
Activation Mechanisms and Diverse Functions of Mammalian Phospholipase C


{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
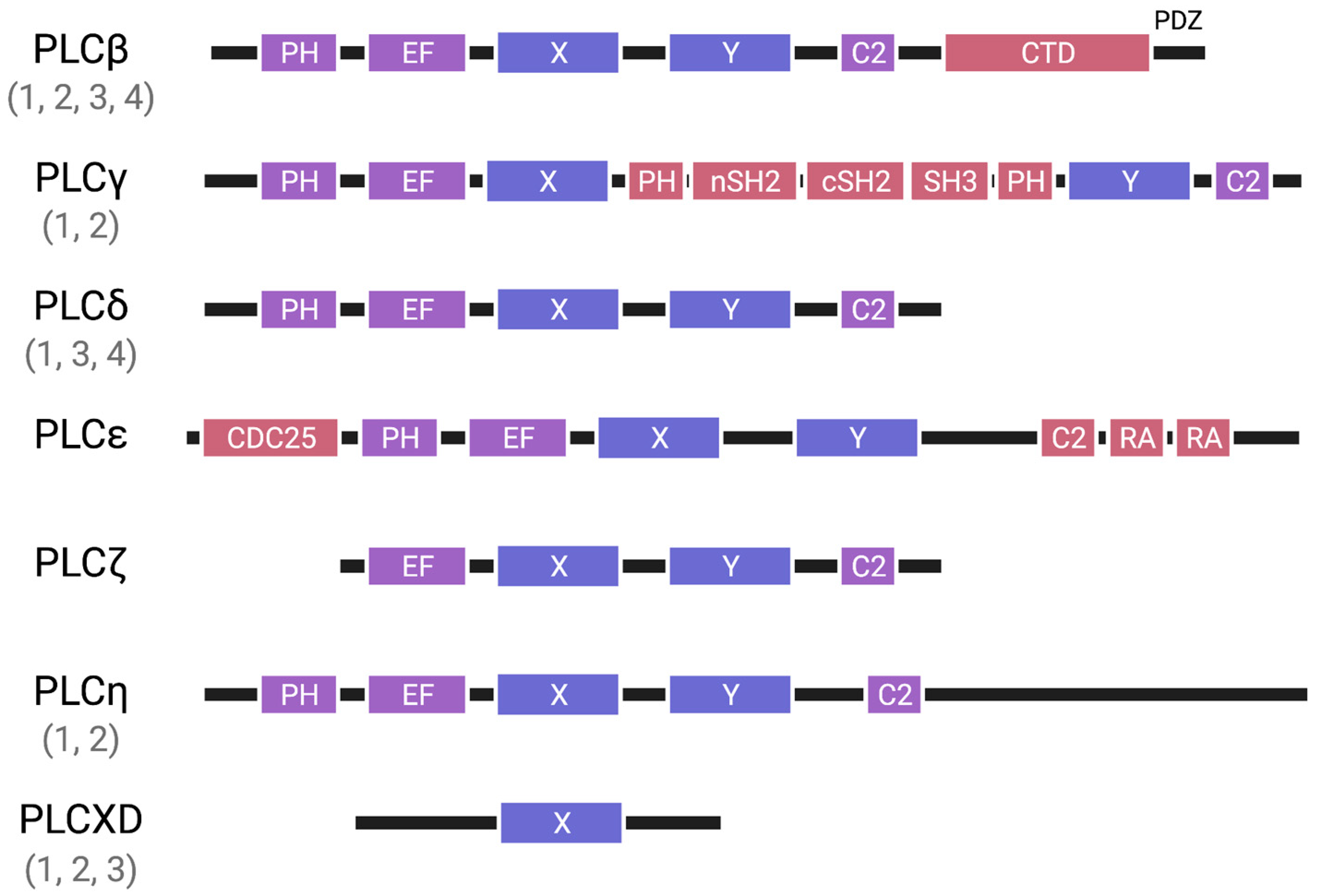
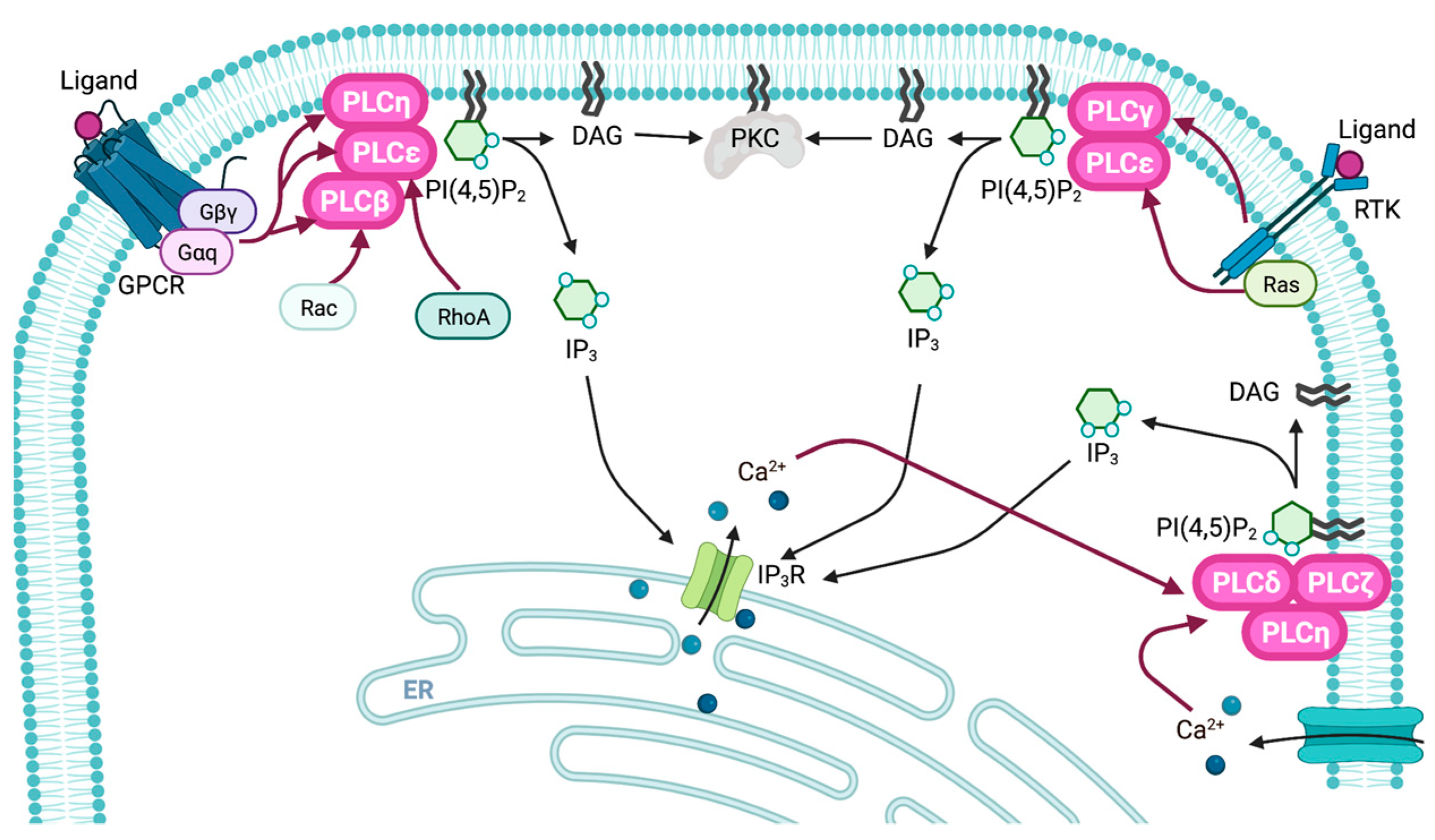
2. Regulatory Mechanisms
2.1. Regulatory Mechanisms of PLCβ
2.2. Regulatory Mechanisms of PLCγ
2.3. Regulatory Mechanisms of PLCδ
2.4. Regulatory Mechanisms of PLCε
2.5. Regulatory Mechanisms of PLCζ, PLCη, and PLCXD
3. Physiological Functions of PLC
3.1. PLCβ
3.2. PLCγ
3.3. PLCδ
3.4. PLCε
3.5. PLCζ
4. Chemical Inhibitors and Activators for PLC
5. Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berridge, M.J.; Irvine, R.F. Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature 1984, 312, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y. The molecular heterogeneity of protein kinase C and its implications for cellular regulation. Nature 1988, 334, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Katan, M.; Cockcroft, S. Phospholipase C families: Common themes and versatility in physiology and pathology. Prog. Lipid Res. 2020, 80, 101065. [Google Scholar] [PubMed]
- Suh, P.G.; Park, J.I.; Manzoli, L.; Cocco, L.; Peak, J.C.; Katan, M.; Fukami, K.; Kataoka, T.; Yun, S.; Ryu, S.H. Multiple roles of phosphoinositide-specific phospholipase C isozymes. BMB Rep. 2008, 41, 415–434. [Google Scholar] [CrossRef]
- Nakamura, Y.; Fukami, K. Regulation and physiological functions of mammalian phospholipase C. J. Biochem. 2017, 161, 315–321. [Google Scholar] [CrossRef]
- Gellatly, S.A.; Kalujnaia, S.; Cramb, G. Cloning, tissue distribution and sub-cellular localisation of phospholipase C X-domain containing protein (PLCXD) isoforms. Biochem. Biophys. Res. Commun. 2012, 424, 651–656. [Google Scholar] [CrossRef]
- Hofmann, T.; Obukhov, A.G.; Schaefer, M.; Harteneck, C.; Gudermann, T.; Schultz, G. Direct activation of human TRPC6 and TRPC3 channels by diacylglycerol. Nature 1999, 397, 259–263. [Google Scholar] [CrossRef]
- Chakraborty, A.; Kim, S.; Snyder, S.H. Inositol pyrophosphates as mammalian cell signals. Sci. Signal. 2011, 4, re1. [Google Scholar] [CrossRef]
- Irvine, R.F.; Schell, M.J. Back in the water: The return of the inositol phosphates. Nat. Rev. Mol. Cell. Biol. 2001, 2, 327–338. [Google Scholar] [CrossRef]
- Laha, D.; Portela-Torres, P.; Desfougères, Y.; Saiardi, A. Inositol phosphate kinases in the eukaryote landscape. Adv. Biol. Regul. 2021, 79, 100782. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kim, Y.R.; Park, J.; Kim, S. Inositol polyphosphate multikinase signaling in the regulation of metabolism. Ann. N. Y. Acad. Sci. 2012, 1271, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Mulugu, S.; Bai, W.; Fridy, P.C.; Bastidas, R.J.; Otto, J.C.; Dollins, D.E.; Haystead, T.A.; Ribeiro, A.A.; York, J.D. A conserved family of enzymes that phosphorylate inositol hexakisphosphate. Science 2007, 316, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Mulugu, S.; York, J.D.; O’Shea, E.K. Regulation of a cyclin-CDK-CDK inhibitor complex by inositol pyrophosphates. Science 2007, 316, 109–112. [Google Scholar] [CrossRef]
- Monserrate, J.P.; York, J.D. Inositol phosphate synthesis and the nuclear processes they affect. Curr. Opin. Cell. Biol. 2010, 22, 365–373. [Google Scholar] [CrossRef]
- Thakur, R.; Naik, A.; Panda, A.; Raghu, P. Regulation of membrane turnover by phosphatidic Acid: Cellular Functions and Disease Implications. Front. Cell. Dev. Biol. 2019, 7, 83. [Google Scholar] [CrossRef]
- Barneda, D.; Cosulich, S.; Stephens, L.; Hawkins, P. How is the acyl chain composition of phosphoinositides created and does it matter? Biochem. Soc. Trans. 2019, 47, 1291–1305. [Google Scholar] [CrossRef]
- Murataeva, N.; Straiker, A.; Mackie, K. Parsing the players: 2-arachidonoylglycerol synthesis and degradation in the CNS. Br. J. Pharmacol. 2014, 171, 1379–1391. [Google Scholar] [CrossRef]
- Tong, J.; Liu, X.; Vickstrom, C.; Li, Y.; Yu, L.; Lu, Y.; Smrcka, A.V.; Liu, Q.S. The Epac-Phospholipase Cε Pathway Regulates Endocannabinoid Signaling and Cocaine-Induced Disinhibition of Ventral Tegmental Area Dopamine Neurons. J. Neurosci. 2017, 37, 3030–3044. [Google Scholar] [CrossRef] [PubMed]
- Di Paolo, G.; De Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef]
- Janetopoulos, C.; Devreotes, P. Phosphoinositide signaling plays a key role in cytokinesis. J. Cell. Biol. 2006, 174, 485–490. [Google Scholar] [CrossRef]
- Martin, T.F. PI(4,5)P(2) regulation of surface membrane traffic. Curr. Opin. Cell. Biol. 2001, 13, 493–499. [Google Scholar] [CrossRef]
- Senju, Y.; Lappalainen, P. Regulation of actin dynamics by PI(4,5)P2 in cell migration and endocytosis. Curr. Opin. Cell. Biol. 2019, 56, 7–13. [Google Scholar] [CrossRef]
- Kanemaru, K.; Shimozawa, M.; Kitamata, M.; Furuishi, R.; Kayano, H.; Sukawa, Y.; Chiba, Y.; Fukuyama, T.; Hasegawa, J.; Nakanishi, H.; et al. Plasma membrane phosphatidylinositol (4,5)-bisphosphate is critical for determination of epithelial characteristics. Nat. Commun. 2022, 13, 2347. [Google Scholar] [CrossRef] [PubMed]
- Manning, B.D.; Cantley, L.C. AKT/PKB signaling: Navigating downstream. Cell 2007, 129, 1261–1274. [Google Scholar] [CrossRef] [PubMed]
- Ellis, M.V.; James, S.R.; Perisic, O.; Downes, C.P.; Williams, R.L.; Katan, M. Catalytic domain of phosphoinositide-specific phospholipase C (PLC). Mutational analysis of residues within the active site and hydrophobic ridge of plcdelta1. J. Biol. Chem. 1998, 273, 11650–11659. [Google Scholar] [CrossRef]
- Zhang, L.; Malik, S.; Pang, J.; Wang, H.; Park, K.M.; Yule, D.I.; Blaxall, B.C.; Smrcka, A.V. Phospholipase Cε hydrolyzes perinuclear phosphatidylinositol 4-phosphate to regulate cardiac hypertrophy. Cell 2013, 153, 216–227. [Google Scholar] [CrossRef]
- Divecha, N.; Banfić, H.; Irvine, R.F. The polyphosphoinositide cycle exists in the nuclei of Swiss 3T3 cells under the control of a receptor (for IGF-I) in the plasma membrane, and stimulation of the cycle increases nuclear diacylglycerol and apparently induces translocation of protein kinase C to the nucleus. EMBO J. 1991, 10, 3207–3214. [Google Scholar]
- Manzoli, L.; Billi, A.M.; Rubbini, S.; Bavelloni, A.; Faenza, I.; Gilmour, R.S.; Rhee, S.G.; Cocco, L. Essential role for nuclear phospholipase C beta1 in insulin-like growth factor I-induced mitogenesis. Cancer Res. 1997, 57, 2137–2139. [Google Scholar] [PubMed]
- Bahk, Y.Y.; Song, H.; Baek, S.H.; Park, B.Y.; Kim, H.; Ryu, S.H.; Suh, P.G. Localization of two forms of phospholipase C-beta1, a and b, in C6Bu-1 cells. Biochim. Biophys. Acta 1998, 1389, 76–80. [Google Scholar] [CrossRef]
- Martelli, A.M.; Fiume, R.; Faenza, I.; Tabellini, G.; Evangelista, C.; Bortul, R.; Follo, M.Y.; Falà, F.; Cocco, L. Nuclear phosphoinositide specific phospholipase C (PI-PLC)-beta 1: A central intermediary in nuclear lipid-dependent signal transduction. Histol. Histopathol. 2005, 20, 1251–1260. [Google Scholar]
- Martelli, A.M.; Follo, M.Y.; Evangelisti, C.; Falà, F.; Fiume, R.; Billi, A.M.; Cocco, L. Nuclear inositol lipid metabolism: More than just second messenger generation? J. Cell. Biochem. 2005, 96, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Gensburger, C.; Freyermuth, S.; Nair, B.C.; Labourdette, G.; Malviya, A.N. A 120 kDa nuclear phospholipase Cgamma1 protein fragment is stimulated in vivo by EGF signal phosphorylating nuclear membrane EGFR. Biochemistry 2004, 43, 15873–15883. [Google Scholar] [CrossRef] [PubMed]
- Yagisawa, H.; Okada, M.; Naito, Y.; Sasaki, K.; Yamaga, M.; Fujii, M. Coordinated intracellular translocation of phosphoinositide-specific phospholipase C-delta with the cell cycle. Biochim. Biophys. Acta 2006, 1761, 522–534. [Google Scholar] [CrossRef]
- Okada, M.; Ishimoto, T.; Naito, Y.; Hirata, H.; Yagisawa, H. Phospholipase Cdelta1 associates with importin beta1 and translocates into the nucleus in a Ca2+-dependent manner. FEBS Lett. 2005, 579, 4949–4954. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Fukami, K.; Yu, H.; Takenawa, T. A new phospholipase C delta 4 is induced at S-phase of the cell cycle and appears in the nucleus. J. Biol. Chem. 1996, 271, 355–360. [Google Scholar] [CrossRef]
- Kunrath-Lima, M.; de Miranda, M.C.; Ferreira, A.D.F.; Faraco, C.C.F.; de Melo, M.I.A.; Goes, A.M.; Rodrigues, M.A.; Faria, J.A.Q.A.; Gomes, D.A. Phospholipase C delta 4 (PLCδ4) is a nuclear protein involved in cell proliferation and senescence in mesenchymal stromal stem cells. Cell. Signal. 2018, 49, 59–67. [Google Scholar] [CrossRef]
- Hicks, S.N.; Jezyk, M.R.; Gershburg, S.; Seifert, J.P.; Harden, T.K.; Sondek, J. General and versatile autoinhibition of PLC isozymes. Mol. Cell. 2008, 31, 383–394. [Google Scholar] [CrossRef]
- Philip, F.; Kadamur, G.; Silos, R.G.; Woodson, J.; Ross, E.M. Synergistic activation of phospholipase C-beta3 by Galpha(q) and Gbetagamma describes a simple two-state coincidence detector. Curr. Biol. 2010, 20, 1327–1335. [Google Scholar] [CrossRef]
- Smrcka, A.V.; Sternweis, P.C. Regulation of purified subtypes of phosphatidylinositol-specific phospholipase C beta by G protein alpha and beta gamma subunits. J. Biol. Chem. 1993, 268, 9667–9674. [Google Scholar] [CrossRef]
- Jezyk, M.R.; Snyder, J.T.; Gershberg, S.; Worthylake, D.K.; Harden, T.K.; Sondek, J. Crystal structure of Rac1 bound to its effector phospholipase C-beta2. Nat. Struct. Mol. Biol. 2006, 13, 1135–1140. [Google Scholar] [CrossRef]
- Illenberger, D.; Walliser, C.; Nurnberg, B.; Diaz Lorente, M.; Gierschik, P. Specificity and structural requirements of phospholipase C-beta stimulation by Rho GTPases versus G protein beta gamma dimers. J. Biol. Chem. 2003, 278, 3006–3014. [Google Scholar] [CrossRef]
- Lyon, A.M.; Dutta, S.; Boguth, C.A.; Skiniotis, G.; Tesmer, J.J. Full-length Gα(q)-phospholipase C-β3 structure reveals interfaces of the C-terminal coiled-coil domain. Nat. Struct. Mol. Biol. 2013, 20, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Fisher, I.J.; Jenkins, M.L.; Tall, G.G.; Burke, J.E.; Smrcka, A.V. Activation of Phospholipase C β by Gβγ and Gαq Involves C-Terminal Rearrangement to Release Autoinhibition. Structure 2020, 28, 810–819.e5. [Google Scholar] [CrossRef] [PubMed]
- Inaba, T.; Kishimoto, T.; Murate, M.; Tajima, T.; Sakai, S.; Abe, M.; Makino, A.; Tomishige, N.; Ishitsuka, R.; Ikeda, Y.; et al. Phospholipase Cβ1 induces membrane tubulation and is involved in caveolae formation. Proc. Natl. Acad. Sci. USA 2016, 113, 7834–7839. [Google Scholar] [CrossRef]
- Oh, Y.S.; Jo, N.W.; Choi, J.W.; Kim, H.S.; Seo, S.W.; Kang, K.O.; Hwang, J.I.; Heo, K.; Kim, S.H.; Kim, Y.H.; et al. NHERF2 specifically interacts with LPA2 receptor and defines the specificity and efficiency of receptor-mediated phospholipase C-beta3 activation. Mol. Cell. Biol. 2004, 24, 5069–5079. [Google Scholar] [CrossRef]
- Berstein, G.; Blank, J.L.; Jhon, D.Y.; Exton, J.H.; Rhee, S.G.; Ross, E.M. Phospholipase C-beta 1 is a GTPase-activating protein for Gq/11, its physiologic regulator. Cell 1992, 70, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Kim, J.W.; Zilberstein, A.; Margolis, B.; Kim, J.G.; Schlessinger, J.; Rhee, S.G. PDGF stimulation of inositol phospholipid hydrolysis requires PLC-gamma 1 phosphorylation on tyrosine residues 783 and 1254. Cell 1991, 65, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.I.; Daniel, T.O.; Carpenter, G. Antiphosphotyrosine recovery of phospholipase C activity after EGF treatment of A-431 cells. Science 1988, 241, 968–970. [Google Scholar] [CrossRef]
- Law, C.L.; Chandran, K.A.; Sidorenko, S.P.; Clark, E.A. Phospholipase C-gamma1 interacts with conserved phosphotyrosyl residues in the linker region of Syk and is a substrate for Syk. Mol. Cell. Biol. 1996, 16, 1305–1315. [Google Scholar] [CrossRef]
- Nakanishi, O.; Shibasaki, F.; Hidaka, M.; Homma, Y.; Takenawa, T. Phospholipase C-gamma 1 associates with viral and cellular src kinases. J. Biol. Chem. 1993, 268, 10754–10759. [Google Scholar] [CrossRef]
- Schaeffer, E.M.; Debnath, J.; Yap, G.; McVicar, D.; Liao, X.C.; Littman, D.R.; Sher, A.; Varmus, H.E.; Lenardo, M.J.; Schwartzberg, P.L. Requirement for Tec kinases Rlk and Itk in T cell receptor signaling and immunity. Science 1999, 284, 638–641. [Google Scholar] [CrossRef]
- Bae, J.H.; Lew, E.D.; Yuzawa, S.; Tomé, F.; Lax, I.; Schlessinger, J. The selectivity of receptor tyrosine kinase signaling is controlled by a secondary SH2 domain binding site. Cell 2009, 138, 514–524. [Google Scholar] [CrossRef] [PubMed]
- Hajicek, N.; Keith, N.C.; Siraliev-Perez, E.; Temple, B.R.; Huang, W.; Zhang, Q.; Harden, T.K.; Sondek, J. Structural basis for the activation of PLC-γ isozymes by phosphorylation and cancer-associated mutations. eLife 2019, 8, e51700. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bunney, T.D.; Khosa, S.; Macé, K.; Beckenbauer, K.; Askwith, T.; Maslen, S.; Stubbs, C.; de Oliveira, T.M.; Sader, K.; et al. Structural insights and activating mutations in diverse pathologies define mechanisms of deregulation for phospholipase C gamma enzymes. EBioMedicine 2020, 51, 102607. [Google Scholar] [CrossRef] [PubMed]
- Piechulek, T.; Rehlen, T.; Walliser, C.; Vatter, P.; Moepps, B.; Gierschik, P. Isozyme-specific stimulation of phospholipase C-gamma2 by Rac GTPases. J. Biol. Chem. 2005, 280, 38923–38931. [Google Scholar] [CrossRef]
- Walliser, C.; Retlich, M.; Harris, R.; Everett, K.L.; Josephs, M.B.; Vatter, P.; Esposito, D.; Driscoll, P.C.; Katan, M.; Gierschik, P.; et al. rac regulates its effector phospholipase Cgamma2 through interaction with a split pleckstrin homology domain. J. Biol. Chem. 2008, 283, 30351–30362. [Google Scholar] [CrossRef]
- Bae, Y.S.; Cantley, L.G.; Chen, C.S.; Kim, S.R.; Kwon, K.S.; Rhee, S.G. Activation of phospholipase C-gamma by phosphatidylinositol 3,4,5-trisphosphate. J. Biol. Chem. 1998, 273, 4465–4469. [Google Scholar] [CrossRef]
- Falasca, M.; Logan, S.K.L.; Lehto, V.P.; Baccante, G.; Lemmon, M.A.; Schlessinger, J. Activation of phospholipase C gamma by PI 3-kinase-induced PH domain-mediated membrane targeting. EMBO J. 1998, 17, 414–422. [Google Scholar] [CrossRef]
- Gratacap, M.P.; Payrastre, B.; Viala, C.; Mauco, G.; Plantavid, M.; Chap, H. Phosphatidylinositol 3,4,5-trisphosphate-dependent stimulation of phospholipase C-gamma2 is an early key event in FcgammaRIIA-mediated activation of human platelets. J. Biol. Chem. 1998, 273, 24314–24321. [Google Scholar] [CrossRef] [PubMed]
- Allen, V.; Swigart, P.; Cheung, R.; Cockcroft, S.; Katan, M. Regulation of inositol lipid-specific phospholipase cdelta by changes in Ca2+ ion concentrations. Biochem. J. 1997, 327, 545–552. [Google Scholar] [CrossRef]
- Kim, Y.H.; Park, T.J.; Lee, Y.H.; Baek, K.J.; Suh, P.G.; Ryu, S.H.; Kim, K.T. Phospholipase C-delta1 is activated by capacitative calcium entry that follows phospholipase C-beta activation upon bradykinin stimulation. J. Biol. Chem. 1999, 274, 26127–26134. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, K.M.; Lemmon, M.A.; Schlessinger, J.; Sigler, P.B. Structure of the high affinity complex of inositol trisphosphate with a phospholipase C pleckstrin homology domain. Cell 1995, 83, 1037–1046. [Google Scholar] [CrossRef]
- Harlan, J.E.; Hajduk, P.J.; Yoon, H.S.; Fesik, S.W. Pleckstrin homology domains bind to phosphatidylinositol-4,5-bisphosphate. Nature 1994, 371, 168–170. [Google Scholar] [CrossRef]
- Hirose, K.; Kadowaki, S.; Tanabe, M.; Takeshima, H.; Iino, M. Spatiotemporal dynamics of inositol 1,4,5-trisphosphate that underlies complex Ca2+ mobilization patterns. Science 1999, 284, 1527–1530. [Google Scholar] [CrossRef]
- Yagisawa, H.; Sakuma, K.; Paterson, H.F.; Cheung, R.; Allen, V.; Hirata, H.; Watanabe, Y.; Hirata, M.; Williams, R.L.; Katan, M. Replacements of single basic amino acids in the pleckstrin homology domain of phospholipase C-delta1 alter the ligand binding, phospholipase activity, and interaction with the plasma membrane. J. Biol. Chem. 1998, 273, 417–424. [Google Scholar] [CrossRef]
- Feng, J.F.; Rhee, S.G.; Im, M.J. Evidence that phospholipase delta1 is the effector in the Gh (transglutaminase II)-mediated signaling. J. Biol. Chem. 1996, 271, 16451–16454. [Google Scholar] [CrossRef]
- Sidhu, R.S.; Clough, R.R.; Bhullar, R.P. Regulation of phospholipase C-delta1 through direct interactions with the small GTPase Ral and calmodulin. J. Biol. Chem. 2005, 280, 21933–21941. [Google Scholar] [CrossRef] [PubMed]
- Kelley, G.G.; Reks, S.E.; Smrcka, A.V. Hormonal regulation of phospholipase Cepsilon through distinct and overlapping pathways involving G12 and Ras family G-proteins. Biochem. J. 2004, 378, 129–139. [Google Scholar] [CrossRef]
- Bunney, T.D.; Katan, M. Phospholipase C epsilon: Linking second messengers and small GTPases. Trends Cell. Biol. 2006, 16, 640–648. [Google Scholar] [CrossRef]
- Wing, M.R.; Houston, D.; Kelley, G.G.; Der, C.J.; Siderovski, D.P.; Harden, T.K. Activation of phospholipase C-epsilon by heterotrimeric G protein betagamma-subunits. J. Biol. Chem. 2001, 276, 48257–48261. [Google Scholar] [CrossRef] [PubMed]
- Bunney, T.D.; Harris, R.; Gandarillas, N.L.; Josephs, M.B.; Roe, S.M.; Sorli, S.C.; Paterson, H.F.; Rodrigues-Lima, F.; Esposito, D.; Ponting, C.P.; et al. Structural and mechanistic insights into ras association domains of phospholipase C epsilon. Mol. Cell. 2006, 21, 495–507. [Google Scholar] [CrossRef] [PubMed]
- Kelley, G.G.; Reks, S.E.; Ondrako, J.M.; Smrcka, A.V. Phospholipase C(epsilon): A novel Ras effector. EMBO J. 2001, 20, 743–754. [Google Scholar] [CrossRef]
- Song, C.; Hu, C.D.; Masago, M.; Kariyai, K.; Yamawaki-Kataoka, Y.; Shibatohge, M.; Wu, D.; Satoh, T.; Kataoka, T. Regulation of a novel human phospholipase C, PLCepsilon, through membrane targeting by Ras. J. Biol. Chem. 2001, 276, 2752–2757. [Google Scholar] [CrossRef]
- Jin, T.G.; Satoh, T.; Liao, Y.; Song, C.; Gao, X.; Kariya, K.; Hu, C.D.; Kataoka, T. Role of the CDC25 homology domain of phospholipase Cepsilon in amplification of Rap1-dependent signaling. J. Biol. Chem. 2001, 276, 30301–30307. [Google Scholar] [CrossRef] [PubMed]
- Wing, M.R.; Snyder, J.T.; Sondek, J.; Harden, T.K. Direct activation of phospholipase C-epsilon by Rho. J. Biol. Chem. 2003, 278, 41253–41258. [Google Scholar] [CrossRef] [PubMed]
- Lopez, I.; Mak, E.C.; Ding, J.; Hamm, H.E.; Lomasney, J.W. A novel bifunctional phospholipase c that is regulated by Galpha 12 and stimulates the Ras/mitogen-activated protein kinase pathway. J. Biol. Chem. 2001, 276, 2758–2765. [Google Scholar] [CrossRef]
- Nomikos, M.; Kashir, J.; Lai, F.A. The role and mechanism of action of sperm PLC-zeta in mammalian fertilization. Biochem. J. 2017, 474, 3659–3673. [Google Scholar] [CrossRef]
- Nomikos, M.; Sanders, J.R.; Parthimos, D.; Buntwal, L.; Calver, B.L.; Stamatiadis, P.; Smith, A.; Clue, M.; Sideratou, Z.; Swann, K.; et al. Essential Role of the EF-hand Domain in Targeting Sperm Phospholipase Cζ to Membrane Phosphatidylinositol 4,5-Bisphosphate (PIP2). J. Biol. Chem. 2015, 290, 29519–29530. [Google Scholar] [CrossRef]
- Yu, Y.; Nomikos, M.; Theodoridou, M.; Nounesis, G.; Lai, F.A.; Swann, K. PLCζ causes Ca(2+) oscillations in mouse eggs by targeting intracellular and not plasma membrane PI(4,5)P(2). Mol. Biol. Cell. 2012, 23, 371–380. [Google Scholar] [CrossRef]
- Nakahara, M.; Shimozawa, M.; Nakamura, Y.; Irino, Y.; Morita, M.; Kudo, Y.; Fukami, K. A novel phospholipase C, PLC(eta)2, is a neuron-specific isozyme. J. Biol. Chem. 2005, 280, 29128–29134. [Google Scholar] [CrossRef]
- Popovics, P.; Lu, J.; Nadia Kamil, L.; Morgan, K.; Millar, R.P.; Schmid, R.; Blindauer, C.A.; Stewart, A.J. A canonical EF-loop directs Ca(2+) -sensitivity in phospholipase C-η2. J. Cell. Biochem. 2014, 115, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wing, M.R.; Sondek, J.; Harden, T.K. Molecular cloning and characterization of PLC-eta2. Biochem. J. 2005, 391, 667–676. [Google Scholar] [CrossRef]
- Zhou, Y.; Sondek, J.; Harden, T.K. Activation of human phospholipase C-eta2 by Gbetagamma. Biochemistry 2008, 47, 4410–4417. [Google Scholar] [CrossRef] [PubMed]
- Böhm, D.; Schwegler, H.; Kotthaus, L.; Nayernia, K.; Rickmann, M.; Köhler, M.; Rosenbusch, J.; Engel, W.; Flügge, G.; Burfeind, P. Disruption of PLC-beta 1-mediated signal transduction in mutant mice causes age-dependent hippocampal mossy fiber sprouting and neurodegeneration. Mol. Cell. Neurosci. 2002, 21, 584–601. [Google Scholar] [CrossRef]
- Desprairies, C.; Valence, S.; Maurey, H.; Helal, S.I.; Weckhuysen, S.; Soliman, H.; Mefford, H.C.; Spentchian, M.; Héron, D.; Leguern, E.; et al. Three novel patients with epileptic encephalopathy due to biallelic mutations in the PLCB1 gene. Clin. Genet. 2020, 97, 477–482. [Google Scholar] [CrossRef]
- Kurian, M.A.; Meyer, E.; Vassallo, G.; Morgan, N.V.; Prakash, N.; Pasha, S.; Hai, N.A.; Shuib, S.; Rahman, F.; Wassmer, E.; et al. Phospholipase C beta 1 deficiency is associated with early-onset epileptic encephalopathy. Brain 2010, 133, 2964–2970. [Google Scholar] [CrossRef]
- Hwang, H.J.; Yang, Y.R.; Kim, H.Y.; Choi, Y.; Park, K.S.; Lee, H.; Ma, J.S.; Yamamoto, M.; Kim, J.; Chae, Y.C.; et al. Phospholipase C-β1 potentiates glucose-stimulated insulin secretion. FASEB J. 2019, 33, 10668–10679. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.J.; Jang, H.J.; Cocco, L.; Suh, P.G. The regulation of insulin secretion via phosphoinositide-specific phospholipase Cβ signaling. Adv. Biol. Regul. 2019, 71, 10–18. [Google Scholar] [CrossRef]
- Ratti, S.; Marvi, M.V.; Mongiorgi, S.; Obeng, E.O.; Rusciano, I.; Ramazzotti, G.; Morandi, L.; Asioli, S.; Zoli, M.; Mazzatenta, D.; et al. Impact of phospholipase C β1 in glioblastoma: A study on the main mechanisms of tumor aggressiveness. Cell. Mol. Life Sci. 2022, 79, 195. [Google Scholar] [CrossRef]
- Jiang, H.; Kuang, Y.; Wu, Y.; Xie, W.; Simon, M.I.; Wu, D. Roles of phospholipase C beta2 in chemoattractant-elicited responses. Proc. Natl. Acad. Sci. USA 1997, 94, 7971–7975. [Google Scholar] [CrossRef]
- Li, Z.; Jiang, H.; Xie, W.; Zhang, Z.; Smrcka, A.V.; Wu, D. Roles of PLC-beta2 and -beta3 and PI3Kgamma in chemoattractant-mediated signal transduction. Science 2000, 287, 1046–1149. [Google Scholar] [CrossRef]
- Zhao, G.Q.; Zhang, Y.; Hoon, M.A.; Chandrashekar, J.; Erlenbach, I.; Ryba, N.J.; Zuker, C.S. The receptors for mammalian sweet and umami taste. Cell 2003, 115, 255–266. [Google Scholar] [PubMed]
- Damak, S.; Rong, M.; Yasumatsu, K.; Kokrashvili, Z.; Pérez, C.A.; Shigemura, N.; Yoshida, R.; Mosinger, B., Jr.; Glendinning, J.I.; Ninomiya, Y.; et al. Trpm5 null mice respond to bitter, sweet, and umami compounds. Chem. Senses 2006, 31, 253–264. [Google Scholar] [CrossRef]
- Hisatsune, C.; Yasumatsu, K.; Takahashi-Iwanaga, H.; Ogawa, N.; Kuroda, Y.; Yoshida, R.; Ninomiya, Y.; Mikoshiba, K. Abnormal taste perception in mice lacking the type 3 inositol 1,4,5-trisphosphate receptor. J. Biol. Chem. 2007, 282, 37225–37231. [Google Scholar]
- Wang, L.; Zhou, Y.; Chen, Z.; Sun, L.; Wu, J.; Li, H.; Liu, F.; Wang, F.; Yang, C.; Yang, J.; et al. PLCβ2 negatively regulates the inflammatory response to virus infection by inhibiting phosphoinositide-mediated activation of TAK1. Nat. Commun. 2019, 10, 746. [Google Scholar] [PubMed]
- Xiao, W.; Kashiwakura, J.; Hong, H.; Yasudo, H.; Ando, T.; Maeda-Yamamoto, M.; Wu, D.; Kawakami, Y.; Kawakami, T. Phospholipase C-β3 regulates FcɛRI-mediated mast cell activation by recruiting the protein phosphatase SHP-1. Immunity 2011, 34, 893–904. [Google Scholar] [CrossRef]
- Ando, T.; Xiao, W.; Gao, P.; Namiranian, S.; Matsumoto, K.; Tomimori, Y.; Hong, H.; Yamashita, H.; Kimura, M.; Kashiwakura, J.; et al. Critical role for mast cell Stat5 activity in skin inflammation. Cell. Rep. 2014, 6, 366–376. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Hong, H.; Kawakami, Y.; Kato, Y.; Wu, D.; Yasudo, H.; Kimura, A.; Kubagawa, H.; Bertoli, L.F.; Davis, R.S.; et al. Tumor suppression by phospholipase C-beta3 via SHP-1-mediated dephosphorylation of Stat5. Cancer Cell. 2009, 16, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, B.; Wang, P.; Dong, X.; Fernandez-Hernando, C.; Li, Z.; Hla, T.; Li, Z.; Claffey, K.; Smith, J.D.; et al. Phospholipase C beta3 deficiency leads to macrophage hypersensitivity to apoptotic induction and reduction of atherosclerosis in mice. J. Clin. Investig. 2008, 118, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Rimessi, A.; Bezzerri, V.; Salvatori, F.; Tamanini, A.; Nigro, F.; Dechecchi, M.C.; Santangelo, A.; Prandini, P.; Munari, S.; Provezza, L.; et al. PLCB3 Loss of Function Reduces Pseudomonas aeruginosa-Dependent IL-8 Release in Cystic Fibrosis. Am. J. Respir. Cell. Mol. Biol. 2018, 59, 428–436. [Google Scholar] [CrossRef]
- Bezzerri, V.; d’Adamo, P.; Rimessi, A.; Lanzara, C.; Crovella, S.; Nicolis, E.; Tamanini, A.; Athanasakis, E.; Tebon, M.; Bisoffi, G.; et al. Phospholipase C-β3 is a key modulator of IL-8 expression in cystic fibrosis bronchial epithelial cells. J. Immunol. 2011, 186, 4946–4958. [Google Scholar] [CrossRef] [PubMed]
- Ben-Salem, S.; Robbins, S.M.; Lm Sobreira, N.; Lyon, A.; Al-Shamsi, A.M.; Islam, B.K.; Akawi, N.A.; John, A.; Thachillath, P.; Al Hamed, S.; et al. Defect in phosphoinositide signalling through a homozygous variant in PLCB3 causes a new form of spondylometaphyseal dysplasia with corneal dystrophy. J. Med. Genet. 2018, 55, 122–130. [Google Scholar] [CrossRef]
- Kim, D.; Jun, K.S.; Lee, S.B.; Kang, N.G.; Min, D.S.; Kim, Y.H.; Ryu, S.H.; Suh, P.G.; Shin, H.S. Phospholipase C isozymes selectively couple to specific neurotransmitter receptors. Nature 1997, 389, 290–293. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Lyubarsky, A.; Dodd, R.; Vardi, N.; Pugh, E.; Baylor, D.; Simon, M.I.; Wu, D. Phospholipase C beta 4 is involved in modulating the visual response in mice. Proc. Natl. Acad. Sci. USA 1996, 93, 14598–14601. [Google Scholar] [CrossRef]
- Cheong, E.; Zheng, Y.; Lee, K.; Lee, J.; Kim, S.; Sanati, M.; Lee, S.; Kim, Y.S.; Shin, H.S. Deletion of phospholipase C beta4 in thalamocortical relay nucleus leads to absence seizures. Proc. Natl. Acad. Sci. USA 2009, 106, 21912–21917. [Google Scholar] [CrossRef]
- Moore, A.R.; Ceraudo, E.; Sher, J.J.; Guan, Y.; Shoushtari, A.N.; Chang, M.T.; Zhang, J.Q.; Walczak, E.G.; Kazmi, M.A.; Taylor, B.S.; et al. Recurrent activating mutations of G-protein-coupled receptor CYSLTR2 in uveal melanoma. Nat. Genet. 2016, 48, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Kido, Y.; Gordon, C.T.; Sakazume, S.; Ben Bdira, E.; Dattani, M.; Wilson, L.C.; Lyonnet, S.; Murakami, N.; Cunningham, M.L.; Amiel, J.; et al. Further characterization of atypical features in auriculocondylar syndrome caused by recessive PLCB4 mutations. Am. J. Med. Genet. A 2013, 161A, 2339–2346. [Google Scholar] [CrossRef]
- Ji, Q.S.; Winnier, G.E.; Niswender, K.D.; Horstman, D.; Wisdom, R.; Magnuson, M.A.; Carpenter, G. Essential role of the tyrosine kinase substrate phospholipase C-gamma1 in mammalian growth and development. Proc. Natl. Acad. Sci. USA 1997, 94, 2999–3003. [Google Scholar] [CrossRef]
- Liao, H.J.; Kume, T.; McKay, C.; Xu, M.J.; Ihle, J.N.; Carpenter, G. Absence of erythrogenesis and vasculogenesis in Plcg1-deficient mice. J. Biol. Chem. 2002, 277, 9335–9341. [Google Scholar] [CrossRef]
- Fu, G.; Chen, Y.; Yu, M.; Podd, A.; Schuman, J.; He, Y.; Di, L.; Yassai, M.; Haribhai, D.; North, P.E.; et al. Phospholipase C{gamma}1 is essential for T cell development, activation, and tolerance. J. Exp. Med. 2010, 207, 309–318. [Google Scholar] [CrossRef]
- Shirane, M.; Sawa, H.; Kobayashi, Y.; Nakano, T.; Kitajima, K.; Shinkai, Y.; Nagashima, K.; Negishi, I. Deficiency of phospholipase C-gamma1 impairs renal development and hematopoiesis. Development 2001, 128, 5173–5180. [Google Scholar] [CrossRef] [PubMed]
- Arteaga, C.L.; Johnson, M.D.; Todderud, G.; Coffey, R.J.; Carpenter, G.; Page, D.L. Elevated content of the tyrosine kinase substrate phospholipase C-gamma 1 in primary human breast carcinomas. Proc. Natl. Acad. Sci. USA 1991, 88, 10435–10439. [Google Scholar] [CrossRef] [PubMed]
- Park, J.G.; Lee, Y.H.; Kim, S.S.; Park, K.J.; Noh, D.Y.; Ryu, S.H.; Suh, P.G. Overexpression of phospholipase C-gamma 1 in familial adenomatous polyposis. Cancer Res. 1994, 54, 2240–2244. [Google Scholar]
- Noh, D.Y.; Lee, Y.H.; Kim, S.S.; Kim, Y.I.; Ryu, S.H.; Suh, P.G.; Park, J.G. Elevated content of phospholipase C-gamma 1 in colorectal cancer tissues. Cancer 1994, 73, 36–41. [Google Scholar] [CrossRef]
- Thomas, S.M.; Coppelli, F.M.; Wells, A.; Gooding, W.E.; Song, J.; Kassis, J.; Drenning, S.D.; Grandis, J.R. Epidermal growth factor receptor-stimulated activation of phospholipase Cgamma-1 promotes invasion of head and neck squamous cell carcinoma. Cancer Res. 2003, 63, 5629–5635. [Google Scholar] [PubMed]
- Sala, G.; Dituri, F.; Raimondi, C.; Previdi, S.; Maffucci, T.; Mazzoletti, M.; Rossi, C.; Iezzi, M.; Lattanzio, R.; Piantelli, M.; et al. Phospholipase Cgamma1 is required for metastasis development and progression. Cancer Res. 2008, 68, 10187–10196. [Google Scholar] [CrossRef] [PubMed]
- Lattanzio, R.; Marchisio, M.; La Sorda, R.; Tinari, N.; Falasca, M.; Alberti, S.; Miscia, S.; Ercolani, C.; Di Benedetto, A.; Perracchio, L.; et al. Overexpression of activated phospholipase Cγ1 is a risk factor for distant metastases in T1-T2, N0 breast cancer patients undergoing adjuvant chemotherapy. Int. J. Cancer. 2013, 132, 1022–1031. [Google Scholar] [CrossRef]
- Lattanzio, R.; Iezzi, M.; Sala, G.; Tinari, N.; Falasca, M.; Alberti, S.; Buglioni, S.; Mottolese, M.; Perracchio, L.; Natali, P.G.; et al. PLC-gamma- 1 phosphorylation status is prognostic of metastatic risk in patients with earlystage Luminal-A and -B breast cancer subtypes. BMC Cancer 2019, 19, 747. [Google Scholar] [CrossRef] [PubMed]
- Behjati, S.; Tarpey, P.S.; Sheldon, H.; Martincorena, I.; Van Loo, P.; Gundem, G.; Wedge, D.C.; Ramakrishna, M.; Cooke, S.L.; Pillay, N.; et al. Recurrent PTPRB and PLCG1 mutations in angiosarcoma. Nat. Genet. 2014, 46, 376–379. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.C.; Zhang, L.; Sung, Y.S.; Chen, C.L.; Kao, Y.C.; Agaram, N.P.; Singer, S.; Tap, W.D.; D’Angelo, S.; Antonescu, C.R. Recurrent CIC Gene Abnormalities in Angiosarcomas: A Molecular Study of 120 Cases With Concurrent Investigation of PLCG1, KDR, MYC, and FLT4 Gene Alterations. Am. J. Surg. Pathol. 2016, 40, 645–655. [Google Scholar] [CrossRef]
- Kunze, K.; Spieker, T.; Gamerdinger, U.; Nau, K.; Berger, J.; Dreyer, T.; Sindermann, J.R.; Hoffmeier, A.; Gattenlöhner, S.; Bräuninger, A. A recurrent activating PLCG1 mutation in cardiac angiosarcomas increases apoptosis resistance and invasiveness of endothelial cells. Cancer Res. 2014, 74, 6173–6183. [Google Scholar] [CrossRef] [PubMed]
- Murali, R.; Chandramohan, R.; Möller, I.; Scholz, S.L.; Berger, M.; Huberman, K.; Viale, A.; Pirun, M.; Socci, N.D.; Bouvier, N.; et al. Targeted massively parallel sequencing of angiosarcomas reveals frequent activation of the mitogen activated protein kinase pathway. Oncotarget 2015, 6, 36041–36052. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, K.; Nagata, Y.; Kitanaka, A.; Shiraishi, Y.; Shimamura, T.; Yasunaga, J.; Totoki, Y.; Chiba, K.; Sato-Otsubo, A.; Nagae, G.; et al. Integrated molecular analysis of adult T cell leukemia/lymphoma. Nat. Genet. 2015, 47, 1304–1315. [Google Scholar] [CrossRef]
- Vaqué, J.P.; Gómez-López, G.; Monsálvez, V.; Varela, I.; Martínez, N.; Pérez, C.; Domínguez, O.; Graña, O.; Rodríguez-Peralto, J.L.; Rodríguez-Pinilla, S.M.; et al. PLCG1 mutations in cutaneous T-cell lymphomas. Blood 2014, 123, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Ni, X.; Covington, K.R.; Yang, B.Y.; Shiu, J.; Zhang, X.; Xi, L.; Meng, Q.; Langridge, T.; Drummond, J.; et al. Genomic profiling of Sézary syndrome identifies alterations of key T cell signaling and differentiation genes. Nat. Genet. 2015, 47, 1426–1434. [Google Scholar] [CrossRef]
- Manso, R.; Rodríguez-Pinilla, S.M.; González-Rincón, J.; Gómez, S.; Monsalvo, S.; Llamas, P.; Rojo, F.; Pérez-Callejo, D.; Cereceda, L.; Limeres, M.A.; et al. Recurrent presence of the PLCG1 S345F mutation in nodal peripheral T-cell lymphomas. Haematologica 2015, 100, e25–e27. [Google Scholar] [CrossRef] [PubMed]
- Vallois, D.; Dobay, M.P.; Morin, R.D.; Lemonnier, F.; Missiaglia, E.; Juilland, M.; Iwaszkiewicz, J.; Fataccioli, V.; Bisig, B.; Roberti, A.; et al. Activating mutations in genes related to TCR signaling in angioimmunoblastic and other follicular helper T-cell-derived lymphomas. Blood 2016, 128, 1490–1502. [Google Scholar] [CrossRef] [PubMed]
- Saliakoura, M.; Rossi Sebastiano, M.; Pozzato, C.; Heidel, F.H.; Schnöder, T.M.; Savic Prince, S.; Bubendorf, L.; Pinton, P.A.; Schmid, R.; Baumgartner, J.; et al. PLCγ1 suppression promotes the adaptation of KRAS-mutant lung adenocarcinomas to hypoxia. Nat. Cell. Biol. 2020, 22, 1382–1395. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.S.; Yang, Y.R.; Lee, C.; Park, B.; Park, K.I.; Seo, J.K.; Seo, Y.K.; Cho, H.; Lucio, C.; Suh, P.G. Netrin-1/DCC-mediated PLCγ1 activation is required for axon guidance and brain structure development. EMBO Rep. 2018, 19, e46250. [Google Scholar] [CrossRef]
- Xie, Y.; Ding, Y.Q.; Hong, Y.; Feng, Z.; Navarre, S.; Xi, C.X.; Zhu, X.J.; Wang, C.L.; Ackerman, S.L.; Kozlowski, D.; et al. Phosphatidylinositol transfer protein-alpha in netrin-1-induced PLC signalling and neurite outgrowth. Nat. Cell. Biol. 2005, 7, 1124–1132. [Google Scholar] [CrossRef]
- Xie, Y.; Hong, Y.; Ma, X.Y.; Ren, X.R.; Ackerman, S.; Mei, L.; Xiong, W.C. DCC-dependent phospholipase C signaling in netrin-1-induced neurite elongation. J. Biol. Chem. 2006, 281, 2605–2611. [Google Scholar] [CrossRef]
- Yang, Y.R.; Jung, J.H.; Kim, S.J.; Hamada, K.; Suzuki, A.; Kim, H.J.; Lee, J.H.; Kwon, O.B.; Lee, Y.K.; Kim, J.; et al. Forebrain-specific ablation of phospholipase Cγ1 causes manic-like behavior. Mol. Psychiatry 2017, 22, 1473–1482. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.Y.; Yang, Y.R.; Hwang, H.; Lee, H.E.; Jang, H.J.; Kim, J.; Yang, E.; Kim, H.; Rhim, H.; Suh, P.G.; et al. Deletion of PLCγ1 in GABAergic neurons increases seizure susceptibility in aged mice. Sci. Rep. 2019, 9, 17761. [Google Scholar] [CrossRef] [PubMed]
- Gu, B.; Huang, Y.Z.; He, X.P.; Joshi, R.B.; Jang, W.; McNamara, J.O. A Peptide Uncoupling BDNF Receptor TrkB from Phospholipase Cγ1 Prevents Epilepsy Induced by Status Epilepticus. Neuron 2015, 88, 484–491. [Google Scholar] [CrossRef]
- Hashimoto, A.; Takeda, K.; Inaba, M.; Sekimata, M.; Kaisho, T.; Ikehara, S.; Homma, Y.; Akira, S.; Kurosaki, T. Cutting edge: Essential role of phospholipase C-gamma 2 in B cell development and function. J. Immunol. 2000, 165, 1738–1742. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Constien, R.; Dear, N.; Katan, M.; Hanke, P.; Bunney, T.D.; Kunder, S.; Quintanilla-Martinez, L.; Huffstadt, U.; Schröder, A.; et al. Autoimmunity and inflammation due to a gain-of-function mutation in phospholipase C gamma 2 that specifically increases external Ca2+ entry. Immunity 2005, 22, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Feng, J.; Wen, R.; Marine, J.C.; Sangster, M.Y.; Parganas, E.; Hoffmeyer, A.; Jackson, C.W.; Cleveland, J.L.; Murray, P.J.; et al. Phospholipase Cgamma2 is essential in the functions of B cell and several Fc receptors. Immunity. 2000, 13, 25–35. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, X.; Di, L.; Fu, G.; Chen, Y.; Bai, L.; Liu, J.; Feng, X.; McDonald, J.M.; Michalek, S.; et al. Phospholipase Cgamma2 mediates RANKL-stimulated lymph node organogenesis and osteoclastogenesis. J. Biol. Chem. 2008, 283, 29593–29601. [Google Scholar] [CrossRef]
- Bunney, T.D.; Baxendale, R.W.; Martins, M.S.; Romberg, N.; Komarow, H.; Aksentijevich, I.; Kim, H.S.; Ho, J.; Cruse, G.; Jung, M.Y.; et al. Cold urticaria, immunodeficiency, and autoimmunity related to PLCG2 deletions. N. Engl. J. Med. 2012, 366, 330–338. [Google Scholar]
- Novice, T.; Kariminia, A.; Del Bel, K.L.; Lu, H.; Sharma, M.; Lim, C.J.; Read, J.; Lugt, M.V.; Hannibal, M.C.; O’Dwyer, D.; et al. A Germline Mutation in the C2 Domain of PLCγ2 Associated with Gain-of-Function Expands the Phenotype for PLCG2-Related Diseases. J. Clin. Immunol. 2020, 40, 267–276. [Google Scholar] [CrossRef]
- Martín-Nalda, A.; Fortuny, C.; Rey, L.; Bunney, T.D.; Alsina, L.; Esteve-Solé, A.; Bull, D.; Anton, M.C.; Basagaña, M.; Casals, F.; et al. Severe Autoinflammatory Manifestations and Antibody Deficiency Due to Novel Hypermorphic PLCG2 Mutations. J. Clin. Immunol. 2020, 40, 987–1000. [Google Scholar] [CrossRef]
- Zhou, Q.; Lee, G.S.; Brady, J.; Datta, S.; Katan, M.; Sheikh, A.; Martins, M.S.; Bunney, T.D.; Santich, B.H.; Moir, S.; et al. A hypermorphic missense mutation in PLCG2, encoding phospholipase Cγ2, causes a dominantly inherited autoinflammatory disease with immunodeficiency. Am. J. Hum. Genet. 2012, 91, 713–720. [Google Scholar] [CrossRef]
- Nakamura, Y.; Fukami, K.; Yu, H.; Takenaka, K.; Kataoka, Y.; Shirakata, Y.; Nishikawa, S.I.; Hashimoto, K.; Yoshida, N.; Takenawa, T. Phospholipase Cdelta1 is required for skin stem cell lineage commitment. EMBO J. 2003, 22, 2981–2991. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Ichinohe, M.; Hirata, M.; Matsuura, H.; Fujiwara, T.; Igarashi, T.; Nakahara, M.; Yamaguchi, H.; Yasugi, S.; Takenawa, T.; et al. Phospholipase C-delta1 is an essential molecule downstream of Foxn1, the gene responsible for the nude mutation, in normal hair development. FASEB J. 2008, 22, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Kiuru, M.; Kurban, M.; Itoh, M.; Petukhova, L.; Shimomura, Y.; Wajid, M.; Christiano, A.M. Hereditary leukonychia, or porcelain nails, resulting from mutations in PLCD1. Am. J. Hum. Genet. 2011, 88, 839–844. [Google Scholar] [CrossRef]
- Khan, A.K.; Khan, S.A.; Muhammad, N.; Muhammad, N.; Ahmad, J.; Nawaz, H.; Nasir, A.; Farman, S.; Khan, S. Mutation in Phospholipase C, δ1 ( PLCD1) Gene Underlies Hereditary Leukonychia in a Pashtun Family and Review of the Literature. Balkan J. Med. Genet. 2018, 21, 69–72. [Google Scholar] [CrossRef]
- Khan, T.; Khan, M.; Yousaf, A.; Khan, S.; Naeem, M.; Shah, A.; Murtaza, G.; Ali, A.; Jabeen, N.; Hussain, H.M.J.; et al. Whole exome sequencing identifies a novel dominant missense mutation underlying leukonychia in a Pakistani family. J. Hum. Genet. 2018, 63, 1071–1076. [Google Scholar] [CrossRef] [PubMed]
- Xue, K.; Zheng, Y.; Shen, C.; Cui, Y. Identification of a novel PLCD1 mutation in Chinese Han pedigree with hereditary leukonychia and koilonychia. J. Cosmet. Dermatol. 2019, 18, 912–915. [Google Scholar] [CrossRef]
- Kanemaru, K.; Nakamura, Y.; Sato, K.; Kojima, R.; Takahashi, S.; Yamaguchi, M.; Ichinohe, M.; Kiyonari, H.; Shioi, G.; Kabashima, K.; et al. Epidermal phospholipase Cδ1 regulates granulocyte counts and systemic interleukin-17 levels in mice. Nat. Commun. 2012, 3, 963. [Google Scholar] [CrossRef]
- Kanemaru, K.; Nakamura, Y.; Totoki, K.; Fukuyama, T.; Shoji, M.; Kaneko, H.; Shiratori, K.; Yoneda, A.; Inoue, T.; Iwakura, Y.; et al. Phospholipase Cδ1 regulates p38 MAPK activity and skin barrier integrity. Cell. Death Differ. 2017, 24, 1079–1090. [Google Scholar] [CrossRef]
- Park, S.J.; Haan, K.D.; Nakamura, Y.; Fukami, K.; Fisher, T.E. PLCδ1 Plays Central Roles in the Osmotic Activation of ΔN-TRPV1 Channels in Mouse Supraoptic Neurons and in Murine Osmoregulation. J. Neurosci. 2021, 41, 3579–3587. [Google Scholar] [CrossRef]
- Haan, K.D.; Park, S.J.; Nakamura, Y.; Fukami, K.; Fisher, T.E. Osmotically evoked PLCδ1-dependent translocation of ΔN-TRPV1 channels in rat supraoptic neurons. iScience 2023, 26, 106258. [Google Scholar] [CrossRef]
- Hu, X.T.; Zhang, F.B.; Fan, Y.C.; Shu, X.S.; Wong, A.H.; Zhou, W.; Shi, Q.L.; Tang, H.M.; Fu, L.; Guan, X.Y.; et al. Phospholipase C delta 1 is a novel 3p22.3 tumor suppressor involved in cytoskeleton organization, with its epigenetic silencing correlated with high-stage gastric cancer. Oncogene 2009, 28, 2466–2475. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.; Wang, N.; Zhao, L.; Li, S.; Li, Q.; Chen, L.; Luo, X.; Qiu, Z.; Li, L.; Ren, G.; et al. Methylation of PLCD1 and adenovirus-mediated PLCD1 overexpression elicits a gene therapy effect on human breast cancer. Exp. Cell. Res. 2015, 332, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Xiang, T.; Li, L.; Fan, Y.; Jiang, Y.; Ying, Y.; Putti, T.C.; Tao, Q.; Ren, G. PLCD1 is a functional tumor suppressor inducing G(2)/M arrest and frequently methylated in breast cancer. Cancer Biol. Ther. 2010, 10, 520–527. [Google Scholar] [CrossRef][Green Version]
- Fu, L.; Qin, Y.R.; Xie, D.; Hu, L.; Kwong, D.L.; Srivastava, G.; Tsao, S.W.; Guan, X.Y. Characterization of a novel tumor-suppressor gene PLC delta 1 at 3p22 in esophageal squamous cell carcinoma. Cancer Res. 2007, 67, 10720–10726. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Hamada, Y.; Fujiwara, T.; Enomoto, H.; Hiroe, T.; Tanaka, S.; Nose, M.; Nakahara, M.; Yoshida, N.; Takenawa, T.; et al. Phospholipase C-delta1 and -delta3 are essential in the trophoblast for placental development. Mol. Cell. Biol. 2005, 25, 10979–10988. [Google Scholar] [CrossRef]
- Nakamura, Y.; Kanemaru, K.; Kojima, R.; Hashimoto, Y.; Marunouchi, T.; Oka, N.; Ogura, T.; Tanonaka, K.; Fukami, K. Simultaneous loss of phospholipase Cδ1 and phospholipase Cδ3 causes cardiomyocyte apoptosis and cardiomyopathy. Cell Death Dis. 2014, 5, e1215. [Google Scholar] [CrossRef] [PubMed]
- Fukami, K.; Nakao, K.; Inoue, T.; Kataoka, Y.; Kurokawa, M.; Fissore, R.A.; Nakamura, K.; Katsuki, M.; Mikoshiba, K.; Yoshida, N.; et al. Requirement of phospholipase Cdelta4 for the zona pellucida-induced acrosome reaction. Science 2001, 292, 920–923. [Google Scholar] [CrossRef]
- Fukami, K.; Yoshida, M.; Inoue, T.; Kurokawa, M.; Fissore, R.A.; Yoshida, N.; Mikoshiba, K.; Takenawa, T. Phospholipase Cdelta4 is required for Ca2+ mobilization essential for acrosome reaction in sperm. J. Cell. Biol. 2003, 161, 79–88. [Google Scholar] [CrossRef]
- Wang, H.; Oestreich, E.A.; Maekawa, N.; Bullard, T.A.; Vikstrom, K.L.; Dirksen, R.T.; Kelley, G.G.; Blaxall, B.C.; Smrcka, A.V. Phospholipase C epsilon modulates beta-adrenergic receptor-dependent cardiac contraction and inhibits cardiac hypertrophy. Circ. Res. 2005, 97, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Nash, C.A.; Wei, W.; Irannejad, R.; Smrcka, A.V. Golgi localized β1-adrenergic receptors stimulate Golgi PI4P hydrolysis by PLCε to regulate cardiac hypertrophy. eLife 2019, 8, e48167. [Google Scholar] [CrossRef]
- Tadano, M.; Edamatsu, H.; Minamisawa, S.; Yokoyama, U.; Ishikawa, Y.; Suzuki, N.; Saito, H.; Wu, D.; Masago-Toda, M.; Yamawaki-Kataoka, Y.; et al. Congenital semilunar valvulogenesis defect in mice deficient in phospholipase C epsilon. Mol. Cell. Biol. 2005, 25, 2191–2199. [Google Scholar] [CrossRef] [PubMed]
- Takenaka, N.; Edamatsu, H.; Suzuki, N.; Saito, H.; Inoue, Y.; Oka, M.; Hu, L.; Kataoka, T. Overexpression of phospholipase Cε in keratinocytes upregulates cytokine expression and causes dermatitis with acanthosis and T-cell infiltration. Eur. J. Immunol. 2011, 41, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Edamatsu, H.; Takenaka, N.; Ikuta, S.; Kataoka, T. Crucial role of phospholipase Cepsilon in induction of local skin inflammatory reactions in the elicitation stage of allergic contact hypersensitivity. J. Immunol. 2010, 184, 993–1002. [Google Scholar] [CrossRef]
- Dusaban, S.S.; Purcell, N.H.; Rockenstein, E.; Masliah, E.; Cho, M.K.; Smrcka, A.V.; Brown, J.H. Phospholipase C epsilon links G protein-coupled receptor activation to inflammatory astrocytic responses. Proc. Natl. Acad. Sci. USA 2013, 110, 3609–3614. [Google Scholar] [CrossRef]
- Hinkes, B.; Wiggins, R.C.; Gbadegesin, R.; Vlangos, C.N.; Seelow, D.; Nürnberg, G.; Garg, P.; Verma, R.; Chaib, H.; Hoskins, B.E.; et al. Positional cloning uncovers mutations in PLCE1 responsible for a nephrotic syndrome variant that may be reversible. Nat. Genet. 2006, 38, 1397–1405. [Google Scholar] [CrossRef]
- Rao, J.; Ashraf, S.; Tan, W.; van der Ven, A.T.; Gee, H.Y.; Braun, D.A.; Fehér, K.; George, S.P.; Esmaeilniakooshkghazi, A.; Choi, W.I.; et al. Advillin acts upstream of phospholipase C ϵ1 in steroid-resistant nephrotic syndrome. J. Clin. Investig. 2017, 127, 4257–4269. [Google Scholar] [CrossRef]
- Saunders, C.M.; Larman, M.G.; Parrington, J.; Cox, L.J.; Royse, J.; Blayney, L.M.; Swann, K.; Lai, F.A. PLC zeta: A sperm-specific trigger of Ca(2+) oscillations in eggs and embryo development. Development 2002, 129, 3533–3544. [Google Scholar] [CrossRef]
- Knott, J.G.; Kurokawa, M.; Fissore, R.A.; Schultz, R.M.; Williams, C.J. Transgenic RNA interference reveals role for mouse sperm phospholipase Czeta in triggering Ca2+ oscillations during fertilization. Biol. Reprod. 2005, 72, 992–996. [Google Scholar] [CrossRef]
- Nozawa, K.; Satouh, Y.; Fujimoto, T.; Oji, A.; Ikawa, M. Sperm-borne phospholipase C zeta-1 ensures monospermic fertilization in mice. Sci. Rep. 2018, 8, 1315. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Amanai, M.; Fukui, T.; Kajikawa, E.; Brahmajosyula, M.; Iwahori, A.; Nakano, Y.; Shoji, S.; Diebold, J.; Hessel, H.; et al. Broad, ectopic expression of the sperm protein PLCZ1 induces parthenogenesis and ovarian tumours in mice. Development 2007, 134, 3941–3952. [Google Scholar] [CrossRef][Green Version]
- Escoffier, J.; Lee, H.C.; Yassine, S.; Zouari, R.; Martinez, G.; Karaouzène, T.; Coutton, C.; Kherraf, Z.E.; Halouani, L.; Triki, C.; et al. Homozygous mutation of PLCZ1 leads to defective human oocyte activation and infertility that is not rescued by the WW-binding protein PAWP. Hum. Mol. Genet. 2016, 25, 878–891. [Google Scholar] [CrossRef] [PubMed]
- Torra-Massana, M.; Cornet-Bartolomé, D.; Barragán, M.; Durban, M.; Ferrer-Vaquer, A.; Zambelli, F.; Rodriguez, A.; Oliva, R.; Vassena, R. Novel phospholipase C zeta 1 mutations associated with fertilization failures after ICSI. Hum. Reprod. 2019, 34, 1494–1504. [Google Scholar] [CrossRef]
- Heytens, E.; Parrington, J.; Coward, K.; Young, C.; Lambrecht, S.; Yoon, S.Y.; Fissore, R.A.; Hamer, R.; Deane, C.M.; Ruas, M.; et al. Reduced amounts and abnormal forms of phospholipase C zeta (PLCzeta) in spermatozoa from infertile men. Hum. Reprod. 2009, 24, 2417–2428. [Google Scholar] [CrossRef]
- Feisst, C.; Albert, D.; Steinhilber, D.; Werz, O. The aminosteroid phospholipase C antagonist U-73122 (1-[6-[[17-beta-3-methoxyestra-1,3,5(10)-trien-17-yl]amino]hexyl]-1H-pyrrole-2,5-dione) potently inhibits human 5-lipoxygenase in vivo and in vitro. Mol. Pharmacol. 2005, 67, 1751–1757. [Google Scholar] [CrossRef] [PubMed]
- Hollywood, M.A.; Sergeant, G.P.; Thornbury, K.D.; McHale, N.G. The PI-PLC inhibitor U-73122 is a potent inhibitor of the SERCA pump in smooth muscle. Br. J. Pharmacol. 2010, 160, 1293–1294. [Google Scholar] [CrossRef]
- Leitner, M.G.; Michel, N.; Behrendt, M.; Dierich, M.; Dembla, S.; Wilke, B.U.; Konrad, M.; Lindner, M.; Oberwinkler, J.; Oliver, D. Direct modulation of TRPM4 and TRPM3 channels by the phospholipase C inhibitor U73122. Br. J. Pharmacol. 2016, 173, 2555–2569. [Google Scholar] [CrossRef]
- Krjukova, J.; Holmqvist, T.; Danis, A.S.; Akerman, K.E.; Kukkonen, J.P. Phospholipase C activator m-3M3FBS affects Ca2+ homeostasis independently of phospholipase C activation. Br. J. Pharmacol. 2004, 143, 3–7. [Google Scholar] [CrossRef]
- Trinquet, E.; Fink, M.; Bazin, H.; Grillet, F.; Maurin, F.; Bourrier, E.; Ansanay, H.; Leroy, C.; Michaud, A.; Durroux, T.; et al. D-myo-inositol 1-phosphate as a surrogate of D-myo-inositol 1,4,5-tris phosphate to monitor G protein-coupled receptor activation. Anal. Biochem. 2006, 358, 126–135. [Google Scholar] [CrossRef]
- Huang, W.; Hicks, S.N.; Sondek, J.; Zhang, Q. A fluorogenic, small molecule reporter for mammalian phospholipase C isozymes. ACS Chem. Biol. 2011, 6, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wang, X.; Endo-Streeter, S.; Barrett, M.; Waybright, J.; Wohlfeld, C.; Hajicek, N.; Harden, T.K.; Sondek, J.; Zhang, Q. A membrane-associated, fluorogenic reporter for mammalian phospholipase C isozymes. J. Biol. Chem. 2018, 293, 1728–1735. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanemaru, K.; Nakamura, Y. Activation Mechanisms and Diverse Functions of Mammalian Phospholipase C. Biomolecules 2023, 13, 915. https://doi.org/10.3390/biom13060915
Kanemaru K, Nakamura Y. Activation Mechanisms and Diverse Functions of Mammalian Phospholipase C. Biomolecules. 2023; 13(6):915. https://doi.org/10.3390/biom13060915
Chicago/Turabian StyleKanemaru, Kaori, and Yoshikazu Nakamura. 2023. "Activation Mechanisms and Diverse Functions of Mammalian Phospholipase C" Biomolecules 13, no. 6: 915. https://doi.org/10.3390/biom13060915
APA StyleKanemaru, K., & Nakamura, Y. (2023). Activation Mechanisms and Diverse Functions of Mammalian Phospholipase C. Biomolecules, 13(6), 915. https://doi.org/10.3390/biom13060915