
Molecular Dynamics Simulations of Plasma–Antifolate Drug Synergy in Cancer Therapy



Abstract
1. Introduction
2. Methods
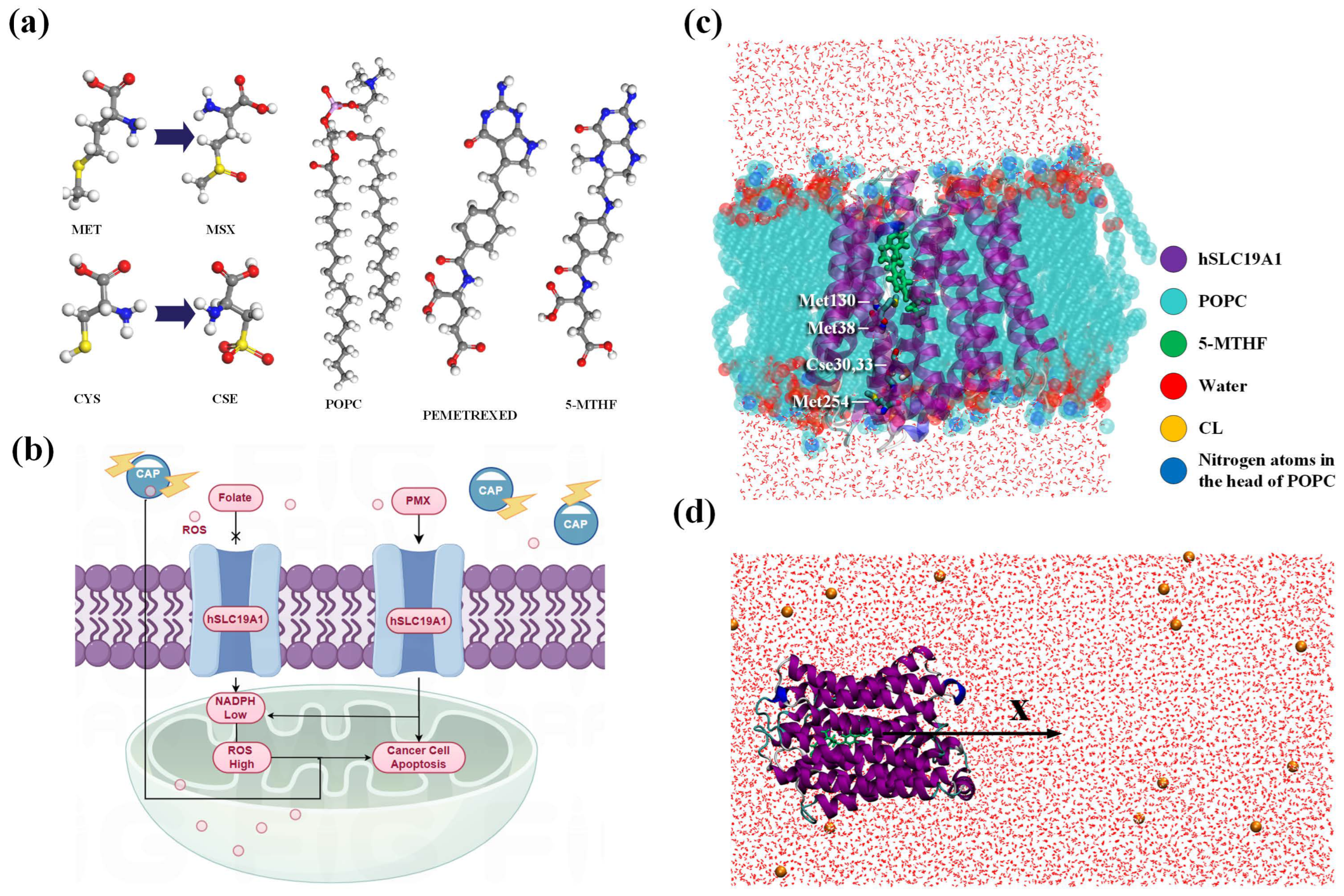
2.1. Protein Modeling
2.2. Simulation Details
2.3. Free Energy Calculations
3. Results and Discussion
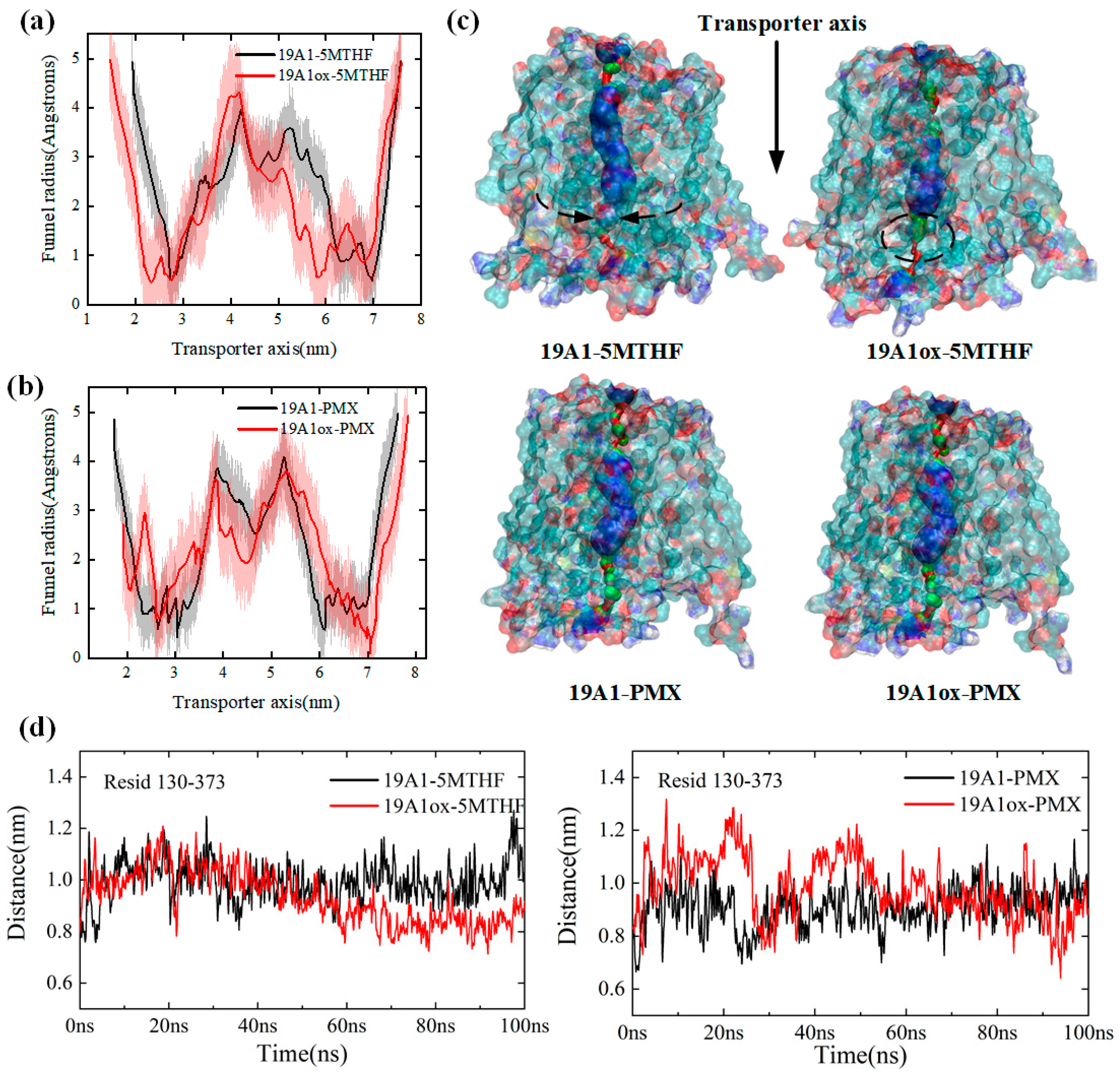
3.1. Changes in Protein Structure
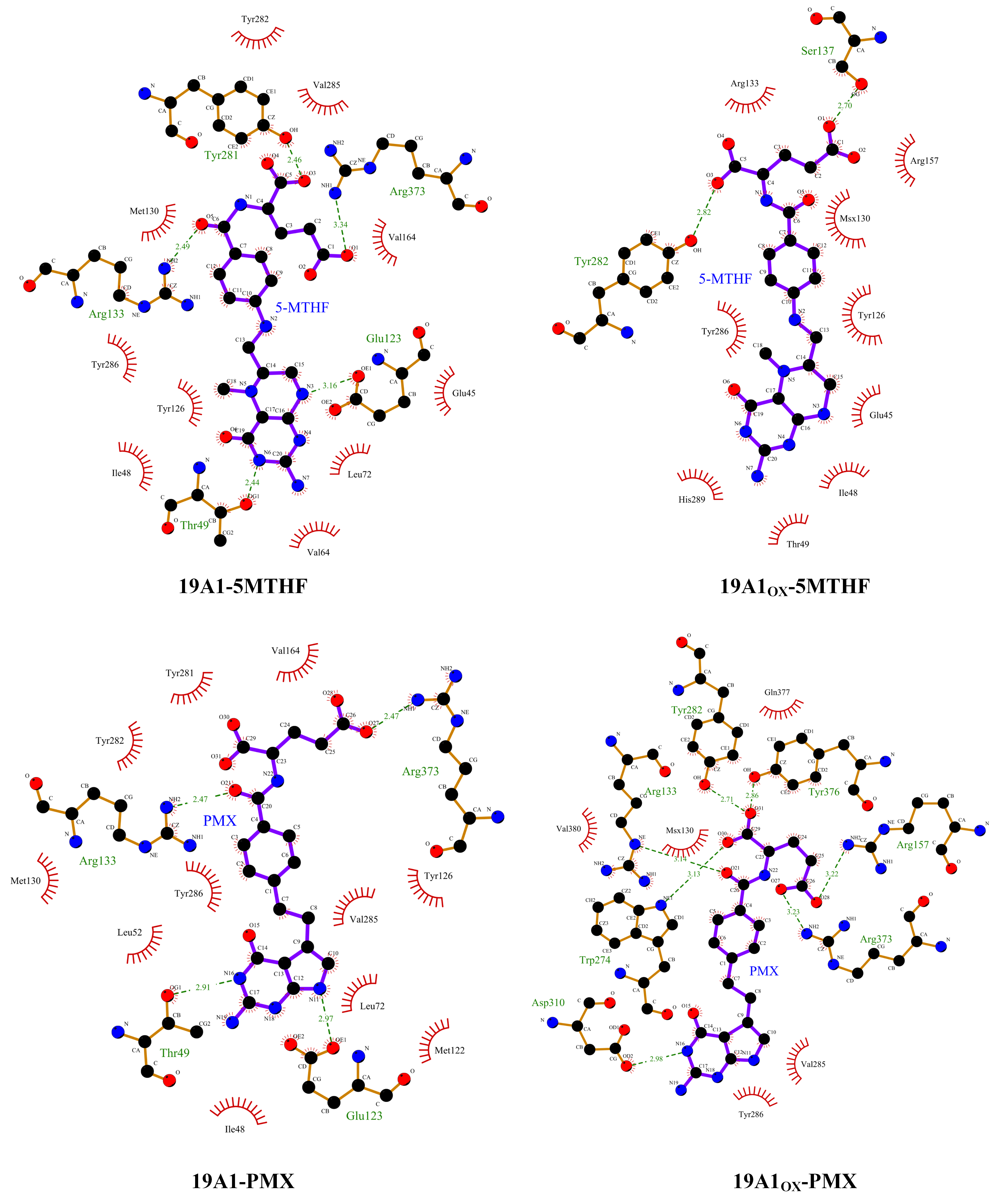
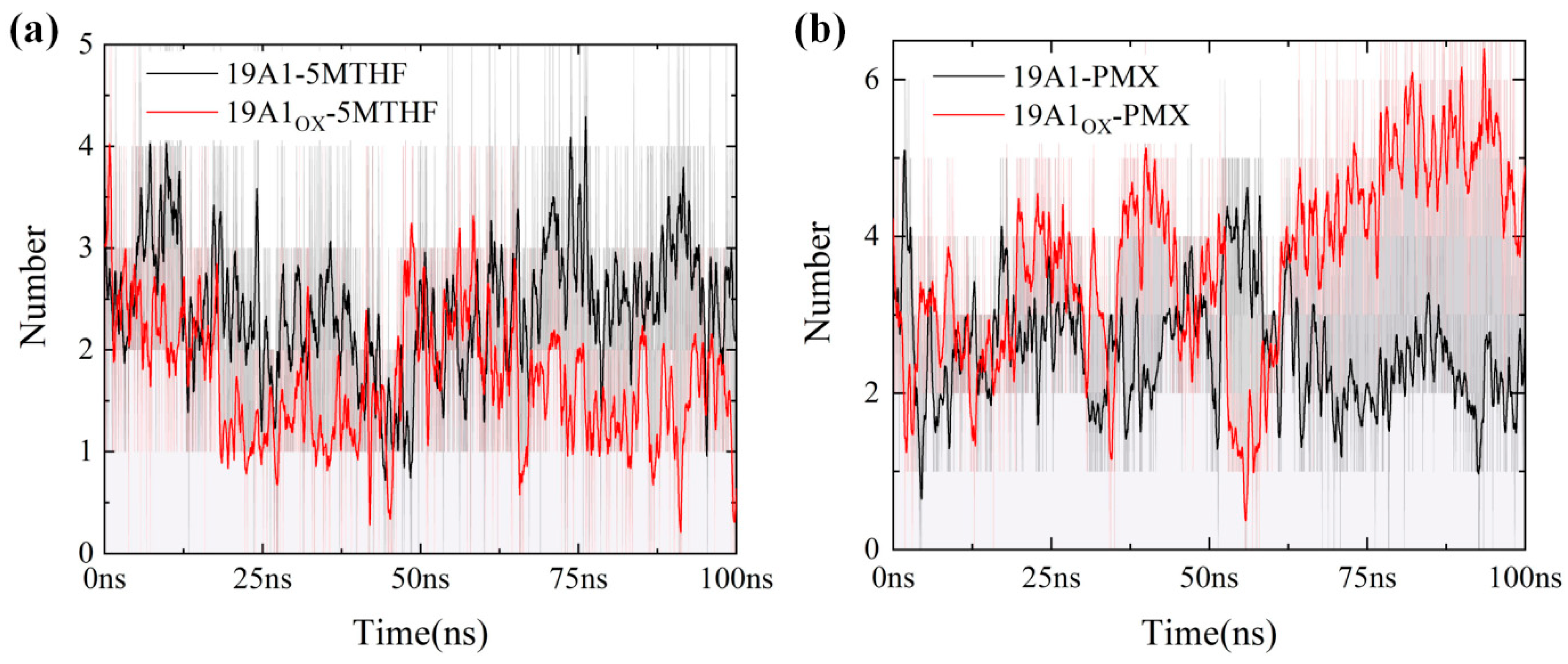
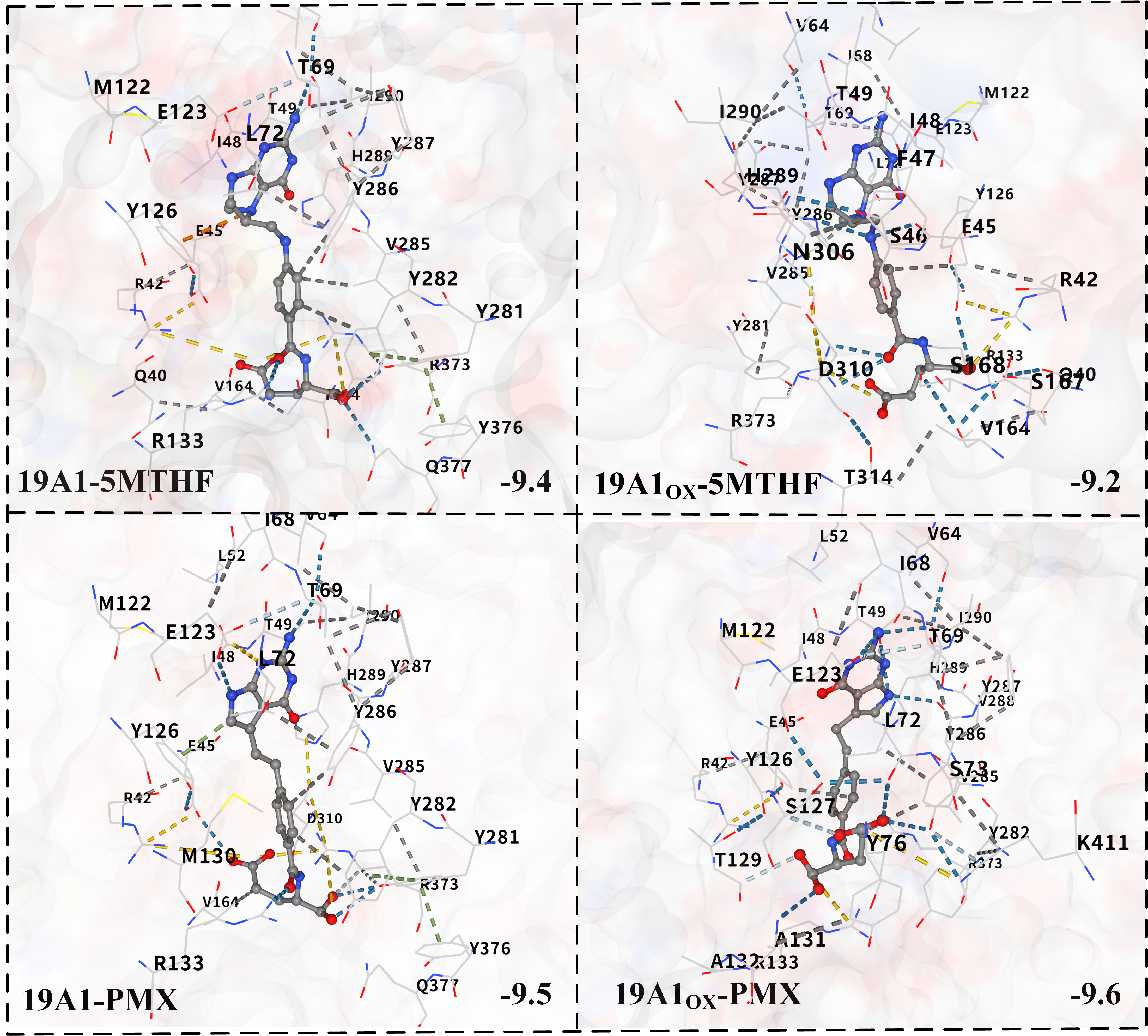
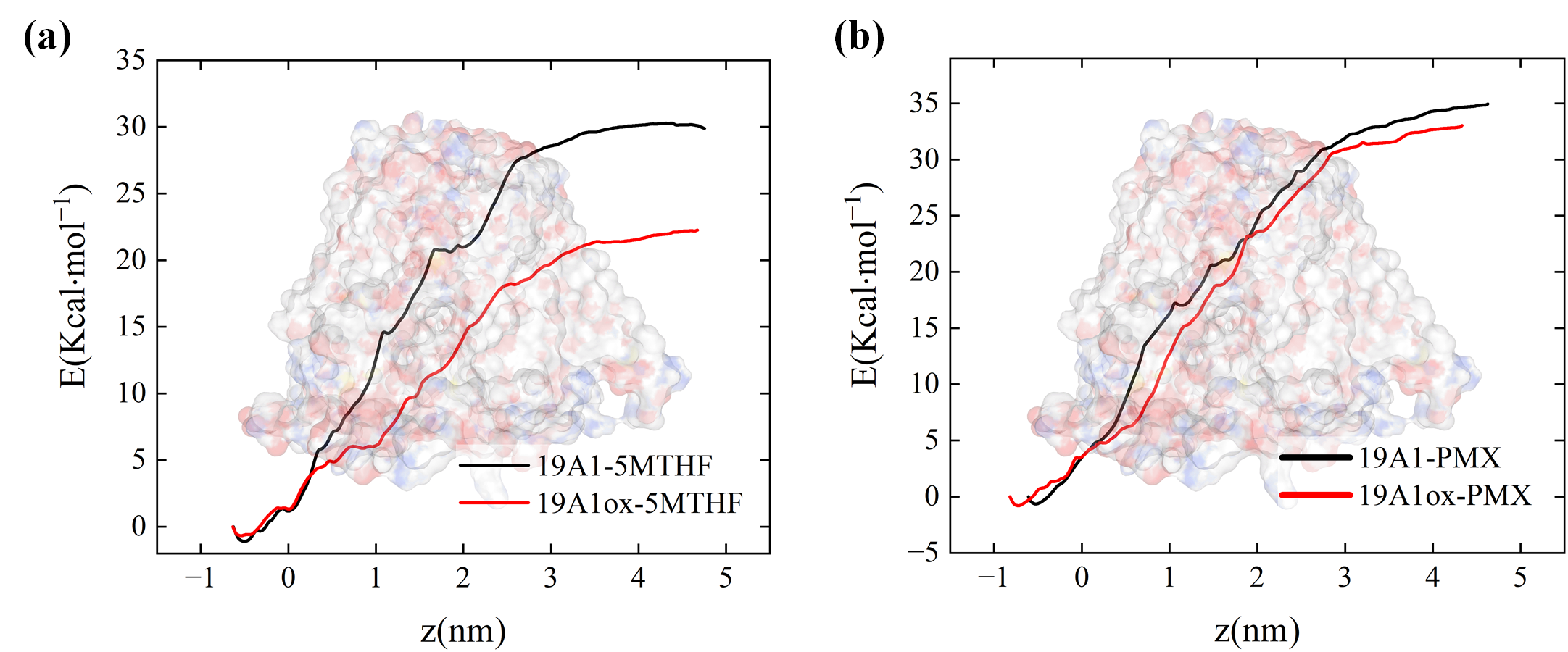
3.2. Analysis of Changes in Protein–Ligand Interactions
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Epstein, J.B.; Thariat, J.; Bensadoun, R.J.; Barasch, A.; Murphy, B.A.; Kolnicket, L.; Popplewell, L. Oral complications of cancer and cancer therapy: From cancer treatment to survivorship. CA Cancer J. Clin. 2012, 62, 400–422. [Google Scholar] [CrossRef] [PubMed]
- Keidar, M.; Walk, R.; Shashurin, A.; Srinivasan, P.; Sandleret, A.; Dasgupta, S.; Ravi, R.; Preston, R.G.; Trinkal, B. Cold plasma selectivity and the possibility of a paradigm shift in cancer therapy. Br. J. Cancer 2011, 105, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- Habor, C.S.; Li, N.Y. Cold spring harbor symposia on quantitative biology. Cold Spring Harb. Symp. Quant. Biol. 1979, 43, 1197–1208. [Google Scholar]
- Lee, H.J.; Shon, C.H.; Kim, Y.S.; Kim, S.; Kim, G.C.; Kong, M.G. Degradation of adhesion molecules of G361 melanoma cells by a non-thermal atmospheric pressure microplasma. New J. Phys. 2009, 11, 115026. [Google Scholar] [CrossRef]
- Utsumi, F.; Kajiyama, H.; Nakamura, K.; Tanaka, H.; Mizuno, M.; Ishikawa, K.; Kondo, H.; Kano, H.; Hori, M.; Kikka, F. Effect of indirect nonequilibrium atmospheric pressure plasma on anti-proliferative activity against chronic chemo-resistant ovarian cancer cells in vitro and in vivo. PLoS ONE 2013, 8, e81576. [Google Scholar] [CrossRef]
- Mohades, S.; Barekzi, N.; Laroussi, M. Efficacy of low temperature plasma against SCaBER cancer cells. Plasma Process. Polym. 2014, 11, 1150–1155. [Google Scholar] [CrossRef]
- Perrotti, V.; Caponio, V.C.A.; Muzio, L.L.; Choi, E.H.; Marcantonio, M.C.; Mazzone, M.; Kaushik, N.K.; Mincione, G. Open questions in cold atmospheric plasma treatment in head and neck cancer: A systematic review. Int. J. Mol. Sci. 2022, 23, 10238. [Google Scholar] [CrossRef]
- Lin, A.; Chernets, N.; Han, J.; Alicea, Y.; Dobrynin, D.; Fridman, G.; Freeman, T.A.; Fridman, A.; Miller, V. Non-equilibrium dielectric barrier discharge treatment of mesenchymal stem cells: Charges and reactive oxygen species play the major role in cell death. Plasma Process. Polym. 2015, 12, 1117–1127. [Google Scholar] [CrossRef]
- Remon, J.; Besse, B.; Aix, S.P.; Callejo, A.; Rabi, K.A.; Bernabe, R.; Greillier, L.; Majem, M.; Reguart, N.; Monnet, I.; et al. Osimertinib treatment based on plasma T790M monitoring in patients with EGFR-mutant non-small-cell lung cancer (NSCLC): EORTC Lung Cancer Group 1613 APPLE phase II randomized clinical trial. Ann. Oncol. 2023, 34, 468–476. [Google Scholar] [CrossRef]
- Yun, J.H.; Yang, Y.H.; Han, C.H.; Kang, S.U.; Kim, C.H. Non-thermal atmospheric pressure plasma induces selective cancer cell apoptosis by modulating redox homeostasis. Cell Commun. Signal. 2024, 22, 452. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.D.; Dinkova-Kostova, A.T.; Tew, K.D. Oxidative stress in cancer. Cancer Cell 2020, 38, 167–197. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Z.; Hoshino, A.; Zheng, H.D.; Morley, M.; Arany, Z.; Rabinowitz, J.D. NADPH production by the oxidative pentose-phosphate pathway supports folate metabolism. Nat. Metab. 2019, 1, 404–415. [Google Scholar] [CrossRef]
- Ebisch, I.M.W.; Thomas, C.M.G.; Peters, W.H.M.; Braat, D.D.M.; Steegers-Theunissen, R.P.M. The importance of folate, zinc and antioxidants in the pathogenesis and prevention of subfertility. Hum. Reprod. Update 2007, 13, 163–174. [Google Scholar] [CrossRef]
- Ducker, G.S.; Rabinowitz, J.D. One-carbon metabolism in health and disease. Cell Metab. 2017, 25, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.C.; Chang, W.M.; Wu, J.Y.; Huang, C.C.; Lu, F.J.; Chuang, Y.W.; Chang, P.J.; Chen, K.H.; Hong, C.Z.; Yeh, R.H.; et al. Folate deficiency triggered apoptosis of synoviocytes: Role of overproduction of reactive oxygen species generated via NADPH oxidase/mitochondrial complex II and calcium perturbation. PLoS ONE 2016, 11, e0146440. [Google Scholar] [CrossRef]
- Lee, D.; Xu, I.M.J.; Chiu, D.K.C.; Lai, R.K.; Tse, A.P.; Li, L.L.; Law, C.; Tsang, F.H.; Wei, L.L.; Chan, C.Y.; et al. Folate cycle enzyme MTHFD1L confers metabolic advantages in hepatocellular carcinoma. J. Clin. Investig. 2017, 127, 1856–1872. [Google Scholar] [CrossRef]
- Ducker, G.S.; Chen, L.; Morscher, R.J.; Ghergurovich, J.M.; Esposito, M.; Teng, X.; Kang, Y.; Rabinowitz, J.D. Reversal of cytosolic one-carbon flux compensates for loss of the mitochondrial folate pathway. Cell Metab. 2016, 23, 1140–1153. [Google Scholar] [CrossRef]
- Ye, Y.L.; Chan, Y.T.; Liu, H.C.; Lu, H.T.; Huang, R.F.S. Depleted folate pool and dysfunctional mitochondria associated with defective mitochondrial folate proteins sensitize Chinese ovary cell mutants to tert-butylhydroperoxide-induced oxidative stress and apoptosis. J. Nutr. Biochem. 2010, 21, 793–800. [Google Scholar] [CrossRef]
- Chern, C.L.; Huang, R.F.S.; Chen, Y.H.; Cheng, J.T.; Liu, T.Z. Folate deficiency-induced oxidative stress and apoptosis are mediated via homocysteine-dependent overproduction of hydrogen peroxide and enhanced activation of NF-κB in human Hep G2 cells. Biomed. Pharmacother. 2001, 55, 434–442. [Google Scholar] [CrossRef]
- Zhao, S.; Xiong, Z.; Mao, X.; Meng, D.; Lei, Q.; Li, Y.; Deng, P.; Chen, M.; Tu, M.; Lu, X.; et al. Atmospheric pressure room temperature plasma jets facilitate oxidative and nitrative stress and lead to endoplasmic reticulum stress dependent apoptosis in HepG2 cells. PLoS ONE 2013, 8, e73665. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G.; Sersenová, D.; Graves, D.B.; Machala, Z. Cold atmospheric plasma and plasma-activated medium trigger RONS-based tumor cell apoptosis. Sci. Rep. 2019, 9, 14210. [Google Scholar] [CrossRef] [PubMed]
- Bekeschus, S.; Eisenmann, S.; Sagwal, S.K.; Bodnar, Y.; Moritz, J.; Poschkamp, B.; Stoffels, I.; Emmert, S.; Madesh, M.; Weltmann, K.; et al. xCT (SLC7A11) expression confers intrinsic resistance to physical plasma treatment in tumor cells. Redox Biol. 2020, 30, 101423. [Google Scholar] [CrossRef]
- Moniruzzaman, R.; Rehman, M.U.; Zhao, Q.; Jawaid, P.; Mitsuhashi, Y.; Imaue, S.; Fujiwara, K.; Ogawa, R.; Tomihara, K.; Saitoh, J.; et al. Roles of intracellular and extracellular ROS formation in apoptosis induced by cold atmospheric helium plasma and X-irradiation in the presence of sulfasalazine. Free Radic. Biol. Med. 2018, 129, 537–547. [Google Scholar] [CrossRef]
- Ghasemitarei, M.; Yusupov, M.; Razzokov, J.; Shokri, B.; Bogaerts, A. Effect of oxidative stress on cystine transportation by xC− antiporter. Arch. Biochem. Biophys. 2019, 674, 108114. [Google Scholar] [CrossRef] [PubMed]
- Matherly, L.H.; Hou, Z.; Deng, Y. Human reduced folate carrier: Translation of basic biology to cancer etiology and therapy. Cancer Metastasis Rev. 2007, 26, 111–128. [Google Scholar] [CrossRef]
- Rezaei, M.; Ghasemitarei, M.; Razzokov, J.; Yusupov, M.; Ghorbanaliluet, M.; Ejtehadi, M.R. In silico study of the impact of oxidation on pyruvate transmission across the hVDAC1 protein channel. Arch. Biochem. Biophys. 2024, 751, 109835. [Google Scholar] [CrossRef]
- Margreitter, C.; Petrov, D.; Zagrovic, B. Vienna-PTM web server: A toolkit for MD simulations of protein post-translational modifications. Nucleic Acids Res. 2013, 41, W422–W426. [Google Scholar] [CrossRef]
- Margreitter, C.; Reif, M.M.; Oostenbrink, C. Update on phosphate and charged post-translationally modified amino acid parameters in the GROMOS force field. J. Comput. Chem. 2017, 38, 714–720. [Google Scholar] [CrossRef]
- Petrov, D.; Margreitter, C.; Grandits, M.; Oostenbrink, C.; Zargovic, B. A systematic framework for molecular dynamics simulations of protein post-translational modifications. PLoS Comput. Biol. 2013, 9, e1003154. [Google Scholar] [CrossRef]
- Xu, G.; Chance, M.R. Hydroxyl radical-mediated modification of proteins as probes for structural proteomics. Chem. Rev. 2007, 107, 3514–3543. [Google Scholar] [CrossRef] [PubMed]
- Takai, E.; Kitamura, T.; Kuwabara, J.; Ikawaet, S.; Yoshizawa, S.; Shiraki, K.; Kawasaki, H.; Arakawa, R.; Kitano, K. Chemical modification of amino acids by atmospheric-pressure cold plasma in aqueous solution. J. Phys. D Appl. Phys. 2014, 47, 285403. [Google Scholar] [CrossRef]
- Bruno, G.; Heusler, T.; Lackmann, J.; Woedtke, T.V.; Weltmann, K.; Wende, K. Cold physical plasma-induced oxidation of cysteine yields reactive sulfur species (RSS). Clin. Plasma Med. 2019, 14, 100083. [Google Scholar] [CrossRef]
- Zhou, R.; Zhou, R.; Zhuang, J.; Zong, Z.; Zhang, X.; Liu, D.; Bazaka, K.; Ostrikov, K. Interaction of atmospheric-pressure air microplasmas with amino acids as fundamental processes in aqueous solution. PLoS ONE 2016, 11, e0155584. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Zhou, D.; Du, X.; Zhao, H.; Lee, C.; Yang, J.; Wang, Y.; Qin, C.; Guo, Z.; Zhang, Z. Molecular mechanism of substrate recognition by folate transporter SLC19A1. Cell Discov. 2022, 8, 141. [Google Scholar] [CrossRef]
- Malde, A.K.; Zuo, L.; Breeze, M.; Stroet, M.; Poger, D.; Nair, P.C.; Oostenbrink, C.; Mark, A.E. An automated force field topology builder (ATB) and repository: Version 1.0. J. Chem. Theory Comput. 2011, 7, 4026–4037. [Google Scholar] [CrossRef]
- Stroet, M.; Caron, B.; Engler, M.S.; Woning, J.V.D.; Kauffmann, A.; Dijk, M.V.; EI-Kebir, M.; Visscher, K.M.; Holownia, J.; Macfarlane, C.; et al. OFraMP: A fragment-based tool to facilitate the parametrization of large molecules. J. Comput.-Aided Mol. Des. 2023, 37, 357–371. [Google Scholar] [CrossRef]
- Stansfeld, P.J.; Sansom, M.S.P. Molecular simulation approaches to membrane proteins. Structure 2011, 19, 1562–1572. [Google Scholar] [CrossRef]
- Nagarajan, S.K.; Babu, S.; Kulkarni, S.A.; Vadivelu, A.; Devaraju, P.; Sohn, H.; Madhavan, T. Understanding the influence of lipid bilayers and ligand molecules in determining the conformational dynamics of somatostatin receptor 2. Sci. Rep. 2021, 11, 7677. [Google Scholar] [CrossRef]
- Frenkel, D.; Smit, B. Understanding Molecular Simulation; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Knapp, B.; Ospina, L.; Deane, C.M. Avoiding false positive conclusions in molecular simulation: The importance of replicas. J. Chem. Theory Comput. 2018, 14, 6127–6138. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.; Bhati, A.P.; Zasada, S.J.; Conveney, P.V. Rapid, accurate, precise and reproducible ligand–protein binding free energy prediction. Interface Focus 2020, 10, 20200007. [Google Scholar] [CrossRef] [PubMed]
- Coveney, P.V.; Wan, S. On the calculation of equilibrium thermodynamic properties from molecular dynamics. Phys. Chem. Chem. Phys. 2016, 18, 30236–30240. [Google Scholar] [CrossRef] [PubMed]
- Hassan, A.M.; Gattan, H.S.; Faizo, A.A.; Alruhaili, M.H.; Alharbi, A.S.; Bajrai, L.H.; AL-Zahrani, I.A.; Dwivedi, V.D.; Azhar, E.I. Evaluating the Binding Potential and Stability of Drug-like Compounds with the Monkeypox Virus VP39 Protein Using Molecular Dynamics Simulations and Free Energy Analysis. Pharmaceuticals 2024, 17, 1617. [Google Scholar] [CrossRef]
- Available online: https://jerkwin.github.io/2019/07/31/gmx_mmpbsa%E4%BD%BF%E7%94%A8%E8%AF%B4%E6%98%8E/ (accessed on 15 December 2024).
- Smart, O.S.; Goodfellow, J.M.; Wallace, B.A. The pore dimensions of gramicidin A. Biophys. J. 1993, 65, 2455–2460. [Google Scholar] [CrossRef]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. Des. Sel. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand–protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, X.; Zhu, Y.; Sun, P.; Zhang, L.; Ma, J.; Zhang, Y.; Zeng, L.; Nie, X.; Gao, Y.; et al. Recognition of cyclic dinucleotides and folates by human SLC19A1. Nature 2022, 612, 170–176. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, X.; Gan, J.; Chen, S.; Xiao, Z.; Cao, Y. CB-Dock2: Improved protein–ligand blind docking by integrating cavity detection, docking and homologous template fitting. Nucleic Acids Res. 2022, 50, W159–W164. [Google Scholar] [CrossRef]
- Motaln, H.; Recek, N.; Rogelj, B. Intracellular responses triggered by cold atmospheric plasma and plasma-activated media in cancer cells. Molecules 2021, 26, 1336. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue Type | Met | Met | Met | Met | Met | Cys | Cys | Cys | Cys |
|---|---|---|---|---|---|---|---|---|---|
| Resid Number | 38 | 119 | 122 | 130 | 254 | 30 | 33 | 365 | 396 |
| SASA | 0.715 | 0.015 | 0.073 | 0.350 | 0.696 | 0.813 | 0.239 | 0.009 | 0.050 |
| 19A1-5MTHF | 19A1ox-5MTHF | 19A1-PMX | 19A1ox-PMX | |
|---|---|---|---|---|
| Protein | hSLC19A1 | hSLC19A1OX | hSLC19A1 | hSLC19A1OX |
| Ligand | 5-MTHF | 5-MTHF | PMX | PMX |
| Hydrogen Bonding Rate | Average Number of Hydrogen Bonds | ||||||
|---|---|---|---|---|---|---|---|
| OX1 | Tyr281 | Gln40 | Tyr282 | Arg133 | Tyr286 | Arg373 | 2.4197 |
| 98.95% | 53.47% | 32.93% | 27.24% | 11.94% | 10.29% | ||
| OX2 | Tyr282 | Tyr281 | Ser137 | Gln377 | Arg133 | Arg157 | 1.7067 |
| 70.36% | 36.28% | 23.54% | 15.74% | 7.05% | 5.95% | ||
| OX3 | Tyr281 | Lys411 | Thr404 | Glu123 | Tyr282 | Glu45 | 2.5364 |
| 82.21% | 68.67% | 32.08% | 27.49% | 11.84% | 8.30% | ||
| OX4 | Tyr282 | Tyr376 | Tyr281 | Gln377 | Arg373 | Glu123 | 3.6851 |
| 79.71% | 78.11% | 71.46% | 52.62% | 30.13% | 21.59% | ||
| 19A1-5MTHF | 19A1ox-5MTHF | 19A1-PMX | 19A1ox-PMX | |
|---|---|---|---|---|
| MM | −532.891 | −570.111 | −591.315 | −573.037 |
| PB | 416.934 | 512.568 | 485.418 | 460.364 |
| SA | −32.761 | −34.905 | −28.211 | −27.893 |
| COU | −331.411 | −365.373 | −449.946 | −423.8 |
| VDW | −201.480 | −204.738 | −141.369 | −149.237 |
| Tds | 25.101 | 21.878 | 16.025 | 19.384 |
| dG(kcal/mol) | −28.023 | −16.866 | −28.223 | −28.963 |
| 19A1-5MTHF | 19A1ox-5MTHF | 19A1-PMX | 19A1ox-PMX | |
|---|---|---|---|---|
| Cys30 | 0.095 | 0.857 | 0.157 | 2.228 |
| Cys33 | 0.361 | 0.995 | 0.47 | 9.619 |
| Met38 | 0.024 | 0.041 | −0.184 | −0.051 |
| Arg42 | −12.945 | 2.088 | −11.012 | −7.685 |
| Glu45 | 25.177 | 15.048 | 11.624 | 7.523 |
| Tyr126 | −8.849 | −7.746 | −0.602 | 0.688 |
| Met130 | −2.145 | −1.131 | 0.397 | −3.177 |
| Arg133 | −4.388 | 3.084 | −15.397 | −16.281 |
| Arg157 | −9.451 | −15.801 | −11.123 | −14.658 |
| Met254 | 0.014 | 0.023 | 0.070 | −0.078 |
| Tyr281 | −4.430 | −0.925 | −5.203 | −2.791 |
| Tyr282 | −5.548 | −9.930 | −6.362 | −6.548 |
| Asp310 | 12.721 | 9.721 | 5.493 | 25.672 |
| Arg373 | −2.116 | −13.064 | −2.269 | −0.414 |
| Lys411 | −5.756 | 17.921 | −7.507 | −9.973 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, Y.; Zhao, T.; Wang, X.; Sun, Y.; Zhang, Y. Molecular Dynamics Simulations of Plasma–Antifolate Drug Synergy in Cancer Therapy. Biomolecules 2025, 15, 773. https://doi.org/10.3390/biom15060773
Niu Y, Zhao T, Wang X, Sun Y, Zhang Y. Molecular Dynamics Simulations of Plasma–Antifolate Drug Synergy in Cancer Therapy. Biomolecules. 2025; 15(6):773. https://doi.org/10.3390/biom15060773
Chicago/Turabian StyleNiu, Yanxiong, Tong Zhao, Xiaolong Wang, Ying Sun, and Yuantao Zhang. 2025. "Molecular Dynamics Simulations of Plasma–Antifolate Drug Synergy in Cancer Therapy" Biomolecules 15, no. 6: 773. https://doi.org/10.3390/biom15060773
APA StyleNiu, Y., Zhao, T., Wang, X., Sun, Y., & Zhang, Y. (2025). Molecular Dynamics Simulations of Plasma–Antifolate Drug Synergy in Cancer Therapy. Biomolecules, 15(6), 773. https://doi.org/10.3390/biom15060773