
The Influence of Topiramate on Morphine Dependence in Mice

 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. General Experimental Conditions
2.2. Drugs
2.3. The Procedures of Behavioral Experiments
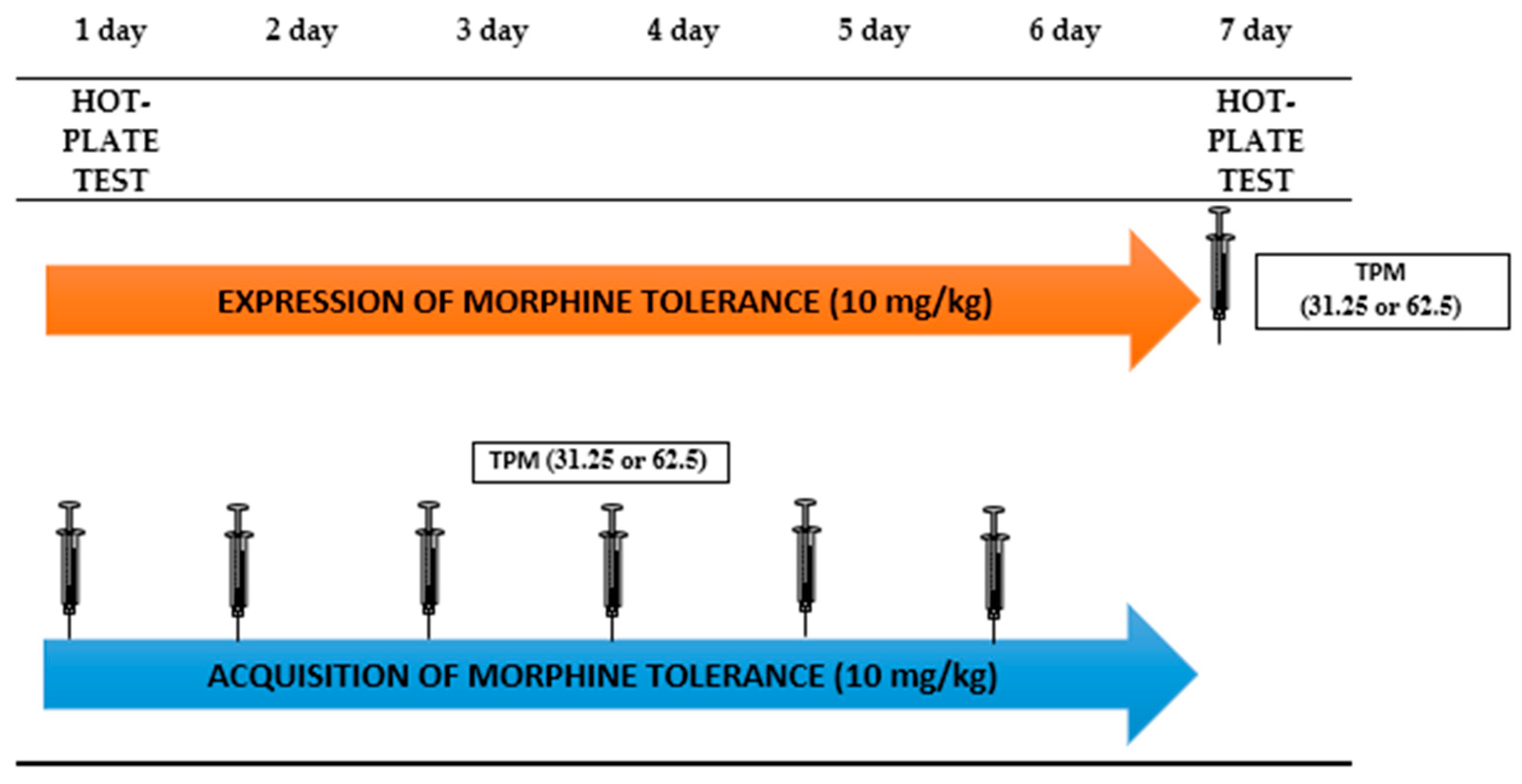
2.3.1. The Effect of Topiramate (12.5; 25 mg/kg, i.p.) on Morphine Tolerance to the Antinociceptive Effects in the Hot-Plate Test in Mice
2.3.2. The Influence of Topiramate (12.5; 25 mg/kg, i.p.) on the Severity of Morphine Withdrawal Symptoms Induced by Naloxone (2 mg/kg, i.p.) in Mice
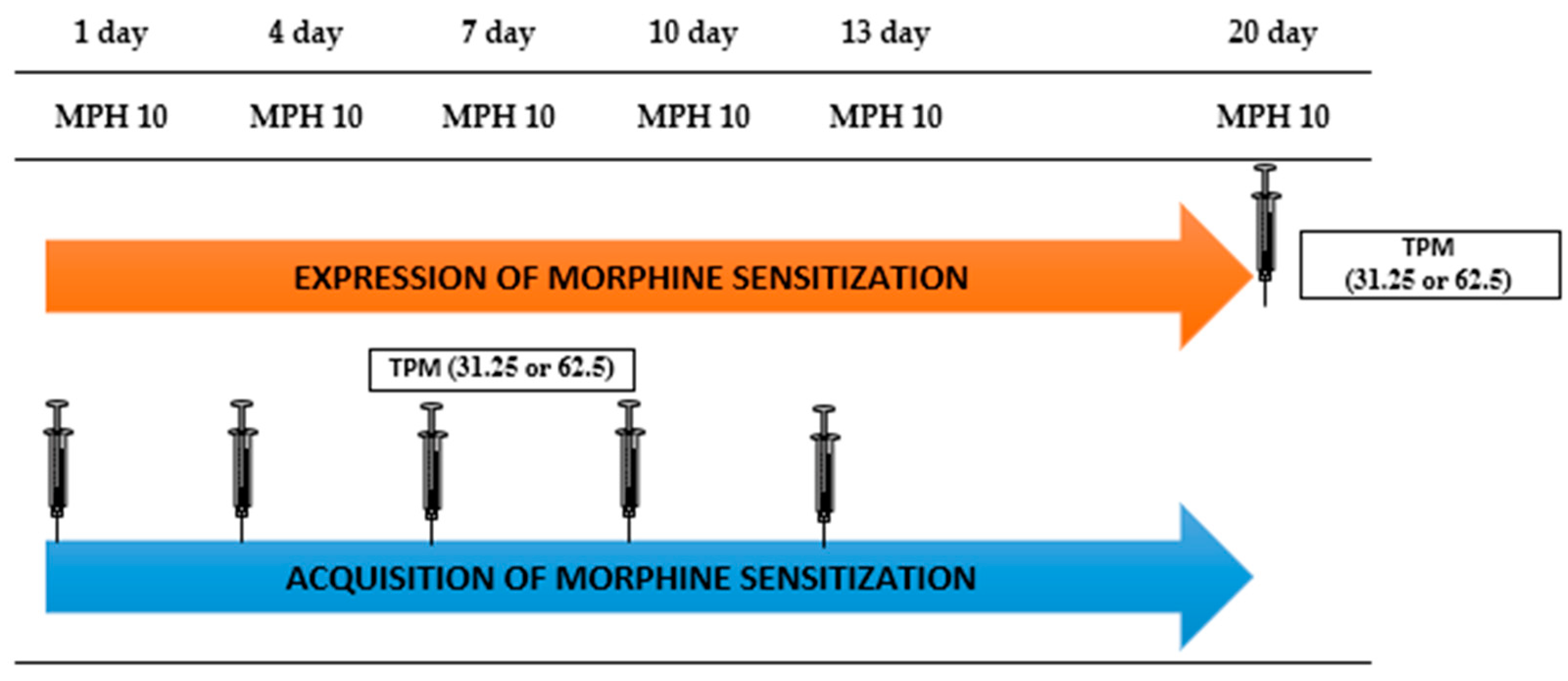
2.3.3. The Influence of Topiramate (12.5; 25 and 50 mg/kg, i.p.) on Morphine-Induced Sensitization to the Locomotor Effect in Mice
2.3.4. Statistical Analysis
3. Results
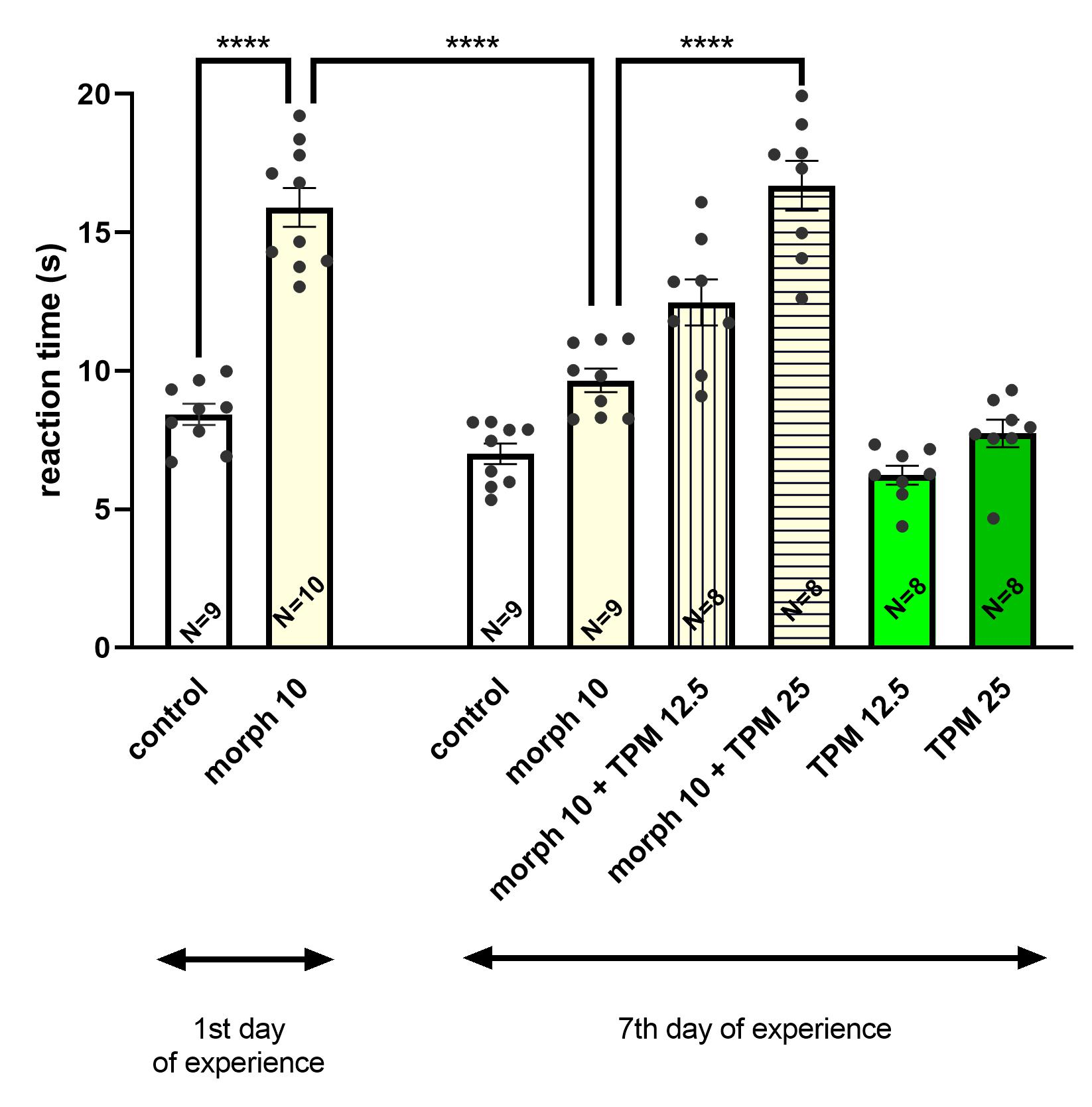
3.1. The Effect of Topiramate (12.5; 25 mg/kg, i.p.) on Morphine Tolerance to the Antinociceptive Effects in the Hot-Plate Test in Mice
3.1.1. The Influence of Topiramate on the Expression of Morphine Tolerance
3.1.2. The Influence of Topiramate on the Acquisition of Morphine Tolerance
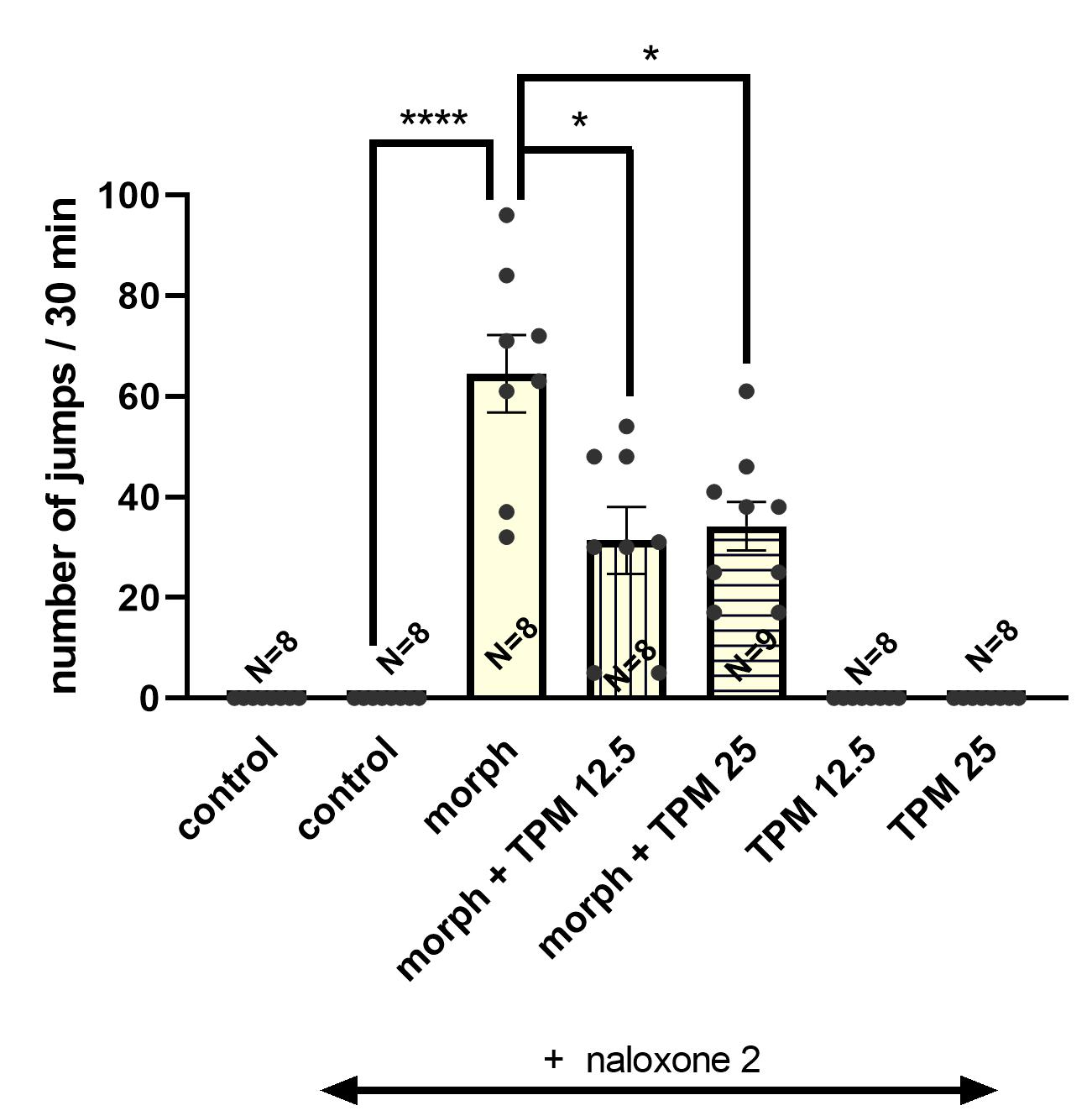
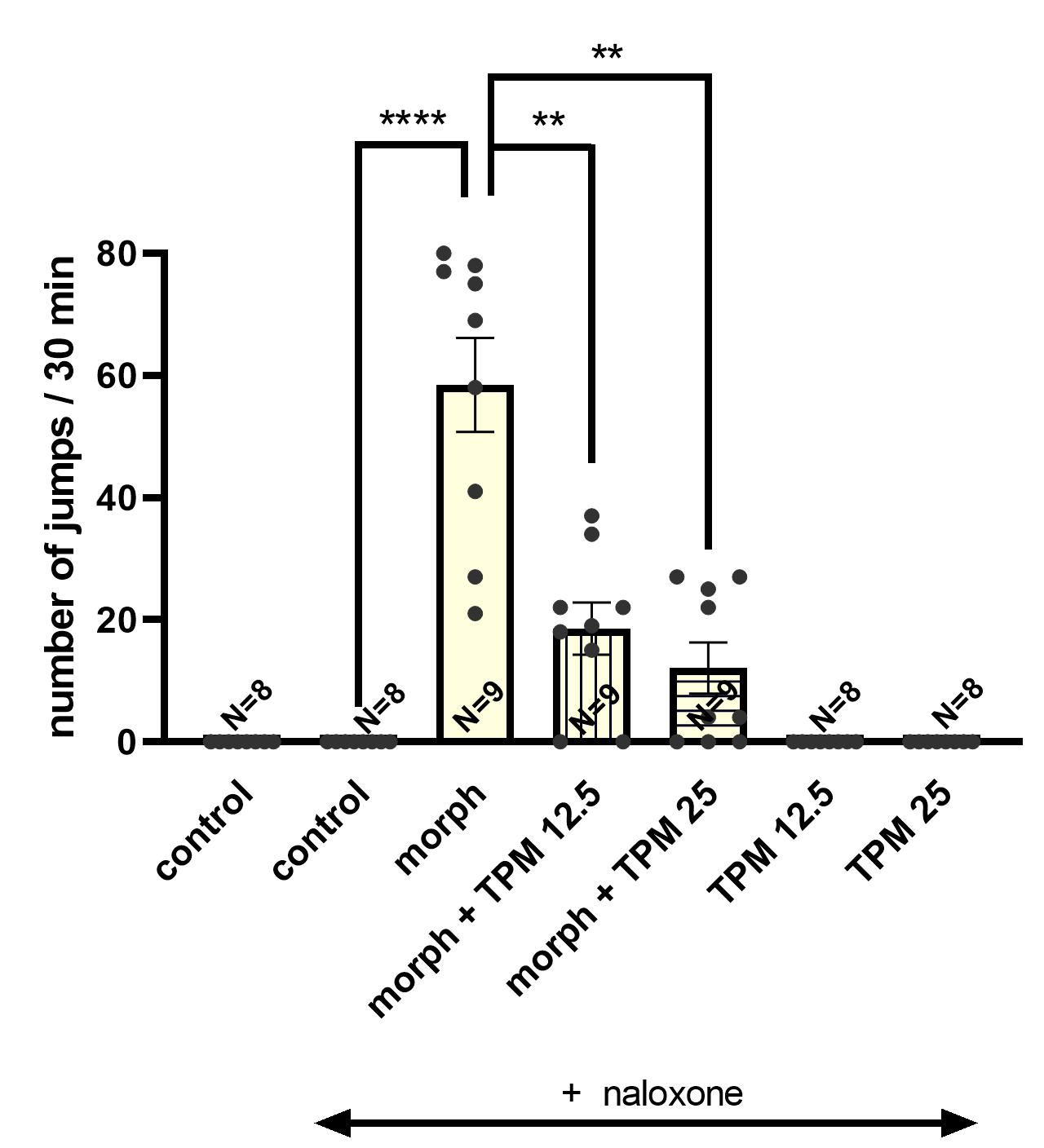
3.2. Effect of Topiramate (12.5; 25 mg/kg, i.p.) on the Severity of Morphine Withdrawal Symptoms Induced by Naloxone (2 mg/kg, i.p.) in Mice
3.2.1. The Influence of Topiramate on the Expression of Morphine Withdrawal Symptoms
3.2.2. The Influence of Topiramate on the Acquisition of Morphine Withdrawal Symptoms
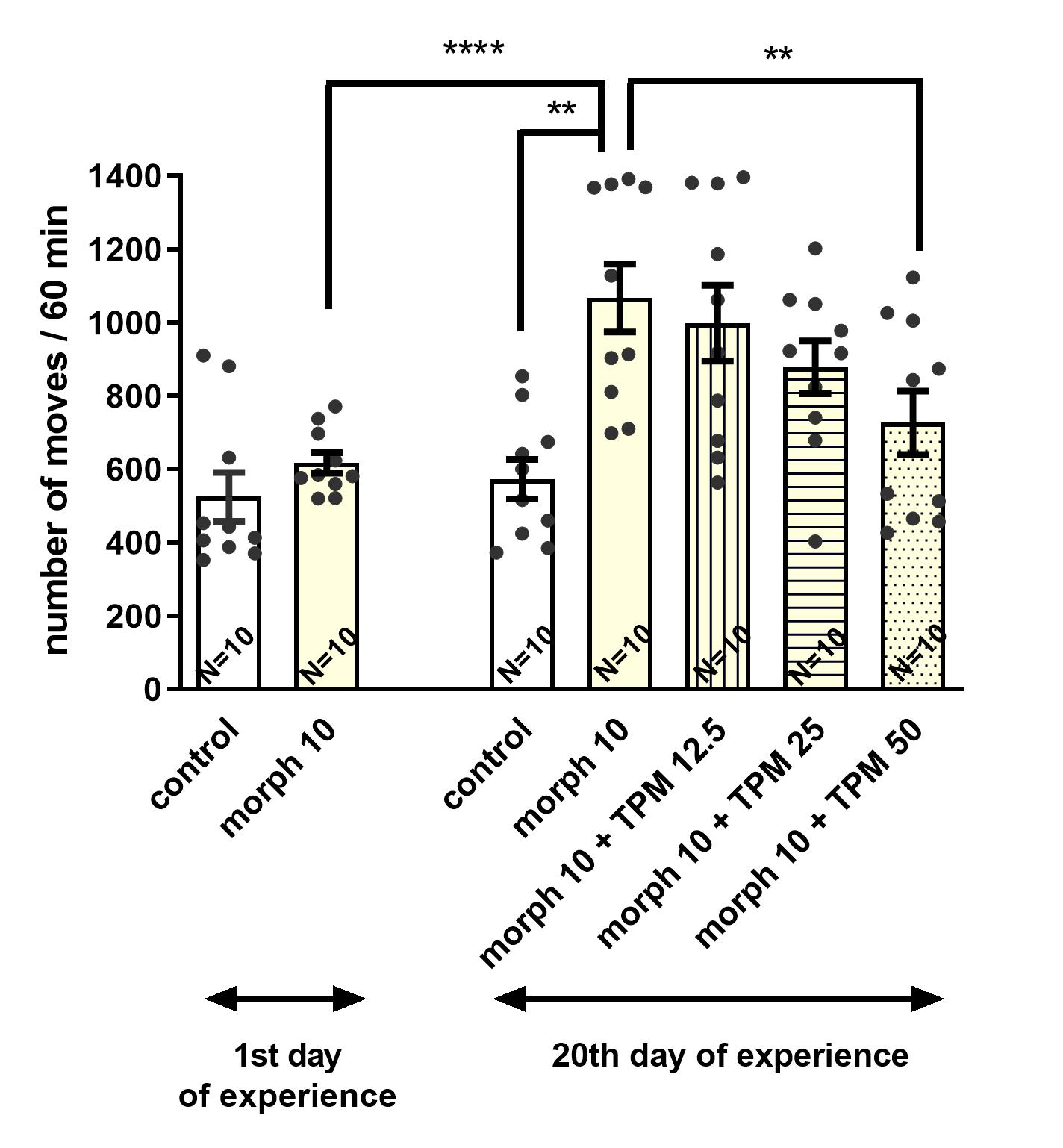
3.3. Effect of Topiramate (12.5; 25; 50 mg/kg, i.p.) on Behavioral Sensitization to the Locomotor Effects of Morphine (10 mg/kg, i.p.) in Mice
3.3.1. The Influence of Topiramate on the Expression of Morphine Sensitization
3.3.2. The Influence of Topiramate on the Acquisition of Morphine Sensitization
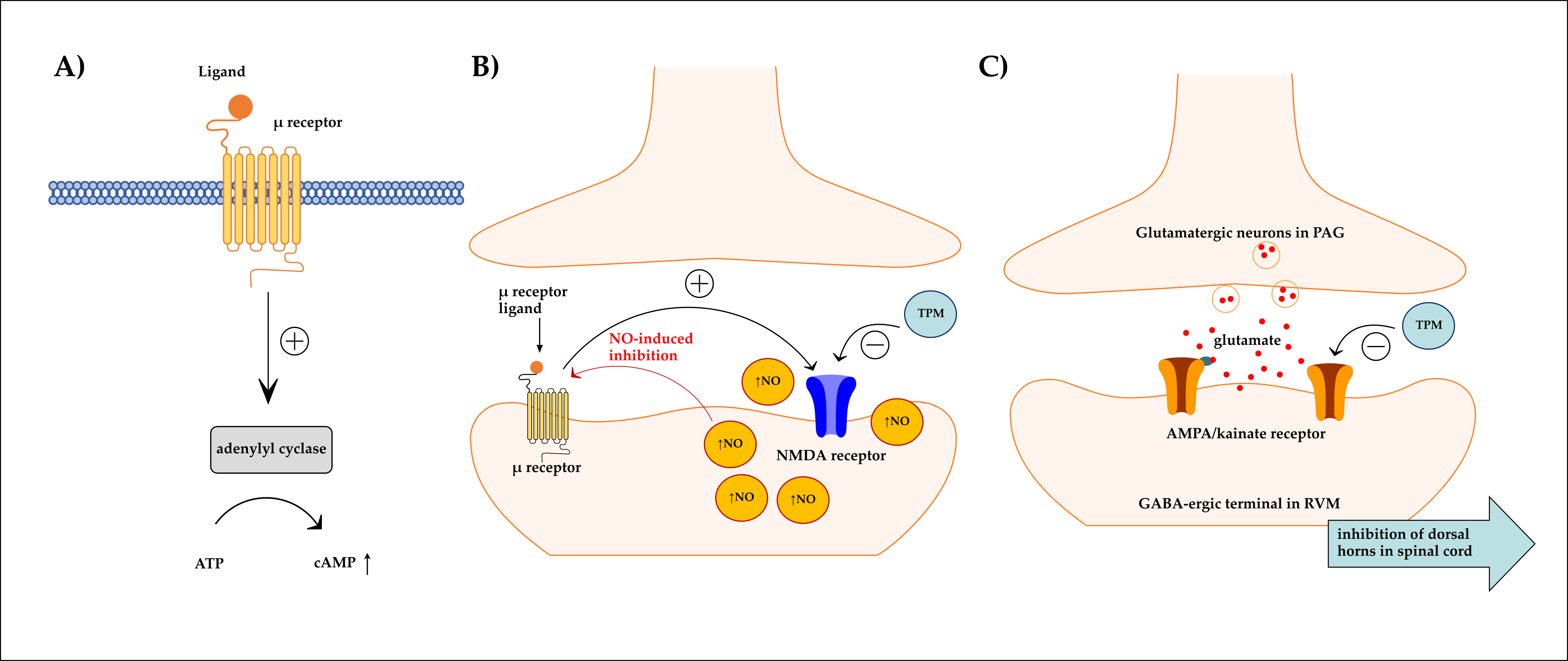
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| i.p. | intraperitoneal injection |
| GABA | gamma-aminobutyric acid |
| NMDA | N-methyl-D-aspartate |
| AMPA | aminomethylphosphonic acid |
| RVM | rostroventral medulla |
| N | the number of animals per group |
| PAG | periaqueductal gray |
| Morph | morphine |
| p | probability ratio |
| TPM | topiramate |
References
- Cherian, A.; Thomas, S.V. Status epilepticus. Ann. Indian Acad. Neurol. 2009, 12, 140–153. [Google Scholar] [PubMed]
- Wasterlain, C.G.; Chen, J.W. Mechanistic and pharmacologic aspects of status epilepticus and its treatment with new antiepileptic drugs. Epilepsia 2008, 49, 63–73. [Google Scholar] [CrossRef]
- Avoli, M.; Kawasaki, H.; Zona, C. Effects induced by topiramate on sodium electrogenesis in mammalian central neurons. Epilepsia 1996, 37, 51–52. [Google Scholar]
- Coulter, D.A.; Sombati, S.; De Lorenzo, R. Selective effects of topiramate on sustained repetitive firing and spontaneous bursting in cultured hippocampal neurons. Epilepsia 1993, 34, 123. [Google Scholar]
- Taverna, S.; Sancini, G.; Mantegazza, M.; Franceschetti, S.; Avanzini, G. Inhibition of transient and persistent Na+ current fractions by the new anticonvulsant topiramate. J. Pharmacol. Exp. Ther. 1999, 288, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.C.; Werkman, T.R.; Battefeld, A.; Clare, J.J.; Wadman, W.J. Carbamazepine and topiramate modulation of transient and persistent sodium currents studied in HEK293 cells expressing the Na(v)1.3 alpha-subunit. Epilepsia 2007, 48, 774–782. [Google Scholar]
- Gibbs, J.W., 3rd; Sombati, S.; DeLorenzo, R.J.; Coulter, D.A. Cellular actions of topiramate: Blockade of kainate-evoked inward currents in cultured hippocampal neurons. Epilepsia 2000, 41, 10–16. [Google Scholar] [CrossRef]
- Gryder, D.S.; Rogawski, M.A. Selective antagonism of GluR5 kainate receptor mediated synaptic currents by topiramate in rat basolateral amygdala neurons. J. Neurosci. 2003, 23, 7069–7074. [Google Scholar] [CrossRef]
- Severt, L.; Coulter, D.A.; Sombati, S.; DeLorenzo, R.J. Topiramate selectively blocks kainate currents in cultured hippocampal neurons. Epilepsia 1995, 36, 38–40. [Google Scholar]
- Skradski, S.; White, H.S. Topiramate blocks kainate evoked cobalt influx into cultured neurons. Epilepsia 2000, 41, 45–47. [Google Scholar] [CrossRef]
- Kanda, T.; Kurokawa, M.; Tamura, S.; Nakamura, J.; Ishii, A.; Kuwana, Y.; Serikawa, T.; Yamada, J.; Ishihara, K.; Sasa, M. Topiramate reduces abnormally high extracellular levels of glutamate and aspartate in the hippocampus of spontaneously epileptic rats (SER). Life Sci. 1996, 59, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, M.; Jiang, L.; Ma, C.; Xi, Z.; Li, X.; He, N. A comparison of extracellular excitatory amino acids release inhibition of acute lamotrigine and topiramate treatment in the hippocampus of PTZ-kindled epileptic rats. J. Biomed. Nanotechnol. 2013, 9, 1123–1128. [Google Scholar] [CrossRef]
- Petroff, O.A.; Hyder, F.; Rothman, D.L.; Mattson, R.H. Topiramate rapidly raises brain GABA in epilepsy patients. Epilepsia 2001, 42, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Gordey, M.; DeLorey, T.M.; Olsen, R.W. Differential sensitivity of recombinant GABA(A) receptors expressed in Xenopus oocytes to modulation by topiramate. Epilepsia 2000, 41, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Simeone, T.A.; Wilcox, K.S.; White, H.S. Subunit selectivity of topiramate modulation of heteromeric GABA(A) receptors. Neuropharmacology 2006, 50, 845–857. [Google Scholar] [CrossRef]
- Brown, S.D.; Wolf, H.H.; Swyniard, E.A.; Twyman, R.E.; White, H.S. The novel anticonvulsant topiramate enhances GABA-mediated chloride flux. Epilepsia 1993, 34, 122–123. [Google Scholar]
- White, H.S.; Brown, S.D.; Woodhead, J.H.; Gwendolyn, A.S.; Wolf, H.H. Topiramate enhances GABA-mediated chloride flux and GABA-evoked chloride currents in murine brain neurons and increases seizure threshold. Epilepsy Res. 1997, 28, 167–179. [Google Scholar] [CrossRef]
- White, H.S.; Brown, S.D.; Woodhead, J.H.; Skeen, G.A.; Wolf, H.H. Topiramate modulates GABA-evoked currents in murine cortical neurons by a nonbenzodiazepine mechanism. Epilepsia 2000, 41, 17–20. [Google Scholar] [CrossRef]
- Hu, C.; Zhang, Y.; Tan, G. Advances in topiramate as prophylactic treatment for migraine. Brain Behav. 2021, 11, e2290. [Google Scholar] [CrossRef]
- Linde, M.; Mulleners, W.M.; Chronicle, E.P.; McCrory, D.C. Topiramate for the prophylaxis of episodic migraine in adults. Cochrane Database Syst. Rev. 2013, 2013, CD010610. [Google Scholar] [CrossRef]
- Ashtari, F.; Shaygannejad, V.; Akbari, M. A double-blind, randomized trial of low-dose topiramate vs propranolol in migraine prophylaxis. Acta Neurol. Scand. 2008, 18, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Bagnato, F.; Good, J. The use of antiepileptics in migraine prophylaxis. Headache 2016, 56, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Storey, J.R.; Calder, C.S.; Hart, D.E.; Potter, D.L. Topiramate in migraine prevention: A double-blind, placebo-controlled study. Headache 2001, 41, 968–975. [Google Scholar] [CrossRef] [PubMed]
- Raskin, P.; Donofrio, P.D.; Rosenthal, N.R.; Hewitt, D.J.; Jordan, D.M.; Xiang, J.; Vinik, A.I. Topiramate vs placebo in painful diabetic neuropaty. Analgesic and metabolic effects. Neurology 2004, 63, 865–873. [Google Scholar] [CrossRef]
- Domingues, B.R.; Kuster, W.G.; Aquino, H.C.C. Treatment of trigeminal neuralgia with low doses of topiramate. Arq. Neuropsiquiatr. 2007, 65, 792–794. [Google Scholar] [CrossRef]
- Zvartau-Hind, M.; Din, M.U.; Gilani, A.; Lisak, R.P.; Khan, O.A. Topiramate relieves refractory trigeminal neuralgia in MS patients. Neurology 2000, 55, 1587–1588. [Google Scholar] [CrossRef]
- Alfaris, N.; Minnick, A.M.; Hopkins, C.M.; Berkowitz, R.I.; Wadden, T.A. Combination phentermine and topiramate extended release in the management of obesity. Expert Opin. Pharmacother. 2015, 16, 1263–1274. [Google Scholar] [CrossRef]
- Khera, R.; Murad, M.H.; Chandar, A.K.; Dulai, P.S.; Wang, Z.; Prokop, L.J.; Loomba, R.; Camilleri, M.; Singh, S. Association of pharmacological treatments for obesity with weight loss and adverse events: A systematic review and meta-analysis. JAMA 2016, 315, 2424–2434. [Google Scholar] [CrossRef]
- Lei, X.G.; Ruan, J.Q.; Lai, C.; Sun, Z.; Yang, X. Efficacy and safety of phentermine/topiramate in adults with overweight or obesity: A systematic review and meta-analysis. Obesity 2021, 29, 985–994. [Google Scholar] [CrossRef]
- Johnson, B.A.; Ait-Daoud, N.; Bowden, C.L.; DiClemente, C.C.; Roache, J.D.; Lawson, K.; Javors, M.A.; Ma, J.Z. Oral topiramate for treatment of alcohol dependence: A randomised controlled trial. Lancet 2003, 361, 1677–1685. [Google Scholar] [CrossRef]
- Johnson, B.A.; Rosenthal, N.; Capece, J.A.; Wiegand, F.; Mao, L.; Beyers, K.; McKay, A.; Ait-Daoud, N.; Anton, R.F.; Ciraulo, D.A.; et al. Topiramate for treating alcohol dependence: A randomized controlled trial. JAMA 2007, 298, 1641–1651. [Google Scholar] [CrossRef] [PubMed]
- Knapp, C.M.; Ciraulo, D.A.; Sarid-Segal, O.; Richardson, M.A.; Devine, E.; Streeter, C.C.; Oscar-Berman, C.M.; Surprise, C.; Colaneri, L.; Putnam, M.; et al. Zonisamide, topiramate, and levetiracetam: Efficacy and neuropsychological effects in alcohol use disorders. J. Clin. Psychopharmacol. 2015, 35, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Kampman, K.M.; Pettinati, H.; Lynch, K.G.; Dackis, C.; Sparkman, T.; Weigley, C.; O’Brien, C.P. A pilot trial of topiramate for the treatment of cocaine dependence. Drug Alcohol Depend. 2004, 75, 233–240. [Google Scholar] [CrossRef]
- Singh, M.; Keer, D.; Klimas, J.; Wood, E.; Werb, D. Topiramate for cocaine dependence: A systematic review and meta-analysis of randomized controlled trials. Addiction 2016, 111, 1337–1346. [Google Scholar] [CrossRef]
- McElroy, S.L.; Arnold, L.M.; Shapira, N.A.; Keck, P.E., Jr.; Rosenthal, N.R.; Karim, M.R.; Kamin, M.; Hudson, J.I. Topiramate in the treatment of binge eating disorder associated with obesity: A randomized, placebo-controlled trial. Am. J. Psychiatry 2003, 160, 255–261. [Google Scholar] [CrossRef]
- Nourredine, M.; Jurek, L.; Angerville, B.; Longuet, Y.; de Ternay, J.; Derveaux, A.; Rolland, B. Use of topiramate in the spectrum of addictive and eating disorders: A systematic review comparing treatment schemes, efficacy, and safety features. CNS Drugs 2021, 35, 177–213. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A.; Rompre, P.P. Brain dopamine and reward. Annu. Rev. Psychol. 1989, 40, 191–225. [Google Scholar] [CrossRef]
- Teoh Bing Fei, J.; Yee, A.; Habil, M.H.; Danaee, M.J. Effectiveness of Methadone Maintenance Therapy and Improvement in Quality of Life Following a Decade of Implementation. Subst. Abuse Treat. 2016, 69, 50–56. [Google Scholar] [CrossRef]
- Coates, S.; Lazarus, P.J. Hydrocodone, Oxycodone, and Morphine Metabolism and Drug-Drug Interactions. Pharmacol. Exp. Ther. 2023, 387, 150–169. [Google Scholar] [CrossRef]
- Koob, G.F.; Ahmed, S.H.; Boutrel, B.; Chen, S.A.; Kenny, P.J.; Markou, A.; O’Dell, L.E.; Parsons, L.H.; Sanna, P.P. Neurobiological mechanisms in the transition from drug use to drug dependence. Neurosci. Biobehav. Rev. 2004, 27, 739–749. [Google Scholar] [CrossRef]
- Diaz, S.L.; Kemmling, A.; Balerio, G.N. Baclofen reestablishes striatal and cortical dopamine concentrations during naloxone-precipitated withdrawal. Neurochem. Int. 2003, 42, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Done, C.; Silverstone, P.; Sharp, T. Effect of naloxone-precipitated morphine withdrawal on noradrenaline release in rat hippocampus in vivo. Eur. J. Pharmacol. 1992, 215, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Robinson, T.E.; Berridge, K.C. The psychology and neurobiology of addiction: An incentive-sensitization view. Addiction 2000, 95, 91–117. [Google Scholar] [CrossRef]
- Guegan, T.; Cebrià, J.P.; Maldonado, R.; Martin, M. Morphine-induced locomotor sensitization produces structural plasticity in the mesocorticolimbic system dependent on CB1-R activity. Addict. Biol. 2016, 21, 1113–1126. [Google Scholar] [CrossRef]
- Liu, X.S.; Hou, Y.; Yan, T.L.; Guo, Y.Y.; Han, W.; Guan, F.L.; Chen, T.; Li, T. Dopamine D3 receptor-regulated NR2B subunits of N-methyl-d-aspartate receptors in the nucleus accumbens involves in morphine-induced locomotor activity. CNS Neurosci. Ther. 2014, 20, 823–829. [Google Scholar] [CrossRef]
- Fish, R.E.; Brown, M.J.; Danneman, P.J.; Karas, A.Z.; Langford, D.J.; Mogil, J.S. Anesthesia and analgesia in laboratory animals. ACLAM 2008, 2, 549–560. [Google Scholar]
- Listos, J.; Baranowska-Bosiacka, I.; Talarek, S.; Listos, P.; Orzelska, J.; Fidecka, S.; Gutowska, I.; Kolasa, A.; Rybicka, M.; Chlubek, D. The effect of perinatal lead exposure on dopamine receptor D2 expression in morphine dependent rats. Toxicology 2013, 310, 73–83. [Google Scholar] [CrossRef]
- Listos, J.; Baranowska-Bosiacka, I.; Wąsik, A.; Talarek, S.; Tarnowski, M.; Listos, P.; Łupina, M.; Antkiewicz-Michaluk, L.; Gutowska, I.; Tkacz, M.; et al. The adenosinergic system is involved in sensitization to morphine withdrawal signs in rats-neurochemical and molecular basis in dopaminergic system. Psychopharmacology 2016, 233, 2383–2397. [Google Scholar] [CrossRef]
- Kuribara, H. Induction of sensitization to hyperactivity caused by morphine in mice: Effects of post-drug environments. Pharmacol. Biochem. Behav. 1997, 57, 341–346. [Google Scholar] [CrossRef]
- Kotlińska, J.; Bocheński, M. Comparison of the effects of mGluR1 and mGluR5 antagonists on the expression of behavioral sensitization to the locomotor effect of morphine and the orphanine withdrawal jumping in mice. Eur. J. Pharmacol. 2007, 558, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Nestler, E.J. Reflections on: “A general role for adaptations in G-Proteins and the cyclic AMP system in mediating the chronic actions of morphine and cocaine on neuronal function”. Brain Res. 2016, 1645, 71–74. [Google Scholar] [CrossRef]
- Garzón, J.; Rodríguez-Muñoz, M.; Sánchez-Blázquez, P. Direct association of Mu-opioid and NMDA glutamate receptors supports their cross-regulation: Molecular implications for opioid tolerance. Curr. Drug Abuse Rev. 2012, 5, 199–226. [Google Scholar] [CrossRef] [PubMed]
- Commons, K.G.; van Bockstaele, E.J.; Pfaff, D.W. Frequent colocalization of mu opioid and NMDA-type glutamate receptors at postsynaptic sites in periaqueductal gray neurons. J. Com. Neurol. 1999, 408, 549–559. [Google Scholar] [CrossRef]
- Trujillo, K.A. The neurobiology of opiate tolerance, dependence and sensitization: Mechanisms of NMDA receptor-dependent synaptic plasticity. Neurotoxicol. Res. 2002, 4, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Joo, D.T. Subpopulation of dorsal horn neurons displays enhanced N-methyl-d-aspartate receptor function after chronic morphine exposure. Anesthesiology 2006, 104, 815–825. [Google Scholar] [CrossRef]
- Morgan, M.M.; Whittier, K.L.; Hegarty, D.M.; Aicher, S.A. Periaqueductal gray neurons project to spinally projecting GABAergic neurons in the rostral ventromedial medulla. Pain 2008, 140, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Medrano, M.C.; Mendiguren, A.; Pineda, J. Effect of ceftriaxone and topiramate treatments on naltrexone-precipitated morphine withdrawal and glutamate receptor desensitization in the rat locus coeruleus. Psychopharmacology 2015, 232, 2795–2809. [Google Scholar] [CrossRef]
- Hajhashemi, V.; Abed-Natanzi, M. Effect of five common anticonvulsant drugs on naloxone-precipitated morphine withdrawal in mice. Res. Pharm. Sci. 2011, 6, 57–62. [Google Scholar]
- Koob, G.F.; Stinus, L.; Le Moal, M.; Bloom, F.E. Opponent process theory of motivation: Neurobiological evidence from studies of opiate dependence. Neurosci. Biobehav. Rev. 1989, 13, 135–140. [Google Scholar] [CrossRef]
- Sepulveda, M.J.; Hernandez, L.; Rada, P.; Tucci, S.; Contreras, E. Effect of precipitated withdrawal on extracellular glutamate and aspartate in the nucleus accumbens of chronically morphine-treated rats: An in vivo microdialysis study. Pharmacol. Biochem. Behav. 1998, 60, 255–262. [Google Scholar] [CrossRef]
- Fox, M.E.; Rodeberg, N.T.; Wightman, R.M. Reciprocal catecholamine changes during opiate exposure and withdrawal. Neuropsychopharmacology 2017, 42, 671–681. [Google Scholar] [CrossRef]
- Liu, Q.F.; Li, L.; Guo, Y.Q.; Li, X.; Mou, Z.D.; Wang, X.; Du, G.Z. Injection of Toll-like receptor 4 siRNA into the ventrolateral periaqueductal gray attenuates withdrawal syndrome in morphine-dependent rats. Arch. Ital. Biol. 2016, 154, 133–142. [Google Scholar] [PubMed]
- Le Moal, M.; Simon, H. Mesocorticolimbic dopaminergic network: Functional and regulatory roles. Physiol. Rev. 1991, 71, 155–234. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. The neural basis of drug craving: An incentive-sensitization theory of addiction. Brain Res. Rev. 1993, 18, 247–291. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. The incentive sensitization theory of addiction: Some current issues. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 3137–3146. [Google Scholar] [CrossRef] [PubMed]
- Vanderschuren, L.J.; Kalivas, P.W. Alterations in dopaminergic and glutamatergic transmission in the induction and expression of behavioral sensitization: A critical review of preclinical studies. Psychopharmacology 2000, 151, 99–120. [Google Scholar] [CrossRef]
- Vanderschuren, L.J.; Pierce, R.C. Sensitization processes in drug addiction. Curr. Top. Behav. Neurosci. 2010, 3, 179–195. [Google Scholar]
- Wolf, M.E. LTP may trigger addiction. Mol. Interv. 2003, 3, 248–252. [Google Scholar] [CrossRef]
- Jeziorski, M.; White, F.J.; Wolf, M.E. MK-801 prevents the development of behavioral sensitization during repeated morphine administration. Synapse 1994, 16, 137–147. [Google Scholar] [CrossRef]
- Carlezon, W.A., Jr.; Rasmussen, K.; Nestler, E.J. AMPA antagonist LY293558 blocks the development, without blocking the expression, of behavioral sensitization to morphine. Synapse 1999, 31, 256–262. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pysiewicz, A.; Mazur, A.; Kotlińska, J.; Baranowska-Bosiacka, I.; Fronc, K.; Łupina, M.; Kruk-Słomka, M.; Listos, J. The Influence of Topiramate on Morphine Dependence in Mice. Biomolecules 2025, 15, 730. https://doi.org/10.3390/biom15050730
Pysiewicz A, Mazur A, Kotlińska J, Baranowska-Bosiacka I, Fronc K, Łupina M, Kruk-Słomka M, Listos J. The Influence of Topiramate on Morphine Dependence in Mice. Biomolecules. 2025; 15(5):730. https://doi.org/10.3390/biom15050730
Chicago/Turabian StylePysiewicz, Adrian, Antonina Mazur, Jolanta Kotlińska, Irena Baranowska-Bosiacka, Krzysztof Fronc, Małgorzata Łupina, Marta Kruk-Słomka, and Joanna Listos. 2025. "The Influence of Topiramate on Morphine Dependence in Mice" Biomolecules 15, no. 5: 730. https://doi.org/10.3390/biom15050730
APA StylePysiewicz, A., Mazur, A., Kotlińska, J., Baranowska-Bosiacka, I., Fronc, K., Łupina, M., Kruk-Słomka, M., & Listos, J. (2025). The Influence of Topiramate on Morphine Dependence in Mice. Biomolecules, 15(5), 730. https://doi.org/10.3390/biom15050730