1. Introduction
Cardiovascular disease (CVD) is a diagnosis of several heart and blood vessel diseases, including coronary artery disease, rhythm disturbances, valvular heart disease, carotid artery disease, congestive cardiomyopathy, and cerebrovascular accidents such as stroke [
1]. CVD remains the leading cause of death globally, and millions of lives are lost prematurely to heart disease each year [
2]. In Poland, CVD, including ischemic heart disease and stroke, is the leading cause of death, accounting for more than one-third of all deaths [
3]. Approximately 1.4 million Polish citizens have heart failure, and ischemic heart disease mortality was more than double compared to other European Union countries, with 131 deaths per 100,000 [
4].
CVD arises from a combination of factors, including socioeconomic status and access to healthcare [
5], psychosocial factors [
6], personal behaviors such as diet, physical activity, and smoking [
7,
8], environmental influences like air pollution [
9,
10], and genetic predisposition [
11]. These elements interact in a complex way, increasing the risk of developing heart disease and related conditions [
5]. Recent genetic studies have identified over 100 single-nucleotide polymorphisms (SNPs) associated with an increased risk of CVD [
12]. However, the exact mechanisms by which these SNPs influence the development and progression of CVD remain unclear [
13].
Since the early 20th century, vitamin D has been recognized for its pivotal role in the prevention and treatment of bone diseases such as rickets and osteomalacia [
14]. While its primary functions involve maintaining calcium and phosphate homeostasis, growing evidence suggests that vitamin D is intricately linked to cardiovascular health and disease [
15,
16]. This connection is rooted in the transcriptional activity of vitamin D, which influences the expression of approximately 3% of the human genome, including genes involved in cardiovascular regulation [
17].
Numerous observational studies have highlighted a significant association between vitamin D deficiency and various cardiovascular conditions [
18,
19]. These include arterial stiffness, hypertension, atherosclerosis, and left ventricular hypertrophy, all of which contribute to an increased risk of cardiovascular morbidity and mortality [
18,
20,
21]. Vitamin D is believed to exert its effects through multiple mechanisms, such as improving endothelial function, supporting cardiomyocyte health, and modulating the calcification processes in valves and vascular tissues [
22,
23,
24,
25]. Notably, the variability across studies and clinical trials investigating the role of vitamin D in cardiovascular health poses substantial challenges to reaching definitive conclusions and establishing universal recommendations [
15,
18,
19].
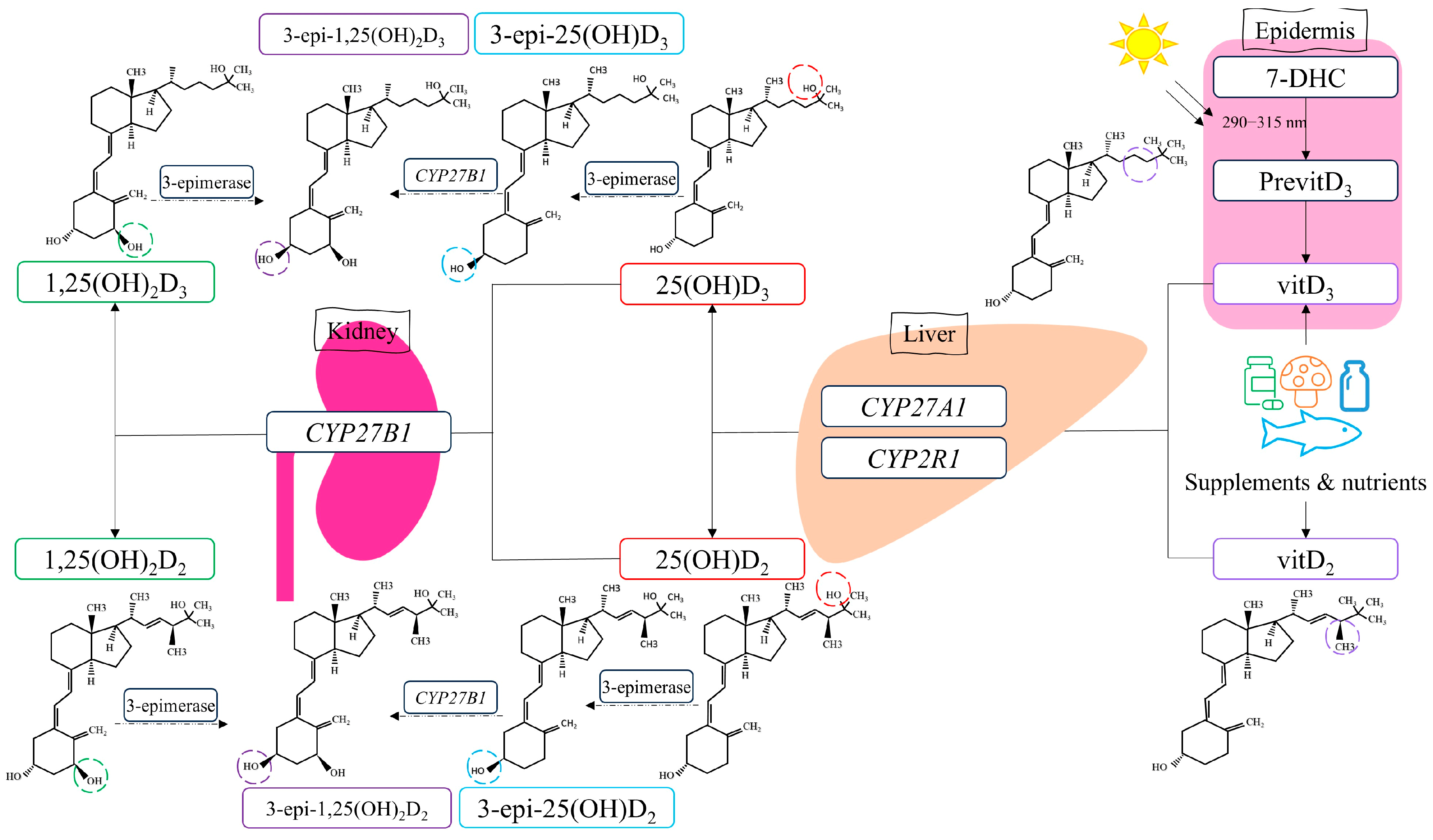
The mechanism of action for vitamin D involves binding the active metabolite, calcitriol, to the vitamin D receptor, which belongs to the steroid hormone family of nuclear receptors accountable for the transcriptional regulation of several hormone-responsive genes [
26]. However, calcitriol is synthesized through metabolic reactions involving several enzymes regulated by vitamin D metabolism genes such as
CYP2R1,
CYP27A1, and
CYP27B1 (
Figure 1).
Our previously published comprehensive review highlighted how the polymorphisms in these genes could influence several disorders [
27]. However, information on the association between
CYP2R1,
CYP27A1, and
CYP27B1 variants and CVD is limited. According to Wang et al., the presence of
CYP2R1-rs10741657 significantly increases the risk of coronary artery disease in men over 60 years old [
28]. This relationship was also observed in studies by Hassanein et al., where the GG genotype was associated with a 6.22-fold higher risk of coronary artery disease than the AA genotype [
29]. Studies in the Polish population, however, showed lower estimated glomerular filtration rate values in AA homozygotes for the
CYP27B1-rs10877012 polymorphism, indicating impaired kidney function and a risk factor for left ventricular heart failure [
30]. For
CYP27A1-rs4674344, carriers of the T allele were found to have a higher risk of developing metabolic syndrome due to disruptions in the levels of pro-inflammatory adipocytokines [
31].
The analyses conducted to date have not provided a conclusive understanding of the role of gene polymorphisms related to vitamin D metabolism in CVD, highlighting the need for further investigation. Our study aims to quantify vitamin D hydroxy metabolites (25(OH)D2 and 25(OH)D3, their epimers 3-epi-25(OH)D2 and 3-epi-25(OH)D3, and the active form 1,25(OH)2D3) in patients with CVD and healthy volunteers. We also identified polymorphisms in the CYP2R1, CYP27A1, and CYP27B1 genes and evaluated their impact on plasma vitamin D metabolite concentrations. To our knowledge, no similar studies have been conducted.
2. Materials and Methods
2.1. Study Population
The study involved 27 adult patients with cardiovascular diseases treated at the Department of Vascular Surgery, Endovascular Surgery, Angiology, and Phlebology of Poznan University of Medical Sciences in Poznań. Inclusion criteria were as follows: age over 18 years, male or female, diagnosis and/or treatment of CVD prior to admission. Exclusion criteria were as follows: supplementation with vitamin D, acute myocardial infarction, a positive history of cancer, impaired renal function determined by serum creatinine concentration >2 mg/dL, and current liver dysfunction.
The patients had venous thrombosis (n = 9), atherosclerosis of the lower extremities (n = 14), atrial fibrillation (n = 19), hypertension (n = 13), and coronary artery disease (n = 5). Other comorbidities were diabetes (n = 7) and respiratory conditions such as bronchial asthma and chronic obstructive pulmonary disease (n = 5). Patients were treated with rivaroxaban (n = 27), statins (n = 14), acetylsalicylic acid (n = 11), clopidogrel (n = 4); other medications were prescribed individually. Recruitment was ongoing throughout the year from December 2021 to January 2025.
The healthy volunteer group comprised 26 healthy adults recruited from the Department of Endocrinology, Metabolism, and Internal Diseases at the University Clinical Hospital in Poznań. Inclusion criteria were as follows: age over 18 years, male or female, and lack of thyroid disease confirmed by biochemical and imaging tests. Exclusion criteria were any diagnosed disease prior to admission, and poor health according to physical examination and laboratory analyses. Recruitment was from 2023 to 2024.
Blood samples were collected and stored at −80 °C until genetic analysis. For UPLC-MS/MS analysis, plasma was separated by centrifuging the blood samples at 1620 g for 10 min. The separated plasma was then stored at −80 °C until further analysis.
The study was planned and conducted according to the ethical principles of the Declaration of Helsinki [
32]. The study received approval from the Bioethics Committee of the Poznań University of Medical Sciences (approval numbers: 873/19, 58/20, 201/21, and 510/21).
2.2. Determining Concentrations of Vitamin D Hydroxy Metabolites in Plasma
We used our validated UPLC-MS/MS method to measure the concentrations of 25(OH)D
2, 25(OH)D
3, 3-epi-25(OH)D
2, and 3-epi-25(OH)D
3 [
33]. Briefly, 200 μL of patient plasma was mixed with 20 μL of methanol and 20 μL of the internal standard, D6-25(OH)D
3. The analytes were extracted using two 1000 μL portions of hexane. After evaporation, the residue was re-dissolved in 200 μL of a methanol–water (80:20,
v/
v) solution, and 10 μL was injected directly into a UPLC Nexera coupled with a triple quadrupole mass spectrometer LCMS-8030 (Shimadzu, Kyoto, Japan). The separation of the analytes was carried out using a Kinetex 2.6 μm F5 analytical column (50 mm × 2.1 mm) (Phenomenex, Torrance, CA, USA) with a mobile phase of methanol–water (80:20,
v/
v) containing 0.1% formic acid. The MS detection was performed in positive electrospray ionization mode. The specific
m/
z transitions for the analytes and chromatograms were provided as the
Supplementary Material (Table S1, Figure S1). The assay was linear from 2 to 100 ng mL⁻
1 (r
2 ≥ 0.995). The LOD and LOQ were 1 ng/mL and 2 ng/mL, respectively. The recovery of the analytes from plasma samples ranged from 70.1% to 97.1%. The precision, both inter- and intra-day, varied between 2.1% and 18.1%, and the accuracy of the method expressed by the relative error was between 0.9% and 14.7%.
Determination of 1,25(OH)2D3 was conducted by ELISA kit [DHVD3 (1,25-Dihydroxyvitamin D3) Competitive ELISA (St. John’s Laboratory, London, UK)] in accordance with the procedure described by the manufacturer. The kit has an identification range of 15.625–1000 pg/mL.
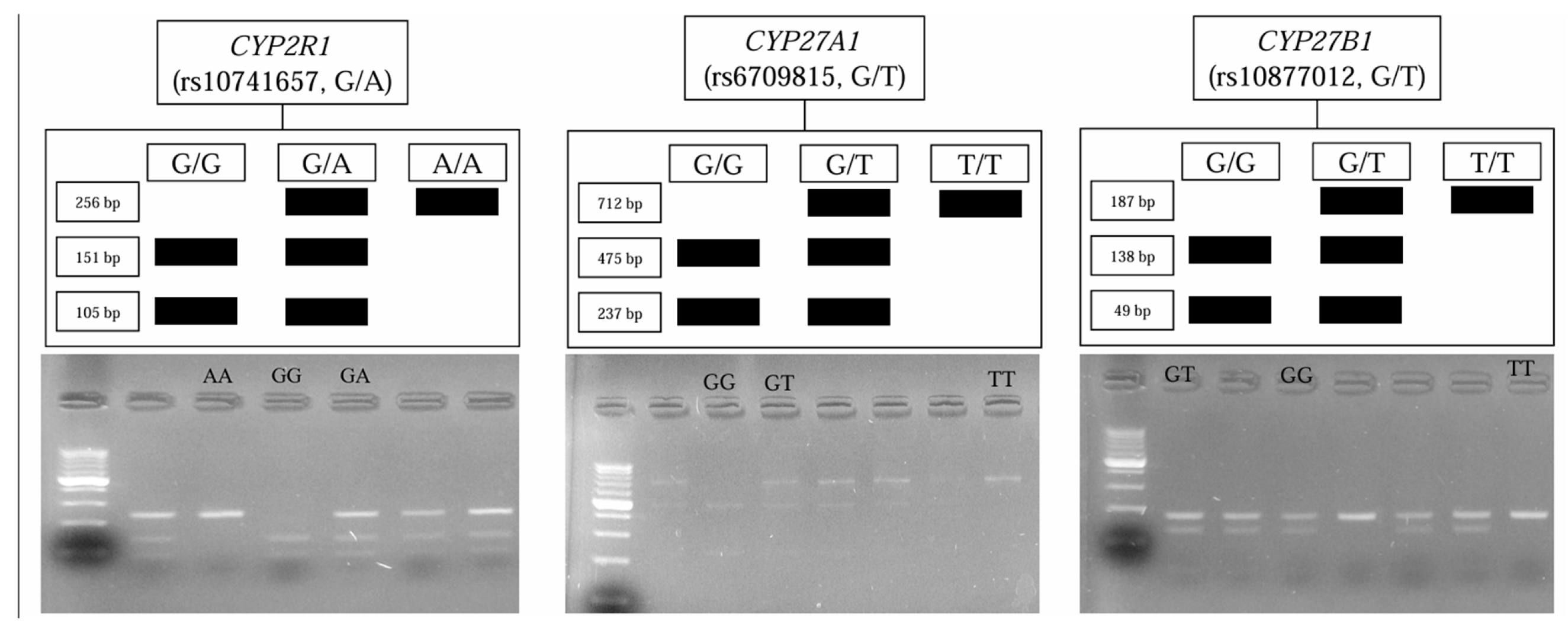
2.3. Genetic Analyses
The isolation of the genomic DNA from 200 μL of blood was performed using a commercially available kit according to the manufacturer’s instructions (GenMATRIX Quick Blood Purification Kit, EURx, Poland). To identify
CYP2R1,
CYP27A1, and
CYP27B1 polymorphisms, polymerase chain reaction (PCR) coupled with restriction fragment length analysis (RFLP) was performed. Primers were used, as reported by Stjepanovic et al. [
34], Giray [
35], and Latacz et al. [
36], respectively. Their sequences were checked by the Primer3Plus software version: 3.3.0 [
37]. The primers’ sequences, their annealing temperatures, the length of the products obtained, and the results of restriction are presented in the
Supplementary Material (Table S2).
2.4. Statistical Analysis
Excel (Microsoft Corp., Redmond, WA, USA) and Statistica 13.3 (StatSoft Inc., Tulsa, OK, USA) were used for statistical analysis. The Shapiro-Wilk test was applied to assess the normality of the variables. Normally distributed data are presented as mean ± SD, while non-normally distributed data are reported as median and interquartile range. A Student’s t-test was used for two groups with normal distribution, and ANOVA for three groups to compare the genotypes of CYP2R1, CYP27A1, and CYP27B1. For non-normal distributions, the Mann-Whitney U test and Kruskal-Wallis ANOVA were applied. The χ2 test assessed allele frequencies’ adherence to Hardy–Weinberg equilibrium, with a critical value of 3.84 (for n = 1 and α = 0.05). We also performed a logistic regression for the predictors of CVD and reported the results as odds ratios (ORs) and 95% confidence intervals (CIs). We considered p-value < 0.05 statistically significant for all tests.
4. Discussion
In the present study, we analyzed the
CYP2R1,
CYP27A1, and
CYP27B1 genetic polymorphisms and their association with anthropometric parameters, concentrations of 25(OH)D
2, 25(OH)D
3, 3-epi-25(OH)D
2, 3-epi-25(OH)D
3, and 1,25(OH)
2D
3, in Polish patients with cardiovascular disease and healthy subjects. To quantify vitamin D metabolites, we used the validated UPLC-MS/MS method, which is considered the gold standard technique for analyzing vitamin D status. This method is suitable for the simultaneous detection of multiple vitamin D metabolites and is not subject to matrix interference, unlike immunoassays [
38]. We found that the genetic distributions of
CYP2R1,
CYP27A1, and
CYP27B1 genotypes aligned with Hardy–Weinberg equilibrium, and allele frequencies were generally similar to the general Polish population, though the TT genotype was significantly more common in thrombosis patients (40%) than in healthy individuals (13.6%). No significant associations were observed between the
rs10741657 CYP2R1 polymorphism and anthropometric or vitamin D metabolite levels. For the
rs6709815 CYP27A1 polymorphism, no significant differences were seen in patients, but in healthy subjects, 3-epi-25(OH)D
3 percentages varied by genotype. Analysis of the
rs10877012 CYP27B1 polymorphism revealed significant genotype-based differences in vitamin D metabolite concentrations, with GT individuals showing the highest 25(OH)D
3 and calcitriol levels, while in TT individuals the values were the lowest. In the healthy volunteers, the TT genotype was associated with significantly higher 25(OH)D
2 concentrations and a trend toward higher calcitriol levels.
In regard to 25(OH)D
3 levels, in both groups, the majority of individuals had concentrations below the optimal value of 30 ng/mL, with the average value being numerically lower in patients with CVD (20.4 ng/mL) than in healthy individuals (25.5 ng/mL). Vitamin D insufficiency (25(OH)D
3 concentration < 20 ng/mL) was found in 44.4% of patients and 34.6% of healthy volunteers, indicating the prevalence of vitamin D deficiency in the analyzed population. Oberoi et al. also demonstrated differences in calcidiol levels between individuals with CVD and healthy individuals, whose study among Indian residents showed vitamin D deficiency (using a different classification, serum level < 30 ng/mL) in 63% of sick individuals and 35% of healthy controls [
39]. Lower concentrations in the study group compared to the healthy volunteers’ group were also observed for epimeric forms of vitamin D. The average 3-epi-25(OH)D
3 concentration was 2.4 ng/mL in the study group and 3.7 ng/mL in the control group, while for 3-epi-25(OH)D
2, the average concentration in the study group was below the lower quantification limit of the method (<2.0 ng/mL), and in the group of healthy volunteers, it was 5.1 ng/mL. Meanwhile, 25(OH)D
2 concentrations were higher in patients with CVD (8.1 ng/mL) compared to healthy individuals (6.5 ng/mL). Differences in 1,25(OH)
2D
3 concentrations were also noted between the two groups, with patients having an average calcitriol concentration of 43.4 pg/mL, compared to 67.9 pg/mL in healthy individuals. However, none of the differences in metabolite concentrations were statistically significant (
Table 1).
For the
rs10741657 CYP2R1 polymorphism, both groups’ genotype and allele frequencies were consistent with Hardy–Weinberg equilibrium. A comparison of genotype frequencies between the study group and the healthy volunteers group showed no statistically significant differences, so there is no basis to suggest an association between the
rs10741657 polymorphism and CVD in the analyzed population. In a study involving German patients with type 2 diabetes and healthy controls, Klahold et al. [
40] also reported that allele frequencies of
rs10741657 were in Hardy–Weinberg equilibrium. However, they noticed that the “A” allele of
CYP2R1 rs10741657 was more frequent among patients (42.1% vs. 36.3%; OR: 1.28; CI: 1.07–1.53;
p = 0.016). Our study did not observe such a relationship, even though it was numerically higher in the patient group than in healthy volunteers (60% vs. 40%; OR: 2.33; CI: 0.78–7.09;
p = 0.132).
Vitamin D metabolite concentrations did not differ significantly based on genotype, either among patients or healthy individuals (
Table 3 and
Table 4). In patients, the lowest 25(OH)D
3 concentration was found in AA homozygotes, with an average of 15.02 ng/mL, while the highest average concentration was observed in heterozygotes (22.4 ng/mL). Similar results were obtained in individuals without CVD, where the lowest average 25(OH)D
3 concentration occurred in volunteers with the AA genotype (21.3 ng/mL) and the highest in GA heterozygotes (26.4 ng/mL). Therefore, this study could not confirm the findings of Duan et al. in the European population [
41] or Hassanein et al. in the Egyptian population [
29], which indicated that the GG genotype is associated with lower 25(OH)D levels. The discrepancy may be due to the small sample size or ethnic differences between populations.
For the
rs6709815 CYP27A1 polymorphism, genotype and allele frequencies among patients and healthy individuals remained in Hardy–Weinberg equilibrium. In the group of individuals with CVD, the T allele was less frequent (T = 0.620) compared to the values defined for the Turkish patients with multiple sclerosis (T = 0.816) [
35], while this allele in our results was less frequent among healthy individuals (T = 0.432). Furthermore, significant differences were observed in the frequency of the
TT genotype between the analyzed groups (
p = 0.044). TT homozygotes were over three times more frequent among individuals with thrombosis than in healthy individuals, as shown in
Table 2. To date, there have been no scientific reports on the
rs6709815 polymorphism as a risk factor for CVD. However, the results of this study indicate a potential association between this polymorphism and CVD.
No significant differences in vitamin D metabolite concentrations were observed in the study group based on genotype. In contrast, in the group of healthy volunteers, significant differences were found in the percentage of the epimeric form of 25-hydroxyvitamin D
3 in the total 25(OH)D
3 pool (
p = 0.0289). The lowest percentage of 3-epi-25(OH)D
3, amounting to 10.3%, was found in individuals with the
CYP7A1-GT genotype, while for the GG and TT genotypes, the percentages were 22.0% and 20.0%, respectively (
Table 6). To date, no studies have demonstrated a relationship between the
rs6709815 polymorphism and changes in vitamin D metabolite concentrations in the body [
42].
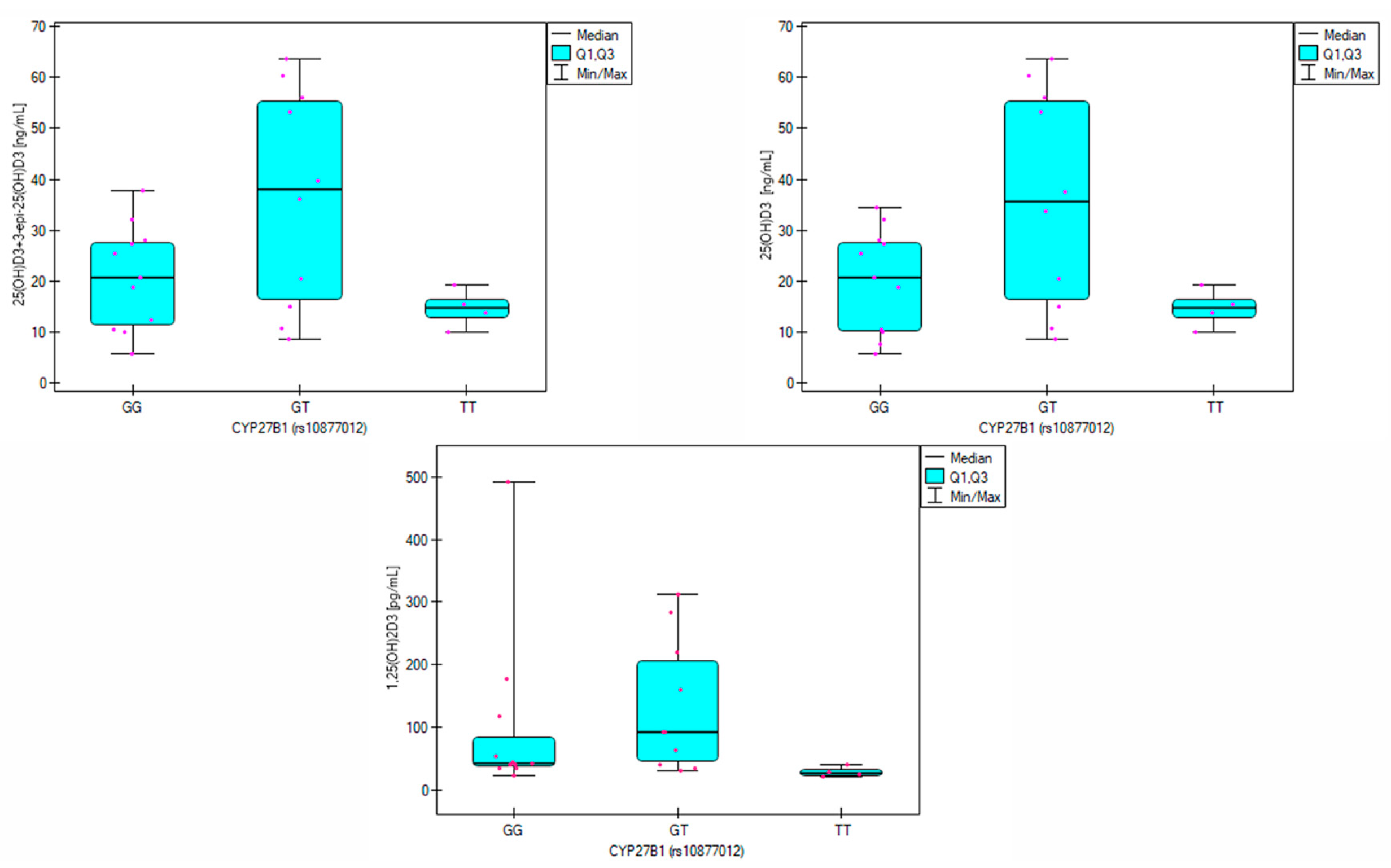
The frequency analysis of the
rs10877012 CYP27B1 polymorphism showed that the gene variation remained in Hardy–Weinberg equilibrium. Allele frequencies in both analyzed groups were similar to those observed in the pan-European population [
43]. The genotypes occurred with similar frequencies in both the study group and the group of healthy volunteers (
Table 2), so no association between the
rs10877012 polymorphism and CVD was identified in the analyzed population. However, significant differences in vitamin D metabolite levels were observed depending on the genotype in the patient group. In heterozygotes, the average 25(OH)D
3 concentration was 35.9 ng/mL, while in GG and TT homozygotes, it was 20.06 ng/mL and 14.66 ng/mL, respectively (
p = 0.034,
Table 7). This is consistent with the findings of Meirina et al., who analyzed 25(OH)D
3 levels in pregnant women and identified the highest levels in those with the GT genotype and the lowest in TT homozygotes. In this group, the correlation between the G allele and reduced vitamin D levels was significant at a statistical trend level (
p = 0.057) [
44].
Additionally, Hu et al. observed the best vitamin D supplementation results in individuals with the GT genotype [
45]. Significant differences in 1,25(OH)
2D
3 levels were also noted among patients (
p = 0.032), with the highest concentrations observed for the GT genotype (92.9 pg/mL) and lower for GG (43.4 pg/mL) and TT (28.1 pg/mL). Post-hoc analysis showed that 1,25(OH)
2D
3 levels were significantly lower in individuals with the TT genotype than the GT genotype (
p = 0.027).
CYP27B1 is the enzyme responsible for 1α-hydroxylation of 25(OH)D
3 to calcitriol. Therefore, these results suggest that the TT genotype may reduce the enzymatic activity of
CYP27B1. In the group of healthy volunteers, statistically significant differences were observed in 25(OH)D
2 concentrations depending on genotype (
p = 0.043). In the case of TT homozygotes, the 25(OH)D
2 level was the highest and amounted to 13.9 ng/mL, while in the case of heterozygotes and GG homozygotes, it was 7.9 ng/mL and 6.3 ng/mL, respectively (
Table 8). Differences in calcitriol concentrations were also observed, with healthy individuals with the TT genotype having an average concentration of 259.4 pg/mL, while in individuals with the GT and GG genotypes, the concentrations were 57.6 pg/mL and 66.1 pg/mL, respectively.
However, it is essential to note that vitamin D metabolism in the body is a complex process, so other factors not analyzed in this study may contribute to the differences in metabolite levels. Nevertheless, identifying the rs10877012 CYP27B1 genotype may help determine appropriate vitamin D supplementation doses for individuals with CVD.
5. Limitations
Our study is the first to report the genetic distributions of
CYP2R1,
CYP27A1, and
CYP27B1 genotypes and their associations with vitamin D hydroxy metabolites, including 25(OH)D
2, 25(OH)D
3, 3-epi-25(OH)D
3, 3-epi-25(OH)D
2, and calcitriol, in healthy Polish volunteers and patients with CVD. However, it is essential to acknowledge the limitations of this study. Firstly, we did not account for seasonal variations, which could have influenced metabolite levels—seasonal analysis was performed in our previous observational study [
46]. Secondly, there was a notable age difference between participants with CVD (median 68 years) and healthy participants (median 35.5 years), primarily due to the challenge of recruiting older individuals without cardiovascular conditions. Therefore, splitting the results reporting for healthy volunteers and patients was also essential. Third,
rs6709815-CYP27A1 could not be identified in two patients and four healthy group members. Hence, despite the chi-square analysis indicating statistical significance (
p < 0.05) and the
CYP27A1-TT genotype approaching the threshold for statistical significance, with an odds ratio of 4.22 (95% CI: 0.983–18.127;
p = 0.053), this result should be considered a proof of concept rather than proof of evidence.
Fourth, we did not adjust
p-values for multiple comparisons because we aimed to explore each genotype separately. However, we acknowledge that this approach may increase the risk of Type I errors, and findings should be interpreted cautiously. Lastly, we did not collect data on patient behaviors and lifestyle factors that may influence vitamin D status. For instance, supplementation patterns or attitudes among healthy volunteers were not identified for all participants. Consequently, the findings of this study should not be generalized and interpreted with caution until confirmed by a larger sample size, because only 53 participants were enrolled; this work should be regarded as a pilot study.Post-hoc power for the three target SNPs did not exceed 25%. A prospective calculation (α = 0.05, 1-β = 0.80, OR = 2.0) and minor allele frequency (rs6709815, 0.47 [
47]; rs10741657, 0.37 [
48]; and rs10877012, 0.47 [
36]) show that 135–155 cases and an equal number of controls would be required for a definitive study.



 ,
, 



{kind=link}
{kind=link}
{kind=link}