
The Role of Soluble Adenylyl Cyclase in the Regulation of Flagellar Motility in Ascidian Sperm


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
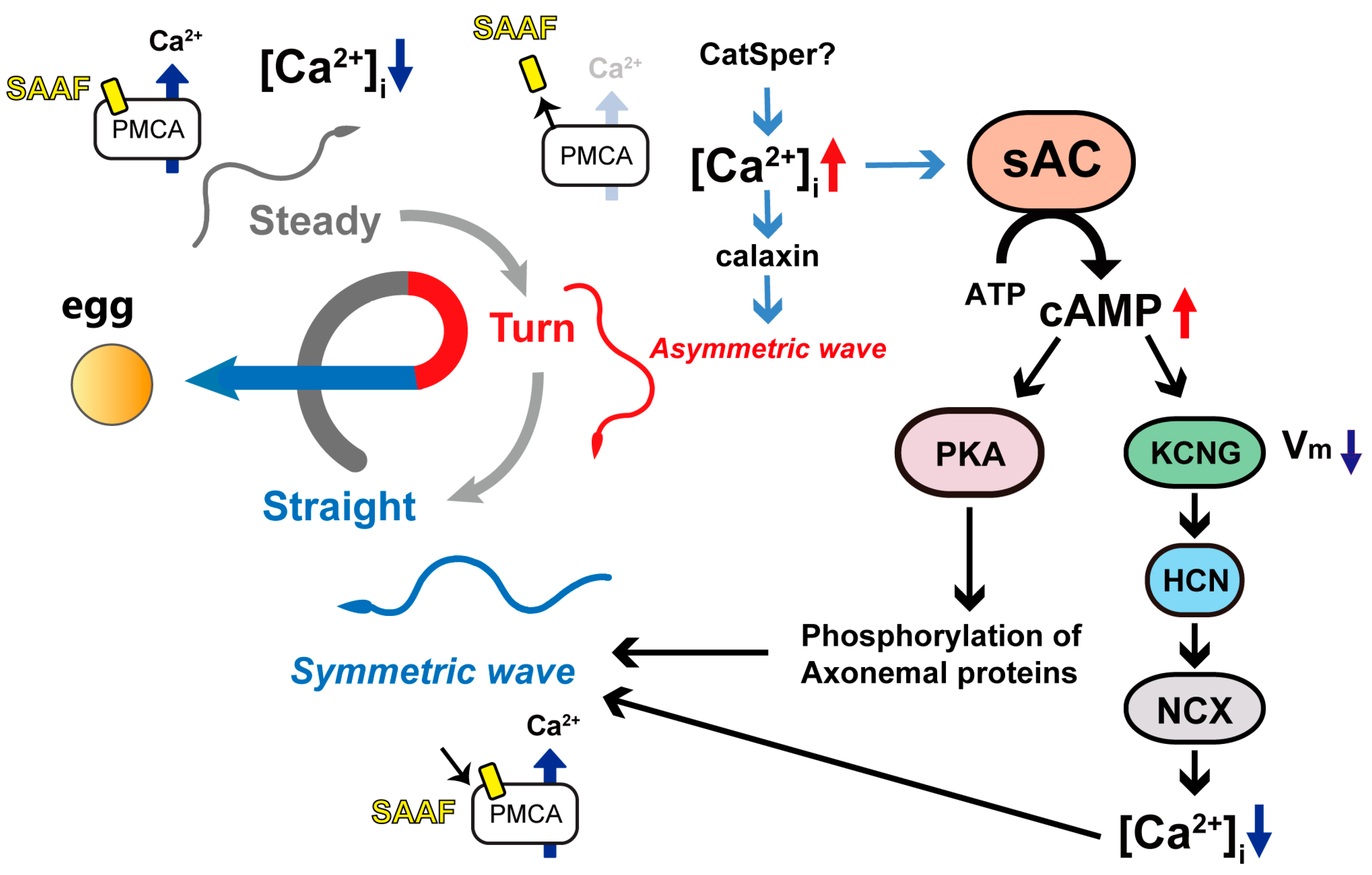
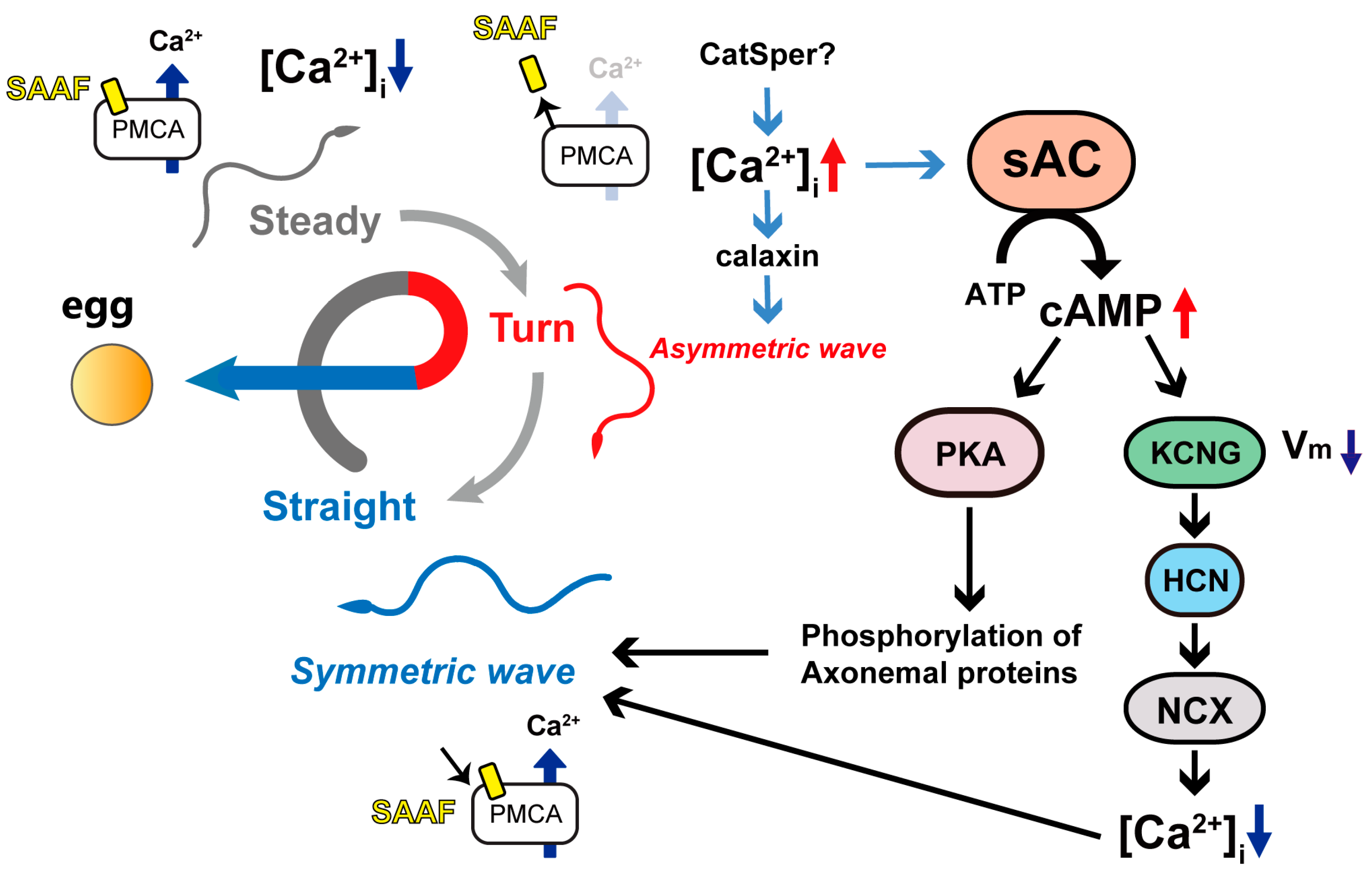
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemical and Solutions
2.3. Preparation of Antibodies and Immunoblotting
2.4. Immunofluorescence Microscopy
2.5. Analysis of Flagellar Waveforms in Sperm Chemotaxis
2.6. Imaging Analysis of [Ca2+]i
2.7. Statistical Analysis
3. Results
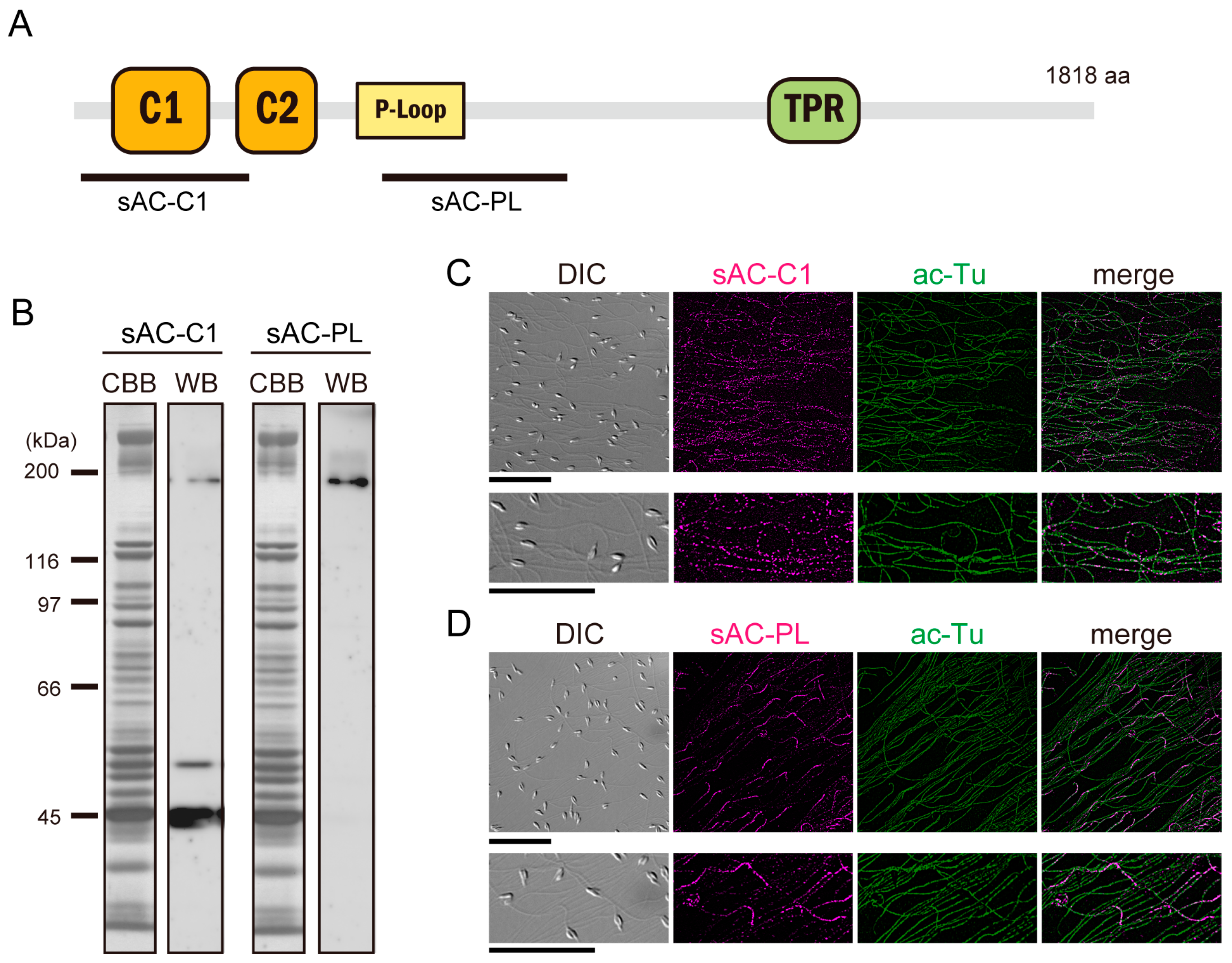
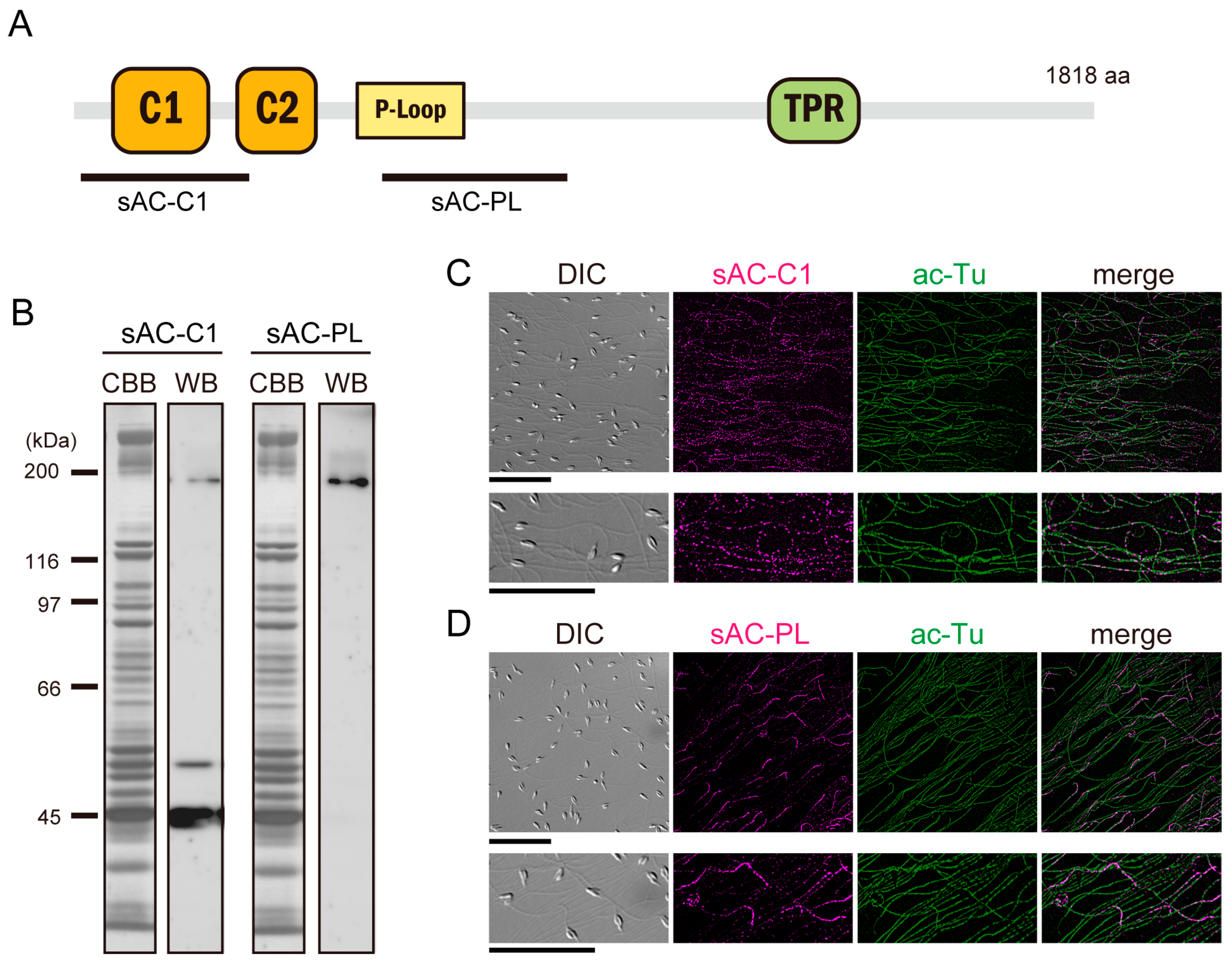
3.1. Expression of Full-Length and Truncated sAC in Ciona Sperm
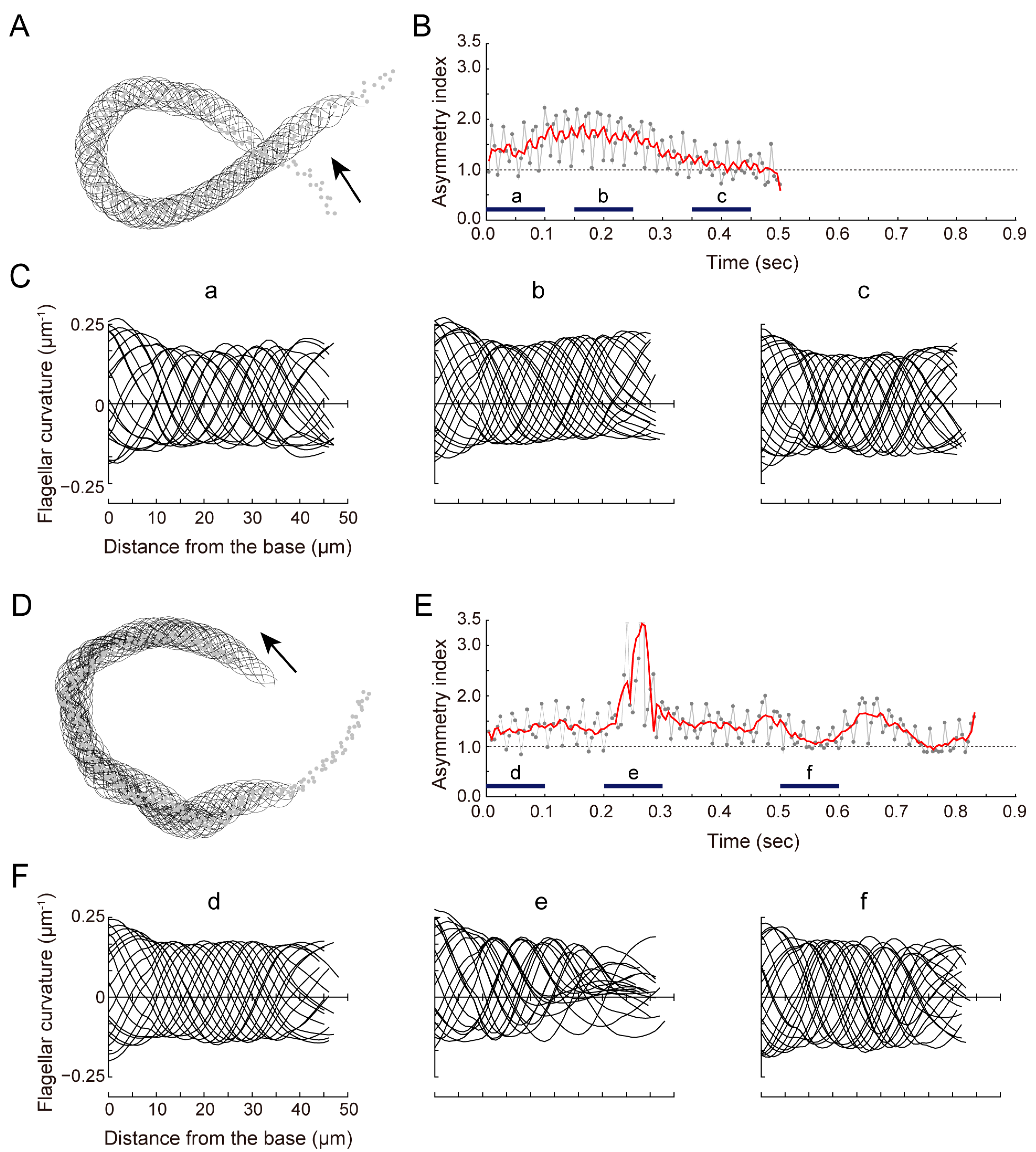
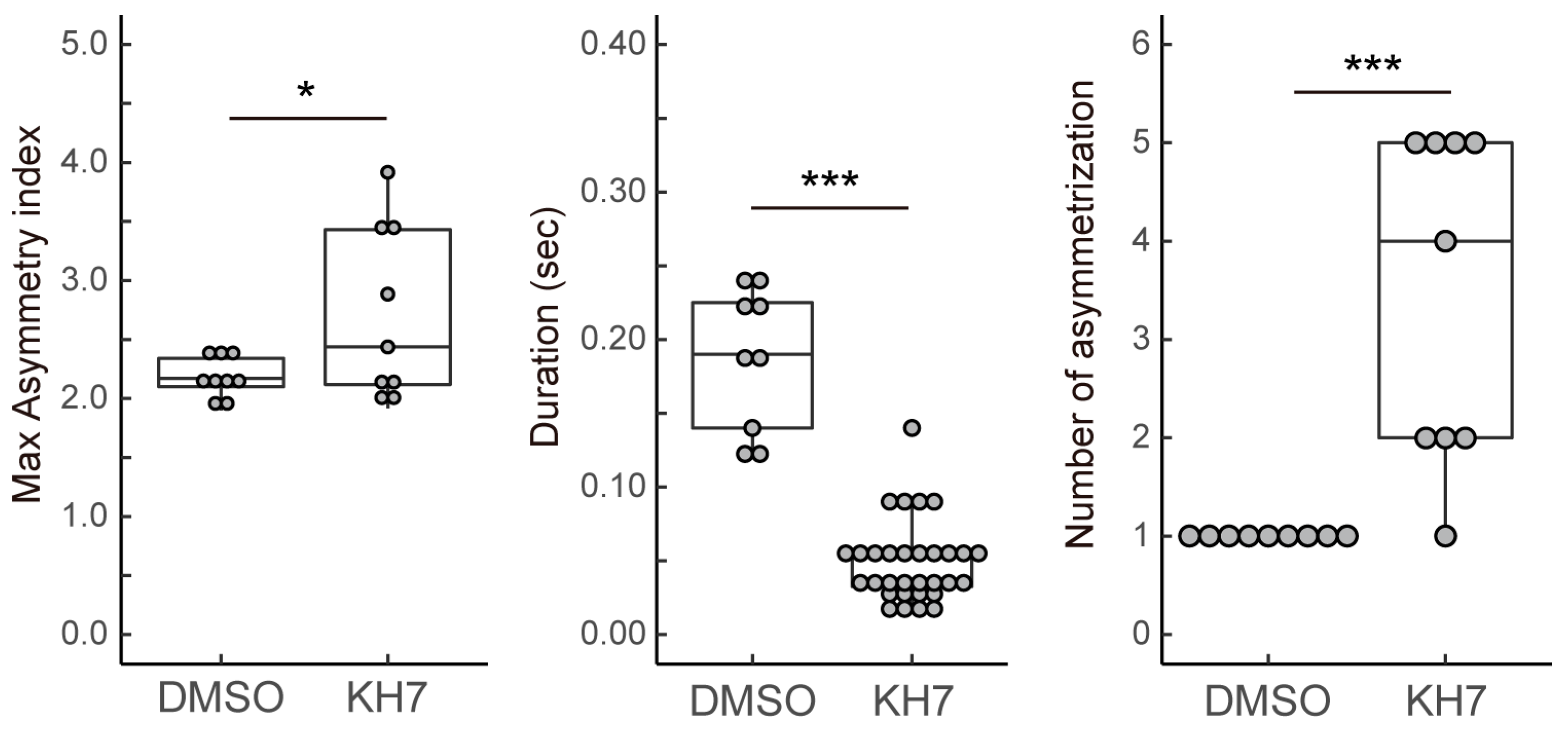
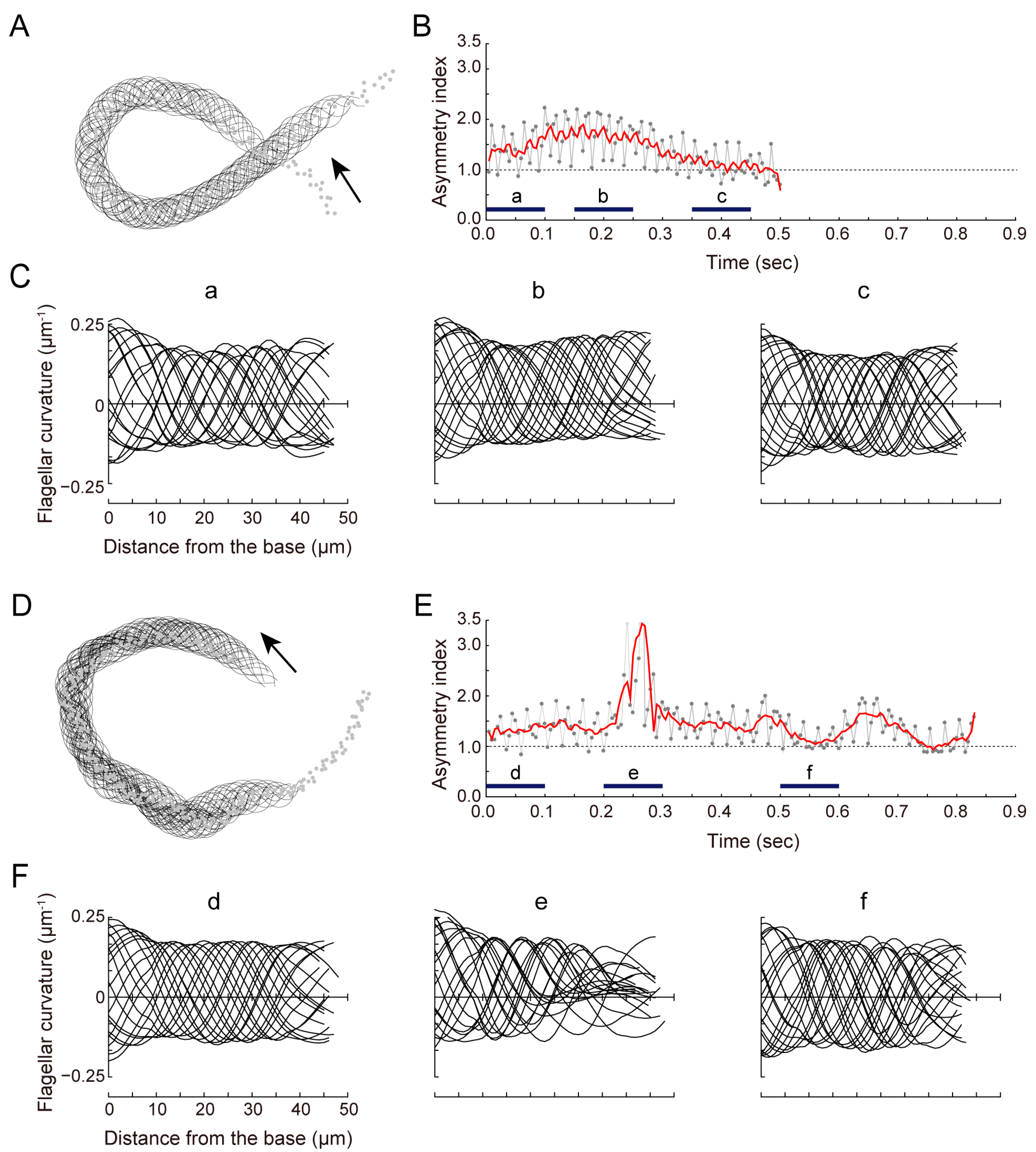
3.2. sAC Is Involved in Turn Movement during Sperm Chemotaxis
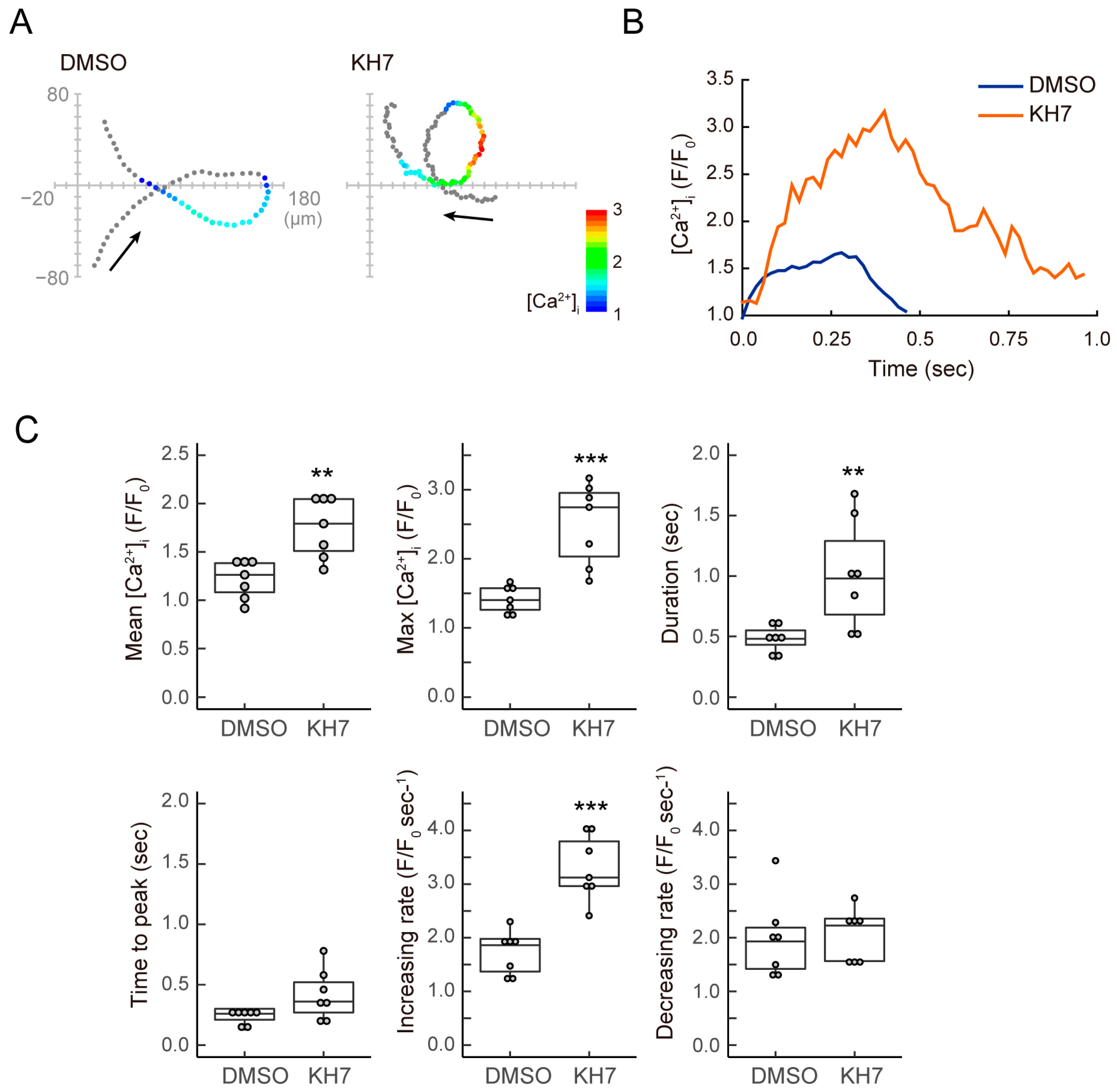
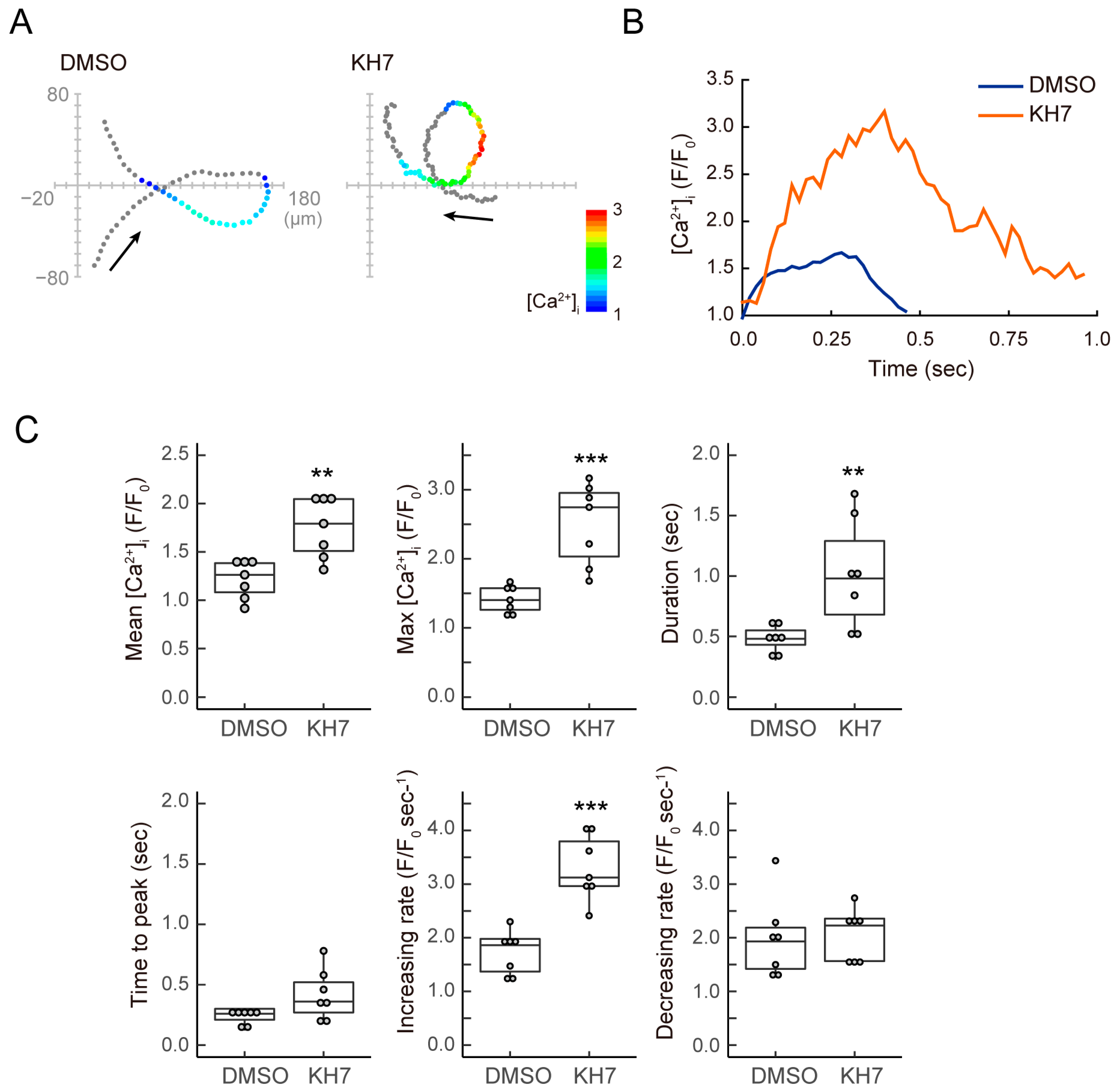
3.3. sAC Is Involved in the Dynamics of [Ca2+]i during Sperm Chemotaxis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Inaba, K. Molecular architecture of the sperm flagella: Molecules for motility and signaling. Zool. Sci. 2003, 20, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.; Nishigaki, T. Comparative genomic analysis suggests that the sperm-specific sodium/proton exchanger and soluble adenylyl cyclase are key regulators of CatSper among the Metazoa. Zool. Lett. 2019, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Speer, K.F.; Allen-Waller, L.; Novikov, D.R.; Barott, K.L. Molecular mechanisms of sperm motility are conserved in an early-branching metazoan. Proc. Natl. Acad. Sci. USA 2021, 118, e2109993118. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, B.S.; Conti, M. Calcium regulation of the soluble adenylyl cyclase expressed in mammalian spermatozoa. Proc. Natl. Acad. Sci. USA 2003, 100, 10676–10681. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Cann, M.J.; Litvin, T.N.; Iourgenko, V.; Sinclair, M.L.; Levin, L.R.; Buck, J. Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor. Science 2000, 289, 625–628. [Google Scholar] [CrossRef] [PubMed]
- Hanoune, J.; Pouille, Y.; Tzavara, E.; Shen, T.; Lipskaya, L.; Miyamoto, N.; Suzuki, Y.; Defer, N. Adenylyl cyclases: Structure, regulation and function in an enzyme superfamily. Mol. Cell. Endocrinol. 1997, 128, 179–194. [Google Scholar] [CrossRef]
- Vacquier, V.D.; Loza-Huerta, A.; Garcia-Rincon, J.; Darszon, A.; Beltran, C. Soluble adenylyl cyclase of sea urchin spermatozoa. Biochim. Biophys. Acta 2014, 1842, 2621–2628. [Google Scholar] [CrossRef]
- Esposito, G.; Jaiswal, B.S.; Xie, F.; Krajnc-Franken, M.A.; Robben, T.J.; Strik, A.M.; Kuil, C.; Philipsen, R.L.; van Duin, M.; Conti, M.; et al. Mice deficient for soluble adenylyl cyclase are infertile because of a severe sperm-motility defect. Proc. Natl. Acad. Sci. USA 2004, 101, 2993–2998. [Google Scholar] [CrossRef]
- Hess, K.C.; Jones, B.H.; Marquez, B.; Chen, Y.; Ord, T.S.; Kamenetsky, M.; Miyamoto, C.; Zippin, J.H.; Kopf, G.S.; Suarez, S.S.; et al. The “soluble” adenylyl cyclase in sperm mediates multiple signaling events required for fertilization. Dev. Cell 2005, 9, 249–259. [Google Scholar] [CrossRef]
- Wertheimer, E.; Krapf, D.; de la Vega-Beltran, J.L.; Sanchez-Cardenas, C.; Navarrete, F.; Haddad, D.; Escoffier, J.; Salicioni, A.M.; Levin, L.R.; Buck, J.; et al. Compartmentalization of distinct cAMP signaling pathways in mammalian sperm. J. Biol. Chem. 2013, 288, 35307–35320. [Google Scholar] [CrossRef]
- Balbach, M.; Ghanem, L.; Rossetti, T.; Kaur, N.; Ritagliati, C.; Ferreira, J.; Krapf, D.; Puga Molina, L.C.; Santi, C.M.; Hansen, J.N.; et al. Soluble adenylyl cyclase inhibition prevents human sperm functions essential for fertilization. Mol. Hum. Reprod. 2021, 27, gaab054. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, B.S.; Conti, M. Identification and functional analysis of splice variants of the germ cell soluble adenylyl cyclase. J. Biol. Chem. 2001, 276, 31698–31708. [Google Scholar] [CrossRef] [PubMed]
- Shiba, K.; Inaba, K. Distinct roles of soluble and transmembrane adenylyl cyclases in the regulation of flagellar motility in Ciona sperm. Int. J. Mol. Sci. 2014, 15, 13192–13208. [Google Scholar] [CrossRef]
- Yoshida, M.; Murata, M.; Inaba, K.; Morisawa, M. A chemoattractant for ascidian spermatozoa is a sulfated steroid. Proc. Natl. Acad. Sci. USA 2002, 99, 14831–14836. [Google Scholar] [CrossRef]
- Nomura, M.; Inaba, K.; Morisawa, M. Cyclic AMP- and calmodulin-dependent phosphorylation of 21 and 26 kDa proteins in axoneme is a prerequisite for SAAF-induced motile activation in ascidian spermatozoa. Dev. Growth Differ. 2000, 42, 129–138. [Google Scholar] [CrossRef]
- Hozumi, A.; Satouh, Y.; Makino, Y.; Toda, T.; Ide, H.; Ogawa, K.; King, S.M.; Inaba, K. Molecular characterization of Ciona sperm outer arm dynein reveals multiple components related to outer arm docking complex protein 2. Cell Motil Cytoskelet. 2006, 63, 591–603. [Google Scholar] [CrossRef]
- Shiba, K.; Inaba, K. The Roles of Two CNG Channels in the Regulation of Ascidian Sperm Chemotaxis. Int. J. Mol. Sci. 2022, 23, 1648. [Google Scholar] [CrossRef]
- Inaba, K.; Padma, P.; Satouh, Y.; Shin, I.T.; Kohara, Y.; Satoh, N.; Satou, Y. EST analysis of gene expression in testis of the ascidian Ciona intestinalis. Mol. Reprod. Dev. 2002, 62, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Oishi, T.; Tsuchikawa, H.; Murata, M.; Yoshida, M.; Morisawa, M. Synthesis and identification of an endogenous sperm activating and attracting factor isolated from eggs of the ascidian Ciona intestinalis; an example of nanomolar-level structure elucidation of novel natural compound. Tetrahedron 2004, 60, 6971–6980. [Google Scholar] [CrossRef]
- Oishi, T.; Tsuchikawa, H.; Murata, M.; Yoshida, M.; Morisawa, M. Synthesis of endogenous sperm-activating and attracting factor isolated from ascidian Ciona intestinalis. Tetrahedron Lett. 2003, 44, 6387–6389. [Google Scholar] [CrossRef]
- Baba, S.A.; Mogami, Y. An approach to digital image analysis of bending shapes of eukaryotic flagella and cilia. Cell Motil. 1985, 5, 475–489. [Google Scholar] [CrossRef]
- Mizuno, K.; Shiba, K.; Okai, M.; Takahashi, Y.; Shitaka, Y.; Oiwa, K.; Tanokura, M.; Inaba, K. Calaxin drives sperm chemotaxis by Ca(2)(+)-mediated direct modulation of a dynein motor. Proc. Natl. Acad. Sci. USA 2012, 109, 20497–20502. [Google Scholar] [CrossRef] [PubMed]
- Shiba, K.; Baba, S.A.; Inoue, T.; Yoshida, M. Ca2+ bursts occur around a local minimal concentration of attractant and trigger sperm chemotactic response. Proc. Natl. Acad. Sci. USA 2008, 105, 19312–19317. [Google Scholar] [CrossRef] [PubMed]
- Beltran, C.; Vacquier, V.D.; Moy, G.; Chen, Y.; Buck, J.; Levin, L.R.; Darszon, A. Particulate and soluble adenylyl cyclases participate in the sperm acrosome reaction. Biochem. Biophys. Res. Commun. 2007, 358, 1128–1135. [Google Scholar] [CrossRef]
- Chaloupka, J.A.; Bullock, S.A.; Iourgenko, V.; Levin, L.R.; Buck, J. Autoinhibitory regulation of soluble adenylyl cyclase. Mol. Reprod. Dev. 2006, 73, 361–368. [Google Scholar] [CrossRef]
- Buck, J.; Sinclair, M.L.; Schapal, L.; Cann, M.J.; Levin, L.R. Cytosolic adenylyl cyclase defines a unique signaling molecule in mammals. Proc. Natl. Acad. Sci. USA 1999, 96, 79–84. [Google Scholar] [CrossRef]
- Yoshida, K.; Shiba, K.; Sakamoto, A.; Ikenaga, J.; Matsunaga, S.; Inaba, K.; Yoshida, M. Ca2+ efflux via plasma membrane Ca2+-ATPase mediates chemotaxis in ascidian sperm. Sci. Rep. 2018, 8, 16622. [Google Scholar] [CrossRef]
- Hozumi, A.; Padma, P.; Toda, T.; Ide, H.; Inaba, K. Molecular characterization of axonemal proteins and signaling molecules responsible for chemoattractant-induced sperm activation in Ciona intestinalis. Cell Motil. Cytoskelet. 2008, 65, 249–267. [Google Scholar] [CrossRef]
- Shiba, K.; Marian, T.; Krasznai, Z.; Baba, S.A.; Morisawa, M.; Yoshida, M. Na+/Ca2+ exchanger modulates the flagellar wave pattern for the regulation of motility activation and chemotaxis in the ascidian spermatozoa. Cell Motil. Cytoskel 2006, 63, 623–632. [Google Scholar] [CrossRef]
- Shiba, K.; Baba, S.A.; Fujiwara, E.; Inaba, K. Calaxin is required for asymmetric bend initiation and propagation in sperm flagella. Front. Cell Dev. Biol. 2023, 11, 1136404. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiba, K.; Inaba, K. The Role of Soluble Adenylyl Cyclase in the Regulation of Flagellar Motility in Ascidian Sperm. Biomolecules 2023, 13, 1594. https://doi.org/10.3390/biom13111594
Shiba K, Inaba K. The Role of Soluble Adenylyl Cyclase in the Regulation of Flagellar Motility in Ascidian Sperm. Biomolecules. 2023; 13(11):1594. https://doi.org/10.3390/biom13111594
Chicago/Turabian StyleShiba, Kogiku, and Kazuo Inaba. 2023. "The Role of Soluble Adenylyl Cyclase in the Regulation of Flagellar Motility in Ascidian Sperm" Biomolecules 13, no. 11: 1594. https://doi.org/10.3390/biom13111594
APA StyleShiba, K., & Inaba, K. (2023). The Role of Soluble Adenylyl Cyclase in the Regulation of Flagellar Motility in Ascidian Sperm. Biomolecules, 13(11), 1594. https://doi.org/10.3390/biom13111594