
The Role of Substance P in Corneal Homeostasis

 , and
, and {kind=link}
Abstract
1. Introduction
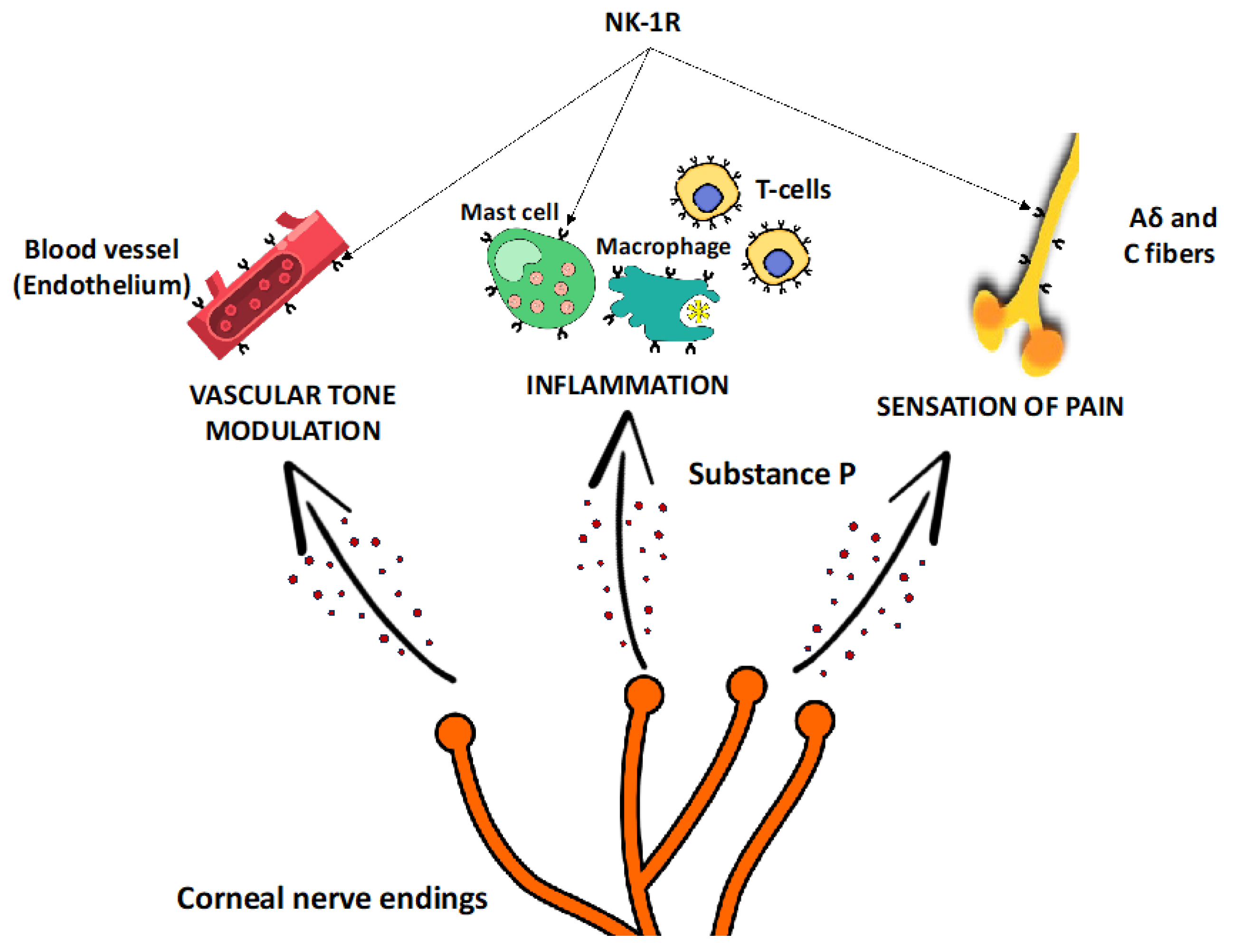
2. The Role of Substance P in Corneal Pain Perception
3. The Role of Substance P in Regulating Immune Response
4. The Role of Substance P in Corneal Neovascularization
5. Substance P and Interactions with Other Cytokines
6. The Role of SP in Epithelial Homeostasis
7. The Role of Substance P in Neurotrophic Keratitis
8. The Role of Substance P in Complications Due to Refractive Procedures
9. Interactions Between Dopamine, Substance P and Corneal Epithelium
10. Discussion
10.1. Corneal Innervation and Homeostasis
10.2. Pain Perception and Inflammatory Response
10.3. Immune Modulation and Inflammation
10.4. Corneal Neovascularization and Wound Healing
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- DelMonte, D.W.; Kim, T. Anatomy and physiology of the cornea. J. Cataract Refract. Surg. 2011, 37, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.J.; Marfurt, C.F.; Kruse, F.; Tervo, T.M.T. Corneal nerves: Structure, contents and function. Exp. Eye Res. 2003, 76, 521–542. [Google Scholar] [CrossRef] [PubMed]
- Sigelman, S.; Friedenwald, J.S. Mitotic and wound-healing activities of the corneal epithelium; effect of sensory denervation. A.M.A. Arch. Ophthalmol. 1954, 52, 46–57. [Google Scholar] [CrossRef]
- Versura, P.; Giannaccare, G.; Pellegrini, M.; Sebastiani, S.; Campos, E.C. Neurotrophic keratitis: Current challenges and future prospects. Eye Brain 2018, 10, 37–45. [Google Scholar] [CrossRef]
- Saad, S.; Abdelmassih, Y.; Saad, R.; Guindolet, D.; Khoury Sel Doan, S.; Cochereau, I.; Gabison, E.E. Neurotrophic keratitis: Frequency, etiologies, clinical management and outcomes. Ocul. Surf. 2020, 18, 231–236. [Google Scholar] [CrossRef]
- Rutledge, L.T. The effects of denervation and stimulation upon synaptic ultrastructure. J. Comp. Neurol. 1978, 178, 117–128. [Google Scholar] [CrossRef]
- Cavanagh, H.D.; Colley, A.M. The molecular basis of neurotrophic keratitis. Acta Ophthalmologica. Suppl. 1989, 192, 115–134. [Google Scholar] [CrossRef] [PubMed]
- v Euler, U.S.; Gaddum, J.H. An unidentified depressor substance in certain tissue extracts. J. Physiol. 1931, 72, 74–87. [Google Scholar] [CrossRef]
- Milner, P.; Bodin, P.; Guiducci, S.; Del Rosso, A.; Kahaleh, M.B.; Matucci-Cerinic, M.; Burnstock, G. Regulation of substance P mRNA expression in human dermal microvascular endothelial cells. Clin. Exp. Rheumatol. 2004, 22 (Suppl. S33), S24–S27. [Google Scholar]
- Lai, J.P.; Douglas, S.D.; Ho, W.Z. Human lymphocytes express substance P and its receptor. J. Neuroimmunol. 1998, 86, 80–86. [Google Scholar] [CrossRef]
- Marriott, I.; Bost, K.L. IL-4 and IFN-gamma up-regulate substance P receptor expression in murine peritoneal macrophages. J. Immunol. 2000, 165, 182–191. [Google Scholar] [CrossRef]
- Suvas, S. Role of Substance P Neuropeptide in Inflammation, Wound Healing, and Tissue Homeostasis. J. Immunol. 2017, 199, 1543–1552. [Google Scholar] [CrossRef]
- Takeda, Y.; Chou, K.B.; Takeda, J.; Sachais, B.S.; Krause, J.E. Molecular cloning, structural characterization and functional expression of the human substance P receptor. Biochem. Biophys. Res. Commun. 1991, 179, 1232–1240. [Google Scholar] [CrossRef]
- Caberlotto, L.; Hurd, Y.L.; Murdock, P.; Wahlin, J.P.; Melotto, S.; Corsi, M.; Carletti, R. Neurokinin 1 receptor and relative abundance of the short and long isoforms in the human brain. Eur. J. Neurosci. 2003, 17, 1736–1746. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.M.; Leeman, S.E.; Niall, H.D. Amino-acid sequence of substance P. Nat. New Biol. 1971, 232, 86–87. [Google Scholar] [CrossRef] [PubMed]
- Mashaghi, A.; Marmalidou, A.; Tehrani, M.; Grace, P.M.; Pothoulakis, C.; Dana, R. Neuropeptide substance P and the immune response. Cell. Mol. Life Sci. 2016, 73, 4249–4264. [Google Scholar] [CrossRef] [PubMed]
- De Koninck, Y.; Henry, J.L. Substance P-mediated slow excitatory postsynaptic potential elicited in dorsal horn neurons in vivo by noxious stimulation. Proc. Natl. Acad. Sci. USA 1991, 88, 11344–11348. [Google Scholar] [CrossRef]
- Yu, M.; Lee, S.M.; Lee, H.; Amouzegar, A.; Nakao, T.; Chen, Y.; Dana, R. Neurokinin-1 Receptor Antagonism Ameliorates Dry Eye Disease by Inhibiting Antigen-Presenting Cell Maturation and T Helper 17 Cell Activation. Am. J. Pathol. 2020, 190, 125–133. [Google Scholar] [CrossRef]
- Lasagni Vitar, R.M.; Bonelli, F.; Atay, A.; Triani, F.; Fonteyne, P.; Di Simone, E.; Rama, P.; Mondino, A.; Ferrari, G. Topical neurokinin-1 receptor antagonist Fosaprepitant ameliorates ocular graft-versus-host disease in a preclinical mouse model. Exp. Eye Res. 2021, 212, 108825. [Google Scholar] [CrossRef]
- Suanno, G.; Pederzolli, M.; Lasagni Vitar, R.M.; Fonteyne, P.; Ferrari, G. Topical fosaprepitant for the treatment of ocular pain and inflammation. Am. J. Ophthalmol. Case Rep. 2023, 32, 101964. [Google Scholar] [CrossRef]
- Pina, R.; Ugarte, G.; Campos, M.; Inigo-Portugues, A.; Olivares, E.; Orio, P.; Belmonte, C.; Bacigalupo, J.; Madrid, R. Role of trpm8 channels in altered cold sensitivity of corneal primary sensory neurons induced by axonal damage. J. Neurosci. 2019, 39, 8177–8192. [Google Scholar] [CrossRef] [PubMed]
- Marfurt, C.F.; Cox, J.; Deek, S.; Dvorscak, L. Anatomy of the human corneal innervation. Exp. Eye Res. 2010, 90, 478–492. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Masuko, S. Peripheral and central distribution of TRPV1, substance P and CGRP of rat corneal neurons. Brain Res. 2006, 1085, 87–94. [Google Scholar] [CrossRef]
- González-González, O.; Bech, F.; Gallar, J.; Merayo-Lloves, J.; Belmonte, C. Functional properties of sensory nerve terminals of the mouse cornea. Investig. Ophthalmol. Vis. Sci. 2017, 58, 404–415. [Google Scholar] [CrossRef]
- Guzmán, M.; Miglio, M.S.; Zgajnar, N.R.; Colado, A.; Almejún, M.B.; Keitelman, I.A.; Sabbione, F.; Fuentes, F.; Trevani, A.S.; Giordano, M.N.; et al. The mucosal surfaces of both eyes are immunologically linked by a neurogenic inflammatory reflex involving TRPV1 and substance P. Mucosal Immunol. 2018, 11, 1441–1453. [Google Scholar] [CrossRef]
- Sumioka, T.; Okada, Y.; Reinach, P.S.; Shirai, K.; Miyajima, M.; Yamanaka, O.; Saika, S. Impairment of corneal epithelial wound healing in a TRPV1-deficient mouse. Investig. Ophthalmol. Vis. Sci. 2014, 55, 3295–3302. [Google Scholar] [CrossRef]
- Chang, C.T.; Jiang, B.Y.; Chen, C.C. Ion Channels Involved in Substance P-Mediated Nociception and Antinociception. Int. J. Mol. Sci. 2019, 20, 1596. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.C.J.; Chen, W.N.; Chen, C.J.; Lin, Y.W.; Zimmer, A.; Chen, C.C. An antinociceptive role for substance P in acid-induced chronic muscle pain. Proc. Natl. Acad. Sci. USA 2012, 109, E76–E83. [Google Scholar] [CrossRef]
- Lorton, D.; Bellinger, D.L.; Felten, S.Y.; Felten, D.L. Substance P innervation of the rat thymus. Peptides 1990, 11, 1269–1275. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Germonpré, P.R.; Germonpré, G.; Everaert, E.G.; Carro-Muino, I.; De Veerman, M.; De Felipe, C.; Hunt, S.P.; Thielemans, K.; Joos, G.F.; et al. Endogenously produced substance P contributes to lymphocyte proliferation induced by dendritic cells and direct TCR ligation. Eur. J. Immunol. 1999, 29, 3815–3825. [Google Scholar] [CrossRef]
- Raap, M.; Rüdrich, U.; Ständer, S.; Gehring, M.; Kapp, A.; Raap, U. Substance P activates human eosinophils. Exp. Dermatol. 2015, 24, 557–559. [Google Scholar] [CrossRef]
- Wozniak, A.; McLennan, G.; Betts, W.H.; Murphy, G.A.; Scicchitano, R. Activation of human neutrophils by substance P: Effect on FMLP-stimulated oxidative and arachidonic acid metabolism and on antibody-dependent cell-mediated cytotoxicity. Immunology 1989, 68, 359. [Google Scholar] [PubMed]
- Lighvani, S.; Huang, X.; Trivedi, P.P.; Swanborg, R.H.; Hazlett, L.D. Substance P regulates natural killer cell interferon-gamma production and resistance to Pseudomonas aeruginosa infection. Eur. J. Immunol. 2005, 35, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Calvo, C.; Chavanel, G.; Senik, A. Substance P enhances IL-2 expression in activated human T cells. J. Immunol. 1992, 148, 3498–3504. [Google Scholar] [CrossRef] [PubMed]
- Bost, K.L. Tachykinin-modulated anti-viral responses. Front. Biosci. A J. Virtual Libr. 2004, 9, 1994–1998. [Google Scholar] [CrossRef]
- Weinstock, J.V. The role of substance P, hemokinin and their receptor in governing mucosal inflammation and granulomatous responses. Front. Biosci. A J. Virtual Libr. 2004, 9, 1936–1943. [Google Scholar] [CrossRef]
- Rameshwar, P.; Gascón, P. Substance P (SP) Mediates Production of Stem Cell Factor and Interleukin-1 in Bone Marrow Stroma: Potential Autoregulatory Role for These Cytokines in SP Receptor Expression and Induction. Blood 1995, 86, 482–490. [Google Scholar] [CrossRef]
- Mei, G.; Xia, L.; Zhou, J.; Zhang, Y.; Tuo, Y.; Fu, S.; Zou, Z.; Wang, Z.; Jin, D. Neuropeptide SP activates the WNT signal transduction pathway and enhances the proliferation of bone marrow stromal stem cells. Cell Biol. Int. 2013, 37, 1225–1232. [Google Scholar] [CrossRef]
- Guo, C.J.; Lai, J.P.; Luo, H.M.; Douglas, S.D.; Ho, W.Z. Substance P up-regulates macrophage inflammatory protein-1β expression in human T lymphocytes. J. Neuroimmunol. 2002, 131, 160–167. [Google Scholar] [CrossRef]
- Mathers, A.R.; Tckacheva, O.A.; Janelsins, B.M.; Shufesky, W.J.; Morelli, A.E.; Larregina, A.T. In vivo signaling through the neurokinin 1 receptor favors transgene expression by Langerhans cells and promotes the generation of Th1- and Tc1-biased immune responses. J. Immunol. 2007, 178, 7006–7017. [Google Scholar] [CrossRef]
- El-Shazly, A.E.; Masuyama, K.; Eura, M.; Ishikawa, T. Immunoregulatory effect of substance P in human eosinophil migratory function. Immunol. Investig. 1996, 25, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shavit, Z.; Goldman, R.; Stabinsky, Y.; Gottlieb, P.; Fridkin, M.; Teichberg, V.I.; Blumberg, S. Enhancement of phagocytosis—A newly found activity of substance P residing in its N-terminal tetrapeptide sequence. Biochem. Biophys. Res. Commun. 1980, 94, 1445–1451. [Google Scholar] [CrossRef]
- Repke, H.; Bienert, M. Structural requirements for mast cell triggering by substance P-like peptides. Agents Actions 1988, 23, 207–210. [Google Scholar] [CrossRef]
- Ziche, M.; Morbidelli, L.; Masini, E.; Amerini, S.; Granger, H.J.; Maggi, C.A.; Geppetti, P.; Ledda, F. Nitric oxide mediates angiogenesis in vivo and endothelial cell growth and migration in vitro promoted by substance P. J. Clin. Investig. 1994, 94, 2036–2044. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Coveñas, R.; Kramer, M. The involvement of the substance P/neurokinin 1 receptor system in viral infection: Focus on the gp120 fusion protein and homologous dipeptide domains. Acta Virol. 2019, 63, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Twardy, B.S.; Channappanavar, R.; Suvas, S. Substance P in the corneal stroma regulates the severity of herpetic stromal keratitis lesions. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8604–8613. [Google Scholar] [CrossRef]
- Bignami, F.; Rama, P.; Ferrari, G. Substance P and its Inhibition in Ocular Inflammation. Curr. Drug Targets 2016, 17, 1265–1274. [Google Scholar] [CrossRef]
- Chui, J.; Di Girolamo, N.; Coroneo, M.T.; Wakefield, D. The role of substance P in the pathogenesis of pterygia. Investig. Ophthalmol. Vis. Sci. 2007, 48, 4482–4489. [Google Scholar] [CrossRef]
- Fujishima, H.; Takeyama, M.; Takeuchi, T.; Saito, I.; Tsubota, K. Elevated levels of substance P in tears of patients with allergic conjunctivitis and vernal keratoconjunctivitis. Clin. Exp. Allergy 1997, 27, 372–378. [Google Scholar] [CrossRef]
- Tamura, T. Olopatadine ophthalmic solution suppresses substance P release in the conjunctivitis models. Asia Pac. Allergy 2012, 2, 115–121. [Google Scholar] [CrossRef]
- Ziche, M.; Morbidelli, L.; Pacini, M.; Geppetti, P.; Alessandri, G.; Maggi, C.A. Substance P stimulates neovascularization in vivo and proliferation of cultured endothelial cells. Microvasc. Res. 1990, 40, 264–278. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, G.; Hajrasouliha, A.R.; Sadrai, Z.; Ueno, H.; Chauhan, S.K.; Dana, R. Nerves and neovessels inhibit each other in the cornea. Investig. Ophthalmol. Vis. Sci. 2013, 54, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Kohara, H.; Tajima, S.; Yamamoto, M.; Tabata, Y. Angiogenesis induced by controlled release of neuropeptide substance P. Biomaterials 2010, 31, 8617–8625. [Google Scholar] [CrossRef]
- Hay, C.W.; Shanley, L.; Davidson, S.; Cowie, P.; Lear, M.; McGuffin, P.; Riedel, G.; McEwan, I.J.; MacKenzie, A. Functional effects of polymorphisms on glucocorticoid receptor modulation of human anxiogenic substance-P gene promoter activity in primary amygdala neurones. Psychoneuroendocrinology 2014, 47, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Garrett, N.E.; Cruwys, S.C.; LKidd, B.; Tomlinson, D.R. Effect of capsaicin on substance P and nerve growth factor in adjuvant arthritic rats. Neurosci. Lett. 1997, 230, 5–8. [Google Scholar] [CrossRef]
- Bignami, F.; Lorusso, A.; Rama, P.; Ferrari, G. Growth inhibition of formed corneal neovascularization following Fosaprepitant treatment. Acta Ophthalmol. 2017, 95, e641–e648. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Hong, H.S.; Kim, J.C.; Shin, J.S.; Son, Y. Inhibitory effect of rapamycin on corneal neovascularization in vitro and in vivo. Investig. Ophthalmol. Vis. Sci. 2005, 46, 454–460. [Google Scholar] [CrossRef]
- Sloniecka, M.; Roux, S.L.E.; Zhou, Q.; Danielson, P. Substance P Enhances Keratocyte Migration and Neutrophil Recruitment through Interleukin-8. Mol. Pharmacol. 2016, 89, 215–225. [Google Scholar] [CrossRef]
- Blanco-Vázquez, M.; Vázquez, A.; Fernández, I.; Novo-Diez, A.; Martínez-Plaza, E.; García-Vázquez, C.; González-García, M.J.; Sobas, E.M.; Calonge, M.; Enríquez-de-Salamanca, A. Inflammation-related molecules in tears of patients with chronic ocular pain and dry eye disease. Exp. Eye Res. 2022, 219, 109057. [Google Scholar] [CrossRef]
- Xu, Q.; Fitzsimmons, B.; Steinauer, J.; O’Neill, A.; Newton, A.C.; Hua, X.Y.; Yaksh, T.L. Spinal phosphinositide 3-kinase-Akt-mammalian target of rapamycin signaling cascades in inflammation-induced hyperalgesia. J. Neurosci. 2011, 31, 2113–2124. [Google Scholar] [CrossRef]
- Kovács, I.; Ludány, A.; Koszegi, T.; Fehér, J.; Kovács, B.; Szolcsányi, J.; Pintér, E. Substance P released from sensory nerve endings influences tear secretion and goblet cell function in the rat. Neuropeptides 2005, 39, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Yamada, M.; Ogata, M.; Kawai, M.; Mashima, Y.; Nishida, T. Substance P in human tears. Cornea 2003, 22, S48–S54. [Google Scholar] [CrossRef] [PubMed]
- Gaddipati, S.; Rao, P.; Jerome, A.D.; Burugula, B.B.; Gerard, N.P.; Suvas, S. Loss of Neurokinin-1 Receptor Alters Ocular Surface Homeostasis and Promotes an Early Development of Herpes Stromal Keratitis. J. Immunol. 2016, 197, 4021–4033. [Google Scholar] [CrossRef]
- Ko, J.A.; Murata, S.; Nishida, T. Up-regulation of the tight-junction protein ZO-1 by substance P and IGF-1 in A431 cells. Cell Biochem. Funct. 2009, 27, 388–394. [Google Scholar] [CrossRef]
- Yamada, N.; Yanai, R.; Inui, M.; Nishida, T. Sensitizing effect of substance P on corneal epithelial migration induced by IGF-1, fibronectin, or interleukin-6. Investig. Ophthalmol. Vis. Sci. 2005, 46, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Di, G.; Qi, X.; Qu, M.; Wang, Y.; Duan, H.; Danielson, P.; Xie, L.; Zhou, Q. Substance P promotes diabetic corneal epithelial wound healing through molecular mechanisms mediated via the neurokinin-1 receptor. Diabetes 2014, 63, 4262–4274. [Google Scholar] [CrossRef]
- Hong, H.S.; Lee, J.; Lee, E.; Kwon, Y.S.; Lee, E.; Ahn, W.; Jiang, M.H.; Kim, J.C.; Son, Y. A new role of substance P as an injury-inducible messenger for mobilization of CD29(+) stromal-like cells. Nat. Med. 2009, 15, 425–435. [Google Scholar] [CrossRef]
- Nishida, T. Neurotrophic mediators and corneal wound healing. Ocul. Surf. 2005, 3, 194–202. [Google Scholar] [CrossRef]
- Chikama, T.I.; Fukuda, K.; Morishige, N.; Nishida, T. Treatment of neurotrophic keratopathy with substance-P-derived peptide (FGLM) and insulin-like growth factor I. Lancet 1998, 351, 1783–1784. [Google Scholar] [CrossRef]
- Nakamura, M.; Ofuji, K.; Chikama, T.I.; Nishida, T. Combined effects of substance P and insulin-like growth factor-1 on corneal epithelial wound closure of rabbit in vivo. Curr. Eye Res. 1997, 16, 275–278. [Google Scholar] [CrossRef]
- Nagano, T.; Nakamura, M.; Nakata, K.; Yamaguchi, T.; Takase, K.; Okahara, A.; Ikuse, T.; Nishida, T. Effects of substance P and IGF-1 in corneal epithelial barrier function and wound healing in a rat model of neurotrophic keratopathy. Investig. Ophthalmol. Vis. Sci. 2003, 44, 3810–3815. [Google Scholar] [CrossRef] [PubMed]
- Guyon, A.; Roques, B.P.; Guyon, F.; Foucault, A.; Perdrisot, R.; Swerts, J.P.; Schwartz, J.C. Enkephalin degradation in mouse brain studied by a new H.P.L.C. method: Further evidence for the involvement of carboxydipeptidase. Life Sci. 1979, 25, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Chikama, T.I.; Nishida, T. Synergistic effect with Phe-Gly-Leu-Met-NH2 of the C-terminal of substance P and insulin-like growth factor-1 on epithelial wound healing of rabbit cornea. Br. J. Pharmacol. 1999, 127, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Sui, W.; Li, Y.; Qi, X.; Wang, Y.; Zhou, Q.; Gao, H. Substance P Inhibits Hyperosmotic Stress-Induced Apoptosis in Corneal Epithelial Cells through the Mechanism of Akt Activation and Reactive Oxygen Species Scavenging via the Neurokinin-1 Receptor. PLoS ONE 2016, 11, e0149865. [Google Scholar] [CrossRef]
- Sacchetti, M.; Lambiase, A. Diagnosis and management of neurotrophic keratitis. Clin. Ophthalmol. 2014, 8, 571. [Google Scholar] [CrossRef]
- Puca, A.; Meglio, M.; Vari, R.; Tamburrini, G.; Tancredi, A. Evaluation of fifth nerve dysfunction in 136 patients with middle and posterior cranial fossae tumors. Eur. Neurol. 1995, 35, 33–37. [Google Scholar] [CrossRef]
- Hyndiuk, R.A.; Kazarian, E.L.; Schultz, R.O.; Seideman, S. Neurotrophic corneal ulcers in diabetes mellitus. Arch. Ophthalmol. 1977, 95, 2193–2196. [Google Scholar] [CrossRef]
- Nakamura, M.; Nishida, T.; Ofuji, K.; Reid, T.W.; Mannis, M.J.; Murphy, C.J. Synergistic effect of substance P with epidermal growth factor on epithelial migration in rabbit cornea. Exp. Eye Res. 1997, 65, 321–329. [Google Scholar] [CrossRef]
- Nakamura, M.; Chikama, T.I.; Nishida, T. Up-regulation of integrin α5 expression by combination of substance P and insulin-like growth factor-1 in rabbit corneal epithelial cells. Biochem. Biophys. Res. Commun. 1998, 246, 777–782. [Google Scholar] [CrossRef]
- Ofuji, K.; Nakamura, M.; Nishida, T. Signaling regulation for synergistic effects of substance P and insulin-like growth factor-1 or epidermal growth factor on corneal epithelial migration. Jpn. J. Ophthalmol. 2000, 44, 1–8. [Google Scholar] [CrossRef]
- Ghiasi, Z.; Gray, T.; Tran, P.; Dubielzig, R.; Murphy, C.; McCartney, D.L.; Reid, T.W. The Effect of Topical Substance-P Plus Insulin-like Growth Factor-1 (IGF-1) on Epithelial Healing After Photorefractive Keratectomy in Rabbits. Transl. Vis. Sci. Technol. 2018, 7, 12. [Google Scholar] [CrossRef] [PubMed]
- Bonini, S.; Lambiase, A.; Rama, P.; Caprioglio, G.; Aloe, L. Topical treatment with nerve growth factor for neurotrophic keratitis. Ophthalmology 2000, 107, 1347–1351. [Google Scholar] [CrossRef]
- Lambiase, A.; Rama, P.; Bonini, S.; Caprioglio, G.; Aloe, L. Topical treatment with nerve growth factor for corneal neurotrophic ulcers. N. Engl. J. Med. 1998, 338, 372–373. [Google Scholar] [CrossRef] [PubMed]
- Donnerer, J.; Amann, R.; Schuligoi, R.; Skofitsch, G. Complete recovery by nerve growth factor of neuropeptide content and function in capsaicin-impaired sensory neurons. Brain Res. 1996, 741, 103–108. [Google Scholar] [CrossRef]
- Schicho, R.; Skofitsch, G.; Donnerer, J. Regenerative effect of human recombinant NGF on capsaicin-lesioned sensory neurons in the adult rat. Brain Res. 1999, 815, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Bonini, S.; Lambiase, A.; Rama, P.; Sinigaglia, F.; Allegretti, M.; Chao, W.; Mantelli, F.; Bonini, S.; Messmer, E.; Aragona, P.; et al. Phase II Randomized, Double-Masked, Vehicle-Controlled Trial of Recombinant Human Nerve Growth Factor for Neurotrophic Keratitis. Ophthalmology 2018, 125, 1332–1343. [Google Scholar] [CrossRef]
- Wirta, D.; Lipsky, W.; Toyos, M.; Martel, J.; Goosey, J.; Verachtert, A.; El-Harazi, S.; Karpecki, P.; Allegretti, M.; Goisis, G.; et al. Recombinant human nerve growth factor (cenegermin) for moderate-to-severe dry eye: Phase II, randomized, vehicle-controlled, dose-ranging trial. BMC Ophthalmol. 2024, 24, 290. [Google Scholar] [CrossRef]
- Singh, R.B.; Naderi, A.; Cho, W.; Ortiz, G.; Musayeva, A.; Dohlman, T.H.; Chen, Y.; Ferrari, G.; Dana, R. Modulating the tachykinin: Role of substance P and neurokinin receptor expression in ocular surface disorders. Ocul. Surf. 2022, 25, 142. [Google Scholar] [CrossRef]
- Sharma, B.; Soni, D.; Saxena, H.; Stevenson, L.J.; Karkhur, S.; Takkar, B.; Vajpayee, R.B. Impact of corneal refractive surgery on the precorneal tear film. Indian J. Ophthalmol. 2020, 68, 2804–2812. [Google Scholar] [CrossRef]
- Chin, J.Y.; Lin, M.T.Y.; Lee, I.X.Y.; Mehta, J.S.; Liu, Y.C. Tear Neuromediator and Corneal Denervation Following SMILE. J. Refract. Surg. 2021, 37, 516–523. [Google Scholar] [CrossRef]
- Liu, Y.C.; Yam, G.H.F.; Lin, M.T.Y.; Teo, E.; Koh, S.K.; Deng, L.; Zhou, L.; Tong, L.; Mehta, J.S. Comparison of tear proteomic and neuromediator profiles changes between small incision lenticule extraction (SMILE) and femtosecond laser-assisted in-situ keratomileusis (LASIK). J. Adv. Res. 2021, 29, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.C.; Se Ji Jung, A.; Chin, J.Y.; Yang, L.W.Y.; Mehta, J.S. Cross-sectional Study on Corneal Denervation in Contralateral Eyes Following SMILE Versus LASIK. J. Refract. Surg. 2020, 36, 653–660. [Google Scholar] [CrossRef]
- Yang, L.; Mehta, J.; Liu, Y.C. Corneal neuromediator profiles following laser refractive surgery. Neural Regen. Res. 2021, 16, 2177–2183. [Google Scholar] [CrossRef]
- Zhao, L.; Zhang, Y.; Duan, H.; Yang, T.; Ma, B.; Zhou, Y.; Chen, J.; Chen, Y.; Qi, H. Clinical Characteristic and Tear Film Biomarkers After Myopic FS-LASIK: 1-Year Prospective Follow-up. J. Refract. Surg. 2024, 40, e508–e519. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.; Golebiowski, B.; Zhao, X.; Chen, S.; Zhou, S.; Stapleton, F. Long-term Effects of LASIK on Corneal Innervation and Tear Neuropeptides and the Associations With Dry Eye. J. Refract. Surg. 2016, 32, 518–524. [Google Scholar] [CrossRef]
- Grueb, M.; Wallenfels-Thilo, B.; Denk, O.; Mielke, J.; Reinthal, E.; Rohrbach, J.M.; Bartz-Schmidt, K.U. Monoamine receptors in human corneal epithelium and endothelium. Acta Ophthalmol. Scand. 2006, 84, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Grüb, M.; Mielke, J.; Rohrbach, M.; Schlote, T. Dopamine receptors of the corneal epithelium and endothelium. Klin. Monatsblatter Fur Augenheilkd. 2012, 229, 822–825. [Google Scholar] [CrossRef]
- Surmeier, D.J.; Ding, J.; Day, M.; Wang, Z.; Shen, W. D1 and D2 dopamine-receptor modulation of striatal glutamatergic signaling in striatal medium spiny neurons. Trends Neurosci. 2007, 30, 228–235. [Google Scholar] [CrossRef]
- Martel, J.C.; Gatti McArthur, S. Dopamine Receptor Subtypes, Physiology and Pharmacology: New Ligands and Concepts in Schizophrenia. Front. Pharmacol. 2020, 11, 1003. [Google Scholar] [CrossRef]
- Lee, S.P.; So, C.H.; Rashid, A.J.; Varghese, G.; Cheng, R.; Lança, A.J.; O’Dowd, B.F.; George, S.R. Dopamine D1 and D2 Receptor Co-activation Generates a Novel Phospholipase C-mediated Calcium Signal. J. Biol. Chem. 2004, 279, 35671–35678. [Google Scholar] [CrossRef]
- Vitar, R.M.L.; Barbariga, M.; Fonteyne, P.; Bignami, F.; Rama, P.; Ferrari, G. Modulating Ocular Surface Pain Through Neurokinin-1 Receptor Blockade. Investig. Ophthalmol. Vis. Sci. 2021, 62, 26. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Nakayasu, K.; Iwatsu, M.; Kanai, A. Endogenous substance P in corneal epithelial cells and keratocytes. Jpn. J. Ophthalmol. 2002, 46, 616–620. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jastrzębska-Miazga, I.; Machna, B.; Wyględowska-Promieńska, D.; Smędowski, A. The Role of Substance P in Corneal Homeostasis. Biomolecules 2025, 15, 729. https://doi.org/10.3390/biom15050729
Jastrzębska-Miazga I, Machna B, Wyględowska-Promieńska D, Smędowski A. The Role of Substance P in Corneal Homeostasis. Biomolecules. 2025; 15(5):729. https://doi.org/10.3390/biom15050729
Chicago/Turabian StyleJastrzębska-Miazga, Irmina, Bartosz Machna, Dorota Wyględowska-Promieńska, and Adrian Smędowski. 2025. "The Role of Substance P in Corneal Homeostasis" Biomolecules 15, no. 5: 729. https://doi.org/10.3390/biom15050729
APA StyleJastrzębska-Miazga, I., Machna, B., Wyględowska-Promieńska, D., & Smędowski, A. (2025). The Role of Substance P in Corneal Homeostasis. Biomolecules, 15(5), 729. https://doi.org/10.3390/biom15050729