
The Need for Novel Asexual Blood-Stage Malaria Vaccine Candidates for Plasmodium falciparum

 , ,
, ,
Abstract
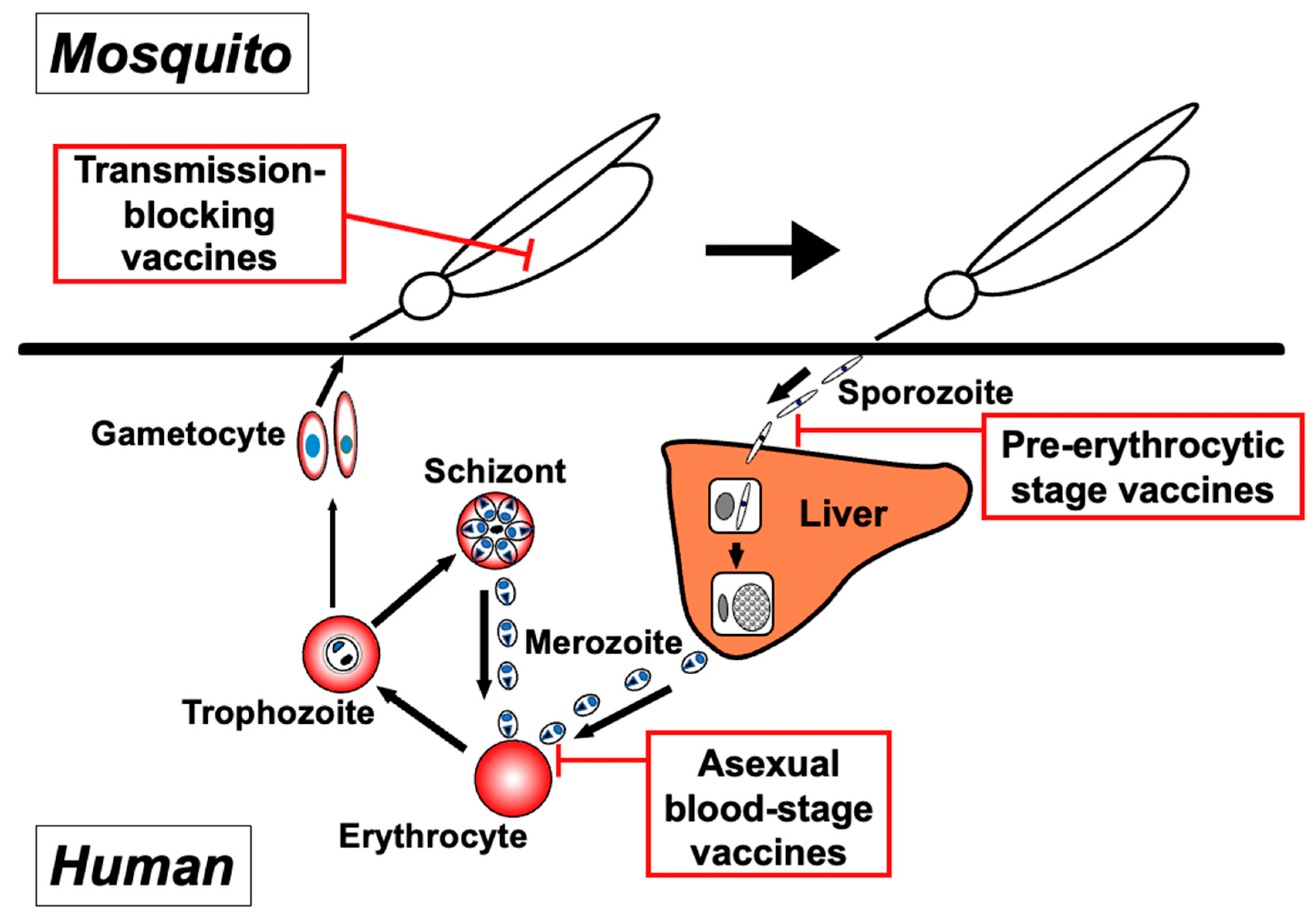
1. Introduction
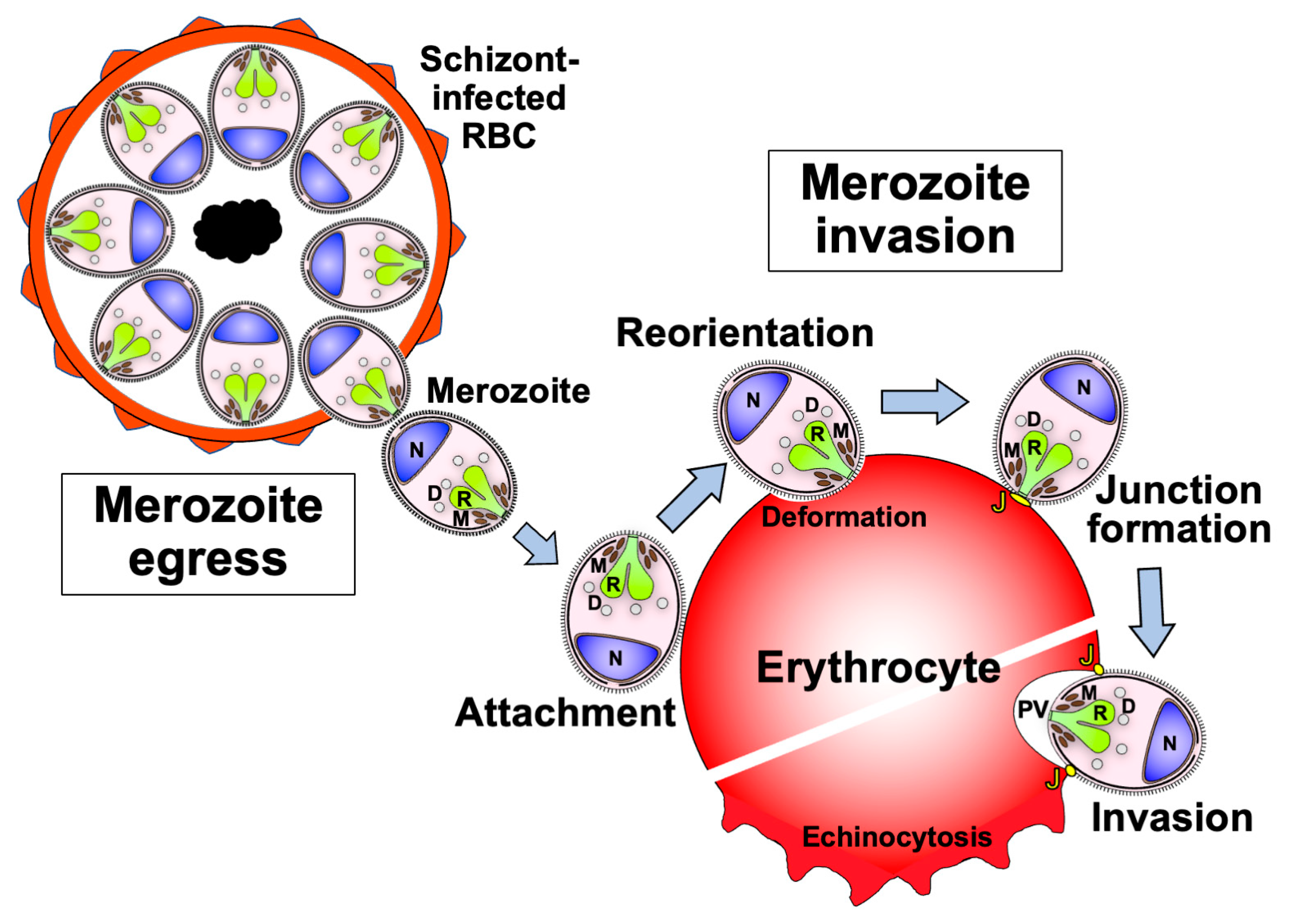
2. Complex Processes of the Asexual Blood-Stage Malaria Lifecycle and the Molecules Involved in These Steps
3. Discovery of Asexual Blood-Stage Malaria Vaccine Candidate Molecules
4. Current Status of Leading Asexual Blood-Stage Malaria Vaccine Candidates
4.1. AMA1
4.2. MSP1
4.3. MSP2 (Combination B Vaccine)
4.4. MSP3 and GMZ2
4.5. EBA175
4.6. SERA5 (BK-SE36 Vaccine)
4.7. Rh5
4.8. Ripr
4.9. CyRPA
4.10. P27A
4.11. PfSEA-1
4.12. PfGARP
4.13. Others
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2023; WHO Press: Geneva, Switzerland, 2023. [Google Scholar]
- Malaria Vaccine Funders Group. Malaria Vaccine Technology Roadmap; WHO Press: Geneva, Switzerland, 2013. [Google Scholar]
- Moorthy, V.S.; Newman, R.D.; Okwo-Bele, J.M. Malaria vaccine technology roadmap. Lancet 2013, 382, 1700–1701. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.M.; McGovern, T.W.; Krzych, U.; Cohen, J.C.; Schneider, I.; LaChance, R.; Heppner, D.G.; Yuan, G.; Hollingdale, M.; Slaoui, M.; et al. Safety, immunogenicity, and efficacy of a recombinantly produced Plasmodium falciparum circumsporozoite protein-hepatitis B surface antigen subunit vaccine. J. Infect. Dis. 1995, 171, 1576–1585. [Google Scholar] [CrossRef]
- Cohen, J.; Nussenzweig, V.; Nussenzweig, R.; Vekemans, J.; Leach, A. From the circumsporozoite protein to the RTS, S/AS candidate vaccine. Hum. Vaccines 2010, 6, 90–96. [Google Scholar] [CrossRef] [PubMed]
- RTS, S.C.T.P. Efficacy and safety of RTS, S/AS01 malaria vaccine with or without a booster dose in infants and children in Africa: Final results of a phase 3, individually randomised, controlled trial. Lancet 2015, 386, 31–45. [Google Scholar] [CrossRef]
- White, M.T.; Verity, R.; Griffin, J.T.; Asante, K.P.; Owusu-Agyei, S.; Greenwood, B.; Drakeley, C.; Gesase, S.; Lusingu, J.; Ansong, D.; et al. Immunogenicity of the RTS, S/AS01 malaria vaccine and implications for duration of vaccine efficacy: Secondary analysis of data from a phase 3 randomised controlled trial. Lancet Infect. Dis. 2015, 15, 1450–1458. [Google Scholar] [CrossRef]
- Adepoju, P. RTS,S malaria vaccine pilots in three African countries. Lancet 2019, 393, 1685. [Google Scholar] [CrossRef]
- Vogel, G. WHO gives first malaria vaccine the green light. Science 2021, 374, 245–246. [Google Scholar] [CrossRef]
- WHO. World Malaria Report 2021; WHO Press: Geneva, Switzerland, 2021. [Google Scholar]
- Datoo, M.S.; Natama, M.H.; Some, A.; Traore, O.; Rouamba, T.; Bellamy, D.; Yameogo, P.; Valia, D.; Tegneri, M.; Ouedraogo, F.; et al. Efficacy of a low-dose candidate malaria vaccine, R21 in adjuvant Matrix-M, with seasonal administration to children in Burkina Faso: A randomised controlled trial. Lancet 2021, 397, 1809–1818. [Google Scholar] [CrossRef]
- Datoo, M.S.; Natama, H.M.; Some, A.; Bellamy, D.; Traore, O.; Rouamba, T.; Tahita, M.C.; Ido, N.F.A.; Yameogo, P.; Valia, D.; et al. Efficacy and immunogenicity of R21/Matrix-M vaccine against clinical malaria after 2 years’ follow-up in children in Burkina Faso: A phase 1/2b randomised controlled trial. Lancet Infect. Dis. 2022, 22, 1728–1736. [Google Scholar] [CrossRef]
- Collins, K.A.; Snaith, R.; Cottingham, M.G.; Gilbert, S.C.; Hill, A.V.S. Enhancing protective immunity to malaria with a highly immunogenic virus-like particle vaccine. Sci. Rep. 2017, 7, 46621. [Google Scholar] [CrossRef]
- Regules, J.A.; Cummings, J.F.; Ockenhouse, C.F. The RTS,S vaccine candidate for malaria. Expert Rev. Vaccines 2011, 10, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, T.; Takashima, E. Antibody titre as a surrogate of protection of the first malaria subunit vaccine, RTS, S/AS01. Lancet Infect. Dis. 2015, 15, 1371–1372. [Google Scholar] [CrossRef] [PubMed][Green Version]
- WHO. Malaria Vaccines: Preferred Product Characteristics and Clinical Development Considerations; WHO Press: Geneva, Switzerland, 2022. [Google Scholar]
- WHO. WHO Guidelines for Malaria; WHO Press: Geneva, Switzerland, 2022. [Google Scholar]
- Aikawa, M.; Miller, L.H.; Johnson, J.; Rabbege, J. Erythrocyte entry by malarial parasites. A moving junction between erythrocyte and parasite. J. Cell Biol. 1978, 77, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Richards, J.S.; Arumugam, T.U.; Reiling, L.; Healer, J.; Hodder, A.N.; Fowkes, F.J.; Cross, N.; Langer, C.; Takeo, S.; Uboldi, A.D.; et al. Identification and prioritization of merozoite antigens as targets of protective human immunity to Plasmodium falciparum malaria for vaccine and biomarker development. J. Immunol. 2013, 191, 795–809. [Google Scholar] [CrossRef]
- Osier, F.H.; Mackinnon, M.J.; Crosnier, C.; Fegan, G.; Kamuyu, G.; Wanaguru, M.; Ogada, E.; McDade, B.; Rayner, J.C.; Wright, G.J.; et al. New antigens for a multicomponent blood-stage malaria vaccine. Sci. Transl. Med. 2014, 6, 247ra102. [Google Scholar] [CrossRef]
- Beeson, J.G.; Drew, D.R.; Boyle, M.J.; Feng, G.; Fowkes, F.J.; Richards, J.S. Merozoite surface proteins in red blood cell invasion, immunity and vaccines against malaria. FEMS Microbiol. Rev. 2016, 40, 343–372. [Google Scholar] [CrossRef]
- Weiss, G.E.; Gilson, P.R.; Taechalertpaisarn, T.; Tham, W.H.; de Jong, N.W.; Harvey, K.L.; Fowkes, F.J.; Barlow, P.N.; Rayner, J.C.; Wright, G.J.; et al. Revealing the sequence and resulting cellular morphology of receptor-ligand interactions during Plasmodium falciparum invasion of erythrocytes. PLoS Pathog. 2015, 11, e1004670. [Google Scholar] [CrossRef] [PubMed]
- Yahata, K.; Hart, M.N.; Davies, H.; Asada, M.; Wassmer, S.C.; Templeton, T.J.; Treeck, M.; Moon, R.W.; Kaneko, O. Gliding motility of Plasmodium merozoites. Proc. Natl. Acad. Sci. USA 2021, 118, e2114442118. [Google Scholar] [CrossRef]
- Riglar, D.T.; Richard, D.; Wilson, D.W.; Boyle, M.J.; Dekiwadia, C.; Turnbull, L.; Angrisano, F.; Marapana, D.S.; Rogers, K.L.; Whitchurch, C.B.; et al. Super-resolution dissection of coordinated events during malaria parasite invasion of the human erythrocyte. Cell Host Microbe 2011, 9, 9–20. [Google Scholar] [CrossRef]
- Tham, W.H.; Healer, J.; Cowman, A.F. Erythrocyte and reticulocyte binding-like proteins of Plasmodium falciparum. Trends Parasitol. 2012, 28, 23–30. [Google Scholar] [CrossRef]
- Tham, W.H.; Kennedy, A.T. Malaria: A master lock for deadly parasites. Nature 2015, 522, 158–159. [Google Scholar] [CrossRef]
- Chen, L.; Lopaticki, S.; Riglar, D.T.; Dekiwadia, C.; Uboldi, A.D.; Tham, W.H.; O’Neill, M.T.; Richard, D.; Baum, J.; Ralph, S.A.; et al. An EGF-like protein forms a complex with PfRh5 and is required for invasion of human erythrocytes by Plasmodium falciparum. PLoS Pathog. 2011, 7, e1002199. [Google Scholar] [CrossRef]
- Reddy, K.S.; Amlabu, E.; Pandey, A.K.; Mitra, P.; Chauhan, V.S.; Gaur, D. Multiprotein complex between the GPI-anchored CyRPA with PfRH5 and PfRipr is crucial for Plasmodium falciparum erythrocyte invasion. Proc. Natl. Acad. Sci. USA 2015, 112, 1179–1184. [Google Scholar] [CrossRef]
- Crosnier, C.; Bustamante, L.Y.; Bartholdson, S.J.; Bei, A.K.; Theron, M.; Uchikawa, M.; Mboup, S.; Ndir, O.; Kwiatkowski, D.P.; Duraisingh, M.T.; et al. Basigin is a receptor essential for erythrocyte invasion by Plasmodium falciparum. Nature 2011, 480, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Scally, S.W.; Triglia, T.; Evelyn, C.; Seager, B.A.; Pasternak, M.; Lim, P.S.; Healer, J.; Geoghegan, N.D.; Adair, A.; Tham, W.H.; et al. PCRCR complex is essential for invasion of human erythrocytes by Plasmodium falciparum. Nat. Microbiol. 2022, 7, 2039–2053. [Google Scholar] [CrossRef]
- Farrell, B.; Alam, N.; Hart, M.N.; Jamwal, A.; Ragotte, R.J.; Walters-Morgan, H.; Draper, S.J.; Knuepfer, E.; Higgins, M.K. The PfRCR complex bridges malaria parasite and erythrocyte during invasion. Nature 2023. [Google Scholar] [CrossRef]
- Good, M.F.; Miller, L.H. Interpreting challenge data from early phase malaria blood stage vaccine trials. Expert Rev. Vaccines 2018, 17, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Freund, J.; Thomson, K.J.; Sommer, H.E.; Walter, A.W.; Pisani, T.M. Immunization of monkeys against malaria by means of killed parasites with adjuvants. Am. J. Trop. Med. Hyg. 1948, 28, 1–22. [Google Scholar] [CrossRef]
- McCarthy, J.S.; Good, M.F. Whole parasite blood stage malaria vaccines: A convergence of evidence. Hum. Vaccines 2010, 6, 114–123. [Google Scholar] [CrossRef]
- Coggeshall, L.T.; Kumm, H.W. Demonstration of Passive Immunity in Experimental Monkey Malaria. J. Exp. Med. 1937, 66, 177–190. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Trager, W.; Jensen, J.B. Human malaria parasites in continuous culture. Science 1976, 193, 673–675. [Google Scholar] [CrossRef]
- Mitchell, G.H.; Richards, W.H.; Butcher, G.A.; Cohen, S. Merozoite vaccination of douroucouli monkeys against falciparum malaria. Lancet 1977, 1, 1335–1338. [Google Scholar] [CrossRef]
- Siddiqui, W.A. An effective immunization of experimental monkeys against a human malaria parasite, Plasmodium falciparum. Science 1977, 197, 388–389. [Google Scholar] [CrossRef]
- Cohen, S.; Mc, G.I.; Carrington, S. Gamma-globulin and acquired immunity to human malaria. Nature 1961, 192, 733–737. [Google Scholar] [CrossRef]
- Bouharoun-Tayoun, H.; Attanath, P.; Sabchareon, A.; Chongsuphajaisiddhi, T.; Druilhe, P. Antibodies that protect humans against Plasmodium falciparum blood stages do not on their own inhibit parasite growth and invasion in vitro, but act in cooperation with monocytes. J. Exp. Med. 1990, 172, 1633–1641. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Ozaki, L.S.; Gwadz, R.W.; Cochrane, A.H.; Nussenzweig, V.; Nussenzweig, R.S.; Godson, G.N. Cloning and expression in E. coli of the malarial sporozoite surface antigen gene from Plasmodium knowlesi. Nature 1983, 302, 536–538. [Google Scholar] [CrossRef]
- Kemp, D.J.; Coppel, R.L.; Cowman, A.F.; Saint, R.B.; Brown, G.V.; Anders, R.F. Expression of Plasmodium falciparum blood-stage antigens in Escherichia coli: Detection with antibodies from immune humans. Proc. Natl. Acad. Sci. USA 1983, 80, 3787–3791. [Google Scholar] [CrossRef]
- Holder, A.A.; Lockyer, M.J.; Odink, K.G.; Sandhu, J.S.; Riveros-Moreno, V.; Nicholls, S.C.; Hillman, Y.; Davey, L.S.; Tizard, M.L.; Schwarz, R.T.; et al. Primary structure of the precursor to the three major surface antigens of Plasmodium falciparum merozoites. Nature 1985, 317, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Smythe, J.A.; Coppel, R.L.; Brown, G.V.; Ramasamy, R.; Kemp, D.J.; Anders, R.F. Identification of two integral membrane proteins of Plasmodium falciparum. Proc. Natl. Acad. Sci. USA 1988, 85, 5195–5199. [Google Scholar] [CrossRef]
- McColl, D.J.; Silva, A.; Foley, M.; Kun, J.F.; Favaloro, J.M.; Thompson, J.K.; Marshall, V.M.; Coppel, R.L.; Kemp, D.J.; Anders, R.F. Molecular variation in a novel polymorphic antigen associated with Plasmodium falciparum merozoites. Mol. Biochem. Parasitol. 1994, 68, 53–67. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.G.; Marshall, V.M.; Smythe, J.A.; Crewther, P.E.; Lew, A.; Silva, A.; Anders, R.F.; Kemp, D.J. Integral membrane protein located in the apical complex of Plasmodium falciparum. Mol. Cell Biol. 1989, 9, 3151–3154. [Google Scholar] [CrossRef] [PubMed]
- Horii, T.; Bzik, D.J.; Inselburg, J. Characterization of antigen-expressing Plasmodium falciparum cDNA clones that are reactive with parasite inhibitory antibodies. Mol. Biochem. Parasitol. 1988, 30, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Borre, M.B.; Dziegiel, M.; Hogh, B.; Petersen, E.; Rieneck, K.; Riley, E.; Meis, J.F.; Aikawa, M.; Nakamura, K.; Harada, M.; et al. Primary structure and localization of a conserved immunogenic Plasmodium falciparum glutamate rich protein (GLURP) expressed in both the preerythrocytic and erythrocytic stages of the vertebrate life cycle. Mol. Biochem. Parasitol. 1991, 49, 119–131. [Google Scholar] [CrossRef]
- Dziegiel, M.; Borre, M.B.; Jepsen, S.; Hogh, B.; Petersen, E.; Vuust, J. Recombinant Plasmodium falciparum glutamate rich protein; purification and use in enzyme-linked immunosorbent assay. Am. J. Trop. Med. Hyg. 1991, 44, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.H.; Hudson, D.E.; Torii, M.; Ward, G.E.; Wellems, T.E.; Aikawa, M.; Miller, L.H. The Duffy receptor family of Plasmodium knowlesi is located within the micronemes of invasive malaria merozoites. Cell 1990, 63, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.D.; Kaslow, D.C.; Adams, J.H.; Miller, L.H. Cloning of the Plasmodium vivax Duffy receptor. Mol. Biochem. Parasitol. 1991, 44, 125–132. [Google Scholar] [CrossRef]
- Sim, B.K.; Orlandi, P.A.; Haynes, J.D.; Klotz, F.W.; Carter, J.M.; Camus, D.; Zegans, M.E.; Chulay, J.D. Primary structure of the 175K Plasmodium falciparum erythrocyte binding antigen and identification of a peptide which elicits antibodies that inhibit malaria merozoite invasion. J. Cell Biol. 1990, 111, 1877–1884. [Google Scholar] [CrossRef]
- Camus, D.; Hadley, T.J. A Plasmodium falciparum antigen that binds to host erythrocytes and merozoites. Science 1985, 230, 553–556. [Google Scholar] [CrossRef]
- Orlandi, P.A.; Sim, B.K.; Chulay, J.D.; Haynes, J.D. Characterization of the 175-kilodalton erythrocyte binding antigen of Plasmodium falciparum. Mol. Biochem. Parasitol. 1990, 40, 285–294. [Google Scholar] [CrossRef]
- Adams, J.H.; Sim, B.K.; Dolan, S.A.; Fang, X.; Kaslow, D.C.; Miller, L.H. A family of erythrocyte binding proteins of malaria parasites. Proc. Natl. Acad. Sci. USA 1992, 89, 7085–7089. [Google Scholar] [CrossRef]
- Mayer, D.C.; Kaneko, O.; Hudson-Taylor, D.E.; Reid, M.E.; Miller, L.H. Characterization of a Plasmodium falciparum erythrocyte-binding protein paralogous to EBA-175. Proc. Natl. Acad. Sci. USA 2001, 98, 5222–5227. [Google Scholar] [CrossRef] [PubMed]
- Narum, D.L.; Fuhrmann, S.R.; Luu, T.; Sim, B.K. A novel Plasmodium falciparum erythrocyte binding protein-2 (EBP2/BAEBL) involved in erythrocyte receptor binding. Mol. Biochem. Parasitol. 2002, 119, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.K.; Triglia, T.; Reed, M.B.; Cowman, A.F. A novel ligand from Plasmodium falciparum that binds to a sialic acid-containing receptor on the surface of human erythrocytes. Mol. Microbiol. 2001, 41, 47–58. [Google Scholar] [CrossRef]
- Galinski, M.R.; Medina, C.C.; Ingravallo, P.; Barnwell, J.W. A reticulocyte-binding protein complex of Plasmodium vivax merozoites. Cell 1992, 69, 1213–1226. [Google Scholar] [CrossRef] [PubMed]
- Keen, J.K.; Sinha, K.A.; Brown, K.N.; Holder, A.A. A gene coding for a high-molecular mass rhoptry protein of Plasmodium yoelii. Mol. Biochem. Parasitol. 1994, 65, 171–177. [Google Scholar] [CrossRef]
- Ogun, S.A.; Holder, A.A. A high molecular mass Plasmodium yoelii rhoptry protein binds to erythrocytes. Mol. Biochem. Parasitol. 1996, 76, 321–324. [Google Scholar] [CrossRef]
- Rayner, J.C.; Vargas-Serrato, E.; Huber, C.S.; Galinski, M.R.; Barnwell, J.W. A Plasmodium falciparum homologue of Plasmodium vivax reticulocyte binding protein (PvRBP1) defines a trypsin-resistant erythrocyte invasion pathway. J. Exp. Med. 2001, 194, 1571–1581. [Google Scholar] [CrossRef][Green Version]
- Miller, L.H.; Baruch, D.I.; Marsh, K.; Doumbo, O.K. The pathogenic basis of malaria. Nature 2002, 415, 673–679. [Google Scholar] [CrossRef]
- Rayner, J.C.; Galinski, M.R.; Ingravallo, P.; Barnwell, J.W. Two Plasmodium falciparum genes express merozoite proteins that are related to Plasmodium vivax and Plasmodium yoelii adhesive proteins involved in host cell selection and invasion. Proc. Natl. Acad. Sci. USA 2000, 97, 9648–9653. [Google Scholar] [CrossRef]
- Triglia, T.; Thompson, J.; Caruana, S.R.; Delorenzi, M.; Speed, T.; Cowman, A.F. Identification of proteins from Plasmodium falciparum that are homologous to reticulocyte binding proteins in Plasmodium vivax. Infect. Immun. 2001, 69, 1084–1092. [Google Scholar] [CrossRef]
- Kaneko, O.; Mu, J.; Tsuboi, T.; Su, X.; Torii, M. Gene structure and expression of a Plasmodium falciparum 220-kDa protein homologous to the Plasmodium vivax reticulocyte binding proteins. Mol. Biochem. Parasitol. 2002, 121, 275–278. [Google Scholar] [CrossRef] [PubMed]
- Cowman, A.F.; Crabb, B.S. Invasion of red blood cells by malaria parasites. Cell 2006, 124, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Hayton, K.; Gaur, D.; Liu, A.; Takahashi, J.; Henschen, B.; Singh, S.; Lambert, L.; Furuya, T.; Bouttenot, R.; Doll, M.; et al. Erythrocyte binding protein PfRH5 polymorphisms determine species-specific pathways of Plasmodium falciparum invasion. Cell Host Microbe 2008, 4, 40–51. [Google Scholar] [CrossRef]
- Rodriguez, M.; Lustigman, S.; Montero, E.; Oksov, Y.; Lobo, C.A. PfRH5: A novel reticulocyte-binding family homolog of plasmodium falciparum that binds to the erythrocyte, and an investigation of its receptor. PLoS ONE 2008, 3, e3300. [Google Scholar] [CrossRef]
- Ragotte, R.J.; Higgins, M.K.; Draper, S.J. The RH5-CyRPA-Ripr Complex as a Malaria Vaccine Target. Trends Parasitol. 2020, 36, 545–559. [Google Scholar] [CrossRef]
- Draper, S.J.; Sack, B.K.; King, C.R.; Nielsen, C.M.; Rayner, J.C.; Higgins, M.K.; Long, C.A.; Seder, R.A. Malaria Vaccines: Recent Advances and New Horizons. Cell Host Microbe 2018, 24, 43–56. [Google Scholar] [CrossRef]
- Douglas, A.D.; Williams, A.R.; Illingworth, J.J.; Kamuyu, G.; Biswas, S.; Goodman, A.L.; Wyllie, D.H.; Crosnier, C.; Miura, K.; Wright, G.J.; et al. The blood-stage malaria antigen PfRH5 is susceptible to vaccine-inducible cross-strain neutralizing antibody. Nat. Commun. 2011, 2, 601. [Google Scholar] [CrossRef]
- Payne, R.O.; Silk, S.E.; Elias, S.C.; Milne, K.H.; Rawlinson, T.A.; Llewellyn, D.; Shakri, A.R.; Jin, J.; Labbe, G.M.; Edwards, N.J.; et al. Human vaccination against Plasmodium vivax Duffy-binding protein induces strain-transcending antibodies. JCI Insight 2017, 2, e93683. [Google Scholar] [CrossRef]
- Tran, T.M.; Ongoiba, A.; Coursen, J.; Crosnier, C.; Diouf, A.; Huang, C.Y.; Li, S.; Doumbo, S.; Doumtabe, D.; Kone, Y.; et al. Naturally acquired antibodies specific for Plasmodium falciparum reticulocyte-binding protein homologue 5 inhibit parasite growth and predict protection from malaria. J. Infect. Dis. 2014, 209, 789–798. [Google Scholar] [CrossRef]
- Villard, V.; Agak, G.W.; Frank, G.; Jafarshad, A.; Servis, C.; Nebie, I.; Sirima, S.B.; Felger, I.; Arevalo-Herrera, M.; Herrera, S.; et al. Rapid identification of malaria vaccine candidates based on alpha-helical coiled coil protein motif. PLoS ONE 2007, 2, e645. [Google Scholar] [CrossRef]
- Raj, D.K.; Nixon, C.P.; Nixon, C.E.; Dvorin, J.D.; DiPetrillo, C.G.; Pond-Tor, S.; Wu, H.W.; Jolly, G.; Pischel, L.; Lu, A.; et al. Antibodies to PfSEA-1 block parasite egress from RBCs and protect against malaria infection. Science 2014, 344, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Raj, D.K.; Das Mohapatra, A.; Jnawali, A.; Zuromski, J.; Jha, A.; Cham-Kpu, G.; Sherman, B.; Rudlaff, R.M.; Nixon, C.E.; Hilton, N.; et al. Anti-PfGARP activates programmed cell death of parasites and reduces severe malaria. Nature 2020, 582, 104–108. [Google Scholar] [CrossRef]
- Duffy, P.E. Current approaches to malaria vaccines. Curr. Opin. Microbiol. 2022, 70, 102227. [Google Scholar] [CrossRef] [PubMed]
- Ntege, E.H.; Takashima, E.; Morita, M.; Nagaoka, H.; Ishino, T.; Tsuboi, T. Blood-stage malaria vaccines: Post-genome strategies for the identification of novel vaccine candidates. Expert Rev. Vaccines 2017, 16, 769–779. [Google Scholar] [CrossRef] [PubMed]
- Thera, M.A.; Doumbo, O.K.; Coulibaly, D.; Laurens, M.B.; Ouattara, A.; Kone, A.K.; Guindo, A.B.; Traore, K.; Traore, I.; Kouriba, B.; et al. A field trial to assess a blood-stage malaria vaccine. N. Engl. J. Med. 2011, 365, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Laurens, M.B.; Thera, M.A.; Coulibaly, D.; Ouattara, A.; Kone, A.K.; Guindo, A.B.; Traore, K.; Traore, I.; Kouriba, B.; Diallo, D.A.; et al. Extended safety, immunogenicity and efficacy of a blood-stage malaria vaccine in malian children: 24-month follow-up of a randomized, double-blinded phase 2 trial. PLoS ONE 2013, 8, e79323. [Google Scholar] [CrossRef]
- Bai, T.; Becker, M.; Gupta, A.; Strike, P.; Murphy, V.J.; Anders, R.F.; Batchelor, A.H. Structure of AMA1 from Plasmodium falciparum reveals a clustering of polymorphisms that surround a conserved hydrophobic pocket. Proc. Natl. Acad. Sci. USA 2005, 102, 12736–12741. [Google Scholar] [CrossRef]
- Ogutu, B.R.; Apollo, O.J.; McKinney, D.; Okoth, W.; Siangla, J.; Dubovsky, F.; Tucker, K.; Waitumbi, J.N.; Diggs, C.; Wittes, J.; et al. Blood stage malaria vaccine eliciting high antigen-specific antibody concentrations confers no protection to young children in Western Kenya. PLoS ONE 2009, 4, e4708. [Google Scholar] [CrossRef]
- Morgan, W.D.; Birdsall, B.; Frenkiel, T.A.; Gradwell, M.G.; Burghaus, P.A.; Syed, S.E.; Uthaipibull, C.; Holder, A.A.; Feeney, J. Solution structure of an EGF module pair from the Plasmodium falciparum merozoite surface protein 1. J. Mol. Biol. 1999, 289, 113–122. [Google Scholar] [CrossRef]
- Genton, B.; Betuela, I.; Felger, I.; Al-Yaman, F.; Anders, R.F.; Saul, A.; Rare, L.; Baisor, M.; Lorry, K.; Brown, G.V.; et al. A recombinant blood-stage malaria vaccine reduces Plasmodium falciparum density and exerts selective pressure on parasite populations in a phase 1-2b trial in Papua New Guinea. J. Infect. Dis. 2002, 185, 820–827. [Google Scholar] [CrossRef]
- Sirima, S.B.; Mordmuller, B.; Milligan, P.; Ngoa, U.A.; Kironde, F.; Atuguba, F.; Tiono, A.B.; Issifou, S.; Kaddumukasa, M.; Bangre, O.; et al. A phase 2b randomized, controlled trial of the efficacy of the GMZ2 malaria vaccine in African children. Vaccine 2016, 34, 4536–4542. [Google Scholar] [CrossRef] [PubMed]
- Koram, K.A.; Adu, B.; Ocran, J.; Karikari, Y.S.; Adu-Amankwah, S.; Ntiri, M.; Abuaku, B.; Dodoo, D.; Gyan, B.; Kronmann, K.C.; et al. Safety and Immunogenicity of EBA-175 RII-NG Malaria Vaccine Administered Intramuscularly in Semi-Immune Adults: A Phase 1, Double-Blinded Placebo Controlled Dosage Escalation Study. PLoS ONE 2016, 11, e0163066. [Google Scholar] [CrossRef] [PubMed]
- Tolia, N.H.; Enemark, E.J.; Sim, B.K.; Joshua-Tor, L. Structural basis for the EBA-175 erythrocyte invasion pathway of the malaria parasite Plasmodium falciparum. Cell 2005, 122, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Ouedraogo, A.; Bougouma, E.C.; Palacpac, N.M.Q.; Houard, S.; Nebie, I.; Sawadogo, J.; Berges, G.D.; Soulama, I.; Diarra, A.; Hien, D.; et al. Safety and immunogenicity of BK-SE36/CpG malaria vaccine in healthy Burkinabe adults and children: A phase 1b randomised, controlled, double-blinded, age de-escalation trial. Front. Immunol. 2023, 14, 1267372. [Google Scholar] [CrossRef]
- Hodder, A.N.; Malby, R.L.; Clarke, O.B.; Fairlie, W.D.; Colman, P.M.; Crabb, B.S.; Smith, B.J. Structural insights into the protease-like antigen Plasmodium falciparum SERA5 and its noncanonical active-site serine. J. Mol. Biol. 2009, 392, 154–165. [Google Scholar] [CrossRef]
- Minassian, A.M.; Silk, S.E.; Barrett, J.R.; Nielsen, C.M.; Miura, K.; Diouf, A.; Loos, C.; Fallon, J.K.; Michell, A.R.; White, M.T.; et al. Reduced blood-stage malaria growth and immune correlates in humans following RH5 vaccination. Med 2021, 2, 701–719.e719. [Google Scholar] [CrossRef]
- Wright, K.E.; Hjerrild, K.A.; Bartlett, J.; Douglas, A.D.; Jin, J.; Brown, R.E.; Illingworth, J.J.; Ashfield, R.; Clemmensen, S.B.; de Jongh, W.A.; et al. Structure of malaria invasion protein RH5 with erythrocyte basigin and blocking antibodies. Nature 2014, 515, 427–430. [Google Scholar] [CrossRef]
- Takashima, E.; Nagaoka, H.; Correia, R.; Alves, P.M.; Roldao, A.; Christensen, D.; Guderian, J.A.; Fukushima, A.; Viebig, N.K.; Depraetere, H.; et al. A novel asexual blood-stage malaria vaccine candidate: PfRipr5 formulated with human-use adjuvants induces potent growth inhibitory antibodies. Front. Immunol. 2022, 13, 1002430. [Google Scholar] [CrossRef]
- Wong, W.; Huang, R.; Menant, S.; Hong, C.; Sandow, J.J.; Birkinshaw, R.W.; Healer, J.; Hodder, A.N.; Kanjee, U.; Tonkin, C.J.; et al. Structure of Plasmodium falciparum Rh5-CyRPA-Ripr invasion complex. Nature 2019, 565, 118–121. [Google Scholar] [CrossRef]
- Tamborrini, M.; Schafer, A.; Hauser, J.; Zou, L.; Paris, D.H.; Pluschke, G. The malaria blood stage antigen PfCyRPA formulated with the TLR-4 agonist adjuvant GLA-SE elicits parasite growth inhibitory antibodies in experimental animals. Malar. J. 2023, 22, 210. [Google Scholar] [CrossRef]
- Favuzza, P.; Guffart, E.; Tamborrini, M.; Scherer, B.; Dreyer, A.M.; Rufer, A.C.; Erny, J.; Hoernschemeyer, J.; Thoma, R.; Schmid, G.; et al. Structure of the malaria vaccine candidate antigen CyRPA and its complex with a parasite invasion inhibitory antibody. eLife 2017, 6, e20383. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Xu, Y.; Wong, W.; Thompson, J.K.; Healer, J.; Goddard-Borger, E.D.; Lawrence, M.C.; Cowman, A.F. Structural basis for inhibition of erythrocyte invasion by antibodies to Plasmodium falciparum protein CyRPA. eLife 2017, 6, e21347. [Google Scholar] [CrossRef] [PubMed]
- Steiner-Monard, V.; Kamaka, K.; Karoui, O.; Roethlisberger, S.; Audran, R.; Daubenberger, C.; Fayet-Mello, A.; Erdmann-Voisin, A.; Felger, I.; Geiger, K.; et al. The Candidate Blood-stage Malaria Vaccine P27A Induces a Robust Humoral Response in a Fast Track to the Field Phase 1 Trial in Exposed and Nonexposed Volunteers. Clin. Infect. Dis. 2019, 68, 466–474. [Google Scholar] [CrossRef] [PubMed]
- Kurtis, J.D.; Raj, D.K.; Michelow, I.C.; Park, S.; Nixon, C.E.; McDonald, E.A.; Nixon, C.P.; Pond-Tor, S.; Jha, A.; Taliano, R.J.; et al. Maternally-derived Antibodies to Schizont Egress Antigen-1 and Protection of Infants From Severe Malaria. Clin. Infect. Dis. 2019, 68, 1718–1724. [Google Scholar] [CrossRef]
- Lamarque, M.H.; Roques, M.; Kong-Hap, M.; Tonkin, M.L.; Rugarabamu, G.; Marq, J.B.; Penarete-Vargas, D.M.; Boulanger, M.J.; Soldati-Favre, D.; Lebrun, M. Plasticity and redundancy among AMA-RON pairs ensure host cell entry of Toxoplasma parasites. Nat. Commun. 2014, 5, 4098. [Google Scholar] [CrossRef] [PubMed]
- Yap, A.; Azevedo, M.F.; Gilson, P.R.; Weiss, G.E.; O’Neill, M.T.; Wilson, D.W.; Crabb, B.S.; Cowman, A.F. Conditional expression of apical membrane antigen 1 in Plasmodium falciparum shows it is required for erythrocyte invasion by merozoites. Cell Microbiol. 2014, 16, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Sagara, I.; Dicko, A.; Ellis, R.D.; Fay, M.P.; Diawara, S.I.; Assadou, M.H.; Sissoko, M.S.; Kone, M.; Diallo, A.I.; Saye, R.; et al. A randomized controlled phase 2 trial of the blood stage AMA1-C1/Alhydrogel malaria vaccine in children in Mali. Vaccine 2009, 27, 3090–3098. [Google Scholar] [CrossRef]
- Ouattara, A.; Mu, J.; Takala-Harrison, S.; Saye, R.; Sagara, I.; Dicko, A.; Niangaly, A.; Duan, J.; Ellis, R.D.; Miller, L.H.; et al. Lack of allele-specific efficacy of a bivalent AMA1 malaria vaccine. Malar. J. 2010, 9, 175. [Google Scholar] [CrossRef]
- Remarque, E.J.; Faber, B.W.; Kocken, C.H.; Thomas, A.W. A diversity-covering approach to immunization with Plasmodium falciparum apical membrane antigen 1 induces broader allelic recognition and growth inhibition responses in rabbits. Infect. Immun. 2008, 76, 2660–2670. [Google Scholar] [CrossRef]
- Kusi, K.A.; Remarque, E.J.; Riasat, V.; Walraven, V.; Thomas, A.W.; Faber, B.W.; Kocken, C.H. Safety and immunogenicity of multi-antigen AMA1-based vaccines formulated with CoVaccine HT and Montanide ISA 51 in rhesus macaques. Malar. J. 2011, 10, 182. [Google Scholar] [CrossRef]
- Dutta, S.; Dlugosz, L.S.; Drew, D.R.; Ge, X.; Ababacar, D.; Rovira, Y.I.; Moch, J.K.; Shi, M.; Long, C.A.; Foley, M.; et al. Overcoming antigenic diversity by enhancing the immunogenicity of conserved epitopes on the malaria vaccine candidate apical membrane antigen-1. PLoS Pathog. 2013, 9, e1003840. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, P.; Ekanem, E.; Diouf, A.; Tonkin, M.L.; Miura, K.; Boulanger, M.J.; Long, C.A.; Narum, D.L.; Miller, L.H. Immunization with a functional protein complex required for erythrocyte invasion protects against lethal malaria. Proc. Natl. Acad. Sci. USA 2014, 111, 10311–10316. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, P.; Baldeviano, G.C.; Miura, K.; Diouf, A.; Ventocilla, J.A.; Leiva, K.P.; Lugo-Roman, L.; Lucas, C.; Orr-Gonzalez, S.; Zhu, D.; et al. A malaria vaccine protects Aotus monkeys against virulent Plasmodium falciparum infection. NPJ Vaccines 2017, 2, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Dai, W.; Srinivasan, P.; McClellan, H.; Braden, D.; Allee-Munoz, A.; Hurtado, P.A.G.; Miller, L.H.; Duffy, P.E. Characterization of AMA1-RON2L complex with native gel electrophoresis and capillary isoelectric focusing. Electrophoresis 2022, 43, 509–515. [Google Scholar] [CrossRef]
- Patel, P.N.; Dickey, T.H.; Diouf, A.; Salinas, N.D.; McAleese, H.; Ouahes, T.; Long, C.A.; Miura, K.; Lambert, L.E.; Tolia, N.H. Structure-based design of a strain transcending AMA1-RON2L malaria vaccine. Nat. Commun. 2023, 14, 5345. [Google Scholar] [CrossRef] [PubMed]
- Boyle, M.J.; Reiling, L.; Feng, G.; Langer, C.; Osier, F.H.; Aspeling-Jones, H.; Cheng, Y.S.; Stubbs, J.; Tetteh, K.K.; Conway, D.J.; et al. Human antibodies fix complement to inhibit Plasmodium falciparum invasion of erythrocytes and are associated with protection against malaria. Immunity 2015, 42, 580–590. [Google Scholar] [CrossRef]
- Douglas, A.D.; Baldeviano, G.C.; Lucas, C.M.; Lugo-Roman, L.A.; Crosnier, C.; Bartholdson, S.J.; Diouf, A.; Miura, K.; Lambert, L.E.; Ventocilla, J.A.; et al. A PfRH5-based vaccine is efficacious against Heterologous strain blood-stage Plasmodium falciparum infection in aotus monkeys. Cell Host Microbe 2015, 17, 130–139. [Google Scholar] [CrossRef]
- Das, S.; Hertrich, N.; Perrin, A.J.; Withers-Martinez, C.; Collins, C.R.; Jones, M.L.; Watermeyer, J.M.; Fobes, E.T.; Martin, S.R.; Saibil, H.R.; et al. Processing of Plasmodium falciparum Merozoite Surface Protein MSP1 Activates a Spectrin-Binding Function Enabling Parasite Egress from RBCs. Cell Host Microbe 2015, 18, 433–444. [Google Scholar] [CrossRef]
- Lin, C.S.; Uboldi, A.D.; Epp, C.; Bujard, H.; Tsuboi, T.; Czabotar, P.E.; Cowman, A.F. Multiple Plasmodium falciparum Merozoite Surface Protein 1 Complexes Mediate Merozoite Binding to Human Erythrocytes. J. Biol. Chem. 2016, 291, 7703–7715. [Google Scholar] [CrossRef]
- Kumar, S.; Yadava, A.; Keister, D.B.; Tian, J.H.; Ohl, M.; Perdue-Greenfield, K.A.; Miller, L.H.; Kaslow, D.C. Immunogenicity and in vivo efficacy of recombinant Plasmodium falciparum merozoite surface protein-1 in Aotus monkeys. Mol. Med. 1995, 1, 325–332. [Google Scholar] [CrossRef]
- Blank, A.; Furle, K.; Jaschke, A.; Mikus, G.; Lehmann, M.; Husing, J.; Heiss, K.; Giese, T.; Carter, D.; Bohnlein, E.; et al. Immunization with full-length Plasmodium falciparum merozoite surface protein 1 is safe and elicits functional cytophilic antibodies in a randomized first-in-human trial. NPJ Vaccines 2020, 5, 10. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, M.; Furle, K.; Hibbert, J.; Ulmer, A.; Ali, A.; Giese, T.; Blank, A.; Haefeli, W.E.; Bohnlein, E.; Lanzer, M.; et al. Multifunctional IgG/IgM antibodies and cellular cytotoxicity are elicited by the full-length MSP1 SumayaVac-1 malaria vaccine. NPJ Vaccines 2023, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Smythe, J.A.; Coppel, R.L.; Day, K.P.; Martin, R.K.; Oduola, A.M.; Kemp, D.J.; Anders, R.F. Structural diversity in the Plasmodium falciparum merozoite surface antigen 2. Proc. Natl. Acad. Sci. USA 1991, 88, 1751–1755. [Google Scholar] [CrossRef] [PubMed]
- Fenton, B.; Clark, J.T.; Khan, C.M.; Robinson, J.V.; Walliker, D.; Ridley, R.; Scaife, J.G.; McBride, J.S. Structural and antigenic polymorphism of the 35- to 48-kilodalton merozoite surface antigen (MSA-2) of the malaria parasite Plasmodium falciparum. Mol. Cell Biol. 1991, 11, 963–971. [Google Scholar] [CrossRef]
- Zhang, X.; Perugini, M.A.; Yao, S.; Adda, C.G.; Murphy, V.J.; Low, A.; Anders, R.F.; Norton, R.S. Solution conformation, backbone dynamics and lipid interactions of the intrinsically unstructured malaria surface protein MSP2. J. Mol. Biol. 2008, 379, 105–121. [Google Scholar] [CrossRef]
- McCarthy, J.S.; Marjason, J.; Elliott, S.; Fahey, P.; Bang, G.; Malkin, E.; Tierney, E.; Aked-Hurditch, H.; Adda, C.; Cross, N.; et al. A phase 1 trial of MSP2-C1, a blood-stage malaria vaccine containing 2 isoforms of MSP2 formulated with Montanide(R) ISA 720. PLoS ONE 2011, 6, e24413. [Google Scholar] [CrossRef]
- Krishnarjuna, B.; Andrew, D.; MacRaild, C.A.; Morales, R.A.; Beeson, J.G.; Anders, R.F.; Richards, J.S.; Norton, R.S. Strain-transcending immune response generated by chimeras of the malaria vaccine candidate merozoite surface protein 2. Sci. Rep. 2016, 6, 20613. [Google Scholar] [CrossRef]
- Adda, C.G.; MacRaild, C.A.; Reiling, L.; Wycherley, K.; Boyle, M.J.; Kienzle, V.; Masendycz, P.; Foley, M.; Beeson, J.G.; Norton, R.S.; et al. Antigenic characterization of an intrinsically unstructured protein, Plasmodium falciparum merozoite surface protein 2. Infect. Immun. 2012, 80, 4177–4185. [Google Scholar] [CrossRef]
- Seow, J.; Das, S.C.; Morales, R.A.V.; Ataide, R.; Krishnarjuna, B.; Silk, M.; Chalmers, D.K.; Richards, J.; Anders, R.F.; MacRaild, C.A.; et al. Guiding the Immune Response to a Conserved Epitope in MSP2, an Intrinsically Disordered Malaria Vaccine Candidate. Vaccines 2021, 9, 855. [Google Scholar] [CrossRef]
- Huber, W.; Felger, I.; Matile, H.; Lipps, H.J.; Steiger, S.; Beck, H.P. Limited sequence polymorphism in the Plasmodium falciparum merozoite surface protein 3. Mol. Biochem. Parasitol. 1997, 87, 231–234. [Google Scholar] [CrossRef]
- Singh, S.; Soe, S.; Mejia, J.P.; Roussilhon, C.; Theisen, M.; Corradin, G.; Druilhe, P. Identification of a conserved region of Plasmodium falciparum MSP3 targeted by biologically active antibodies to improve vaccine design. J. Infect. Dis. 2004, 190, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Druilhe, P.; Spertini, F.; Soesoe, D.; Corradin, G.; Mejia, P.; Singh, S.; Audran, R.; Bouzidi, A.; Oeuvray, C.; Roussilhon, C. A malaria vaccine that elicits in humans antibodies able to kill Plasmodium falciparum. PLoS Med. 2005, 2, e344. [Google Scholar] [CrossRef]
- Sirima, S.B.; Cousens, S.; Druilhe, P. Protection against malaria by MSP3 candidate vaccine. N. Engl. J. Med. 2011, 365, 1062–1064. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, M.P.; Jogdand, P.S.; Singh, S.K.; Esen, M.; Christiansen, M.; Issifou, S.; Hounkpatin, A.B.; Ateba-Ngoa, U.; Kremsner, P.G.; Dziegiel, M.H.; et al. The malaria vaccine candidate GMZ2 elicits functional antibodies in individuals from malaria endemic and non-endemic areas. J. Infect. Dis. 2013, 208, 479–488. [Google Scholar] [CrossRef]
- Mordmuller, B.; Szywon, K.; Greutelaers, B.; Esen, M.; Mewono, L.; Treut, C.; Murbeth, R.E.; Chilengi, R.; Noor, R.; Kilama, W.L.; et al. Safety and immunogenicity of the malaria vaccine candidate GMZ2 in malaria-exposed, adult individuals from Lambarene, Gabon. Vaccine 2010, 28, 6698–6703. [Google Scholar] [CrossRef]
- Belard, S.; Issifou, S.; Hounkpatin, A.B.; Schaumburg, F.; Ngoa, U.A.; Esen, M.; Fendel, R.; de Salazar, P.M.; Murbeth, R.E.; Milligan, P.; et al. A randomized controlled phase Ib trial of the malaria vaccine candidate GMZ2 in African children. PLoS ONE 2011, 6, e22525. [Google Scholar] [CrossRef] [PubMed]
- Nouatin, O.; Ibanez, J.; Fendel, R.; Ngoa, U.A.; Lorenz, F.R.; Dejon-Agobe, J.C.; Edoa, J.R.; Flugge, J.; Bruckner, S.; Esen, M.; et al. Cellular and antibody response in GMZ2-vaccinated Gabonese volunteers in a controlled human malaria infection trial. Malar. J. 2022, 21, 191. [Google Scholar] [CrossRef]
- Alves, K.C.S.; Guimaraes, J.M.; Almeida, M.E.M.; Mariuba, L.A.M. Plasmodium falciparum merozoite surface protein 3 as a vaccine candidate: A brief review. Rev. Inst. Med. Trop. Sao Paulo 2022, 64, e23. [Google Scholar] [CrossRef]
- Sim, B.K.; Chitnis, C.E.; Wasniowska, K.; Hadley, T.J.; Miller, L.H. Receptor and ligand domains for invasion of erythrocytes by Plasmodium falciparum. Science 1994, 264, 1941–1944. [Google Scholar] [CrossRef]
- El Sahly, H.M.; Patel, S.M.; Atmar, R.L.; Lanford, T.A.; Dube, T.; Thompson, D.; Sim, B.K.; Long, C.; Keitel, W.A. Safety and immunogenicity of a recombinant nonglycosylated erythrocyte binding antigen 175 Region II malaria vaccine in healthy adults living in an area where malaria is not endemic. Clin. Vaccine Immunol. 2010, 17, 1552–1559. [Google Scholar] [CrossRef]
- Bei, A.K.; Membi, C.D.; Rayner, J.C.; Mubi, M.; Ngasala, B.; Sultan, A.A.; Premji, Z.; Duraisingh, M.T. Variant merozoite protein expression is associated with erythrocyte invasion phenotypes in Plasmodium falciparum isolates from Tanzania. Mol. Biochem. Parasitol. 2007, 153, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mitamura, T.; Fox, B.A.; Bzik, D.J.; Horii, T. Differential localization of processed fragments of Plasmodium falciparum serine repeat antigen and further processing of its N-terminal 47 kDa fragment. Parasitol. Int. 2002, 51, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Aoki, S.; Li, J.; Itagaki, S.; Okech, B.A.; Egwang, T.G.; Matsuoka, H.; Palacpac, N.M.; Mitamura, T.; Horii, T. Serine repeat antigen (SERA5) is predominantly expressed among the SERA multigene family of Plasmodium falciparum, and the acquired antibody titers correlate with serum inhibition of the parasite growth. J. Biol. Chem. 2002, 277, 47533–47540. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.K.; Good, R.T.; Drew, D.R.; Delorenzi, M.; Sanders, P.R.; Hodder, A.N.; Speed, T.P.; Cowman, A.F.; de Koning-Ward, T.F.; Crabb, B.S. A subset of Plasmodium falciparum SERA genes are expressed and appear to play an important role in the erythrocytic cycle. J. Biol. Chem. 2002, 277, 47524–47532. [Google Scholar] [CrossRef]
- McCoubrie, J.E.; Miller, S.K.; Sargeant, T.; Good, R.T.; Hodder, A.N.; Speed, T.P.; de Koning-Ward, T.F.; Crabb, B.S. Evidence for a common role for the serine-type Plasmodium falciparum serine repeat antigen proteases: Implications for vaccine and drug design. Infect. Immun. 2007, 75, 5565–5574. [Google Scholar] [CrossRef] [PubMed]
- Stallmach, R.; Kavishwar, M.; Withers-Martinez, C.; Hackett, F.; Collins, C.R.; Howell, S.A.; Yeoh, S.; Knuepfer, E.; Atid, A.J.; Holder, A.A.; et al. Plasmodium falciparum SERA5 plays a non-enzymatic role in the malarial asexual blood-stage lifecycle. Mol. Microbiol. 2015, 96, 368–387. [Google Scholar] [CrossRef] [PubMed]
- Palacpac, N.M.; Arisue, N.; Tougan, T.; Ishii, K.J.; Horii, T. Plasmodium falciparum serine repeat antigen 5 (SE36) as a malaria vaccine candidate. Vaccine 2011, 29, 5837–5845. [Google Scholar] [CrossRef]
- Tanabe, K.; Arisue, N.; Palacpac, N.M.; Yagi, M.; Tougan, T.; Honma, H.; Ferreira, M.U.; Farnert, A.; Bjorkman, A.; Kaneko, A.; et al. Geographic differentiation of polymorphism in the Plasmodium falciparum malaria vaccine candidate gene SERA5. Vaccine 2012, 30, 1583–1593. [Google Scholar] [CrossRef]
- Arisue, N.; Palacpac, N.M.Q.; Ntege, E.H.; Yeka, A.; Balikagala, B.; Kanoi, B.N.; Bougouma, E.C.; Tiono, A.B.; Nebie, I.; Diarra, A.; et al. African-specific polymorphisms in Plasmodium falciparum serine repeat antigen 5 in Uganda and Burkina Faso clinical samples do not interfere with antibody response to BK-SE36 vaccination. Front. Cell Infect. Microbiol. 2022, 12, 1058081. [Google Scholar] [CrossRef]
- Yagi, M.; Bang, G.; Tougan, T.; Palacpac, N.M.; Arisue, N.; Aoshi, T.; Matsumoto, Y.; Ishii, K.J.; Egwang, T.G.; Druilhe, P.; et al. Protective epitopes of the Plasmodium falciparum SERA5 malaria vaccine reside in intrinsically unstructured N-terminal repetitive sequences. PLoS ONE 2014, 9, e98460. [Google Scholar] [CrossRef]
- Palacpac, N.M.; Ntege, E.; Yeka, A.; Balikagala, B.; Suzuki, N.; Shirai, H.; Yagi, M.; Ito, K.; Fukushima, W.; Hirota, Y.; et al. Phase 1b randomized trial and follow-up study in Uganda of the blood-stage malaria vaccine candidate BK-SE36. PLoS ONE 2013, 8, e64073. [Google Scholar] [CrossRef] [PubMed]
- Yagi, M.; Palacpac, N.M.; Ito, K.; Oishi, Y.; Itagaki, S.; Balikagala, B.; Ntege, E.H.; Yeka, A.; Kanoi, B.N.; Katuro, O.; et al. Antibody titres and boosting after natural malaria infection in BK-SE36 vaccine responders during a follow-up study in Uganda. Sci. Rep. 2016, 6, 34363. [Google Scholar] [CrossRef]
- Bougouma, E.C.; Palacpac, N.M.Q.; Tiono, A.B.; Nebie, I.; Ouedraogo, A.; Houard, S.; Yagi, M.; Coulibaly, S.A.; Diarra, A.; Tougan, T.; et al. Safety and immunogenicity of BK-SE36 in a blinded, randomized, controlled, age de-escalating phase Ib clinical trial in Burkinabe children. Front. Immunol. 2022, 13, 978591. [Google Scholar] [CrossRef] [PubMed]
- Ezoe, S.; Palacpac, N.M.Q.; Tetsutani, K.; Yamamoto, K.; Okada, K.; Taira, M.; Nishida, S.; Hirata, H.; Ogata, A.; Yamada, T.; et al. First-in-human randomised trial and follow-up study of Plasmodium falciparum blood-stage malaria vaccine BK-SE36 with CpG-ODN(K3). Vaccine 2020, 38, 7246–7257. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.D.; Ahouidi, A.D.; Bei, A.K.; Dieye, T.N.; Mboup, S.; Harrison, S.C.; Duraisingh, M.T. Plasmodium falciparum merozoite surface antigen, PfRH5, elicits detectable levels of invasion-inhibiting antibodies in humans. J. Infect. Dis. 2013, 208, 1679–1687. [Google Scholar] [CrossRef]
- Williams, A.R.; Douglas, A.D.; Miura, K.; Illingworth, J.J.; Choudhary, P.; Murungi, L.M.; Furze, J.M.; Diouf, A.; Miotto, O.; Crosnier, C.; et al. Enhancing blockade of Plasmodium falciparum erythrocyte invasion: Assessing combinations of antibodies against PfRH5 and other merozoite antigens. PLoS Pathog. 2012, 8, e1002991. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, L.Y.; Bartholdson, S.J.; Crosnier, C.; Campos, M.G.; Wanaguru, M.; Nguon, C.; Kwiatkowski, D.P.; Wright, G.J.; Rayner, J.C. A full-length recombinant Plasmodium falciparum PfRH5 protein induces inhibitory antibodies that are effective across common PfRH5 genetic variants. Vaccine 2013, 31, 373–379. [Google Scholar] [CrossRef]
- Reddy, K.S.; Pandey, A.K.; Singh, H.; Sahar, T.; Emmanuel, A.; Chitnis, C.E.; Chauhan, V.S.; Gaur, D. Bacterially expressed full-length recombinant Plasmodium falciparum RH5 protein binds erythrocytes and elicits potent strain-transcending parasite-neutralizing antibodies. Infect. Immun. 2014, 82, 152–164. [Google Scholar] [CrossRef]
- Payne, R.O.; Silk, S.E.; Elias, S.C.; Miura, K.; Diouf, A.; Galaway, F.; de Graaf, H.; Brendish, N.J.; Poulton, I.D.; Griffiths, O.J.; et al. Human vaccination against RH5 induces neutralizing antimalarial antibodies that inhibit RH5 invasion complex interactions. JCI Insight 2017, 2, e96381. [Google Scholar] [CrossRef]
- Campeotto, I.; Goldenzweig, A.; Davey, J.; Barfod, L.; Marshall, J.M.; Silk, S.E.; Wright, K.E.; Draper, S.J.; Higgins, M.K.; Fleishman, S.J. One-step design of a stable variant of the malaria invasion protein RH5 for use as a vaccine immunogen. Proc. Natl. Acad. Sci. USA 2017, 114, 998–1002. [Google Scholar] [CrossRef]
- Alanine, D.G.W.; Quinkert, D.; Kumarasingha, R.; Mehmood, S.; Donnellan, F.R.; Minkah, N.K.; Dadonaite, B.; Diouf, A.; Galaway, F.; Silk, S.E.; et al. Human Antibodies that Slow Erythrocyte Invasion Potentiate Malaria-Neutralizing Antibodies. Cell 2019, 178, 216–228.e221. [Google Scholar] [CrossRef] [PubMed]
- Healer, J.; Wong, W.; Thompson, J.K.; He, W.; Birkinshaw, R.W.; Miura, K.; Long, C.A.; Soroka, V.; Sogaard, T.M.M.; Jorgensen, T.; et al. Neutralising antibodies block the function of Rh5/Ripr/CyRPA complex during invasion of Plasmodium falciparum into human erythrocytes. Cell Microbiol. 2019, 21, e13030. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, T.; Takeo, S.; Iriko, H.; Jin, L.; Tsuchimochi, M.; Matsuda, S.; Han, E.T.; Otsuki, H.; Kaneko, O.; Sattabongkot, J.; et al. Wheat germ cell-free system-based production of malaria proteins for discovery of novel vaccine candidates. Infect. Immun. 2008, 76, 1702–1708. [Google Scholar] [CrossRef]
- Ntege, E.H.; Arisue, N.; Ito, D.; Hasegawa, T.; Palacpac, N.M.Q.; Egwang, T.G.; Horii, T.; Takashima, E.; Tsuboi, T. Identification of Plasmodium falciparum reticulocyte binding protein homologue 5-interacting protein, PfRipr, as a highly conserved blood-stage malaria vaccine candidate. Vaccine 2016, 34, 5612–5622. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, H.; Kanoi, B.N.; Ntege, E.H.; Aoki, M.; Fukushima, A.; Tsuboi, T.; Takashima, E. Antibodies against a short region of PfRipr inhibit Plasmodium falciparum merozoite invasion and PfRipr interaction with Rh5 and SEMA7A. Sci. Rep. 2020, 10, 6573. [Google Scholar] [CrossRef] [PubMed]
- Correia, R.; Fernandes, B.; Castro, R.; Nagaoka, H.; Takashima, E.; Tsuboi, T.; Fukushima, A.; Viebig, N.K.; Depraetere, H.; Alves, P.M.; et al. Asexual Blood-Stage Malaria Vaccine Candidate PfRipr5: Enhanced Production in Insect Cells. Front. Bioeng. Biotechnol. 2022, 10, 908509. [Google Scholar] [CrossRef]
- Valmaseda, A.; Macete, E.; Nhabomba, A.; Guinovart, C.; Aide, P.; Bardaji, A.; Bassat, Q.; Nhampossa, T.; Maculuve, S.; Casellas, A.; et al. Identifying Immune Correlates of Protection against Plasmodium falciparum Through a Novel Approach to Account for Heterogeneity in Malaria Exposure. Clin. Infect. Dis. 2018, 66, 586–593. [Google Scholar] [CrossRef]
- Mian, S.Y.; Somanathan, A.; Chaddha, K.; Pandey, A.K.; Singh, H.; Krishna, S.; Chaturvedi, N.; Uchoi, S.; Shukla, M.M.; Bharti, P.K.; et al. Plasmodium falciparum Cysteine-Rich Protective Antigen (CyRPA) Elicits Detectable Levels of Invasion-Inhibitory Antibodies during Natural Infection in Humans. Infect. Immun. 2022, 90, e0037721. [Google Scholar] [CrossRef]
- Tamborrini, M.; Hauser, J.; Schafer, A.; Amacker, M.; Favuzza, P.; Kyungtak, K.; Fleury, S.; Pluschke, G. Vaccination with virosomally formulated recombinant CyRPA elicits protective antibodies against Plasmodium falciparum parasites in preclinical in vitro and in vivo models. NPJ Vaccines 2020, 5, 9. [Google Scholar] [CrossRef]
- Somanathan, A.; Mian, S.Y.; Chaddha, K.; Uchoi, S.; Bharti, P.K.; Tandon, R.; Gaur, D.; Chauhan, V.S. Process development and preclinical evaluation of a major Plasmodium falciparum blood stage vaccine candidate, Cysteine-Rich Protective Antigen (CyRPA). Front. Immunol. 2022, 13, 1005332. [Google Scholar] [CrossRef]
- Fernandes, B.; Sousa, M.; Castro, R.; Schafer, A.; Hauser, J.; Schulze, K.; Amacker, M.; Tamborrini, M.; Pluschke, G.; Alves, P.M.; et al. Scalable Process for High-Yield Production of PfCyRPA Using Insect Cells for Inclusion in a Malaria Virosome-Based Vaccine Candidate. Front. Bioeng. Biotechnol. 2022, 10, 879078. [Google Scholar] [CrossRef] [PubMed]
- Olugbile, S.; Kulangara, C.; Bang, G.; Bertholet, S.; Suzarte, E.; Villard, V.; Frank, G.; Audran, R.; Razaname, A.; Nebie, I.; et al. Vaccine potentials of an intrinsically unstructured fragment derived from the blood stage-associated Plasmodium falciparum protein PFF0165c. Infect. Immun. 2009, 77, 5701–5709. [Google Scholar] [CrossRef] [PubMed]
- Kulangara, C.; Luedin, S.; Dietz, O.; Rusch, S.; Frank, G.; Mueller, D.; Moser, M.; Kajava, A.V.; Corradin, G.; Beck, H.P.; et al. Cell biological characterization of the malaria vaccine candidate trophozoite exported protein 1. PLoS ONE 2012, 7, e46112. [Google Scholar] [CrossRef]
- Almukadi, H.; Schwake, C.; Kaiser, M.M.; Mayer, D.C.G.; Schiemer, J.; Baldwin, M.R.; Hegde, S.; Lu, Y.; Hanada, T.; Chishti, A.H. Human erythrocyte band 3 is a host receptor for Plasmodium falciparum glutamic acid-rich protein. Blood 2019, 133, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Rojrung, R.; Kuamsab, N.; Putaporntip, C.; Jongwutiwes, S. Analysis of sequence diversity in Plasmodium falciparum glutamic acid-rich protein (PfGARP), an asexual blood stage vaccine candidate. Sci. Rep. 2023, 13, 3951. [Google Scholar] [CrossRef] [PubMed]
- Doritchamou, J.Y.A.; Renn, J.P.; Hviid, L.; Duffy, P.E. A conformational epitope in placental malaria vaccine antigen VAR2CSA: What does it teach us? PLoS Pathog. 2023, 19, e1011370. [Google Scholar] [CrossRef]
- Doritchamou, J.Y.A.; Suurbaar, J.; Tuikue Ndam, N. Progress and new horizons toward a VAR2CSA-based placental malaria vaccine. Expert Rev. Vaccines 2021, 20, 215–226. [Google Scholar] [CrossRef]
- Tomlinson, A.; Semblat, J.P.; Gamain, B.; Chene, A. VAR2CSA-Mediated Host Defense Evasion of Plasmodium falciparum Infected Erythrocytes in Placental Malaria. Front. Immunol. 2020, 11, 624126. [Google Scholar] [CrossRef]
- Wang, K.; Dagil, R.; Lavstsen, T.; Misra, S.K.; Spliid, C.B.; Wang, Y.; Gustavsson, T.; Sandoval, D.R.; Vidal-Calvo, E.E.; Choudhary, S.; et al. Cryo-EM reveals the architecture of placental malaria VAR2CSA and provides molecular insight into chondroitin sulfate binding. Nat. Commun. 2021, 12, 2956. [Google Scholar] [CrossRef]
- Raja, A.I.; Stanisic, D.I.; Good, M.F. Chemical Attenuation in the Development of a Whole-Organism Malaria Vaccine. Infect. Immun. 2017, 85, 10-1128. [Google Scholar] [CrossRef]
- Stanisic, D.I.; Fink, J.; Mayer, J.; Coghill, S.; Gore, L.; Liu, X.Q.; El-Deeb, I.; Rodriguez, I.B.; Powell, J.; Willemsen, N.M.; et al. Vaccination with chemically attenuated Plasmodium falciparum asexual blood-stage parasites induces parasite-specific cellular immune responses in malaria-naive volunteers: A pilot study. BMC Med. 2018, 16, 184. [Google Scholar] [CrossRef] [PubMed]
- Low, L.M.; Ssemaganda, A.; Liu, X.Q.; Ho, M.F.; Ozberk, V.; Fink, J.; Sundac, L.; Alcorn, K.; Morrison, A.; O’Callaghan, K.; et al. Controlled Infection Immunization Using Delayed Death Drug Treatment Elicits Protective Immune Responses to Blood-Stage Malaria Parasites. Infect. Immun. 2019, 87, 10-1128. [Google Scholar] [CrossRef] [PubMed]
- Stanisic, D.I.; Ho, M.F.; Nevagi, R.; Cooper, E.; Walton, M.; Islam, M.T.; Hussein, W.M.; Skwarczynski, M.; Toth, I.; Good, M.F. Development and Evaluation of a Cryopreserved Whole-Parasite Vaccine in a Rodent Model of Blood-Stage Malaria. mBio 2021, 12, e0265721. [Google Scholar] [CrossRef] [PubMed]
- Stanisic, D.I.; Good, M.F. Malaria Vaccines: Progress to Date. BioDrugs 2023, 37, 737–756. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Antigen a | Vaccine b | Most Advanced Outcome c | Structure d |
|---|---|---|---|
| AMA1 | FMP2.1/AS02A | P2b: Strain-specific [80,81] | Solved [82] |
| MSP1 | FMP1/AS02A | P2b: Ineffective [83] | Solved [84] |
| MSP2 | Combination B | P2b: Strain-specific [85] | ND |
| MSP3 | GMZ2 | P2b: Ineffective [86] | ND |
| EBA175 | EBA175 RII-NG | P1b: Safe, immunogenic [87] | Solved [88] |
| SERA5 | BK-SE36/CpG | P1b: Safe, immunogenic [89] | Solved [90] |
| Rh5 | RH5.1/AS01B | P2a: Reduction of PMR [91] | Solved [31,92] |
| Ripr | PfRipr5/CAF01 | Pre-clinical [93] | Solved [31,94] |
| CyRPA | CyRPA/GLA-SE | Pre-clinical [95] | Solved [96,97] |
| P27A | P27A/GLA-SE | P1b: Safe, immunogenic [98] | ND |
| PfSEA-1 | PfSEA-1/TiterMax | Experimental [99] | ND |
| PfGARP | PfGARP/mRNA | Pre-clinical [77] | ND |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takashima, E.; Otsuki, H.; Morita, M.; Ito, D.; Nagaoka, H.; Yuguchi, T.; Hassan, I.; Tsuboi, T. The Need for Novel Asexual Blood-Stage Malaria Vaccine Candidates for Plasmodium falciparum. Biomolecules 2024, 14, 100. https://doi.org/10.3390/biom14010100
Takashima E, Otsuki H, Morita M, Ito D, Nagaoka H, Yuguchi T, Hassan I, Tsuboi T. The Need for Novel Asexual Blood-Stage Malaria Vaccine Candidates for Plasmodium falciparum. Biomolecules. 2024; 14(1):100. https://doi.org/10.3390/biom14010100
Chicago/Turabian StyleTakashima, Eizo, Hitoshi Otsuki, Masayuki Morita, Daisuke Ito, Hikaru Nagaoka, Takaaki Yuguchi, Ifra Hassan, and Takafumi Tsuboi. 2024. "The Need for Novel Asexual Blood-Stage Malaria Vaccine Candidates for Plasmodium falciparum" Biomolecules 14, no. 1: 100. https://doi.org/10.3390/biom14010100
APA StyleTakashima, E., Otsuki, H., Morita, M., Ito, D., Nagaoka, H., Yuguchi, T., Hassan, I., & Tsuboi, T. (2024). The Need for Novel Asexual Blood-Stage Malaria Vaccine Candidates for Plasmodium falciparum. Biomolecules, 14(1), 100. https://doi.org/10.3390/biom14010100