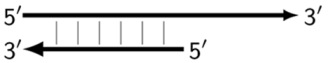
2.1. Nomenclature of Target Zones in the LAMP Reaction
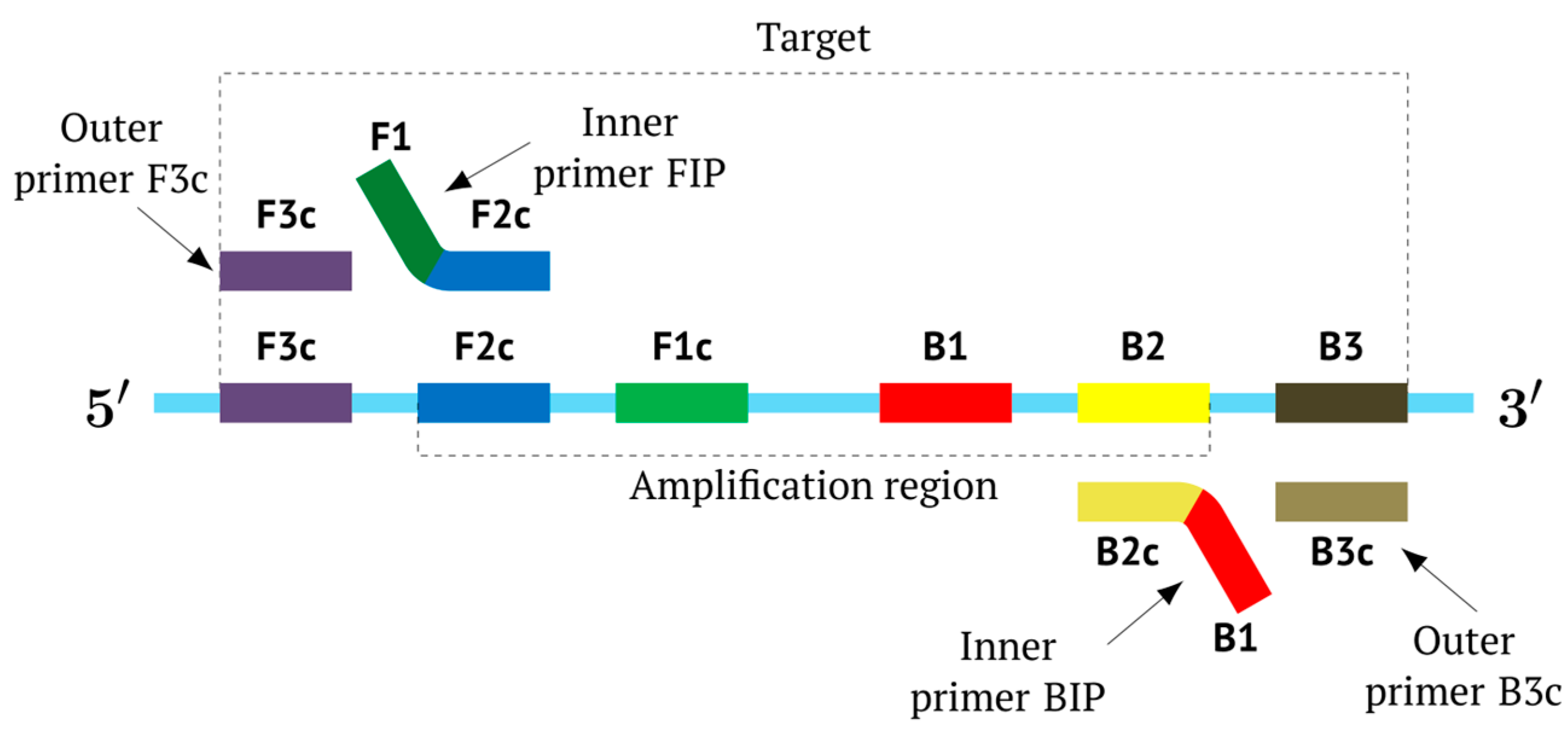
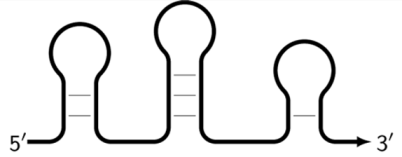
As noted above, the structure of lamplicons in the LAMP reaction is extremely complex, and their diversity cannot be described simply. The first obstacle is the nomenclature of zones in a target, which was proposed in 2000 (
Figure 1). According to this, the zones on one DNA strand are designated both with and without the letter ‘c’ (meaning ‘complementary’). This approach cannot be considered highly correct. To make it easier to operate with primers and lamplicons, the names of primer annealing zones should be modified. We propose single-letter symbols for all primer annealing zones. Instead of zones F3c, F2c, F1c, B1, B2, and B3, the following should be used: F, A, B, C, D, and E and, respectively f, a, b, c, d, and e (
Figure 3). The annealing zones of loop primers LF and LB can be labeled, for example, as X, Y and x, y. In general, all these zones can be labeled as Z and z (meaning a zone), supplied with a number corresponding to an ID number of such zones in a particular lamplicon. The primers used in the LAMP reaction will have the following letter designations, either uppercase or lowercase (reflecting their annealing on the starting or complementary strands): outer primer F3c will be “F”, the other outer primer B3 will then appear as “e”, while loop primers LF and LBc will be “X” and “y”, respectively. As for composite primers FIP and BIP, given that they carry two annealing zones (F1c|F2 and B1c|B2), they can each be labeled with two letters: 5′-bA-3′ and 5′-Cd-3′, respectively. Furthermore, where a direction of a zone sequence is not specifically indicated (including for primers), it will be 5′→3′ by default. The distinct feature of our proposed nomenclature is that all zones belong to one strand, and the letter ‘c’ is no longer required. This is because on a presumed upper strand, letters designating the zones are capitalized, whereas on the complementary (lower) strand (not shown in
Figure 1), they are lowercase.
Nevertheless, we by no means propose replacing the already customary names of primer bands in LAMP reactions, as will become evident in the description below. The replacement by a one-letter designation has only a technical purpose and is necessary only for a better understanding of processes taking place in this very complicated reaction. On the other hand, it is necessary to recognize that many additional designations have to be introduced, without which it is impossible to paint a complete picture of the processes occurring during LAMP, but these names do not detract from the general logic of this reaction.
Thus, all zones on the initial DNA strand serving as a target (T1) or templates T2, T3, and T4 for primer annealing in a LAMP reaction without loop primers can be depicted as 5′-FABCDE-3′, where dotted lines symbolize flanking sequences of indefinite length. When a LAMP reaction is carried out with loop primers (which are outside the scope of this paper), zones in the T1 target may be labeled 5′-FAXBCYDE-3′.
It should be noted that the structures described below may occur with certain probabilities during LAMP, in accordance with the competition between primer annealing and self-priming, which in turn depends on specific nucleotide sequences of the various zones in the target. Because they may differ greatly among different objects under study (targets), we will have to consider here all possible types of formation of hairpin structures and annealing zones both for primers and for 3′ ends of the generated lamplicons, which provide self-priming.
2.3. Types of Products in the LAMP Reaction
The first dumbbell-like structure 5′-
bABCDc-3′ Z6_dmb(1) generated during the preamplification phase is one of basic elements of any LAMP reaction and starts the cascade of subsequent events. Nevertheless, before proceeding to a detailed review of the processes within the LAMP cycle as well as outside it, it is necessary to focus on different types of lamplicons arising during LAMP. Namely, previously, Kaur et al. [
17] have identified four categories among LAMP reaction products: SL (single loop), T (terminated), SS (single-stranded), and PDS (partially double-stranded), and affixed Roman numerals to some of them: I, II, and III. In another article [
18], investigators analyzed the second phase of the LAMP reaction starting from dumbbell-like structures, calling it IDEA (isothermal dumbbell exponential amplification), using slightly different starting structures, which were designated as a short dumbbell, middle dumbbell, and long dumbbell; all differences among them consisted of different lengths of small additional nucleotide sequences in the region between zones F1c and B1, according to
Figure 1. Self-priming of the initial dumbbell results in the formation of hairpin B2c = Hp1b. After several displacements of strands, a hairpin emerges that is symmetric to it; this hairpin was named hairpin 1 F2c = Hp1f.
Nonetheless, some other types of lamplicons also arise during the LAMP reaction, which also need to be examined (
Table 1).
Table 1.
Types of products in classical loop isothermal amplification—LAMP.
Table 1.
Types of products in classical loop isothermal amplification—LAMP.
| Picture | Name | Brief Description |
|---|
| LAMP reaction intermediates |
![Biomolecules 15 00690 i003]() | Z_dmb(1)Z_dmb(2) | Dumbbell-like structures in the form of single- or double-stranded DNA. The letter Z (zone) is accompanied by a number corresponding to the ID number of zones in specific products. Z6_dmb(1) and Z6_dmb(2) are the main types of lamplicons that trigger cascades of subsequent processes of enzymatic construction of DNA strands and ensure exponential amplification. In fact, strands bABCDc and CdcbaB are dual mirror twins with respect to each other. |
![Biomolecules 15 00690 i004]() | Z9_dmb(1)Z9_dmb(2) | Elongated dmb structures in the form of single/double-stranded DNA similar to Z6_dmb but differing in the length owing to a different number of zones, e.g., Z9_dmb or Z15_dmb. |
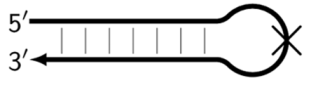
![Biomolecules 15 00690 i005]() | Z_hp | Hairpin-like structure of single-stranded DNA (loop) and double-stranded DNA (stem). Arises from Z_dmb, e.g., Z6_dmb gives rise to Z9_hp. |
![Biomolecules 15 00690 i006]() | Z_li | Single-/double-stranded DNA structure (linear) resulting from linearization of hairpin structure Z_hp and then evolving into a new, larger Z_hp structure and also generating a smaller Z_dmb structure(s). |
| Terminated (stop) lamplicons in LAMP reaction |
![Biomolecules 15 00690 i007]() | ZzS | Double-stranded DNA structures generating by polymerase from structures like Z_li or Z_dmb (here, generated from Z6_dmb, Z9_li or Z9_dmb, respectively) and not participating in further amplification. |
![Biomolecules 15 00690 i008]() | Z_hpS | Hairpin-like structure in the form of double-stranded DNA (stem) and single-stranded DNA (loop), on which (on whose loop) the next primer cannot anneal, as indicated by the crossing out of the loop. |
| Other unstable products of LAMP reaction |
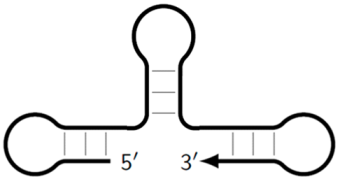
![Biomolecules 15 00690 i009]() | Z_dmb_hp | Dumbbell-like structure with looping in the stem part, on whose loop one of the inner primers is also able to anneal, followed by elongation and strand displacement. |
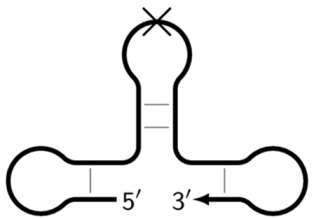
![Biomolecules 15 00690 i010]() | Z_dmb_hpS | Dumbbell-like structure with a hollowing out in the stem part, on whose loop the inner primers are unable to anneal, as indicated by the crossing out of the loop. |
![Biomolecules 15 00690 i011]() | Z_mtlp | Late-onset single-/double-stranded DNA containing multiple different loops that can also participate in further amplification. |
![Biomolecules 15 00690 i012]() | cauliflower-like | High-molecular-weight single-/double-stranded DNA arising at late stages, carrying strands annealed on each other in a chaotic manner. Limitedly suitable for further amplification. |
The probability of formation of certain types of products depends on their GC content and on stacking interactions of neighboring nucleotides in specific zones and cannot be predicted.
It should also be pointed out that in the types of LAMP reaction involving loop primers and stem primers, other types of products appear as semi-dumbbell (sdmb) structures, but we will not touch upon them here. Among the final products of the LAMP reaction are DNA molecules with double-stranded regions, representing the target with flanking single-stranded regions (shown by dotted lines) of unknown length:
![Biomolecules 15 00690 i013]()
The number of such molecules will approximately correspond to the initial number of copies of the desired sequence, and they will not make a serious contribution to the total amount of DNA produced during the LAMP reaction.
2.4. The Basic LAMP Cycle
After the appearance of the first dumbbell-like structure Z6_dmb(1), exponential amplification begins as the LAMP cycle, which continues by releasing a number of products into the outer contour, in which the next generation of extended lamplicons (NGEL) takes place and similar dumbbell-like Z6_dmb structures are formed, initiating similar LAMP cycles. The processes taking place are summarized in
Figure 4.
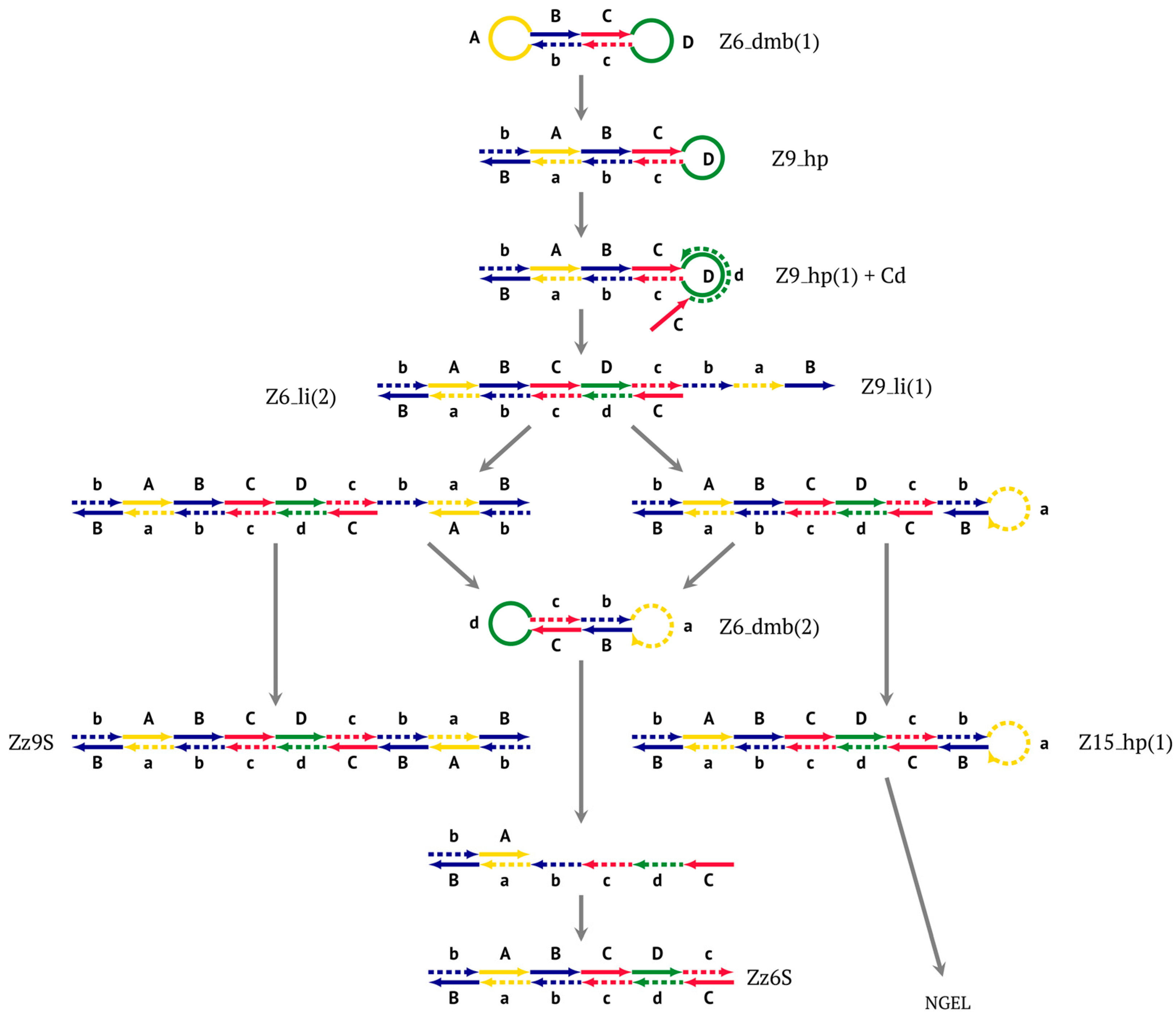
The most interesting is the LAMP cycle, which repeats many times and occurs at the next stages in the course of the formation of larger lamplicons during NGEL when they return to the main lamplicon Z6_dmb. The sequence of events in the LAMP cycle is depicted in
Figure 5.
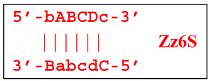
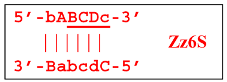
Upon entering the LAMP cycle, the
bABCDc strand appears suitable for annealing with the Cd primer, leading to the terminated Zz6S lamplicon (shown hereafter in red), the amount of which will increase during the LAMP reaction:
![Biomolecules 15 00690 i014]()
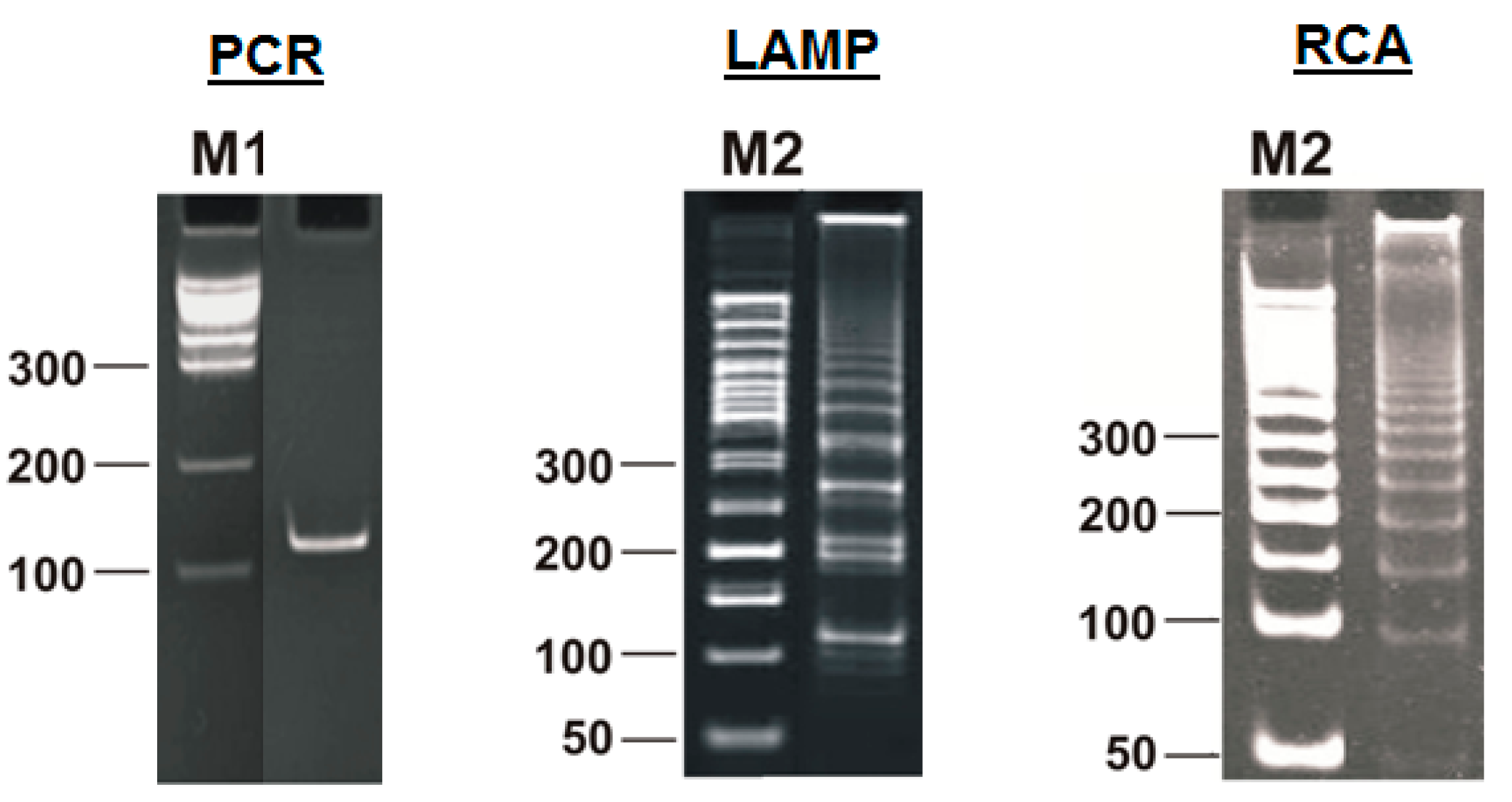
Moreover, only Zz6S and similar terminated double-strand lamplicons ZZS and also Z_hpS lamplicons are visible when products of the LAMP reaction are separated by gel electrophoresis. As for some of the following single-/double-stranded lamplicons, due to the isothermal nature of the process, at every moment among many similar structures there will be those differing from each other by an unequal number of nucleotides. Therefore, this situation will interfere with the formation of discrete DNA bands of equal or similar sizes visible to the naked eye in the gel. It is also worth noting that the probability of formation of the above-mentioned terminated double-stranded Zz6S lamplicon is rather high, if we take into account that the Cd primer will be annealed on the bABCDc structure in two zones (italicized here) at once, thus representing serious competition to self-priming, which means annealing only by a single zone.
Nevertheless, the dumbbell-like structure of
bABCDc—owing to self-priming (annealing of zone ‘
c’ on zone ‘C’) and subsequent strand elongation—gives rise to a new hairpin structure (Z9_hp_D, where hp is a hairpin) with a loop in the form of zone D:
![Biomolecules 15 00690 i015]()
At the same time, primer Cd can be annealing (at first with one zone d) on the single-stranded loop formed by the D zone, whose elongation by DNA polymerase will cause the displacement of the strand and will transform the Z9_hp_D strand into a linearized version [Z9_li (linear)], which will have a double-stranded section carrying the Z6_dmb(2) strand:
![Biomolecules 15 00690 i016]()
On the Z9_li(1) strand, the bA primer is also able to anneal (with two zones), leading to both a terminated lamplicon
![Biomolecules 15 00690 i017]()
and to the displacement of the
CdcbaB strand, forming in turn the second main dumbbell-like structure Z6_dmb(2) with a symmetric sequence in relation to Z6_dmb(1). They are in fact dual mirror twins. Indeed, the main dumbbell-like lamplicons in the form of strands Z6_dmb(1) and Z6_dmb(2) are fully complementary; if they are present in the reaction mixture in large amounts at late stages of the LAMP reaction, then they can form a double-stranded terminated structure Zz6S by simply annealing on each other. Dumbbell-like structure Z6_dmb(2) is able to elongate itself by means of polymerase to form a new hairpin with a loop in the form of the “a” zone:
![Biomolecules 15 00690 i018]()
Annealing of primer bA (initially with one zone A) on the a loop of the Z9_hp_a strand and its elongation will enable the annealing of primer Cd (with two zones) and the formation of a number of structures, among which is terminated lamplicon Zz9S(2), which has a sequence different from the similar previous Zz9S(1) lamplicon:
![Biomolecules 15 00690 i019]()
The
bABCDc strand (built from primer bA)—displaced by polymerase as a result of primer Cd elongation—will repeat the structure of the first base lamplicon Z6_dmb(1), which will start a new cycle:
![Biomolecules 15 00690 i020]()
Thus, within one LAMP cycle in the classic version of this reaction, six intermediate products and four terminated lamplicons will be generated as well as two elongated hairpin structures Z15_hp, from which other structures of both larger and smaller sizes will arise later. Here, we can also note that both within the LAMP cycle and during the subsequent multiplication processes, only “c” and “B” act as self-priming zones. This phenomenon should be taken into account when primer systems are designed, with special attention to the 5′ ends of primers FIP (bA) and BIP (Cd) because the 3′ ends of their complementary strands will be elongated by polymerase during self-priming.
2.5. Continuation of the LAMP Reaction Beyond the Basic LAMP Cycle
Thus, two different Z15_hp strands, once outside the LAMP loop, will lead to further growth of DNA strands during the LAMP reaction and form new self-extending hairpin structures; because they are symmetric, it is enough to trace descendants of only one:
![Biomolecules 15 00690 i021]()
The Cd primer will anneal on the D loop formed in Z15_hp (initially with one zone d) followed by linearization of the strand, resulting in a new single/double-stranded structure:
![Biomolecules 15 00690 i022]()
On the remaining single-stranded site, another primer, Cd, will be annealed, causing the previous CdcbABCDc (Z9_dmb) strand to be displaced and causing the formation of a terminated double-stranded lamplicon, which already consists of 15 zones:
![Biomolecules 15 00690 i023]()
As for the fate of the CdcbABCDc strand (Z9_dmb), it may become a Z15_hp lamplicon or form another lamplicon as a Z_hpS hairpin structure owing to self-annealing:
![Biomolecules 15 00690 i024]()
This Z9_hpS lamplicon is a terminated structure because it is a hairpin on which none of the primers present in the reaction mixture can be annealed. This and similar structures will move disproportionately to the location corresponding to the total length of the nucleotide sequence during gel electrophoresis owing to the presence of a single-stranded region with different electrophoretic mobility. Thus, as stated above, during the LAMP reaction, terminated lamplicons are formed, representing not only double-stranded DNA fragments but also some hairpin structures unsuitable for primer annealing on them. This notion also explains the presence in the electrophoretic image of binary and even triple DNA bands with similar sizes and characterized by slightly different mobility in the gel.
When hairpin structure Z15_hp_D is generated, another self-priming of the created structure will occur with intermediate linearization and subsequent formation of a new hairpin:
![Biomolecules 15 00690 i025]()
Upon annealing of another Cd primer on the Z27_hp_D hairpin structure (initially with a single d zone), hairpin opening and subsequent linearization of the strand will again occur:
![Biomolecules 15 00690 i026]()
Two more primers, bA and Cd, can anneal on the Z27_li(2) strand, respectively, leading to this structure:
![Biomolecules 15 00690 i027]()
and in the latter case with the Cd primer, to a terminated lamplicon with 27 zones:
![Biomolecules 15 00690 i028]()
and also to displaced strands Z15_dmb and Z18_dmb.
At the same time, for the displaced strands Z15_dmb and Z18_dmb, there are already three types of self-forming hairpin structures, with subsequent completion of some of them. Thus, the following conversions will occur with the Z15_dmb strand:
![Biomolecules 15 00690 i029]()
If the first two lamplicons, Z15_hpS and Z21_hpS, are terminated, then lamplicon Z27_hp will turn into structure Z27_li owing to annealing of primer Cd on it (at first by one zone d), which will then become structure Z51_hp and start a new NGEL stage of the LAMP reaction.
The transformation of the Z15_dmb strand will not finish here: two primers, Cd and bA, will be able to anneal on it, and if the former forms a terminated Zz15S lamplicon, the latter will give rise to another dumbbell-like structure bABCDc Z6_dmb(1), which will serve as an entry structure into a new LAMP cycle:
![Biomolecules 15 00690 i030]()
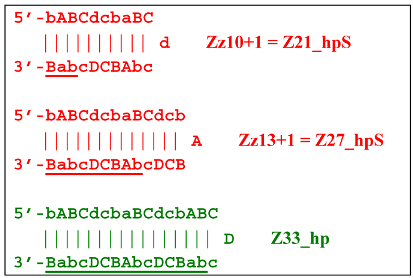
As for lamplicon Z18_dmb’s behavior, it will be similar to that of the Z15_dmb lamplicon, but with some differences. Lamplicon Z18_dmb will also form three hairpin structures:
![Biomolecules 15 00690 i031]()
If the first two structures, Z21_hpS and Z27_hpS, are terminated, the last Z33_hp will continue the multiplication of DNA strands, entering a new NGEL stage of the LAMP reaction, and will continue to develop lamplicons of its size series. It can be observed here that the Z21_hpS structures that formed from strands Z15_dmb and Z18_dmb, while having the same size, have different compositions, including different loops, A and d, respectively.
In addition to all kinds of hairpin structures arising due to self-annealing, primers bA and Cd can be annealed on the Z18_dmb strand. In this case, elongation of the latter primer will lead to a terminated Zz18S lamplicon. In contrast, elongation of primer bA will lead to a dumbbell-like Z9_dmb lamplicon and through it both to the Z15_hp strand in the direction of increasing product length and to the Z6_dmb(2) lamplicon in the direction of decreasing length, thereby starting its LAMP cycle:
![Biomolecules 15 00690 i032]()
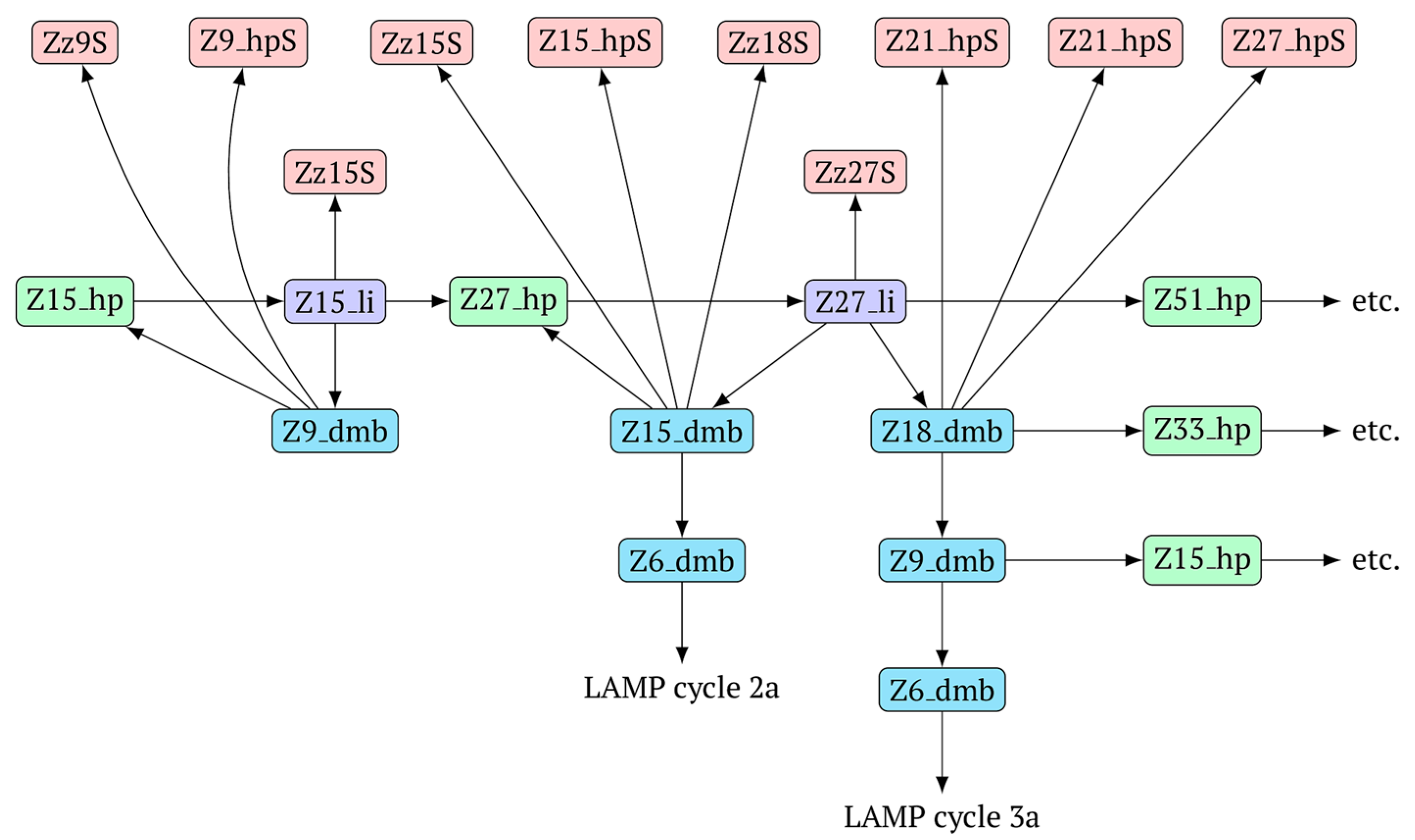
The above-mentioned amplification processes in the LAMP reaction for one lamplicon Z15_hp(1) formed in the first LAMP cycle can also be displayed as a scheme. This scheme implies that for a second (similar) lamplicon Z15_hp(2) formed in the same LAMP cycle 1, the outcomes will be symmetric, with the only difference being that the loop zones will be different (
Figure 6).
As evident from the scheme in
Figure 6, after the formation of lamplicon Z27_li and the displacement of strands Z15_dmb and Z18_dmb, two new main dumbbell-like structures Z6_dmb are formed, which will start new LAMP cycles. Considering that the reactions are symmetric, in the scheme, they are marked 2a and 3a, implying that there will also be similar LAMP cycles 2b and 3b during this exponential NGEL phase.
Regarding the growth of strands during the LAMP reaction, owing to self-priming, the increase in the number of zones in the lamplicons obeys the following formula:
Thus, the main pathway of generation of lamplicons in the direction of their elongation looks as follows: Z6 → Z9 → Z15 → Z27 → Z51 → Z99 and so forth. For the other pathway, the following series is lined up: Z18 → Z33 → Z63 → Z123 and so forth. In addition to them—because there are a lot of places for primer annealing in lamplicons of increased size, including on the displaced strands—a variety of lamplicons can be formed whose lengths are also multiples of three. Terminated lamplicons will also have a number of zones that is a multiple of three. Nonetheless, it should be kept in mind that not all possible lamplicons will be produced in appreciable quantities, and whether they will be produced at all depends on specific features of nucleotide sequences of these or other zones in the selected target.
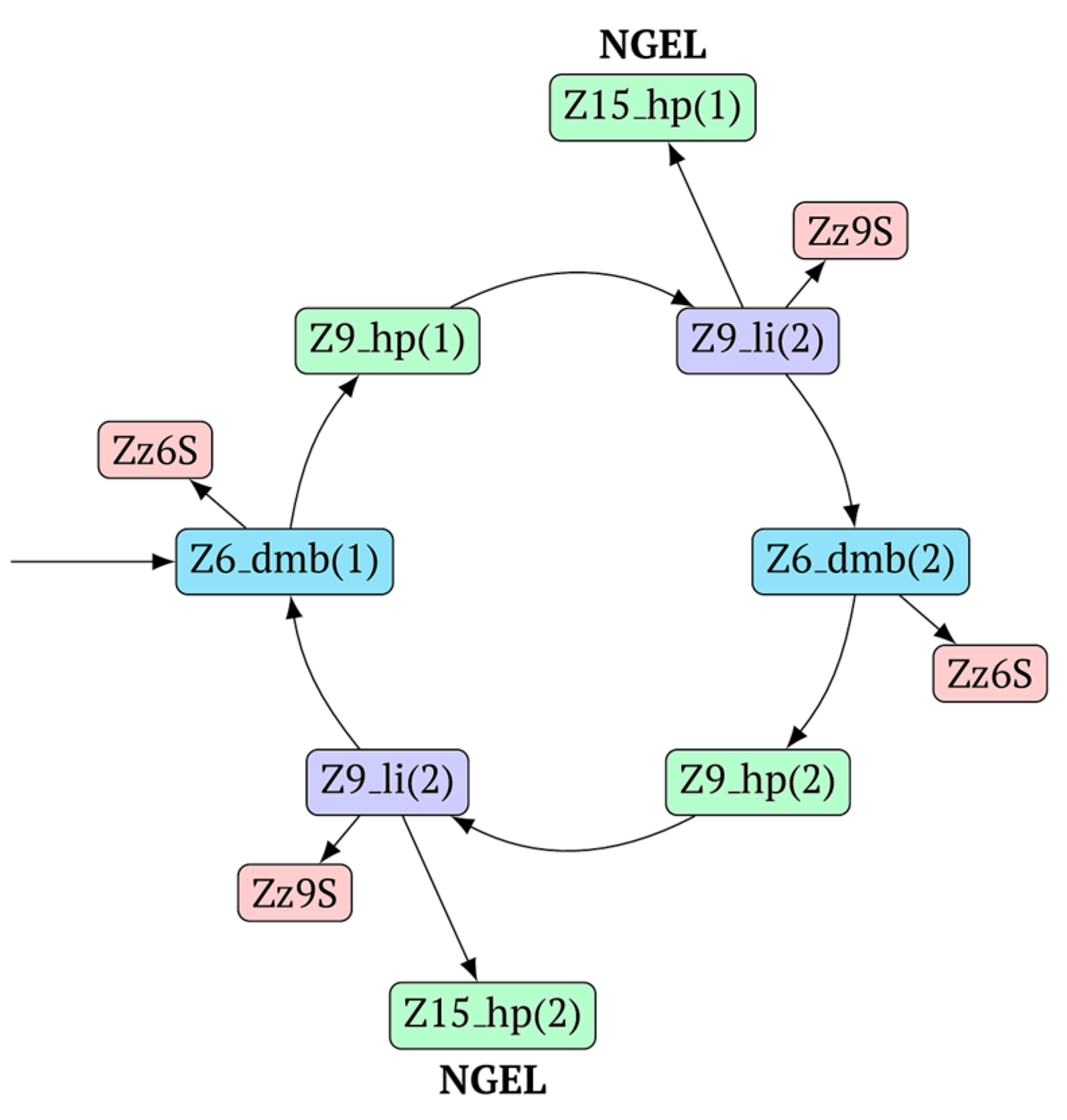
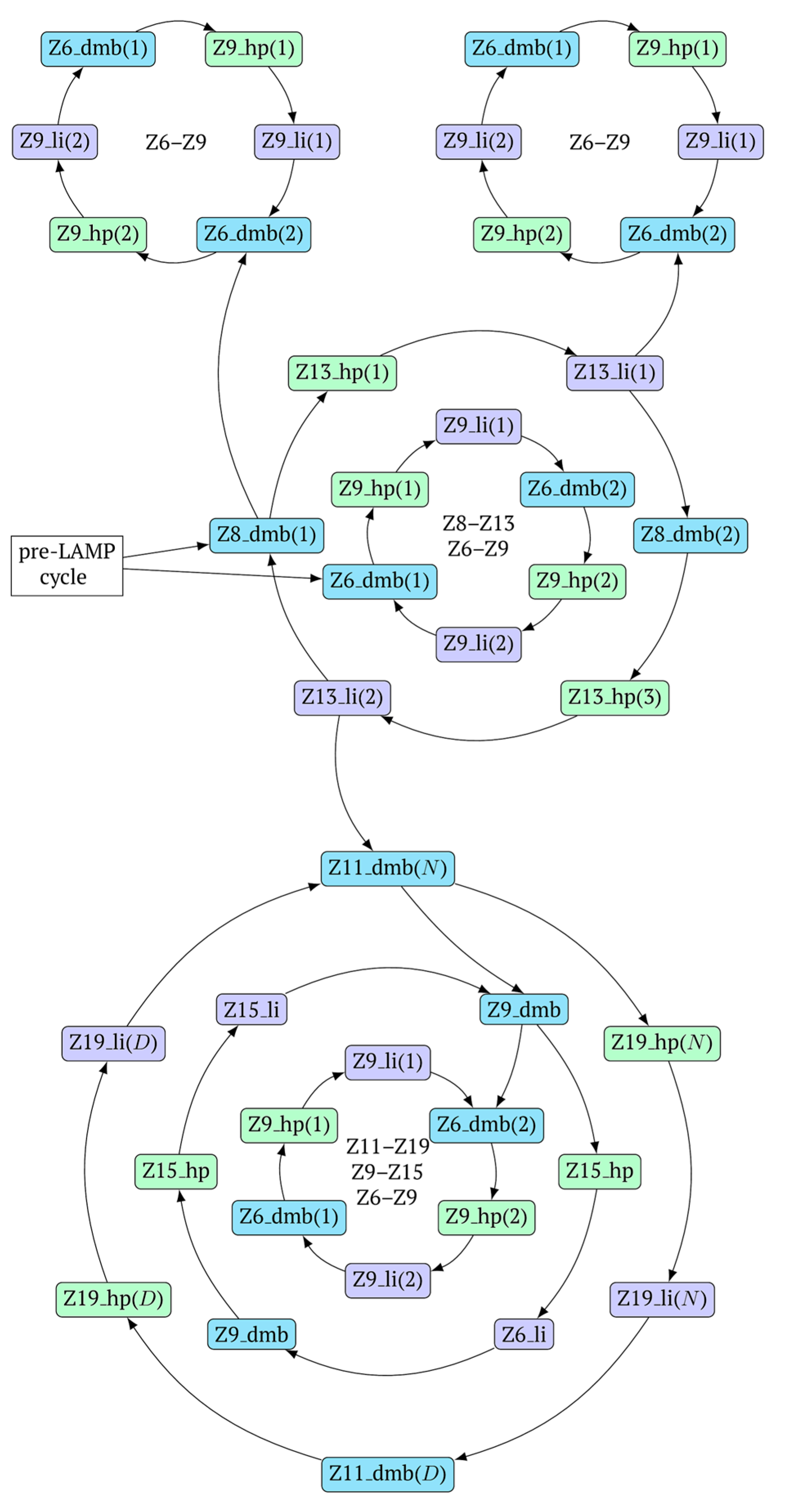
In general, during the LAMP reaction in its exponential phase, the following cyclic processes occur in the main size series, as shown in
Figure 7. At the same time, for each of these types (Z_dmb, Z_hp, and Z_li), the composition of the zones will slightly change, but their number within specific dimensional boundaries will be unchanged.
As one can see in
Figure 6 and
Figure 7, Z_dmb, Z_hp, and Z_li types of lamplicons are capable of transforming into each other, while generating many other lamplicons of different types.
Furthermore, during the exponential phase of the LAMP reaction up to its plateau, various other variants with different sizes of lamplicons can be formed owing to possible hairpin looping due to regularly repeating elements in the form of different Z_mtlp structures, and also due to the formation of other unpredictable structures. This is because in the reaction mixture after some time, a huge number of single-stranded DNA molecules emerge over large regions complementary to each other; these are called “cauliflower-like” structures. At the same time, all these diverse DNA molecules contribute to the overall DNA amplification when they are detected by means of nonspecific fluorescence with intercalating dyes such as SYBR Green I or with the help of byproducts of the DNA polymerization reaction. In these cases, turns of the DNA helix or consumed dNTPs, which transformed into dNMPs and pyrophosphates with a proton release, “work” for detection.
2.6. Pseudo-Hemi-Nested LAMP(phn-LAMP) Reaction
Of separate interest are LAMP reactions with an accelerated type of amplification due to additional primers. Neither loop nor stem primers give a very significant increment, and LAMP reactions with multiple inner primers, some which have been proposed earlier [
7,
20,
21,
22], look more promising. Nonetheless, apparently due to a significant increase in target length and elevated probability of primer dimerization, these types of LAMP reactions have not become popular. In particular, during the pandemic of novel coronavirus SARS-CoV-2, one paper described the RT-MIPLAMP method. When two pairs of inner primers were used, this method showed an advantage in sensitivity and speed of obtaining results as compared to the conventional RT-LAMP reaction; however, when the number of inner primers was increased to three pairs, nonspecific amplification began [
22]. Also, a serious disadvantage of such LAMP reactions with an increased number of inner primers is the greater length of the target.
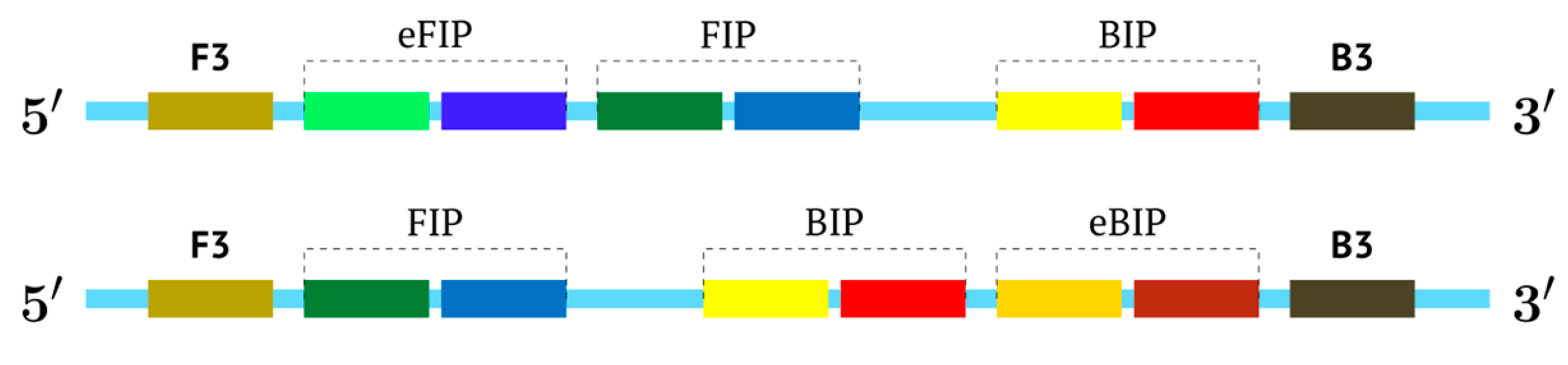
An intermediate option (a compromise) could be the use of only one additional inner primer, and this approach would resemble semi-nested PCR. In the process, however, nested primers enter amplification later, which arises due to temperature differences in primer annealing or by some barrier method, if the tubes are not opened. For the LAMP reaction, these approaches are not acceptable, and therefore we named such a reaction pseudo-hemi-nested LAMP (phn-LAMP). Its advantage over MIP-LAMP variants is that only two annealing sites are added (
Figure 8); this approach does not significantly increase the length of the target and makes it comparable to LAMP variants involving loop or stem primers but at the same time provides a much higher level of DNA amplification, as readers can see from the following modeling of this reaction.
In this case, the character of DNA amplification during the LAMP reaction will be the same both for primers FIP and eFIP (extra) together with a single BIP and vice versa: with a single FIP together with BIP and eBIP.
Nevertheless, for a better explanation of this type of LAMP reaction, it is also necessary to switch to the previously used coding of zones and primers. Thus, in phn-LAMP with the single-letter coding in one of the possible types of FIP and BIP primers’ arrangement, the target would look like this: 5′-…FABCDMNE…-3′, while the primers would be F, bA, Cd, Mn, and e. The initial steps of the preamplification phase can be displayed as follows:
![Biomolecules 15 00690 i033]()
As a result, two types of dumbbell-like structures,
bABCDc [Z6_dmb(1)] and
bABCDMNm [Z8_dmb(1)], will be generated during the preamplification phase of the phn-LAMP reaction. When they enter the LAMP cycle through a series of processes, structures symmetric to them—
CdcbaB [Z6_dmb(2)] and
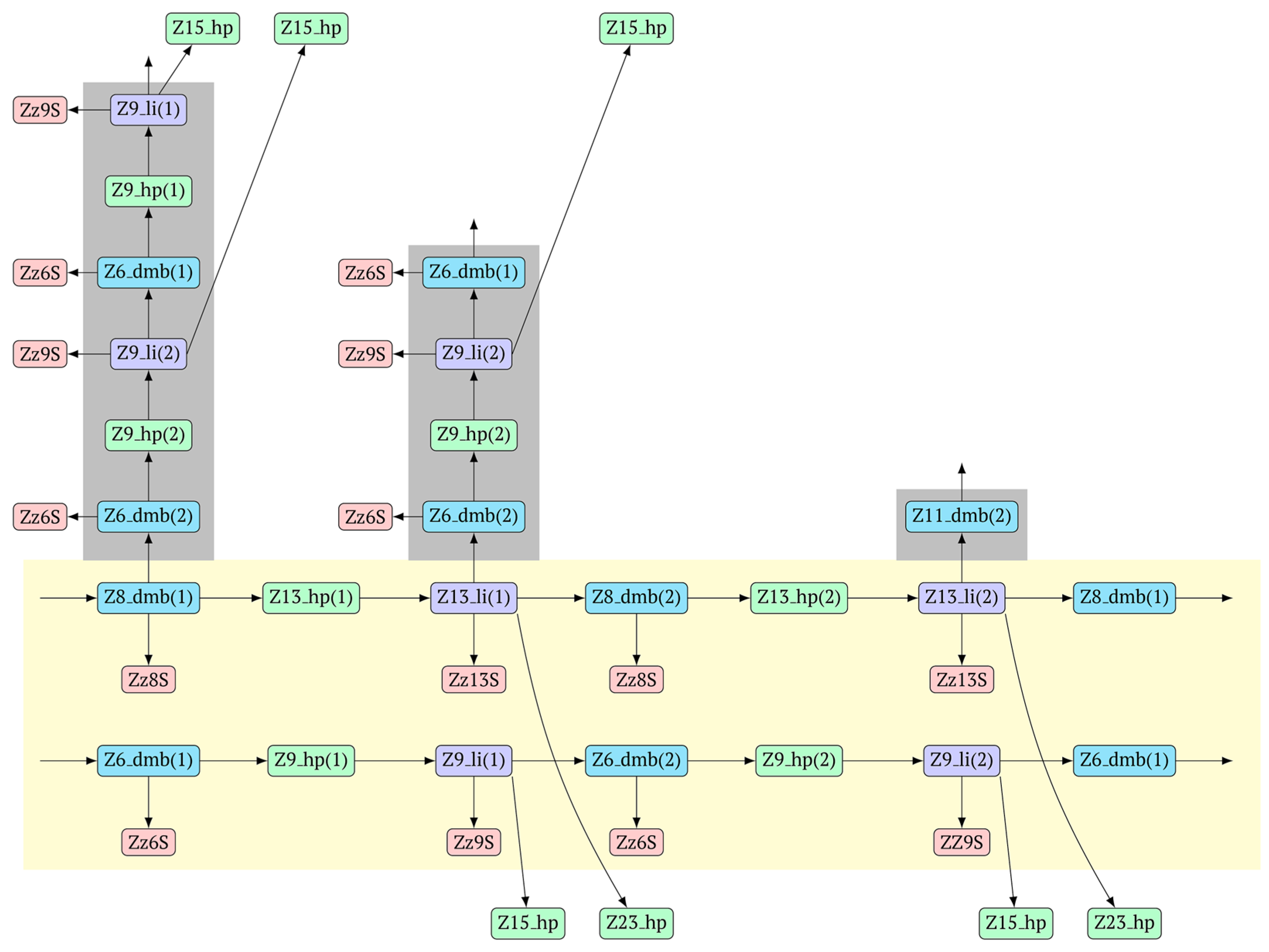
MnmdcbaB [Z8_dmb(2)]—will appear. The increment of lamplicons during one main LAMP cycle in phn-LAMP is illustrated in
Figure 9.
In fact, the transition of the phn-LAMP reaction to the exponential phase results in two LAMP cycles in one, accompanied by subcycles, as depicted in the simplified scheme of these processes in
Figure 10. The Z11_dmb(2) lamplicon, which generates its own subcycles, deserves special attention.
The phn-LAMP subtype designed for detection of eight zones in a target proceeds in a way similar to the classic LAMP reaction, with a difference being that after preamplification, dmb structures of two sizes—Z8_dmb(1) and as its derivative Z6_dmb(1)—will initially emerge in the reaction mixture. The Z6_dmb(1) structure behaves similarly to that in the classic LAMP reaction, whereas Z8_dmb(1) will immediately generate another Z6_dmb(2) structure, which will come again to dmb(2) via dmb(1).Z8_dmb will then engender three more Z6_dmb-like structures.
In the phn-LAMP reaction, taking into account some presumed delay of strand elongation (which is highlighted with the gray background), 18 terminated lamplicons are formed (Zz6S: eight, Zz8S: two, Zz9S: five, and Zz13S: two) and 7 other lamplicons (Z15_hp: five and Z23_hp: two) go out into the NGEL phase. In contrast, in the classic LAMP reaction with a pair of inner primers (FIP and BIP), the number of lamplicons is much smaller: Zz6S, Zz9S, and Z15_hp, two of each. The number of larger lamplicons entering the next loop is also important for subsequent increased multiplication of strands (in the LAMP reaction), which further enhances the exponential nature of such amplification. As for the main type of Z6_dmb lamplicon, in the classic LAMP reaction, the number of copies of such lamplicons that entered each LAMP loop is the same as the number of such lamplicons that exit. There is the same situation with Z8_dmb lamplicons in phn-LAMP, but in this reaction, instead of one Z6_dmb lamplicon that entered, five times more of them come out of each LAMP cycle. In addition to them, another new structure, Z11_dmb, emerges in the next cycle.
With the help of the single-letter coding, it is convenient to trace the development of phn-LAMP in a LAMP cycle. Because the
bABCDc lamplicon [Z6_dmb(1)] that formed during the preamplification phase of phn-LAMP behaves similarly to that in the normal LAMP reaction, its transformations will not be discussed here. As for the
bABCDMNm [Z8_dmb(1)] lamplicon, Z6_dmb lamplicons periodically arise from it and will also repeat the standard LAMP cycle in
Figure 9; they are highlighted by means of the gray background of different heights in accordance with the processes occurring in the main LAMP subcycle.
Thus, the following structures emerge from Z8_dmb(1):
![Biomolecules 15 00690 i034]()
When the Mn primer is annealed, a terminated Zz8S lamplicon is produced, and when the Cd primer is annealed before it, a displaced Z6_dmb(2) strand is formed that can initiate a new LAMP cycle:
![Biomolecules 15 00690 i035]()
At the same time, the bA primer can be annealed on the Z6_dmb(2) strand, leading to a terminated Zz6S lamplicon:
![Biomolecules 15 00690 i036]()
Additionally, for the Z8_dmb(1) structure, there is a probability of self-priming, giving lamplicon Z13_hp(1):
![Biomolecules 15 00690 i037]()
on the hairpin of which primer Mn is able to anneal (initially with one n zone), leading to the opening of the hairpin and emergence of the Z13_li structure. It also contains the Z8_dmb(2) strand, which can be displaced by the elongation strand from primer bA annealed upstream or by self-priming, leading to the NGEL stage and to lamplicon Z23_hp:
![Biomolecules 15 00690 i038]()
Once the Z8_dmb(2) structure is displaced, it can engender a terminated lamplicon:
![Biomolecules 15 00690 i039]()
where Z8_dmb(1) and Z8_dmb(2) are dual mirror twins with respect to each other.
The Z8_dmb(2) strand can also form a new hairpin structure, Z13_hp(2):
![Biomolecules 15 00690 i040]()
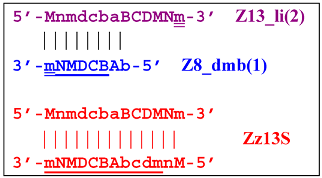
Moreover, the Z13_hp(2) structure—with a loop in the form of zone “a” owing to primer bA annealing on it (first via one zone A)—will cause the opening of the hairpin structure of lamplicon Z13_li(2) and then to terminated lamplicon Zz13S:
![Biomolecules 15 00690 i041]()
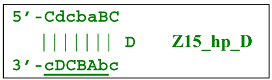
A peculiarity of the Z13_li(2) strand, in contrast to its counterpart Z13_li(1), is that the Cd primer is able to anneal on it, and this event will ensure the formation of a new size range of the lamplicon lineage starting from the Z11_dmb(N) structure, where the type of loop is indicated by a letter instead of numbering “1” or “2”:
![Biomolecules 15 00690 i042]()
Moreover, within the same LAMP cycle, a new displaceable dumbbell-like Z11_dmb(1) strand will emerge, which will cause the following: from the first LAMP cycle, in the phn-LAMP reaction, an additional somewhat elongated dmb structure will emerge, which already in the next such cycle will turn into Z19_hp, entering the next NGEL step. Through temporary structure Z9_dmb, this molecule will give rise to another basic lamplicon, Z6_dmb(1), initiating a new LAMP cycle.
Regarding strand Z13_li(2), its self-priming will produce hairpin structure Z23_hp, which will exit to the next NGEL contour:
![Biomolecules 15 00690 i043]()
and derivatives of Z11_dmb will continue transformations already in the next LAMP cycle:
![Biomolecules 15 00690 i044]()
Aside from the formation of the Z6_dmb(1) lamplicon, the Z9_dmb(D) lamplicon will turn into the Z15_hp(D) hairpin structure by annealing on its primer loop. Then, after annealing of the Cd primer (initially with one zone d), it will become the Z15_li(D) structure containing Z9_dmb(D), unable to become Z6_dmb:
![Biomolecules 15 00690 i045]()
When the LAMP subcycle with Z11_dmb(N) is initiated after annealing and elongation of the Mn primer, a terminated Zz11S lamplicon will emerge:
![Biomolecules 15 00690 i046]()
whereas self-framing will result in a Z19_hp hairpin structure:
It is then linearized, thereby yielding the following structures after annealing and elongation of primers Mn and bA:
![Biomolecules 15 00690 i047]()
After annealing of the Cd primer, a terminated Zz19S lamplicon will appear:
![Biomolecules 15 00690 i048]()
As for the Z16_dmb(D) lamplicon, it will undergo the following transformations—by annealing with three primers (Cd, bA, and Cd) on it—as well as self-priming:
![Biomolecules 15 00690 i049]()
The increase in the size of the lamplicons in the phn-LAMP reaction follows the same logic. When the Z_dmb structure is transformed into its derivative structure Z_hp, the increment takes place by actually doubling the zones minus three zones. That is, starting from Z8_dmb, sizes of subsequent lamplicons in terms of the number of zones will be as follows: Z8 → Z13 → Z23 → Z43 → Z83 → Z163 and so forth. This is the main size series of lamplicons, but as already mentioned above, subsequently, there will be others, starting with lamplicons with an initially larger number of zones.
In this case, the use of one additional inner primer gives a certain advantage, because on the target, there will be a choice of annealing sites or two FIPs and one BIP, or vice versa. It depends on specific sequences of the target. We have previously created primer design software LAMPrimers_iQ (
https://github.com/Restily/LAMPrimers-iQ) [
23], into which we just added a special option for the selection of primers for phn-LAMP; this feature is designed to select the optimal locations of zones for the three inner primers for this or that type of reaction, as shown in
Figure 8. For the selection of annealing sites and for the design of the corresponding primers, it is easier to choose sites for either two FIPs (FIP and eFIP) or two BIPs (BIP and eBIP) than for two loop or stem primers. This is because they must occupy specific locations in the target, thereby narrowing the freedom of choice.

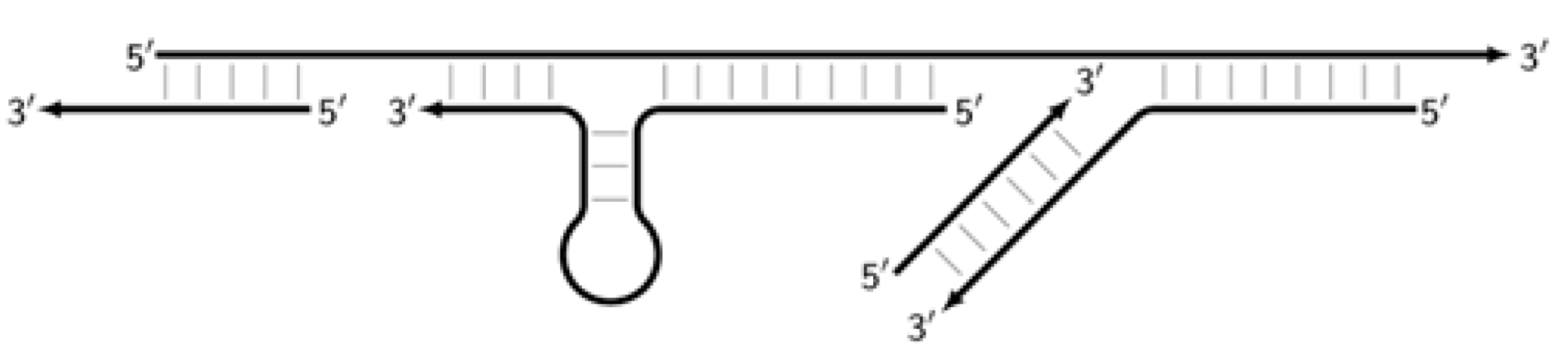
 ,
, 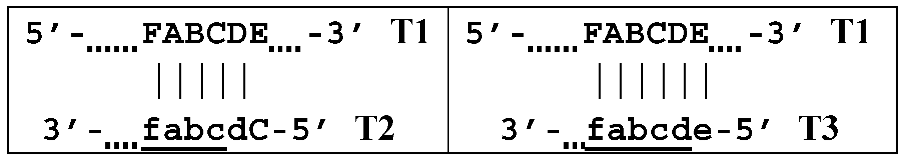 where vertical lines collectively symbolize hydrogen bonds between complementary nucleotides in specific zones, and underlining indicates new strands being built from the primer, which itself remains underlined. The primer zones that served for self-priming will not be underlined either. Such strands together with the primer, being a whole, serve as new templates and are presented without underlining. Alphanumeric names denote the type of DNA strand built from the annealed primer. A similar coding will be used to describe processes occurring in the exponential phase of LAMP, including the main LAMP cycle, as well as the phn-LAMP reaction.
where vertical lines collectively symbolize hydrogen bonds between complementary nucleotides in specific zones, and underlining indicates new strands being built from the primer, which itself remains underlined. The primer zones that served for self-priming will not be underlined either. Such strands together with the primer, being a whole, serve as new templates and are presented without underlining. Alphanumeric names denote the type of DNA strand built from the annealed primer. A similar coding will be used to describe processes occurring in the exponential phase of LAMP, including the main LAMP cycle, as well as the phn-LAMP reaction.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




 and to the displacement of the CdcbaB strand, forming in turn the second main dumbbell-like structure Z6_dmb(2) with a symmetric sequence in relation to Z6_dmb(1). They are in fact dual mirror twins. Indeed, the main dumbbell-like lamplicons in the form of strands Z6_dmb(1) and Z6_dmb(2) are fully complementary; if they are present in the reaction mixture in large amounts at late stages of the LAMP reaction, then they can form a double-stranded terminated structure Zz6S by simply annealing on each other. Dumbbell-like structure Z6_dmb(2) is able to elongate itself by means of polymerase to form a new hairpin with a loop in the form of the “a” zone:
and to the displacement of the CdcbaB strand, forming in turn the second main dumbbell-like structure Z6_dmb(2) with a symmetric sequence in relation to Z6_dmb(1). They are in fact dual mirror twins. Indeed, the main dumbbell-like lamplicons in the form of strands Z6_dmb(1) and Z6_dmb(2) are fully complementary; if they are present in the reaction mixture in large amounts at late stages of the LAMP reaction, then they can form a double-stranded terminated structure Zz6S by simply annealing on each other. Dumbbell-like structure Z6_dmb(2) is able to elongate itself by means of polymerase to form a new hairpin with a loop in the form of the “a” zone: 







 and in the latter case with the Cd primer, to a terminated lamplicon with 27 zones:
and in the latter case with the Cd primer, to a terminated lamplicon with 27 zones:  and also to displaced strands Z15_dmb and Z18_dmb.
and also to displaced strands Z15_dmb and Z18_dmb.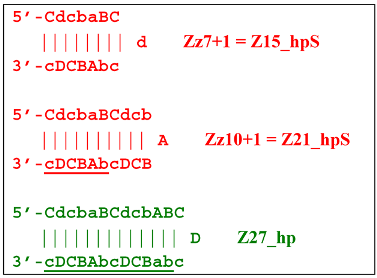





 on the hairpin of which primer Mn is able to anneal (initially with one n zone), leading to the opening of the hairpin and emergence of the Z13_li structure. It also contains the Z8_dmb(2) strand, which can be displaced by the elongation strand from primer bA annealed upstream or by self-priming, leading to the NGEL stage and to lamplicon Z23_hp:
on the hairpin of which primer Mn is able to anneal (initially with one n zone), leading to the opening of the hairpin and emergence of the Z13_li structure. It also contains the Z8_dmb(2) strand, which can be displaced by the elongation strand from primer bA annealed upstream or by self-priming, leading to the NGEL stage and to lamplicon Z23_hp: 
 where Z8_dmb(1) and Z8_dmb(2) are dual mirror twins with respect to each other.
where Z8_dmb(1) and Z8_dmb(2) are dual mirror twins with respect to each other.

 and derivatives of Z11_dmb will continue transformations already in the next LAMP cycle:
and derivatives of Z11_dmb will continue transformations already in the next LAMP cycle: 

 whereas self-framing will result in a Z19_hp hairpin structure:
whereas self-framing will result in a Z19_hp hairpin structure: