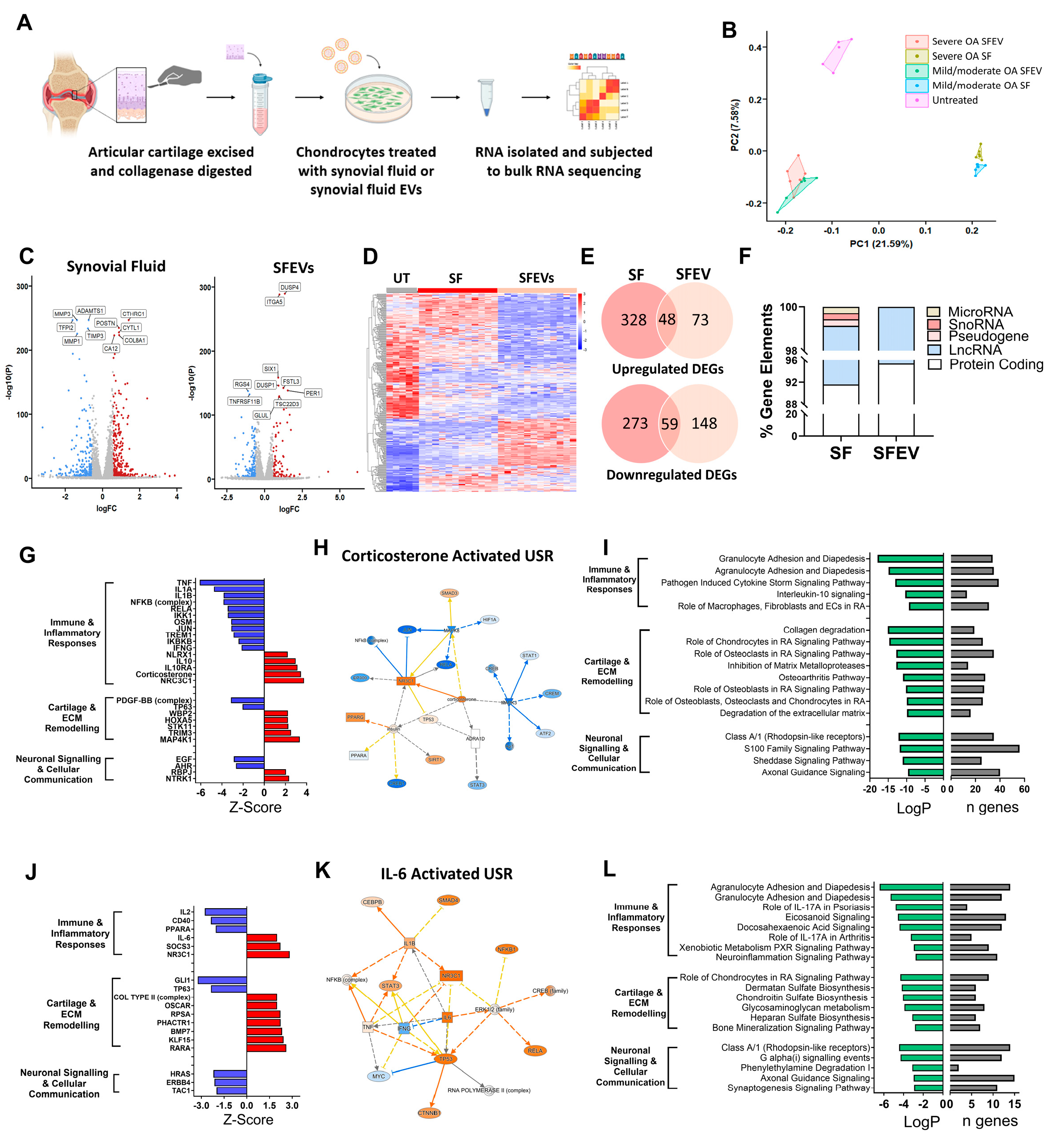
3.3. Synovial Fluid and Synovial Fluid EVs Induce Distinct Transcriptomic Profiles in Human Articular Chondrocytes
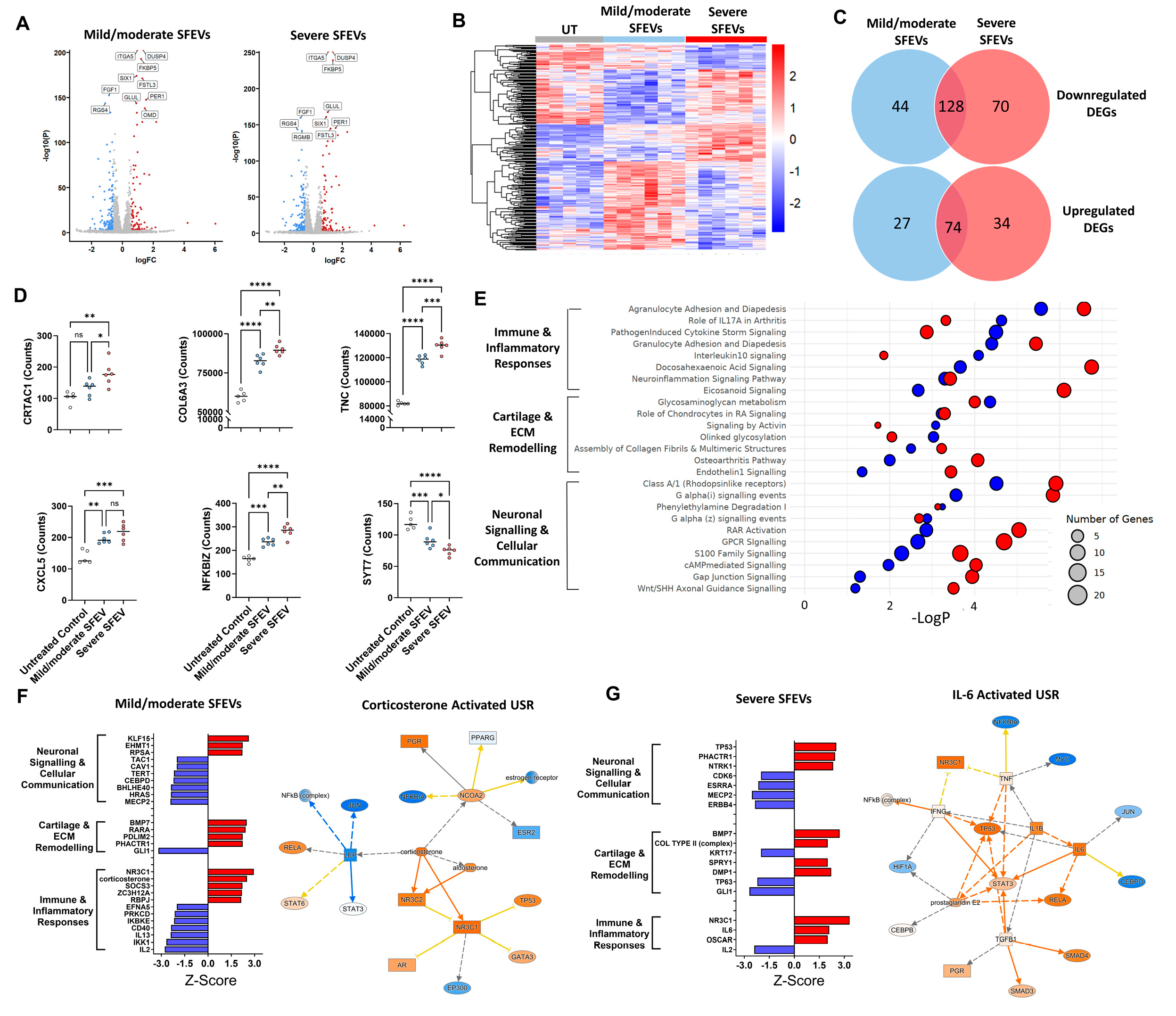
3.4. Synovial Fluid EVs from Patients with Severe OA Elicit Differential Catabolic and Inflammatory Transcriptomic Responses in Human Articular Chondrocytes
3.5. Synovial Fluid EVs from Patients with Severe OA Induce the Differential Release of Catabolic and Inflammatory Mediators in Human Articular Chondrocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| MDPI | Multidisciplinary Digital Publishing Institute |
| EVs | Extracellular vesicles |
| SFEVs | Synovial Fluid Extracellular Vesicles |
| OKS | Oxford Knee Score |
| VAS | Visual Analogue Scale |
| EQ5D | EuroQol 5-Dimensions (quality of life questionnaire) |
| NTA | Nanoparticle Tracking Analysis |
| DEGs | Differentially Expressed Genes |
| RNA-seq | RNA sequencing |
| IPA | Ingenuity Pathway Analysis |
| ECM | Extracellular Matrix |
| MMP | Matrix Metalloproteinase |
| FCS | Foetal Calf Serum |
| NEAA | Non-Essential Amino Acids |
| DMEM | Dulbecco’s Modified Eagle Medium |
| PBS | Phosphate-Buffered Saline |
| ELISA | Enzyme-Linked Immunosorbent Assay |
| FC | Fold Change |
| Log2FC | Log2 Fold Change |
| FDR | False Discovery Rate |
| PCA | Principal Component Analysis |
| USRs | Upstream Regulators |
| IQR | Interquartile Range |
| BMI | Body mass index |
| hg38 | Human Genome Reference Build 38 |
| DNBSEQ | DNA Nanoball Sequencing Platform |
| DESEQ | Differential gene expression analysis tool |
References
- Cross, M.; Smith, E.; Hoy, D.; Nolte, S.; Ackerman, I.; Fransen, M.; Bridgett, L.; Williams, S.; Guillemin, F.; Hill, C.L.; et al. The global burden of hip and knee osteoarthritis: Estimates from the global burden of disease 2010 study. Ann. Rheum. Dis. 2014, 73, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Hawker, G.A.; King, L.K. The Burden of Osteoarthritis in Older Adults. Clin. Geriatr. Med. 2022, 38, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Coaccioli, S.; Sarzi-Puttini, P.; Zis, P.; Rinonapoli, G.; Varrassi, G. Osteoarthritis: New Insight on Its Pathophysiology. J. Clin. Med. 2022, 11, 6013. [Google Scholar] [CrossRef]
- Tonge, D.P.; Pearson, M.J.; Jones, S.W. The hallmarks of osteoarthritis and the potential to develop personalised disease-modifying pharmacological therapeutics. Osteoarthr. Cartil. 2014, 22, 609–621. [Google Scholar] [CrossRef]
- Mobasheri, A.; Loeser, R. Clinical phenotypes, molecular endotypes and theratypes in OA therapeutic development. Nat. Rev. Rheumatol. 2024, 20, 525–526. [Google Scholar] [CrossRef]
- Angelini, F.; Widera, P.; Mobasheri, A.; Blair, J.; Struglics, A.; Uebelhoer, M.; Henrotin, Y.; Marijnissen, A.C.; Kloppenburg, M.; Blanco, F.J.; et al. Osteoarthritis endotype discovery via clustering of biochemical marker data. Ann. Rheum. Dis. 2022, 81, 666–675. [Google Scholar] [CrossRef]
- Nanus, D.E.; Badoume, A.; Wijesinghe, S.N.; Halsey, A.M.; Hurley, P.; Ahmed, Z.; Botchu, R.; Davis, E.T.; Lindsay, M.A.; Jones, S.W. Synovial tissue from sites of joint pain in knee osteoarthritis patients exhibits a differential phenotype with distinct fibroblast subsets. EBioMedicine 2021, 72, 103618. [Google Scholar] [CrossRef] [PubMed]
- Welsh, J.A.; Goberdhan, D.C.I.; O’Driscoll, L.; Buzas, E.I.; Blenkiron, C.; Bussolati, B.; Cai, H.; Di Vizio, D.; Driedonks, T.A.P.; Erdbrugger, U.; et al. Minimal information for studies of extracellular vesicles (MISEV2023): From basic to advanced approaches. J. Extracell. Vesicles 2024, 13, e12404. [Google Scholar] [CrossRef]
- Zeng, G.; Deng, G.; Xiao, S.; Li, F. Fibroblast-like Synoviocytes-derived Exosomal PCGEM1 Accelerates IL-1β-induced Apoptosis and Cartilage Matrix Degradation by miR-142-5p/RUNX2 in Chondrocytes. Immunol. Investig. 2022, 51, 1284–1301. [Google Scholar] [CrossRef]
- Cao, Y.; Ruan, J.; Kang, J.; Nie, X.; Lan, W.; Ruan, G.; Li, J.; Zhu, Z.; Han, W.; Tang, S.; et al. Extracellular Vesicles in Infrapatellar Fat Pad from Osteoarthritis Patients Impair Cartilage Metabolism and Induce Senescence. Adv. Sci. 2024, 11, e2303614. [Google Scholar] [CrossRef]
- Gao, K.; Zhu, W.; Li, H.; Ma, D.; Liu, W.; Yu, W.; Wang, L.; Cao, Y.; Jiang, Y. Association between cytokines and exosomes in synovial fluid of individuals with knee osteoarthritis. Mod. Rheumatol. 2020, 30, 758–764. [Google Scholar] [CrossRef] [PubMed]
- Bakinowska, E.; Kielbowski, K.; Pawlik, A. The Role of Extracellular Vesicles in the Pathogenesis and Treatment of Rheumatoid Arthritis and Osteoarthritis. Cells 2023, 12, 2716. [Google Scholar] [CrossRef]
- Asghar, S.; Litherland, G.J.; Cole, J.J.; McInnes, I.B.; Meek, R.M.D.; Lockhart, J.C.; Goodyear, C.S.; Crilly, A. Small extracellular vesicles derived from synovial fibroblasts contain distinct miRNA profiles and contribute to chondrocyte damage in osteoarthritis. Arthritis Res. Ther. 2024, 26, 167. [Google Scholar] [CrossRef]
- Distler, J.H.; Jungel, A.; Huber, L.C.; Seemayer, C.A.; Reich, C.F., 3rd; Gay, R.E.; Michel, B.A.; Fontana, A.; Gay, S.; Pisetsky, D.S.; et al. The induction of matrix metalloproteinase and cytokine expression in synovial fibroblasts stimulated with immune cell microparticles. Proc. Natl. Acad. Sci. USA 2005, 102, 2892–2897. [Google Scholar] [CrossRef] [PubMed]
- Guermazi, A.; Roemer, F.W.; Hayashi, D.; Crema, M.D.; Niu, J.; Zhang, Y.; Marra, M.D.; Katur, A.; Lynch, J.A.; El-Khoury, G.Y.; et al. Assessment of synovitis with contrast-enhanced MRI using a whole-joint semiquantitative scoring system in people with, or at high risk of, knee osteoarthritis: The MOST study. Ann. Rheum. Dis. 2011, 70, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Boere, J.; van de Lest, C.H.; Libregts, S.F.; Arkesteijn, G.J.; Geerts, W.J.; Nolte-‘t Hoen, E.N.; Malda, J.; van Weeren, P.R.; Wauben, M.H. Synovial fluid pretreatment with hyaluronidase facilitates isolation of CD44+ extracellular vesicles. J. Extracell. Vesicles 2016, 5, 31751. [Google Scholar] [CrossRef]
- Price, J.M.J.; Hisada, Y.; Hazeldine, J.; Bae-Jump, V.; Luther, T.; Mackman, N.; Harrison, P. Detection of tissue factor-positive extracellular vesicles using the ExoView R100 system. Res. Pract. Thromb. Haemost. 2023, 7, 100177. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, S.; Naz, S.I.; Jain, V.; Soderblom, E.J.; Aliferis, C.; Kraus, V.B. Comprehensive characterization of pathogenic synovial fluid extracellular vesicles from knee osteoarthritis. Clin. Immunol. 2023, 257, 109812. [Google Scholar] [CrossRef]
- Li, Z.; Chen, B. DUSP4 alleviates LPS-induced chondrocyte injury in knee osteoarthritis via the MAPK signaling pathway. Exp. Ther. Med. 2021, 22, 1401. [Google Scholar] [CrossRef]
- Zheng, L.; Gu, M.; Li, X.; Hu, X.; Chen, C.; Kang, Y.; Pan, B.; Chen, W.; Xian, G.; Wu, X.; et al. ITGA5+ synovial fibroblasts orchestrate proinflammatory niche formation by remodelling the local immune microenvironment in rheumatoid arthritis. Ann. Rheum. Dis. 2024, 84, 232–252. [Google Scholar] [CrossRef]
- Bonnet, C.S.; Walsh, D.A. Osteoarthritis, angiogenesis and inflammation. Rheumatology 2005, 44, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.A.; McWilliams, D.F.; Turley, M.J.; Dixon, M.R.; Franses, R.E.; Mapp, P.I.; Wilson, D. Angiogenesis and nerve growth factor at the osteochondral junction in rheumatoid arthritis and osteoarthritis. Rheumatology 2010, 49, 1852–1861. [Google Scholar] [CrossRef]
- Ashraf, S.; Walsh, D.A. Angiogenesis in osteoarthritis. Curr. Opin. Rheumatol. 2008, 20, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Yudoh, K.; Masuko, K. The potential role of vascular endothelial growth factor (VEGF) in cartilage: How the angiogenic factor could be involved in the pathogenesis of osteoarthritis? Osteoarthr. Cartil. 2008, 16, 279–286. [Google Scholar] [CrossRef]
- Hamilton, J.L.; Nagao, M.; Levine, B.R.; Chen, D.; Olsen, B.R.; Im, H.J. Targeting VEGF and Its Receptors for the Treatment of Osteoarthritis and Associated Pain. J. Bone Miner. Res. 2016, 31, 911–924. [Google Scholar] [CrossRef]
- Bujia, J.; Behrends, U.; Rotter, N.; Pitzke, P.; Wilmes, E.; Hammer, C. Expression of ICAM-1 on intact cartilage and isolated chondrocytes. In Vitro Cell. Dev. Biol. Anim. 1996, 32, 116–122. [Google Scholar] [CrossRef]
- Ootake, T.; Ishii, T.; Sueishi, K.; Watanabe, A.; Ishizuka, Y.; Amano, K.; Nagao, M.; Nishimura, K.; Nishii, Y. Effects of mechanical stress and deficiency of dihydrotestosterone or 17β-estradiol on Temporomandibular Joint Osteoarthritis in mice. Osteoarthr. Cartil. 2021, 29, 1575–1589. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Branicky, R.; Noe, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Kim, Y.W.; Byzova, T.V. Oxidative stress in angiogenesis and vascular disease. Blood 2014, 123, 625–631. [Google Scholar]
- Kim, Y.W.; West, X.Z.; Byzova, T.V. Inflammation and oxidative stress in angiogenesis and vascular disease. J. Mol. Med. 2013, 91, 323–328. [Google Scholar]
- Scott, J.L.; Gabrielides, C.; Davidson, R.K.; Swingler, T.E.; Clark, I.M.; Wallis, G.A.; Boot-Handford, R.P.; Kirkwood, T.B.; Taylor, R.W.; Young, D.A. Superoxide dismutase downregulation in osteoarthritis progression and end-stage disease. Ann. Rheum. Dis. 2010, 69, 1502–1510. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Romero, C.; Calamia, V.; Mateos, J.; Carreira, V.; Martinez-Gomariz, M.; Fernandez, M.; Blanco, F.J. Mitochondrial dysregulation of osteoarthritic human articular chondrocytes analyzed by proteomics: A decrease in mitochondrial superoxide dismutase points to a redox imbalance. Mol. Cell. Proteom. 2009, 8, 172–189. [Google Scholar] [CrossRef] [PubMed]
- Buzas, E.I.; Gyorgy, B.; Nagy, G.; Falus, A.; Gay, S. Emerging role of extracellular vesicles in inflammatory diseases. Nat. Rev. Rheumatol. 2014, 10, 356–364. [Google Scholar] [PubMed]
- Baker, K.; Grainger, A.; Niu, J.; Clancy, M.; Guermazi, A.; Crema, M.; Hughes, L.; Buckwalter, J.; Wooley, A.; Nevitt, M.; et al. Relation of synovitis to knee pain using contrast-enhanced MRIs. Ann. Rheum. Dis. 2010, 69, 1779–1783. [Google Scholar] [CrossRef]
- de Lange-Brokaar, B.J.; Ioan-Facsinay, A.; Yusuf, E.; Visser, A.W.; Kroon, H.M.; van Osch, G.J.; Zuurmond, A.M.; Stojanovic-Susulic, V.; Bloem, J.L.; Nelissen, R.G.; et al. Association of pain in knee osteoarthritis with distinct patterns of synovitis. Arthritis Rheumatol. 2015, 67, 733–740. [Google Scholar] [CrossRef]
- Wijesinghe, S.N.; Ditchfield, C.; Flynn, S.; Agrawal, J.; Davis, E.T.; Dajas-Bailador, F.; Chapman, V.; Jones, S.W. Immunomodulation and fibroblast dynamics driving nociceptive joint pain within inflammatory synovium: Unravelling mechanisms for therapeutic advancements in osteoarthritis. Osteoarthr. Cartil. 2024, 32, 1358–1370. [Google Scholar] [CrossRef]
- Orita, S.; Koshi, T.; Mitsuka, T.; Miyagi, M.; Inoue, G.; Arai, G.; Ishikawa, T.; Hanaoka, E.; Yamashita, K.; Yamashita, M.; et al. Associations between proinflammatory cytokines in the synovial fluid and radiographic grading and pain-related scores in 47 consecutive patients with osteoarthritis of the knee. BMC Musculoskelet. Disord. 2011, 12, 144. [Google Scholar] [CrossRef]
- Junger, H.; Sorkin, L.S. Nociceptive and inflammatory effects of subcutaneous TNFα. Pain 2000, 85, 145–151. [Google Scholar] [CrossRef]
- Shang, X.; Wang, Z.; Tao, H. Mechanism and therapeutic effectiveness of nerve growth factor in osteoarthritis pain. Ther. Clin. Risk Manag. 2017, 13, 951–956. [Google Scholar] [CrossRef]
- Schmelz, M.; Mantyh, P.; Malfait, A.M.; Farrar, J.; Yaksh, T.; Tive, L.; Viktrup, L. Nerve growth factor antibody for the treatment of osteoarthritis pain and chronic low-back pain: Mechanism of action in the context of efficacy and safety. Pain 2019, 160, 2210–2220. [Google Scholar]
- Gao, Y.; Hu, Z.; Huang, Y.; Liu, W.; Ren, C. Efficacy and Safety of Anti-Nerve Growth Factor Antibody Therapy for Hip and Knee Osteoarthritis: A Meta-analysis. Orthop. J. Sports Med. 2022, 10, 23259671221088590. [Google Scholar] [CrossRef]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide substance P and the immune response. Cell Mol. Life Sci. 2016, 73, 4249–4264. [Google Scholar] [CrossRef]
- Menkes, C.J.; Renoux, M.; Laoussadi, S.; Mauborgne, A.; Bruxelle, J.; Cesselin, F. Substance P levels in the synovium and synovial fluid from patients with rheumatoid arthritis and osteoarthritis. J. Rheumatol. 1993, 20, 714–717. [Google Scholar] [PubMed]
- Ko, K.R.; Lee, H.; Han, S.H.; Ahn, W.; Kim, D.K.; Kim, I.S.; Jung, B.S.; Lee, S. Substance P, A Promising Therapeutic Target in Musculoskeletal Disorders. Int. J. Mol. Sci. 2022, 23, 2583. [Google Scholar] [CrossRef]
- Warner, S.C.; Walsh, D.A.; Laslett, L.L.; Maciewicz, R.A.; Soni, A.; Hart, D.J.; Zhang, W.; Muir, K.R.; Dennison, E.M.; Leaverton, P.; et al. Pain in knee osteoarthritis is associated with variation in the neurokinin 1/substance P receptor (TACR1) gene. Eur. J. Pain 2017, 21, 1277–1284. [Google Scholar] [CrossRef]
- Conaghan, P.G.; Emerton, M.; Tennant, A. Internal construct validity of the Oxford Knee Scale: Evidence from Rasch measurement. Arthritis Rheum. 2007, 57, 1363–1367. [Google Scholar] [CrossRef]
- Neufeld, M.E.; Masri, B.A. Can the Oxford Knee and Hip Score identify patients who do not require total knee or hip arthroplasty? Bone Jt. J. 2019, 101-B, 23–30. [Google Scholar] [CrossRef]
- Wailoo, A.; Hernandez Alava, M.; Escobar Martinez, A. Modelling the relationship between the WOMAC Osteoarthritis Index and EQ-5D. Health Qual. Life Outcomes 2014, 12, 37. [Google Scholar] [CrossRef]
- Hunter, D.J.; Guermazi, A.; Roemer, F.; Zhang, Y.; Neogi, T. Structural correlates of pain in joints with osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1170–1178. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mild/Moderate OA | Severe OA | p Value | |
|---|---|---|---|
| n = 7 | n = 4 | ||
| OKS 1 | 26.8 ± 3.3 | 15.25 ± 3.38 | * p = 0.044 |
| EQ5D(sum) 2 | 7.6 ± 0.48 | 8.750 ± 0.63 | p = 0.172 |
| VAS 3 | 6.6 ± 0.93 | 5.667 ± 1.86 | p = 0.610 |
| Synovitis 4 | 16.0 ± 1.07 | 16.5 ± 0.65 | p = 0.698 |
| Age | 64.0 ± 6 | 57.5 ± 6 | p = 0.503 |
| Sex (male:female) | 5:2 | 1:3 | |
| BMI 5 | 32.2 ± 2.3 | 35.2 ± 2.5 | p = 0.421 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ditchfield, C.; Price, J.; Davis, E.T.; Jones, S.W. Synovial Fluid Extracellular Vesicles from Patients with Severe Osteoarthritis Differentially Promote a Pro-Catabolic, Inflammatory Chondrocyte Phenotype. Biomolecules 2025, 15, 829. https://doi.org/10.3390/biom15060829
Ditchfield C, Price J, Davis ET, Jones SW. Synovial Fluid Extracellular Vesicles from Patients with Severe Osteoarthritis Differentially Promote a Pro-Catabolic, Inflammatory Chondrocyte Phenotype. Biomolecules. 2025; 15(6):829. https://doi.org/10.3390/biom15060829
Chicago/Turabian StyleDitchfield, Caitlin, Joshua Price, Edward T. Davis, and Simon W. Jones. 2025. "Synovial Fluid Extracellular Vesicles from Patients with Severe Osteoarthritis Differentially Promote a Pro-Catabolic, Inflammatory Chondrocyte Phenotype" Biomolecules 15, no. 6: 829. https://doi.org/10.3390/biom15060829
APA StyleDitchfield, C., Price, J., Davis, E. T., & Jones, S. W. (2025). Synovial Fluid Extracellular Vesicles from Patients with Severe Osteoarthritis Differentially Promote a Pro-Catabolic, Inflammatory Chondrocyte Phenotype. Biomolecules, 15(6), 829. https://doi.org/10.3390/biom15060829