
The Ubiquitin-Proteasome System Participates in Sperm Surface Subproteome Remodeling during Boar Sperm Capacitation



Abstract
1. Introduction
2. Materials and Methods
2.1. Antibodies and Reagents
2.2. Semen Processing, Sperm In Vitro Capacitation (IVC), and Mouse Tissue Collection
2.3. Biotin-Directed Affinity Purification of Sperm Surface Protein
2.4. Proteomics
2.5. Immunofluorescence
2.6. Epifluorescence Microscopy
2.7. Image-Based Flow Cytometry (IBFC)
2.8. Protein Isolation from Spermatozoa and Tissues
2.9. Immunoprecipitation
2.10. Western Blotting
2.11. Statistical Analysis
3. Results
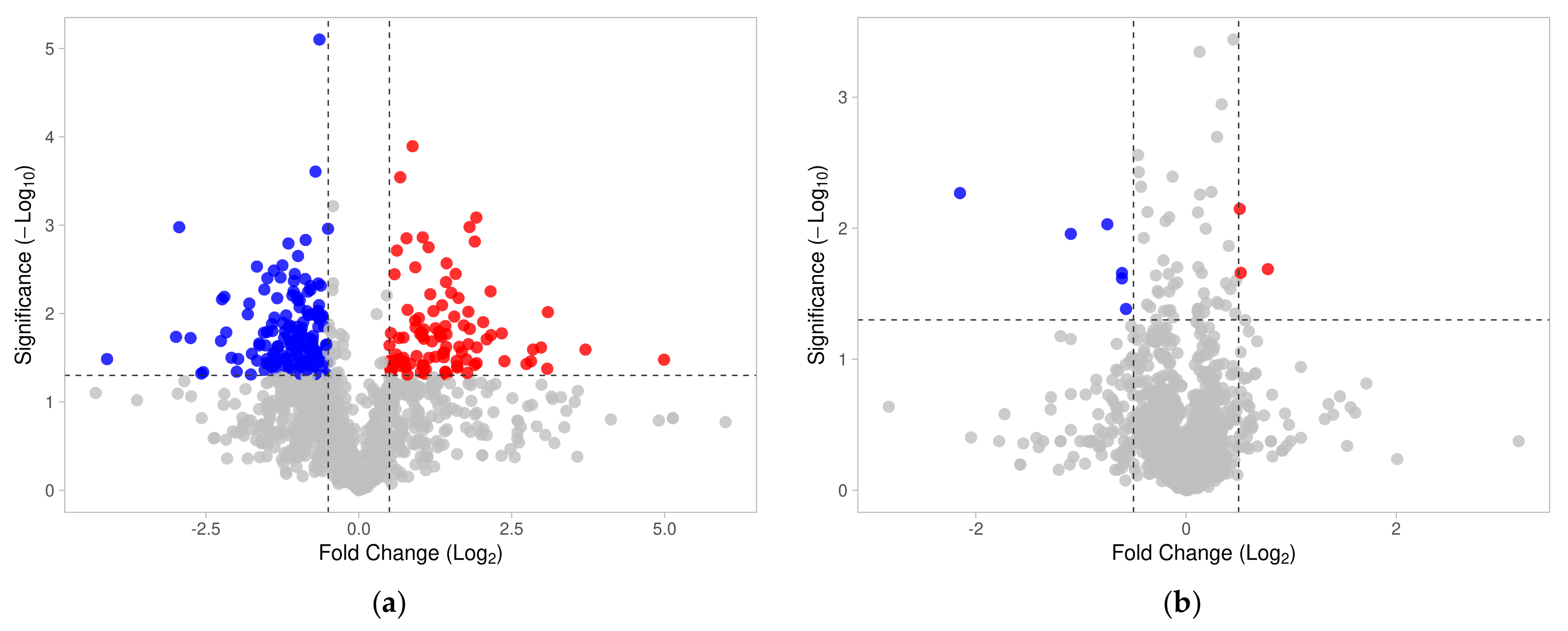
3.1. Identified Sperm Surface Proteins of Non-Capacitated and Capacitated Spermatozoa
3.2. Changes in Sperm Surface Subproteome after In Vitro Capacitation (Non-Capacitated vs. Capacitated Sperm Surface Subproteomes)
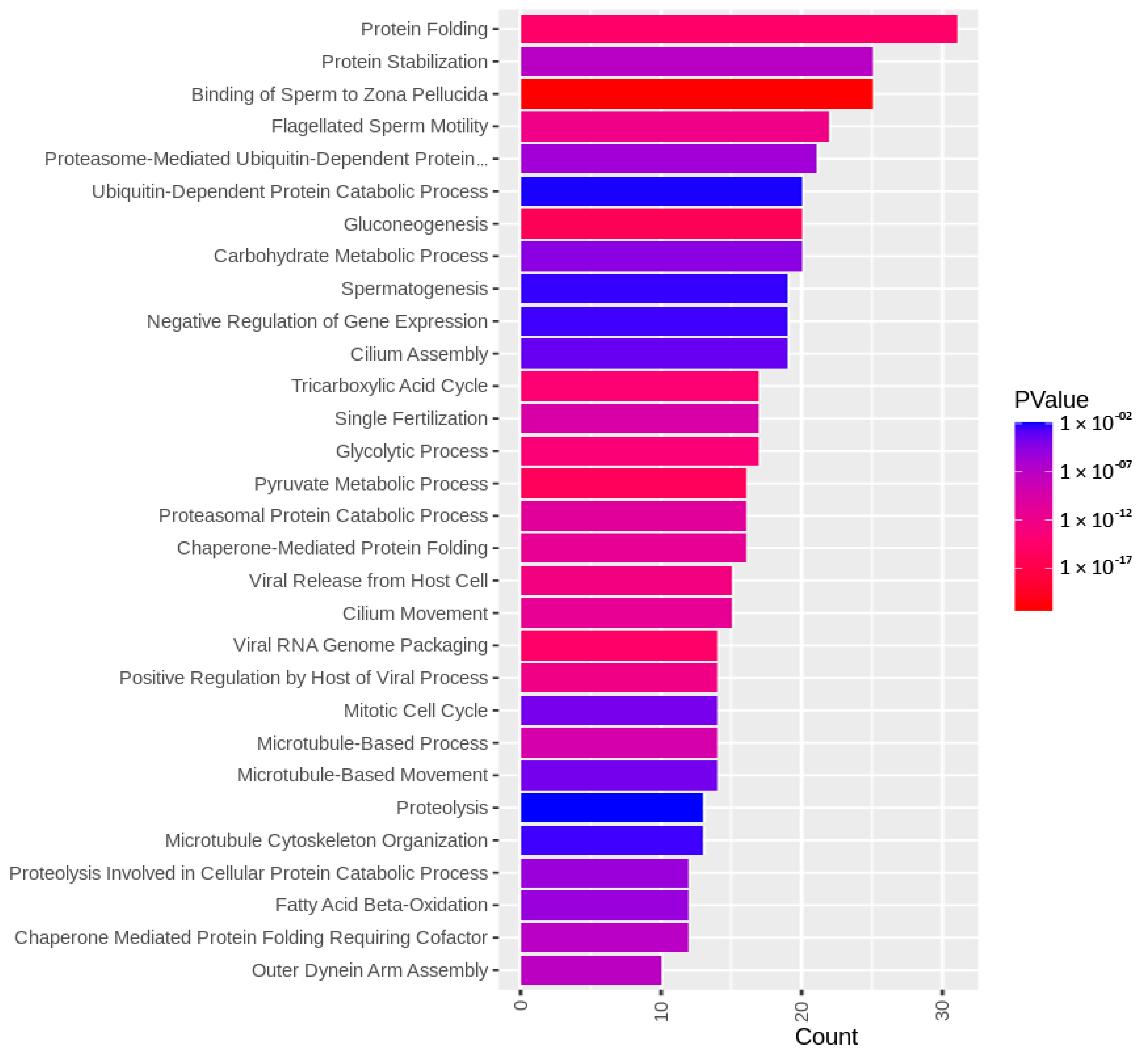
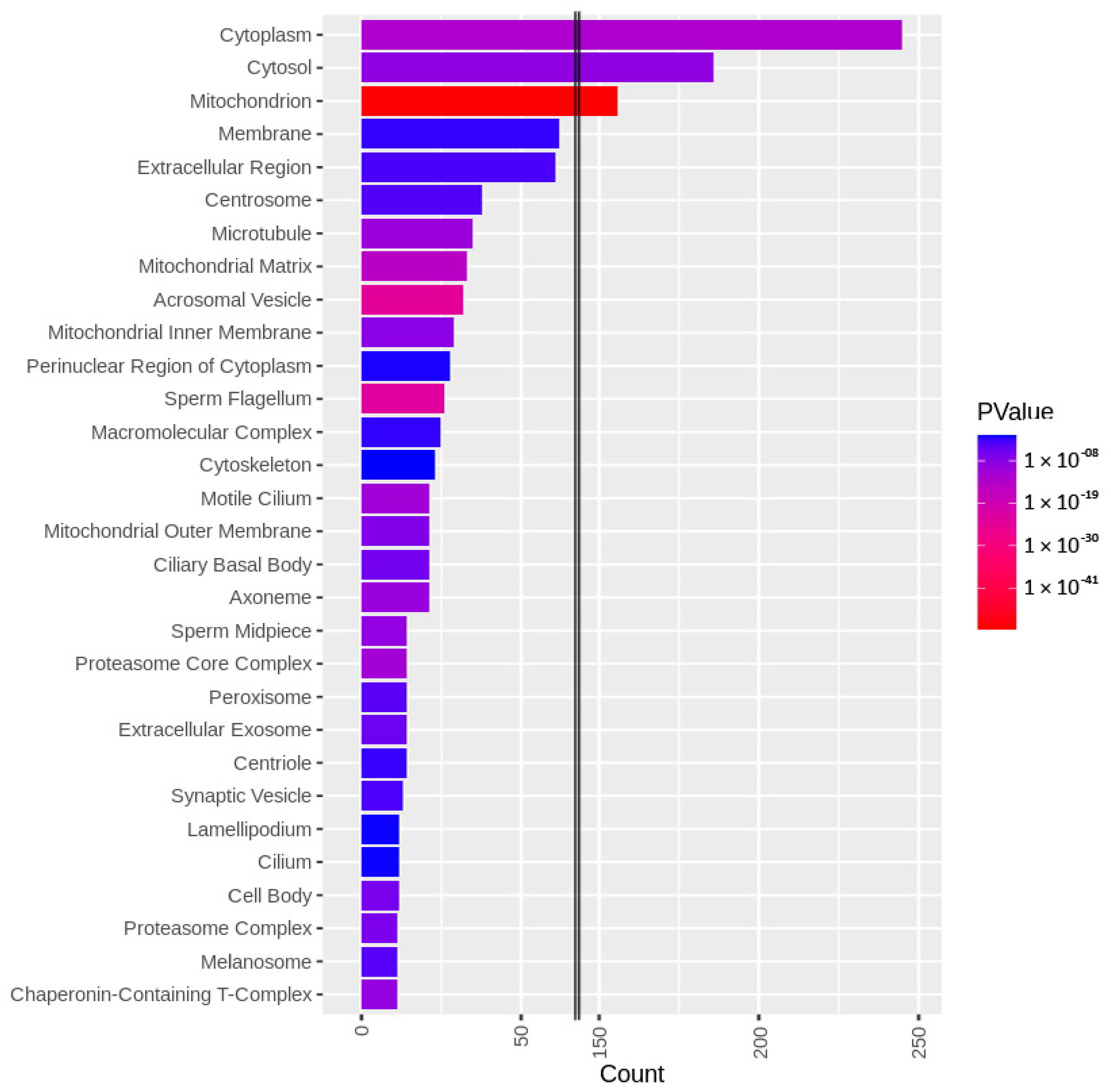
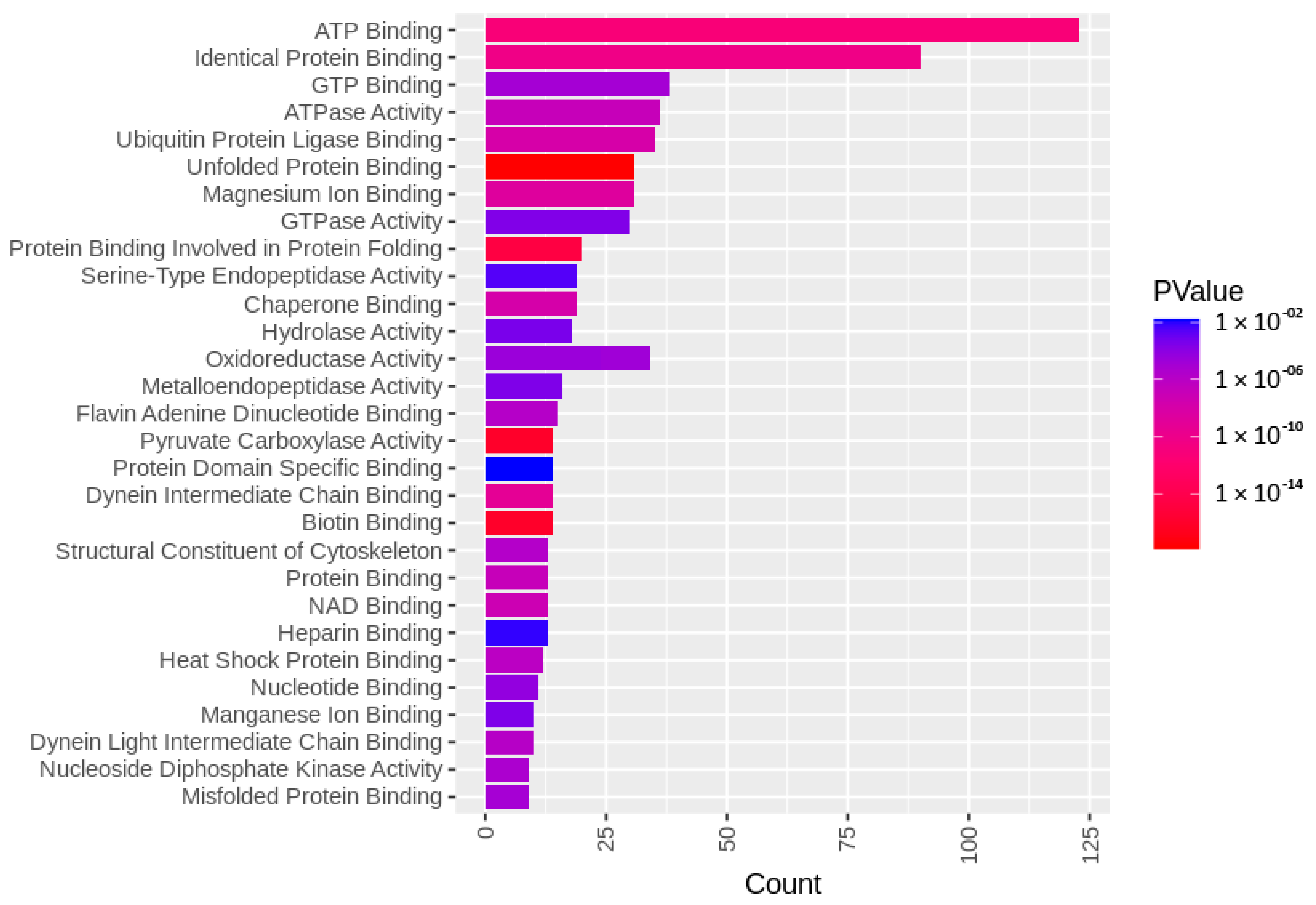
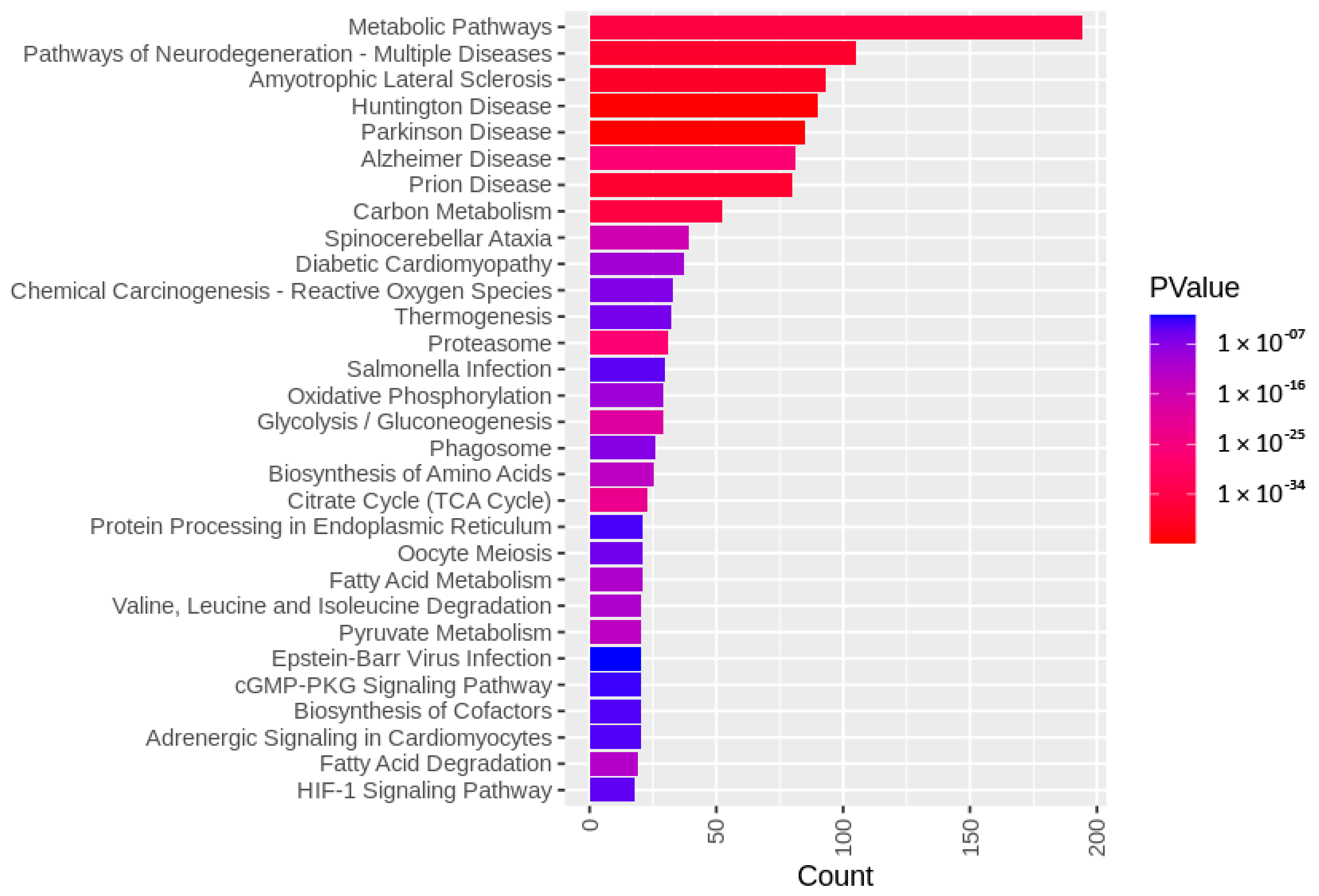
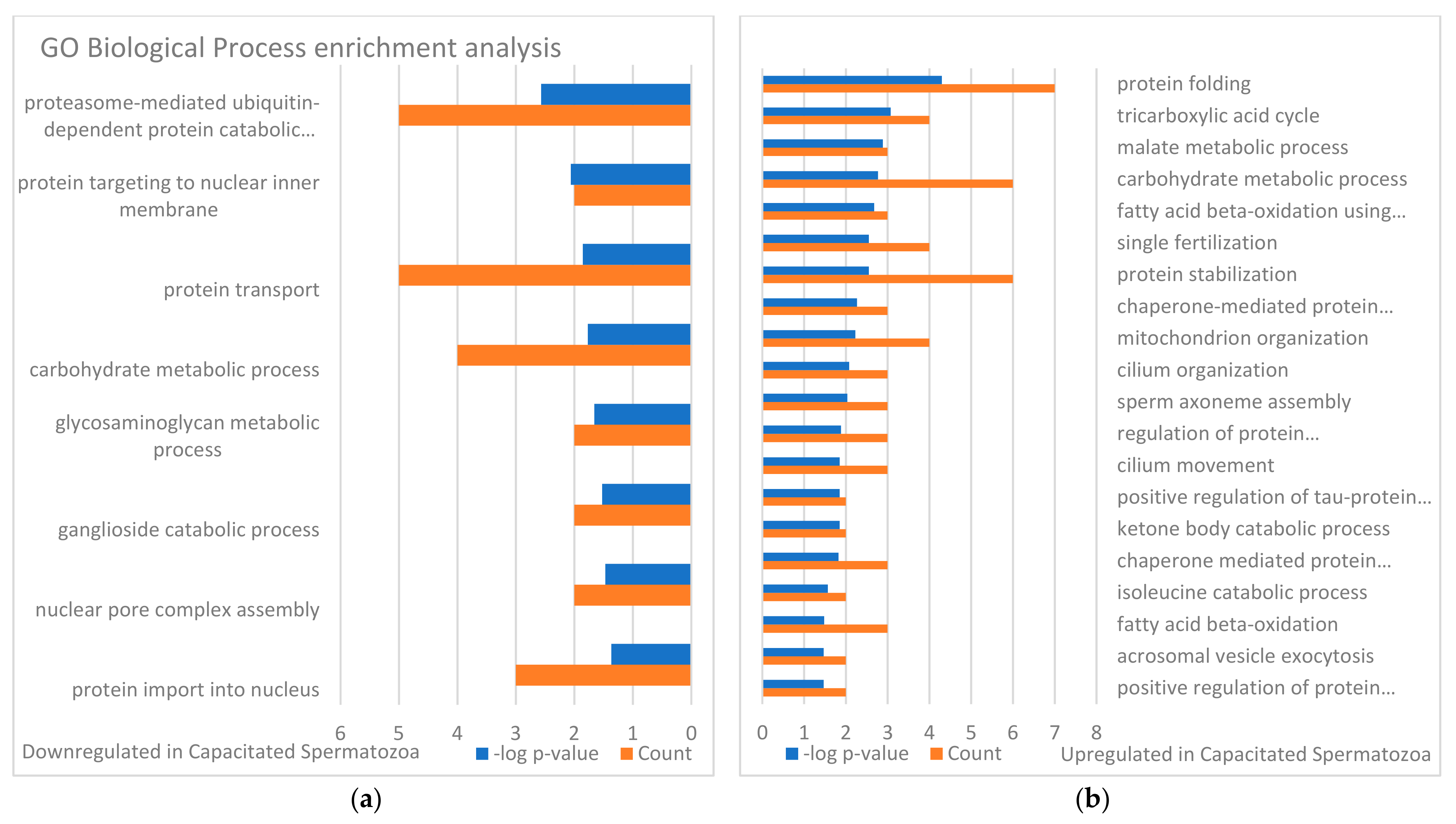
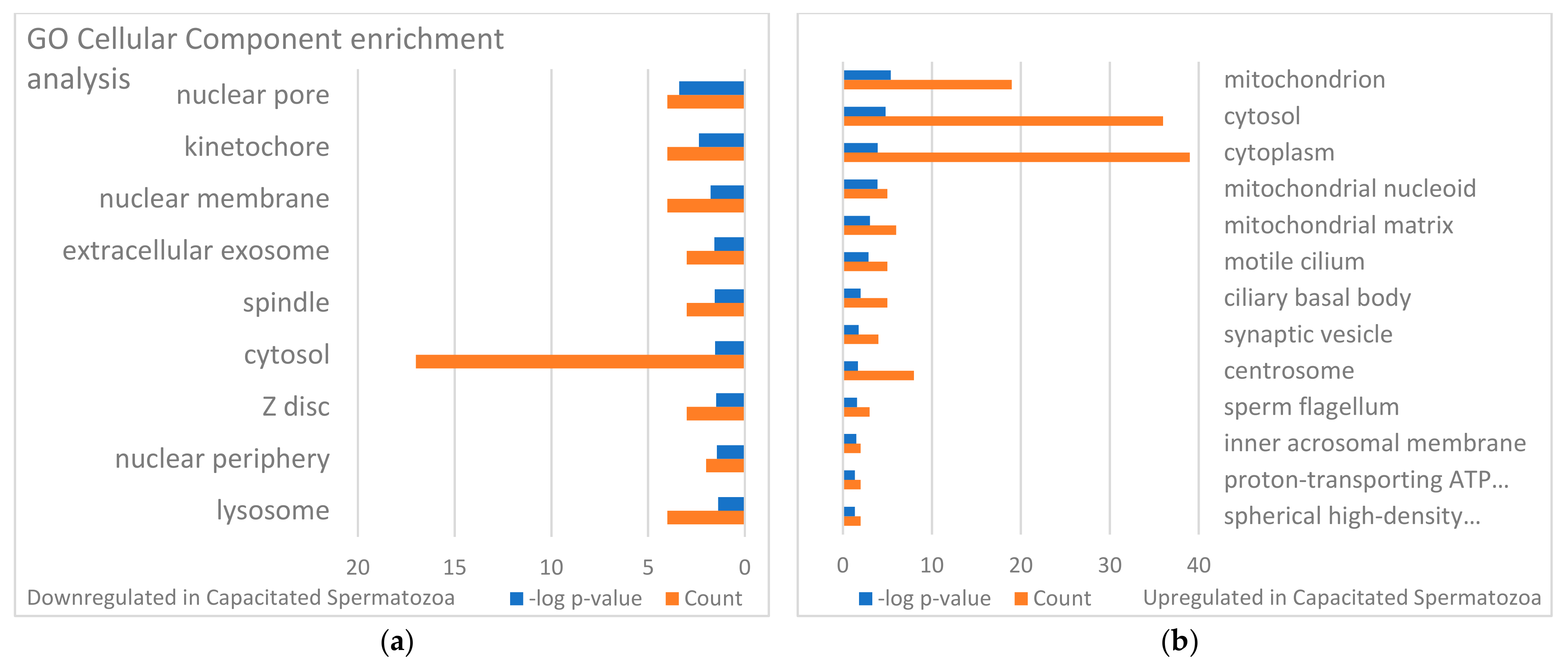
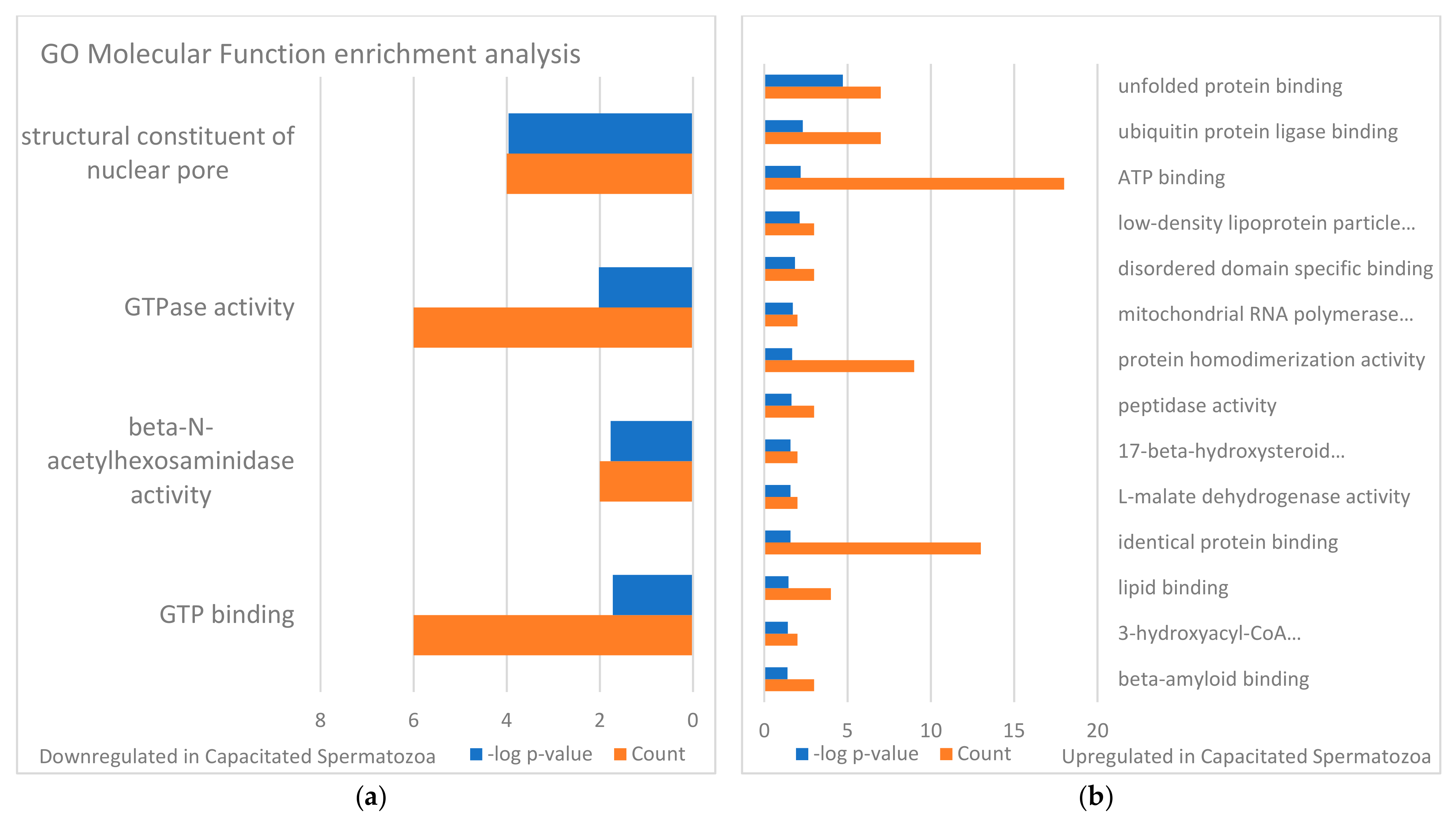
3.3. Targets of the Ubiquitin–Proteasome System on the Sperm Surface during In Vitro Capacitation (Sperm Surface Subproteomes of Capacitated, Proteasomally Inhibited vs. Capacitated, Vehicle Control Spermatozoa)
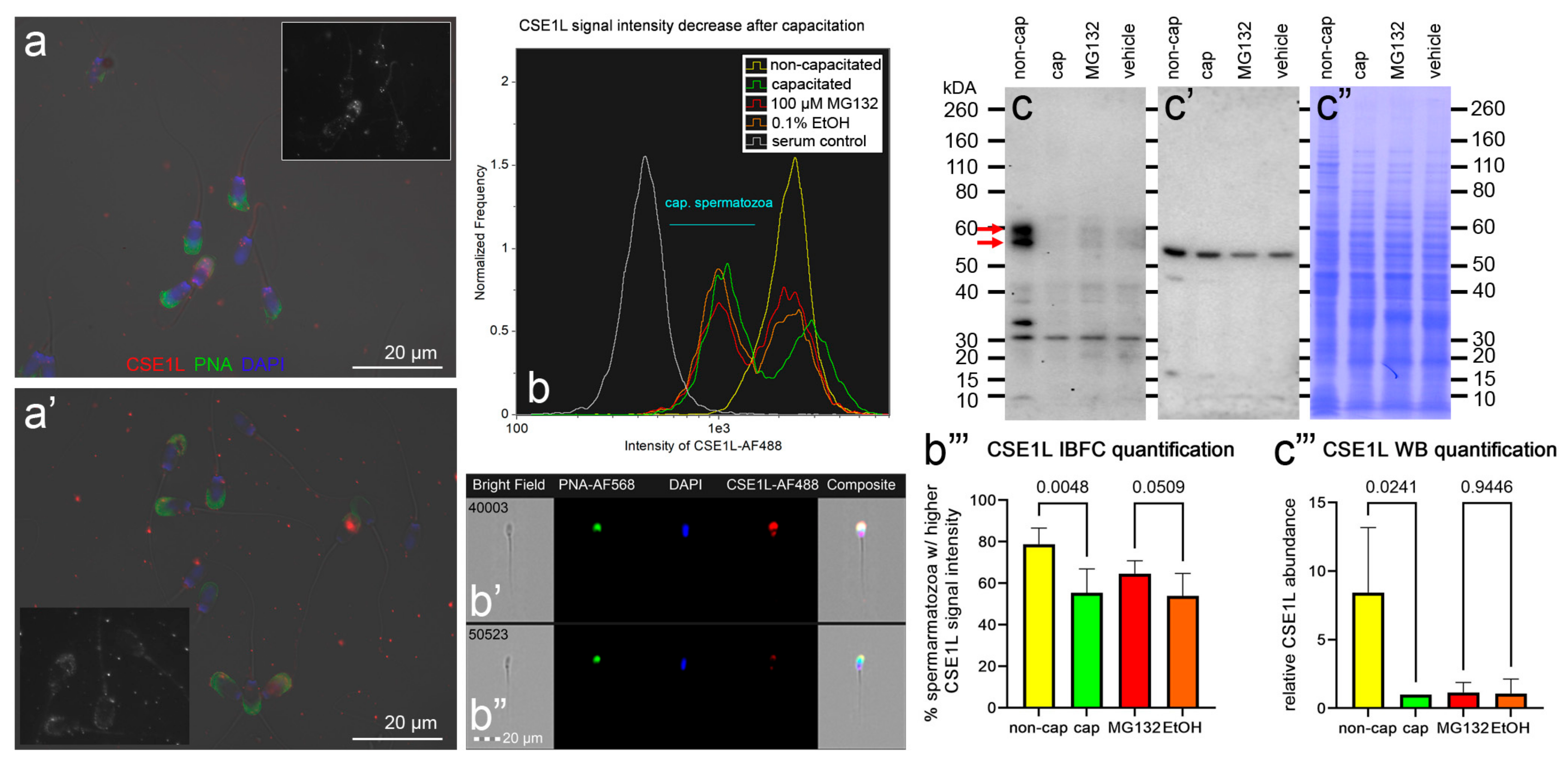
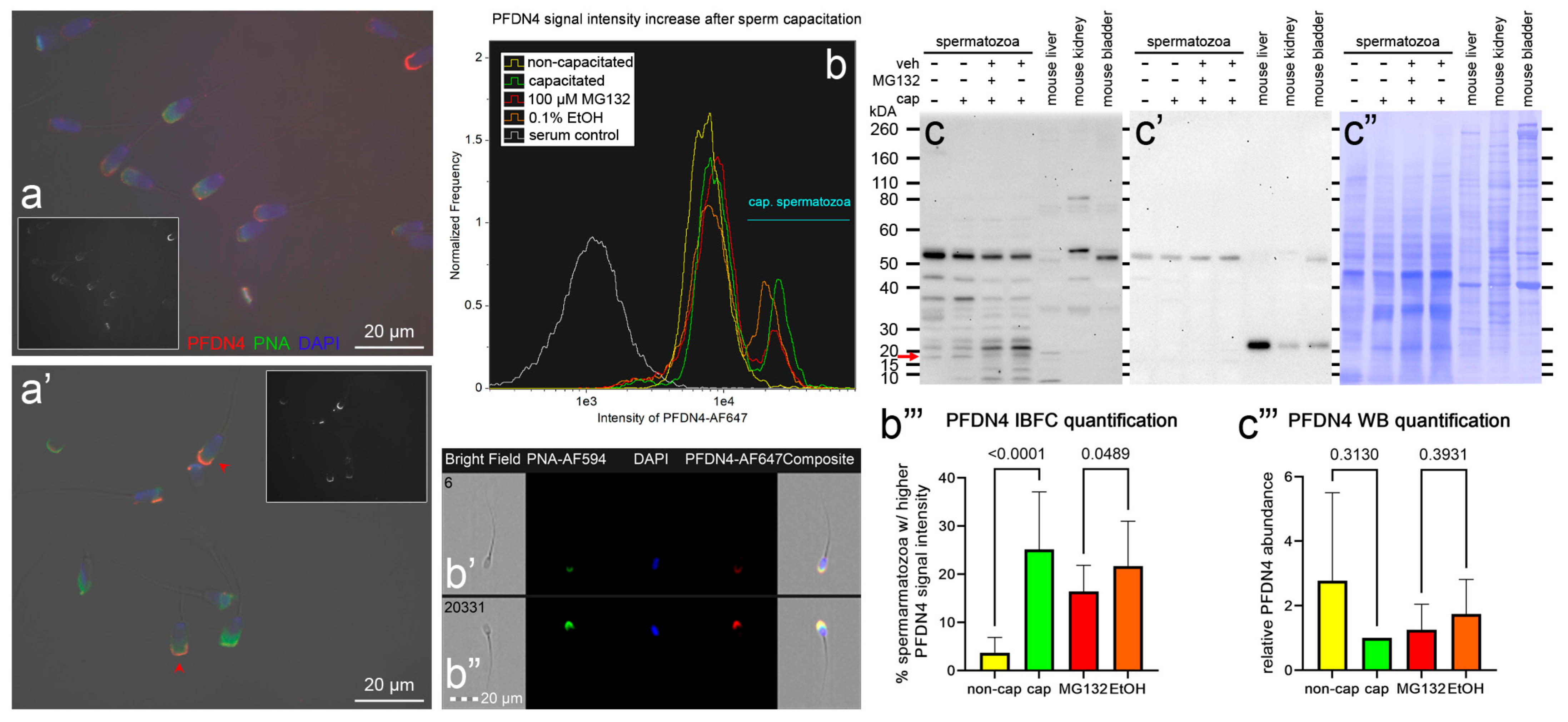
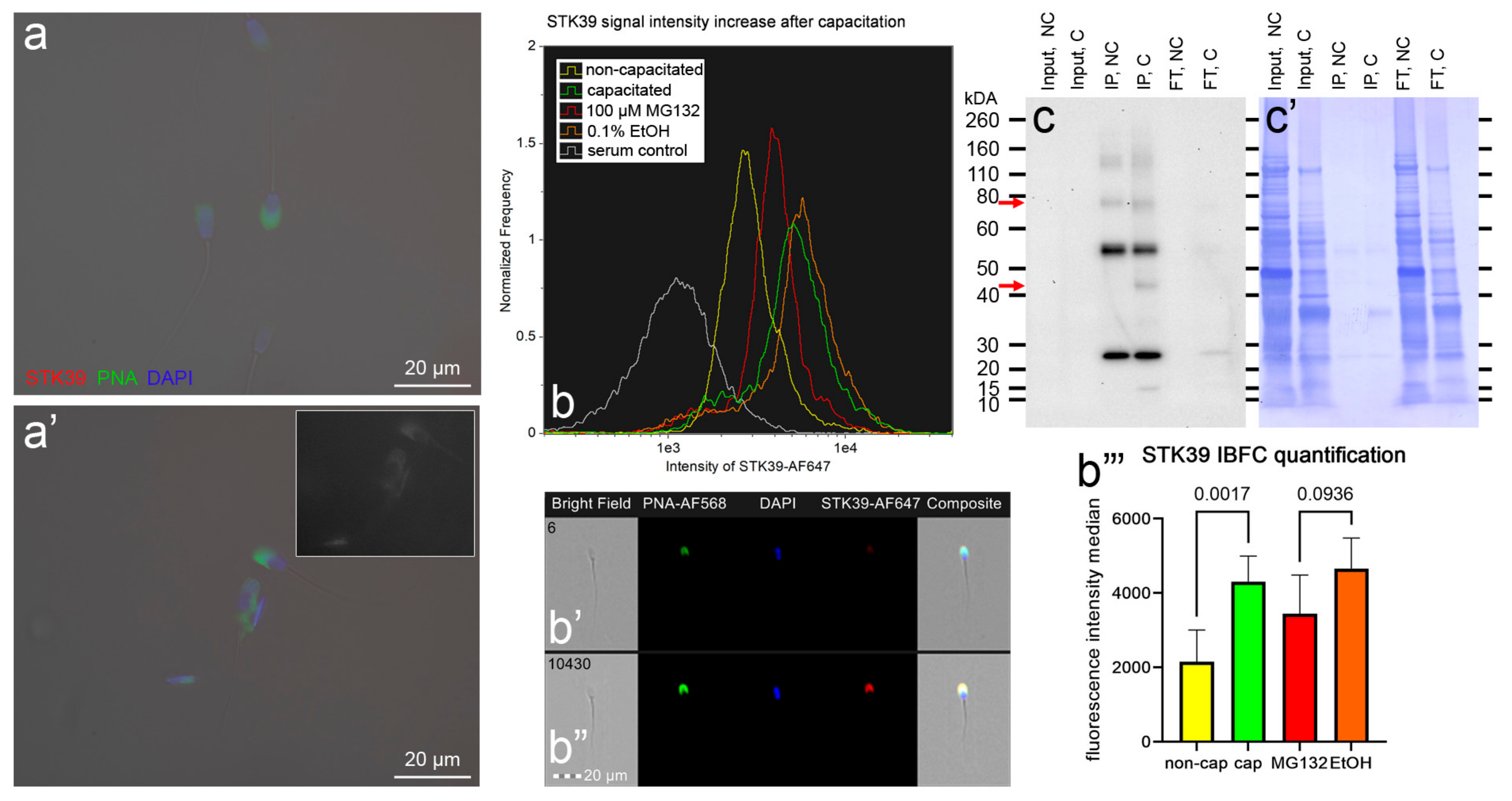
3.4. Localization and Dynamics of Selected Sperm Surface UPS-Regulated Proteins during In Vitro Sperm Capacitation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunt, L.T.; Dayhoff, M.O. Amino-terminal sequence identity of ubiquitin and the nonhistone component of nuclear protein A24. Biochem. Biophys. Res. Commun. 1977, 74, 650–655. [Google Scholar] [CrossRef]
- Hershko, A.; Heller, H. Occurrence of a polyubiquitin structure in ubiquitin-protein conjugates. Biochem. Biophys. Res. Commun. 1985, 128, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Hough, R.; Pratt, G.; Rechsteiner, M. Ubiquitin-lysozyme conjugates. Identification and characterization of an ATP-dependent protease from rabbit reticulocyte lysates. J. Biol. Chem. 1986, 261, 2400–2408. [Google Scholar] [CrossRef] [PubMed]
- Waxman, L.; Fagan, J.M.; Goldberg, A.L. Demonstration of two distinct high molecular weight proteases in rabbit reticulocytes, one of which degrades ubiquitin conjugates. J. Biol. Chem. 1987, 262, 2451–2457. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A.; Elias, S.; Heller, H.; Hershko, A. “Covalent affinity” purification of ubiquitin-activating enzyme. J. Biol. Chem. 1982, 257, 2537–2542. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Heller, H.; Elias, S.; Ciechanover, A. Components of ubiquitin-protein ligase system. Resolution, affinity purification, and role in protein breakdown. J. Biol. Chem. 1983, 258, 8206–8214. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. Intracellular protein degradation: From a vague idea thru the lysosome and the ubiquitin-proteasome system and onto human diseases and drug targeting. Best Pract. Res. Clin. Haematol. 2017, 30, 341–355. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P. Sperm proteasome and fertilization. Reproduction 2011, 142, 1–14. [Google Scholar] [CrossRef]
- Glickman, M.H.; Ciechanover, A. The ubiquitin-proteasome proteolytic pathway: Destruction for the sake of construction. Physiol. Rev. 2002, 82, 373–428. [Google Scholar] [CrossRef]
- Pickart, C.M.; Cohen, R.E. Proteasomes and their kin: Proteases in the machine age. Nat. Rev. Mol. Cell Biol. 2004, 5, 177–187. [Google Scholar] [CrossRef]
- Ciechanover, A. Proteolysis: From the lysosome to ubiquitin and the proteasome. Nat. Rev. Mol. Cell Biol. 2005, 6, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Austin, C.R. Observations on the penetration of the sperm in the mammalian egg. Aust. J. Sci. Res. Ser. B Biol. Sci. 1951, 4, 581–596. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.C. Fertilizing capacity of spermatozoa deposited into the fallopian tubes. Nature 1951, 168, 697–698. [Google Scholar] [CrossRef] [PubMed]
- Suarez, S.S. The oviductal sperm reservoir in mammals: Mechanisms of formation. Biol. Reprod. 1998, 58, 1105–1107. [Google Scholar] [CrossRef] [PubMed]
- Brukman, N.G.; Nunez, S.Y.; Puga Molina, L.D.C.; Buffone, M.G.; Darszon, A.; Cuasnicu, P.S.; Da Ros, V.G. Tyrosine phosphorylation signaling regulates Ca(2+) entry by affecting intracellular pH during human sperm capacitation. J. Cell. Physiol. 2019, 234, 5276–5288. [Google Scholar] [CrossRef]
- Kirichok, Y.; Navarro, B.; Clapham, D.E. Whole-cell patch-clamp measurements of spermatozoa reveal an alkaline-activated Ca2+ channel. Nature 2006, 439, 737–740. [Google Scholar] [CrossRef]
- Gadella, B.M.; Van Gestel, R.A. Bicarbonate and its role in mammalian sperm function. Anim. Reprod. Sci. 2004, 82–83, 307–319. [Google Scholar] [CrossRef]
- Visconti, P.E.; Bailey, J.L.; Moore, G.D.; Pan, D.; Olds-Clarke, P.; Kopf, G.S. Capacitation of mouse spermatozoa. I. Correlation between the capacitation state and protein tyrosine phosphorylation. Development 1995, 121, 1129–1137. [Google Scholar] [CrossRef]
- Visconti, P.E.; Moore, G.D.; Bailey, J.L.; Leclerc, P.; Connors, S.A.; Pan, D.; Olds-Clarke, P.; Kopf, G.S. Capacitation of mouse spermatozoa. II. Protein tyrosine phosphorylation and capacitation are regulated by a cAMP-dependent pathway. Development 1995, 121, 1139–1150. [Google Scholar] [CrossRef]
- Davis, B.K. Timing of fertilization in mammals: Sperm cholesterol/phospholipid ratio as a determinant of the capacitation interval. Proc. Natl. Acad. Sci. USA 1981, 78, 7560–7564. [Google Scholar] [CrossRef]
- Fraser, L.R. Mouse sperm capacitation in vitro involves loss of a surface-associated inhibitory component. J. Reprod. Fertil. 1984, 72, 373–384. [Google Scholar] [CrossRef]
- Kerns, K.; Zigo, M.; Drobnis, E.Z.; Sutovsky, M.; Sutovsky, P. Zinc ion flux during mammalian sperm capacitation. Nat. Commun. 2018, 9, 2061. [Google Scholar] [CrossRef]
- Leemans, B.; Stout, T.A.E.; De Schauwer, C.; Heras, S.; Nelis, H.; Hoogewijs, M.; Van Soom, A.; Gadella, B.M. Update on mammalian sperm capacitation: How much does the horse differ from other species? Reproduction 2019, 157, R181–R197. [Google Scholar] [CrossRef]
- Puga Molina, L.; Luque, G.; Balestrini, P.; Marín-Briggiler, C.; Romarowski, A.; Romarowski, A. Molecular Basis of Human Sperm Capacitation. Front. Cell Dev. Biol. 2018, 6, 72. [Google Scholar] [CrossRef]
- Harrison, R.A. Capacitation mechanisms, and the role of capacitation as seen in eutherian mammals. Reprod. Fertil. Dev. 1996, 8, 581–594. [Google Scholar] [CrossRef]
- Kerns, K.; Morales, P.; Sutovsky, P. Regulation of Sperm Capacitation by the 26S Proteasome: An Emerging New Paradigm in Spermatology. Biol. Reprod. 2016, 94, 117. [Google Scholar] [CrossRef]
- Kerns, K.; Zigo, M.; Sutovsky, P. Zinc: A Necessary Ion for Mammalian Sperm Fertilization Competency. Int. J. Mol. Sci. 2018, 19, 4097. [Google Scholar] [CrossRef]
- Manandhar, G.; Schatten, H.; Sutovsky, P. Centrosome reduction during gametogenesis and its significance. Biol. Reprod. 2005, 72, 2–13. [Google Scholar] [CrossRef] [PubMed]
- Mtango, N.R.; Latham, K.E.; Sutovsky, P. Deubiquitinating enzymes in oocyte maturation, fertilization and preimplantation embryo development. Adv. Exp. Med. Biol. 2014, 759, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Song, W.H.; Ballard, J.W.; Yi, Y.J.; Sutovsky, P. Regulation of mitochondrial genome inheritance by autophagy and ubiquitin-proteasome system: Implications for health, fitness, and fertility. BioMed Res. Int. 2014, 2014, 981867. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P. Ubiquitin-dependent proteolysis in mammalian spermatogenesis, fertilization, and sperm quality control: Killing three birds with one stone. Microsc. Res. Technol. 2003, 61, 88–102. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P. Sperm-egg adhesion and fusion in mammals. Expert Rev. Mol. Med. 2009, 11, e11. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P.; Song, W.H. Post-fertilisation sperm mitophagy: The tale of Mitochondrial Eve and Steve. Reprod. Fertil. Dev. 2017, 30, 56–63. [Google Scholar] [CrossRef]
- Sutovsky, P.; Van Leyen, K.; McCauley, T.; Day, B.N.; Sutovsky, M. Degradation of paternal mitochondria after fertilization: Implications for heteroplasmy, assisted reproductive technologies and mtDNA inheritance. Reprod. Biomed. Online 2004, 8, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Tumova, L.; Zigo, M.; Sutovsky, P.; Sedmikova, M.; Postlerova, P. Ligands and Receptors Involved in the Sperm-Zona Pellucida Interactions in Mammals. Cells 2021, 10, 133. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.J.; Manandhar, G.; Oko, R.J.; Breed, W.G.; Sutovsky, P. Mechanism of sperm-zona pellucida penetration during mammalian fertilization: 26S proteasome as a candidate egg coat lysin. Soc. Reprod. Fertil. Suppl. 2007, 63, 385–408. [Google Scholar]
- Zigo, M.; Manaskova-Postlerova, P.; Zuidema, D.; Kerns, K.; Jonakova, V.; Tumova, L.; Bubenickova, F.; Sutovsky, P. Porcine model for the study of sperm capacitation, fertilization and male fertility. Cell Tissue Res. 2020, 380, 237–262. [Google Scholar] [CrossRef]
- Zimmerman, S.; Sutovsky, P. The sperm proteasome during sperm capacitation and fertilization. J. Reprod. Immunol. 2009, 83, 19–25. [Google Scholar] [CrossRef]
- Hillman, P.; Ickowicz, D.; Vizel, R.; Breitbart, H. Dissociation between AKAP3 and PKARII promotes AKAP3 degradation in sperm capacitation. PLoS ONE 2013, 8, e68873. [Google Scholar] [CrossRef]
- Zapata-Carmona, H.; Barón, L.; Kong, M.; Morales, P. Protein Kinase A (PRKA) Activity Is Regulated by the Proteasome at the Onset of Human Sperm Capacitation. Cells 2021, 10, 3501. [Google Scholar] [CrossRef]
- Qu, X.; Han, Y.; Chen, X.; Lv, Y.; Zhang, Y.; Cao, L.; Zhang, J.; Jin, Y. Inhibition of 26S proteasome enhances AKAP3-mediated cAMP-PKA signaling during boar sperm capacitation. Anim. Reprod. Sci. 2022, 247, 107079. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, R.; Deppe, M.; Schulz, M.; Bravo, P.; Villegas, J.; Morales, P.; Risopatron, J. Participation of the sperm proteasome during in vitro fertilisation and the acrosome reaction in cattle. Andrologia 2011, 43, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Zigo, M.; Manaskova-Postlerova, P.; Jonakova, V.; Kerns, K.; Sutovsky, P. Compartmentalization of the proteasome-interacting proteins during sperm capacitation. Sci. Rep. 2019, 9, 12583. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.J.; Sutovsky, M.; Kennedy, C.; Sutovsky, P. Identification of the inorganic pyrophosphate metabolizing, ATP substituting pathway in mammalian spermatozoa. PLoS ONE 2012, 7, e34524. [Google Scholar] [CrossRef] [PubMed]
- Hackerova, L.; Klusackova, B.; Zigo, M.; Zelenkova, N.; Havlikova, K.; Krejcirova, R.; Sedmikova, M.; Sutovsky, P.; Komrskova, K.; Postlerova, P.; et al. Modulatory effect of MG-132 proteasomal inhibition on boar sperm motility during in vitro capacitation. Front. Vet. Sci. 2023, 10, 1116891. [Google Scholar] [CrossRef]
- Zigo, M.; Jonakova, V.; Manaskova-Postlerova, P.; Kerns, K.; Sutovsky, P. Ubiquitin-proteasome system participates in the de-aggregation of spermadhesin and DQH protein during boar sperm capacitation. Reproduction 2019, 157, 283–295. [Google Scholar] [CrossRef]
- Sharif, M.; Kerns, K.; Sutovsky, P.; Bovin, N.; Miller, D.J. Progesterone induces porcine sperm release from oviduct glycans in a proteasome-dependent manner. Reproduction 2021, 161, 449–457. [Google Scholar] [CrossRef]
- Sharif, M.; Hickl, V.; Juarez, G.; Di, X.; Kerns, K.; Sutovsky, P.; Bovin, N.; Miller, D.J. Hyperactivation is sufficient to release porcine sperm from immobilized oviduct glycans. Sci. Rep. 2022, 12, 6446. [Google Scholar] [CrossRef]
- Taraschi, A.; Cimini, C.; Capacchietti, G.; Ramal-Sanchez, M.; Valbonetti, L.; Machado-Simoes, J.; Moussa, F.; Tagaram, I.; Mokh, S.; Al Iskandarani, M.; et al. Two-Player Game in a Complex Landscape: 26S Proteasome, PKA, and Intracellular Calcium Concentration Modulate Mammalian Sperm Capacitation by Creating an Integrated Dialogue—A Computational Analysis. Int. J. Mol. Sci. 2020, 21, 6256. [Google Scholar] [CrossRef]
- Zigo, M.; Kerns, K.; Sutovsky, M.; Sutovsky, P. Modifications of the 26S proteasome during boar sperm capacitation. Cell Tissue Res. 2018, 372, 591–601. [Google Scholar] [CrossRef]
- Kong, M.; Diaz, E.S.; Morales, P. Participation of the human sperm proteasome in the capacitation process and its regulation by protein kinase A and tyrosine kinase. Biol. Reprod. 2009, 80, 1026–1035. [Google Scholar] [CrossRef] [PubMed]
- Zapata-Carmona, H.; Barón, L.; Zuñiga, L.M.; Díaz, E.S.; Kong, M.; Drobnis, E.Z.; Sutovsky, P.; Morales, P. The activation of the chymotrypsin-like activity of the proteasome is regulated by soluble adenyl cyclase/cAMP/protein kinase A pathway and required for human sperm capacitation. Mol. Hum. Reprod. 2019, 25, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Zigo, M.; Jonakova, V.; Sulc, M.; Manaskova-Postlerova, P. Characterization of sperm surface protein patterns of ejaculated and capacitated boar sperm, with the detection of ZP binding candidates. Int. J. Biol. Macromol. 2013, 61, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, C.E.; Krieger, K.B.; Sutovsky, M.; Xu, W.; Vargovic, P.; Didion, B.A.; Ellersieck, M.R.; Hennessy, M.E.; Verstegen, J.; Oko, R.; et al. Protein expression pattern of PAWP in bull spermatozoa is associated with sperm quality and fertility following artificial insemination. Mol. Reprod. Dev. 2014, 81, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Blighe, K.; Rana, S.; Lewis, M. EnhancedVolcano: Publication-Ready Volcano Plots with Enhanced Colouring and Labeling. R Package Version 1.8.0. Available online: https://bioconductor.org/packages/release/bioc/vignettes/EnhancedVolcano/inst/doc/EnhancedVolcano.html (accessed on 15 March 2021).
- Kim, I.; Chu, X.Y.; Kim, S.; Provoda, C.J.; Lee, K.D.; Amidon, G.L. Identification of a human valacyclovirase: Biphenyl hydrolase-like protein as valacyclovir hydrolase. J. Biol. Chem. 2003, 278, 25348–25356. [Google Scholar] [CrossRef]
- Marsillach, J.; Suzuki, S.M.; Richter, R.J.; McDonald, M.G.; Rademacher, P.M.; MacCoss, M.J.; Hsieh, E.J.; Rettie, A.E.; Furlong, C.E. Human valacyclovir hydrolase/biphenyl hydrolase-like protein is a highly efficient homocysteine thiolactonase. PLoS ONE 2014, 9, e110054. [Google Scholar] [CrossRef]
- Sharma, R.; Agarwal, A.; Mohanty, G.; Hamada, A.J.; Gopalan, B.; Willard, B.; Yadav, S.; du Plessis, S. Proteomic analysis of human spermatozoa proteins with oxidative stress. Reprod. Biol. Endocrinol. RBE 2013, 11, 48. [Google Scholar] [CrossRef]
- Shi, J.; Qiao, F.; Ye, M.; Jiang, T.; Liu, J.; Zhang, M.; Xie, G.; Fok, K.L.; Li, X.; Chen, H. CSE1L/CAS regulates cell proliferation through CDK signalling in mouse spermatogenesis. Cell Prolif. 2022, 55, e13334. [Google Scholar] [CrossRef]
- Finkelstein, M.; Megnagi, B.; Ickowicz, D.; Breitbart, H. Regulation of sperm motility by PIP2(4,5) and actin polymerization. Dev. Biol. 2013, 381, 62–72. [Google Scholar] [CrossRef]
- Finkelstein, M.; Etkovitz, N.; Breitbart, H. Role and regulation of sperm gelsolin prior to fertilization. J. Biol. Chem. 2010, 285, 39702–39709. [Google Scholar] [CrossRef]
- Lee, J.A.; Park, J.E.; Lee, D.H.; Park, S.G.; Myung, P.K.; Park, B.C.; Cho, S. G1 to S phase transition protein 1 induces apoptosis signal-regulating kinase 1 activation by dissociating 14-3-3 from ASK1. Oncogene 2008, 27, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, R.; Wanschers, B.F.; Nabuurs, S.B.; Nouws, J.; Nijtmans, L.G.; Huynen, M.A. NDUFB7 and NDUFA8 are located at the intermembrane surface of complex I. FEBS Lett. 2011, 585, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Tascou, S.; Uedelhoven, J.; Dixkens, C.; Nayernia, K.; Engel, W.; Burfeind, P. Isolation and characterization of a novel human gene, NIF3L1, and its mouse ortholog, Nif3l1, highly conserved from bacteria to mammals. Cytogenet. Cell Genet. 2000, 90, 330–336. [Google Scholar] [CrossRef] [PubMed]
- Tascou, S.; Kang, T.W.; Trappe, R.; Engel, W.; Burfeind, P. Identification and characterization of NIF3L1 BP1, a novel cytoplasmic interaction partner of the NIF3L1 protein. Biochem. Biophys. Res. Commun. 2003, 309, 440–448. [Google Scholar] [CrossRef]
- Siegert, R.; Leroux, M.R.; Scheufler, C.; Hartl, F.U.; Moarefi, I. Structure of the molecular chaperone prefoldin: Unique interaction of multiple coiled coil tentacles with unfolded proteins. Cell 2000, 103, 621–632. [Google Scholar] [CrossRef]
- Sahlan, M.; Zako, T.; Yohda, M. Prefoldin, a jellyfish-like molecular chaperone: Functional cooperation with a group II chaperonin and beyond. Biophys. Rev. 2018, 10, 339–345. [Google Scholar] [CrossRef]
- Amaral, A.; Castillo, J.; Estanyol, J.M.; Ballescà, J.L.; Ramalho-Santos, J.; Oliva, R. Human sperm tail proteome suggests new endogenous metabolic pathways. Mol. Cell. Proteom. MCP 2013, 12, 330–342. [Google Scholar] [CrossRef]
- Han, F.; Dong, M.Z.; Lei, W.L.; Xu, Z.L.; Gao, F.; Schatten, H.; Wang, Z.B.; Sun, X.F.; Sun, Q.Y. Oligoasthenoteratospermia and sperm tail bending in PPP4C-deficient mice. Mol. Hum. Reprod. 2021, 27, gaaa083. [Google Scholar] [CrossRef]
- Beaton, C. Protein Phosphatase 4 in Human Spermatozoa: Investigations into Location, Regulatory Subunits and Role in Motility. Master’s Thesis, University of Dundee, Dundee, UK, 2012. [Google Scholar]
- Liu, Y.L.; Yang, S.S.; Chen, S.J.; Lin, Y.C.; Chu, C.C.; Huang, H.H.; Chang, F.W.; Yu, M.H.; Lin, S.H.; Wu, G.J.; et al. OSR1 and SPAK cooperatively modulate Sertoli cell support of mouse spermatogenesis. Sci. Rep. 2016, 6, 37205. [Google Scholar] [CrossRef]
- Qi, H.; Labrie, Y.; Grenier, J.; Fournier, A.; Fillion, C.; Labrie, C. Androgens induce expression of SPAK, a STE20/SPS1-related kinase, in LNCaP human prostate cancer cells. Mol. Cell Endocrinol. 2001, 182, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhu, W.; Xiong, L.; Yu, X.; Chen, X.; Lin, Q. Role of high expression levels of STK39 in the growth, migration and invasion of non-small cell type lung cancer cells. Oncotarget 2016, 7, 61366–61377. [Google Scholar] [CrossRef]
- Hinton, S.D. The role of pseudophosphatases as signaling regulators. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 167–174. [Google Scholar] [CrossRef]
- Wishart, M.J.; Dixon, J.E. The archetype STYX/dead-phosphatase complexes with a spermatid mRNA-binding protein and is essential for normal sperm production. Proc. Natl. Acad. Sci. USA 2002, 99, 2112–2117. [Google Scholar] [CrossRef]
- Paschen, S.A.; Neupert, W. Protein import into mitochondria. IUBMB Life 2001, 52, 101–112. [Google Scholar] [CrossRef]
- Zigo, M.; Kerns, K.; Sen, S.; Essien, C.; Oko, R.; Xu, D.; Sutovsky, P. Zinc is a master-regulator of sperm function associated with binding, motility, and metabolic modulation during porcine sperm capacitation. Commun. Biol. 2022, 5, 538. [Google Scholar] [CrossRef]
- Hamilton, L.E.; Zigo, M.; Mao, J.; Xu, W.; Sutovsky, P.; O’Flaherty, C.; Oko, R. GSTO2 Isoforms Participate in the Oxidative Regulation of the Plasmalemma in Eutherian Spermatozoa during Capacitation. Antioxidants 2019, 8, 601. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, W.; Kho, Y.; Zhao, Y. Proteomic analysis of integral plasma membrane proteins. Anal. Chem. 2004, 76, 1817–1823. [Google Scholar] [CrossRef]
- Kasvandik, S.; Sillaste, G.; Velthut-Meikas, A.; Mikelsaar, A.V.; Hallap, T.; Padrik, P.; Tenson, T.; Jaakma, Ü.; Kõks, S.; Salumets, A. Bovine sperm plasma membrane proteomics through biotinylation and subcellular enrichment. Proteomics 2015, 15, 1906–1920. [Google Scholar] [CrossRef] [PubMed]
- Song, W.H.; Yi, Y.J.; Sutovsky, M.; Meyers, S.; Sutovsky, P. Autophagy and ubiquitin-proteasome system contribute to sperm mitophagy after mammalian fertilization. Proc. Natl. Acad. Sci. USA 2016, 113, E5261–E5270. [Google Scholar] [CrossRef] [PubMed]
- Zuidema, D.; Jones, A.; Song, W.H.; Zigo, M.; Sutovsky, P. Identification of Candidate Mitochondrial Inheritance Determinants Using the Mammalian Cell-Free System. Elife 2023. [Google Scholar] [CrossRef]
- Wassarman, P.M. Mammalian fertilization: The strange case of sperm protein 56. BioEssays News Rev. Mol. Cell. Dev. Biol. 2009, 31, 153–158. [Google Scholar] [CrossRef]
- Tanphaichitr, N.; Kongmanas, K.; Kruevaisayawan, H.; Saewu, A.; Sugeng, C.; Fernandes, J.; Souda, P.; Angel, J.B.; Faull, K.F.; Aitken, R.J.; et al. Remodeling of the plasma membrane in preparation for sperm-egg recognition: Roles of acrosomal proteins. Asian J. Androl. 2015, 17, 574–582. [Google Scholar] [CrossRef]
- López-Salguero, J.B.; Fierro, R.; Michalski, J.C.; Jiménez-Morales, I.; Lefebvre, T.; Mondragón-Payne, O.; Baldini, S.F.; Vercoutter-Edouart, A.S.; González-Márquez, H. Identification of lipid raft glycoproteins obtained from boar spermatozoa. Glycoconj. J. 2020, 37, 499–509. [Google Scholar] [CrossRef]
- Zhang, M.; Chiozzi, R.Z.; Skerrett-Byrne, D.A.; Veenendaal, T.; Klumperman, J.; Heck, A.J.R.; Nixon, B.; Helms, J.B.; Gadella, B.M.; Bromfield, E.G. High Resolution Proteomic Analysis of Subcellular Fractionated Boar Spermatozoa Provides Comprehensive Insights Into Perinuclear Theca-Residing Proteins. Front. Cell Dev. Biol. 2022, 10, 836208. [Google Scholar] [CrossRef]
- Lawlor, M.; Zigo, M.; Kerns, K.; Cho, I.K.; Easley IV, C.A.; Sutovsky, P. Spermatozoan Metabolism as a Non-Traditional Model for the Study of Huntington’s Disease. Int. J. Mol. Sci. 2022, 23, 7163. [Google Scholar] [CrossRef]
- Meizel, S. The sperm, a neuron with a tail: ‘neuronal’ receptors in mammalian sperm. Biol. Rev. Camb. Philos. Soc. 2004, 79, 713–732. [Google Scholar] [CrossRef] [PubMed]
- Dun, M.D.; Smith, N.D.; Baker, M.A.; Lin, M.; Aitken, R.J.; Nixon, B. The chaperonin containing TCP1 complex (CCT/TRiC) is involved in mediating sperm-oocyte interaction. J. Biol. Chem. 2011, 286, 36875–36887. [Google Scholar] [CrossRef] [PubMed]
- Kongmanas, K.; Kruevaisayawan, H.; Saewu, A.; Sugeng, C.; Fernandes, J.; Souda, P.; Angel, J.B.; Faull, K.F.; Aitken, R.J.; Whitelegge, J.; et al. Proteomic Characterization of Pig Sperm Anterior Head Plasma Membrane Reveals Roles of Acrosomal Proteins in ZP3 Binding. J. Cell. Physiol. 2015, 230, 449–463. [Google Scholar] [CrossRef]
- Redgrove, K.A.; Anderson, A.L.; Dun, M.D.; McLaughlin, E.A.; O’Bryan, M.K.; Aitken, R.J.; Nixon, B. Involvement of multimeric protein complexes in mediating the capacitation-dependent binding of human spermatozoa to homologous zonae pellucidae. Dev. Biol. 2011, 356, 460–474. [Google Scholar] [CrossRef]
- Brewis, I.A.; Gadella, B.M. Sperm surface proteomics: From protein lists to biological function. Mol. Hum. Reprod. 2010, 16, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P.; Ramalho-Santos, J.; Moreno, R.D.; Oko, R.; Hewitson, L.; Schatten, G. On-stage selection of single round spermatids using a vital, mitochondrion-specific fluorescent probe MitoTracker(TM) and high resolution differential interference contrast microscopy. Hum. Reprod. 1999, 14, 2301–2312. [Google Scholar] [CrossRef]
- Zigo, M.; Dorosh, A.; Pohlova, A.; Jonakova, V.; Sulc, M.; Manaskova-Postlerova, P. Panel of monoclonal antibodies to sperm surface proteins as a tool for monitoring localization and identification of sperm-zona pellucida receptors. Cell Tissue Res. 2015, 359, 895–908. [Google Scholar] [CrossRef] [PubMed]
- Kisselev, A.F.; Goldberg, A.L. Proteasome inhibitors: From research tools to drug candidates. Chem. Biol. 2001, 8, 739–758. [Google Scholar] [CrossRef]
- Liao, C.F.; Lin, S.H.; Chen, H.C.; Tai, C.J.; Chang, C.C.; Li, L.T.; Yeh, C.M.; Yeh, K.T.; Chen, Y.C.; Hsu, T.H.; et al. CSE1L, a novel microvesicle membrane protein, mediates Ras-triggered microvesicle generation and metastasis of tumor cells. Mol. Med. 2012, 18, 1269–1280. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.C. CAS (CSE1L) signaling pathway in tumor progression and its potential as a biomarker and target for targeted therapy. Tumour. Biol. 2016, 37, 13077–13090. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J. Reactive oxygen species as mediators of sperm capacitation and pathological damage. Mol. Reprod. Dev. 2017, 84, 1039–1052. [Google Scholar] [CrossRef]
- Jin, M.; Liu, C.; Han, W.; Cong, Y. TRiC/CCT Chaperonin: Structure and Function. Subcell. Biochem. 2019, 93, 625–654. [Google Scholar] [CrossRef]
- Miyoshi, N.; Ishii, H.; Mimori, K.; Nishida, N.; Tokuoka, M.; Akita, H.; Sekimoto, M.; Doki, Y.; Mori, M. Abnormal expression of PFDN4 in colorectal cancer: A novel marker for prognosis. Ann. Surg. Oncol. 2010, 17, 3030–3036. [Google Scholar] [CrossRef]
- Sadeghi, H.; Nazemalhosseini-Mojarad, E.; Sahebi, U.; Fazeli, E.; Azizi-Tabesh, G.; Yassaee, V.R.; Savabkar, S.; Asadzadeh Aghdaei, H.; Zali, M.R.; Mirfakhraie, R. Novel long noncoding RNAs upregulation may have synergistic effects on the CYP24A1 and PFDN4 biomarker role in human colorectal cancer. J. Cell. Physiol. 2021, 236, 2051–2057. [Google Scholar] [CrossRef]
- Wang, D.; Zhu, Z.Z.; Jiang, H.; Zhu, J.; Cong, W.M.; Wen, B.J.; He, S.Q.; Liu, S.F. Multiple genes identified as targets for 20q13.12-13.33 gain contributing to unfavorable clinical outcomes in patients with hepatocellular carcinoma. Hepatol. Int. 2015, 9, 438–446. [Google Scholar] [CrossRef]
- Feng, M.Y.; Wang, K.; Shi, Q.T.; Yu, X.W.; Geng, J.S. Gene expression profiling in TWIST-depleted gastric cancer cells. Anat. Rec. 2009, 292, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Collins, C.; Volik, S.; Kowbel, D.; Ginzinger, D.; Ylstra, B.; Cloutier, T.; Hawkins, T.; Predki, P.; Martin, C.; Wernick, M.; et al. Comprehensive genome sequence analysis of a breast cancer amplicon. Genome Res. 2001, 11, 1034–1042. [Google Scholar] [CrossRef]
- Liu, J.; Xu, W.; Li, S.; Sun, R.; Cheng, W. Multi-omics analysis of tumor mutational burden combined with prognostic assessment in epithelial ovarian cancer based on TCGA database. Int. J. Med. Sci. 2020, 17, 3200–3213. [Google Scholar] [CrossRef]
- Liu, C.; Wei, J.; Xu, K.; Sun, X.; Zhang, H.; Xiong, C. CSE1L participates in regulating cell mitosis in human seminoma. Cell Prolif. 2019, 52, e12549. [Google Scholar] [CrossRef]
- Delpire, E.; Gagnon, K.B. SPAK and OSR1: STE20 kinases involved in the regulation of ion homoeostasis and volume control in mammalian cells. Biochem. J. 2008, 409, 321–331. [Google Scholar] [CrossRef]
- Gagnon, K.B.; Delpire, E. Molecular physiology of SPAK and OSR1: Two Ste20-related protein kinases regulating ion transport. Physiol. Rev. 2012, 92, 1577–1617. [Google Scholar] [CrossRef]
- Alessi, D.R.; Zhang, J.; Khanna, A.; Hochdörfer, T.; Shang, Y.; Kahle, K.T. The WNK-SPAK/OSR1 pathway: Master regulator of cation-chloride cotransporters. Sci. Signal. 2014, 7, re3. [Google Scholar] [CrossRef]
- Huang, T.; Zhou, Y.; Cao, Y.; Tao, J.; Zhou, Z.H.; Hang, D.H. STK39, overexpressed in osteosarcoma, regulates osteosarcoma cell invasion and proliferation. Oncol. Lett. 2017, 14, 4599–4604. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Dong, B.; Guo, W.; Piotr, R.; Longmore, G.; Yang, X.; Yu, Z.; Deng, J.; Evers, B.M.; Wu, Y. STK39 promotes breast cancer invasion and metastasis by increasing SNAI1 activity upon phosphorylation. Theranostics 2021, 11, 7658–7670. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, X.; Fang, D.; Xu, P.; Mo, X.; Hu, C.; Abdelatty, A.; Wang, M.; Xu, H.; Sun, Q.; et al. STK39 is a novel kinase contributing to the progression of hepatocellular carcinoma by the PLK1/ERK signaling pathway. Theranostics 2021, 11, 2108–2122. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Zhang, Y.; Lu, Y.; Han, G.; Rong, D.; Sun, G.; Sun, G.; Tang, W.; Wu, J.; Wang, X. STK39 enhances the progression of Cholangiocarcinoma via PI3K/AKT pathway. iScience 2021, 24, 103223. [Google Scholar] [CrossRef] [PubMed]
- Wertheimer, E.V.; Salicioni, A.M.; Liu, W.; Trevino, C.L.; Chavez, J.; Hernández-González, E.O.; Darszon, A.; Visconti, P.E. Chloride Is essential for capacitation and for the capacitation-associated increase in tyrosine phosphorylation. J. Biol. Chem. 2008, 283, 35539–35550. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | UniProtKB Entry | Protein Name | Mass | Up/Down Regulated * | Biological Function in Spermatozoa | Localization in Spermatozoa | Reference |
|---|---|---|---|---|---|---|---|
| BPHL | F1RWY7 A0A5G2R9J7 | Biphenyl hydrolase like | 34,119 | Down | Detoxification (assumed) | Midpiece, mitochondria | [56,57] |
| CSE1L | I3LPP4 A0A287BD56 I3L918 | Chromosome segregation 1-like protein (Exportin-2) (Importin-alpha re-exporter) | 110,649 64,319 109,335 | Up | Unknown | Head | [58,59] |
| GSN | P20305 A0A287A6P1 | Gelsolin | 84,775 85,683 | Down | Actin polymerization during capacitation | Head and flagellum | [60,61] |
| GSPT1 | I3LNK5 A0A287AMP7 | G1 to S phase transition protein 1 (Eukaryotic peptide chain release factor GTP-binding subunit ERF3A isoform 1) | 69,025 67,988 | Down | Unknown | Unknown | [62] |
| NDUFB7 | F1SCH1 A0A4X1V532 | NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 7 | 16,458 | Up | Oxidative phosphorylation | Midpiece and mitochondria | [63] |
| NIF3L1 | A0A287ASH3 A0A287B238 | NGG1 interacting factor 3 like 1 | 48,992 | Up | Unknown | Unknown | [64,65] |
| PFDN4 | A5GHK3 A0A5G2QBU9 A0A8W4FGC6 | Prefoldin subunit 4 | 15,270 14,692 14,712 | Down | Molecular chaperone (assumed) | Head | [66,67] |
| PGLS | A0A287APK0 A0A4X1UE53 A0A287B5V9 | 6-phosphogluconolactonase EC:3.1.1.31 | 22,496 23,495 25,491 | Up | Gluconeogenesis (assumed) | Unknown | [68] |
| PPP4C | M3VK32 | Serine/threonine-protein phosphatase EC:3.1.3.16 | 35,080 | Up | Cell signaling (assumed) | Unknown | [69,70] |
| STK39, SPAK | I3LIX4 I3LF98 A0A5G2QZU5 | Non-specific serine/threonine protein kinase 39 EC:2.7.11.1 | 55,762 52,445 63,740 | Up | Cell signaling (assumed) | Head | [71,72,73] |
| STYXL1 | A0A4X1UHH1 A0A4X1UJM6 A0A4X1UL86 | Serine/threonine/tyrosine interacting like 1 | 24,717 36,791 35,538 | Down | Cell signaling (assumed) | Unknown | [74,75] |
| TIMM10 | A0A286ZVD0 | Mitochondrial import inner membrane translocase subunit | 10,333 | Down | Protein import into mitochondria (assumed) | Midpiece and mitochondria | [76] |
| TPRG1L | F1RJA8 A0A5G2QRM0 | Tumor protein p63 regulated 1 like | 25,586 30,487 | Up | Unknown | Unknown | |
| UBXN4 | F1S0D8 A0A5G2QFE2 A0A4X1SWJ6 | UBX domain protein 4 | 56,593 52,942 56,593 | Down | Unknown | Unknown |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zigo, M.; Kerns, K.; Sutovsky, P. The Ubiquitin-Proteasome System Participates in Sperm Surface Subproteome Remodeling during Boar Sperm Capacitation. Biomolecules 2023, 13, 996. https://doi.org/10.3390/biom13060996
Zigo M, Kerns K, Sutovsky P. The Ubiquitin-Proteasome System Participates in Sperm Surface Subproteome Remodeling during Boar Sperm Capacitation. Biomolecules. 2023; 13(6):996. https://doi.org/10.3390/biom13060996
Chicago/Turabian StyleZigo, Michal, Karl Kerns, and Peter Sutovsky. 2023. "The Ubiquitin-Proteasome System Participates in Sperm Surface Subproteome Remodeling during Boar Sperm Capacitation" Biomolecules 13, no. 6: 996. https://doi.org/10.3390/biom13060996
APA StyleZigo, M., Kerns, K., & Sutovsky, P. (2023). The Ubiquitin-Proteasome System Participates in Sperm Surface Subproteome Remodeling during Boar Sperm Capacitation. Biomolecules, 13(6), 996. https://doi.org/10.3390/biom13060996