
Functional Expression of IP, 5-HT4, D1, A2A, and VIP Receptors in Human Odontoblast Cell Line

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Immunofluorescence
2.3. Solutions and Reagents
2.4. Measurement of Intracellular cAMP Level
2.5. Statistical Analysis
3. Results
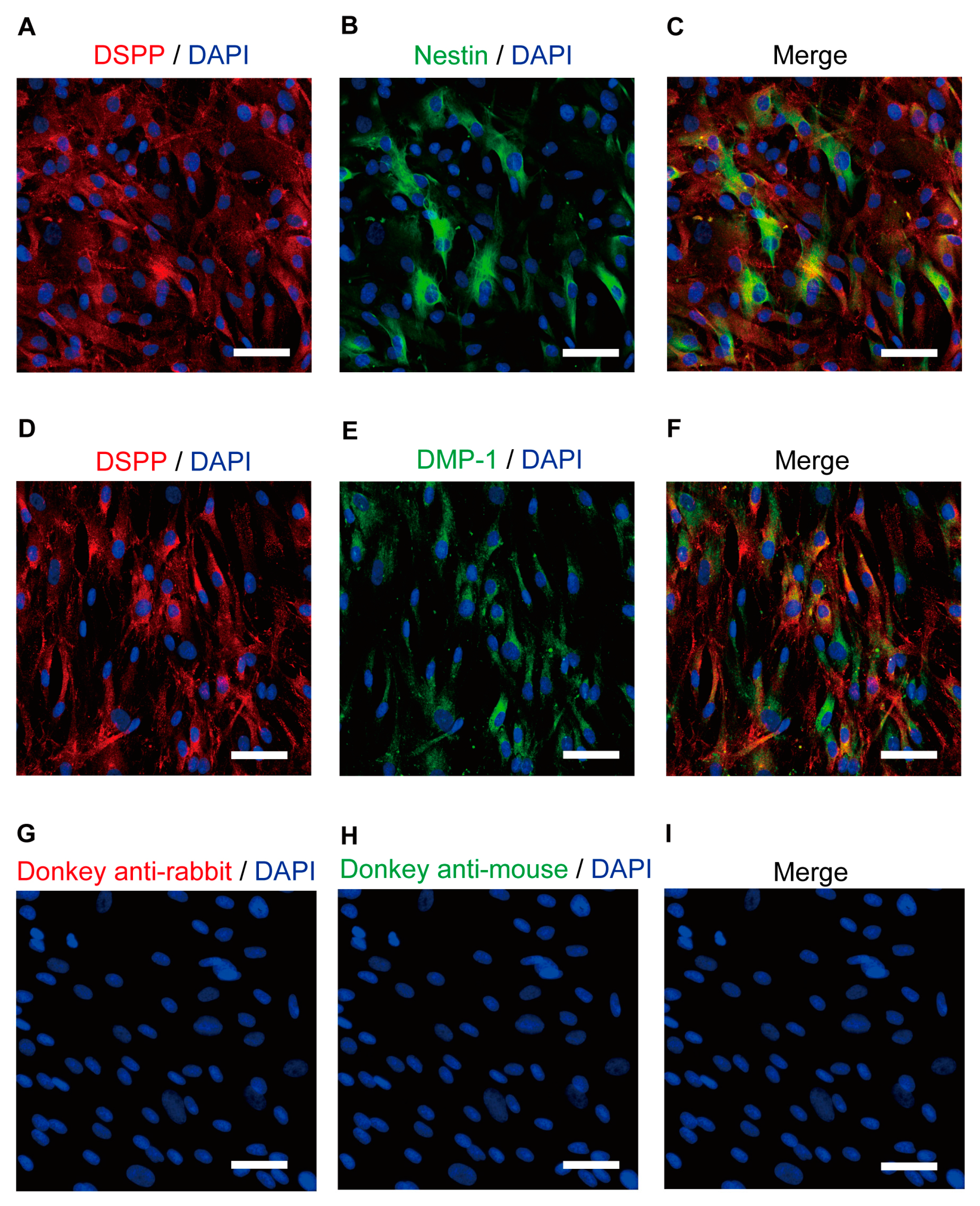
3.1. Immunofluorescence Analysis of Human Odontoblast Markers
3.2. Human Odontoblasts Were Immunopositive for Gαs Protein and IP, 5-HT4, D1, A2A, and VIP Receptor Antibodies
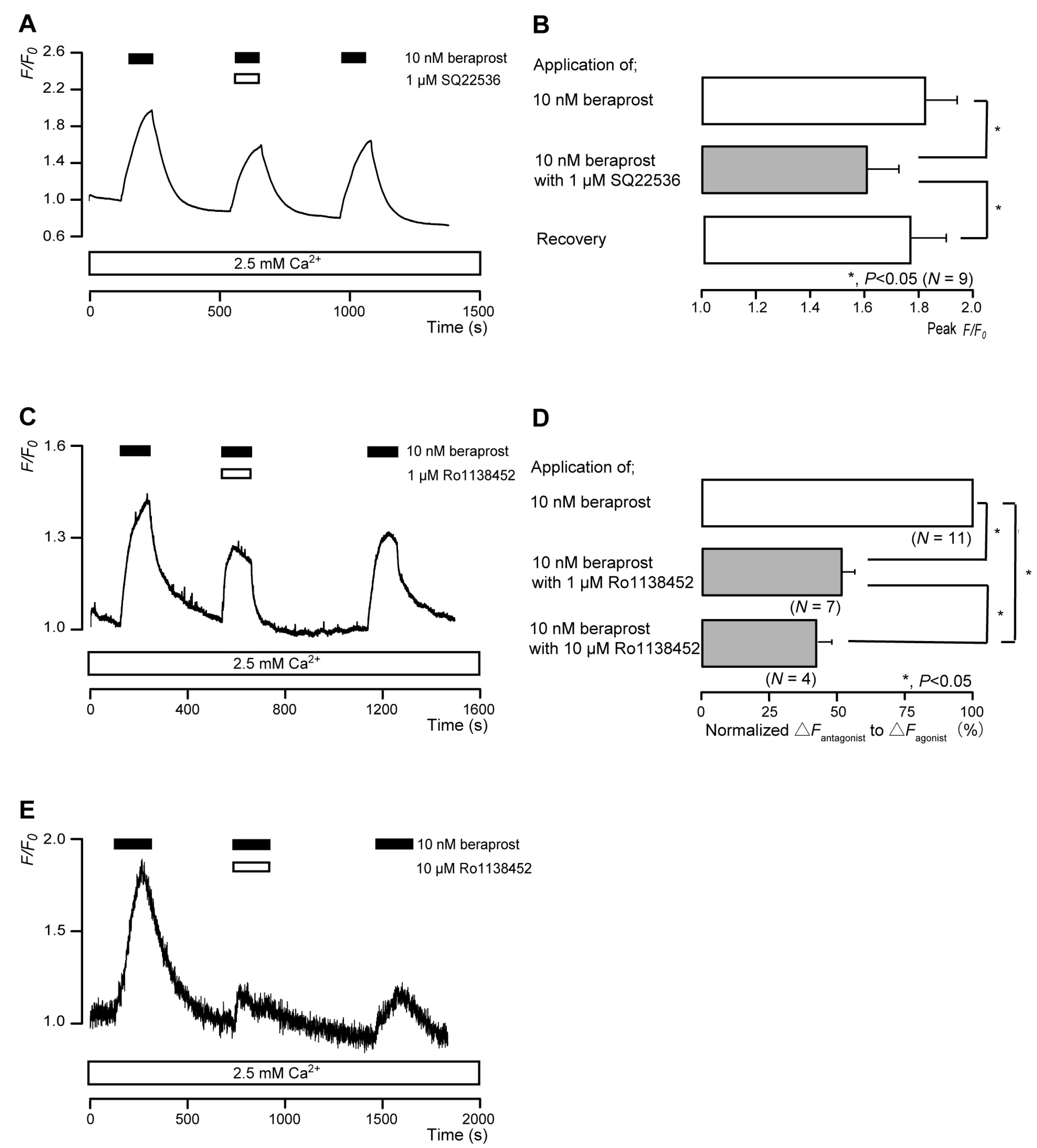
3.3. The IP Receptor Agonist Increased Intracellular cAMP Levels in Odontoblasts
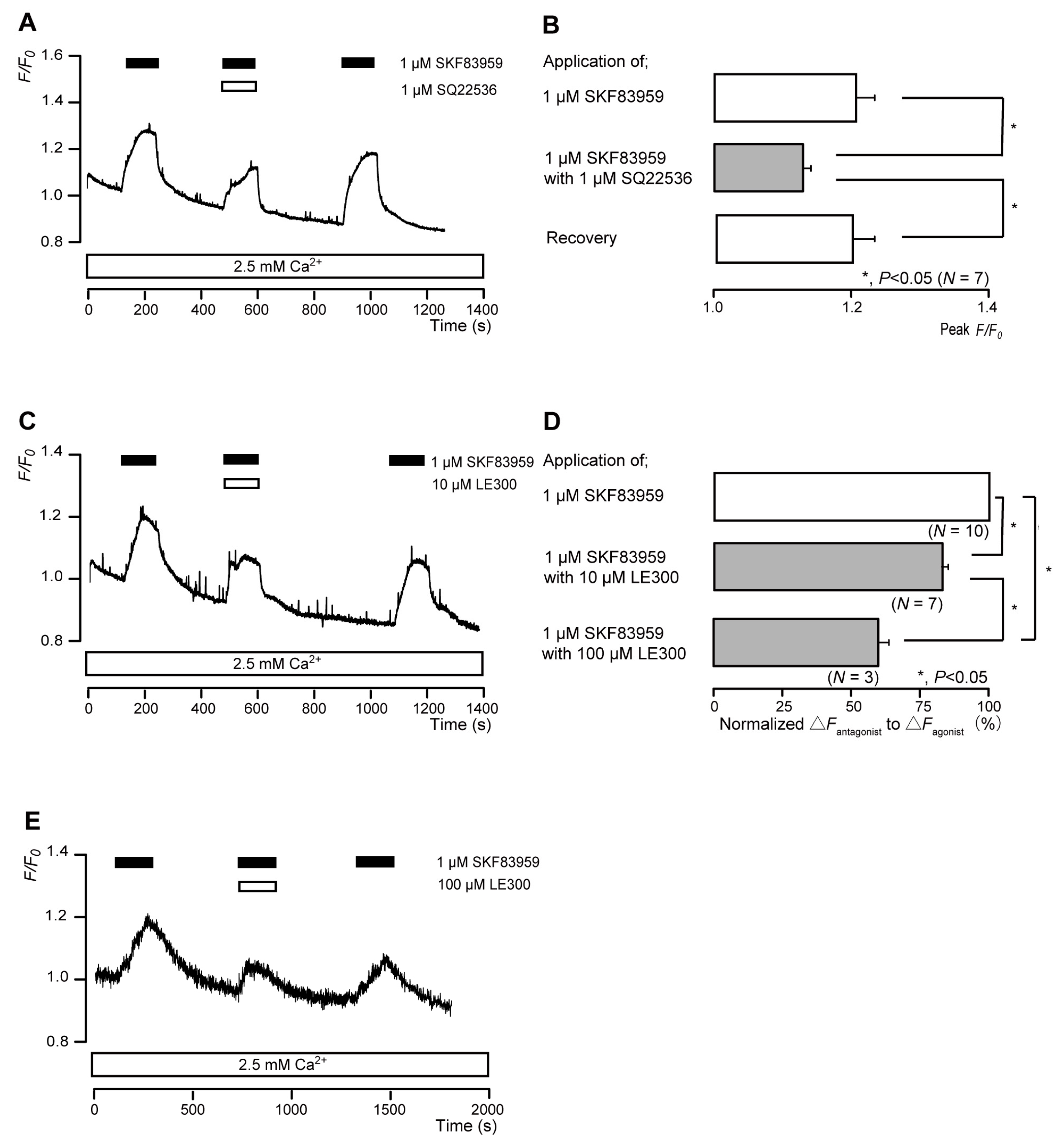
3.4. The 5-HT4 Receptor Agonist Increased Intracellular cAMP Levels in Odontoblasts
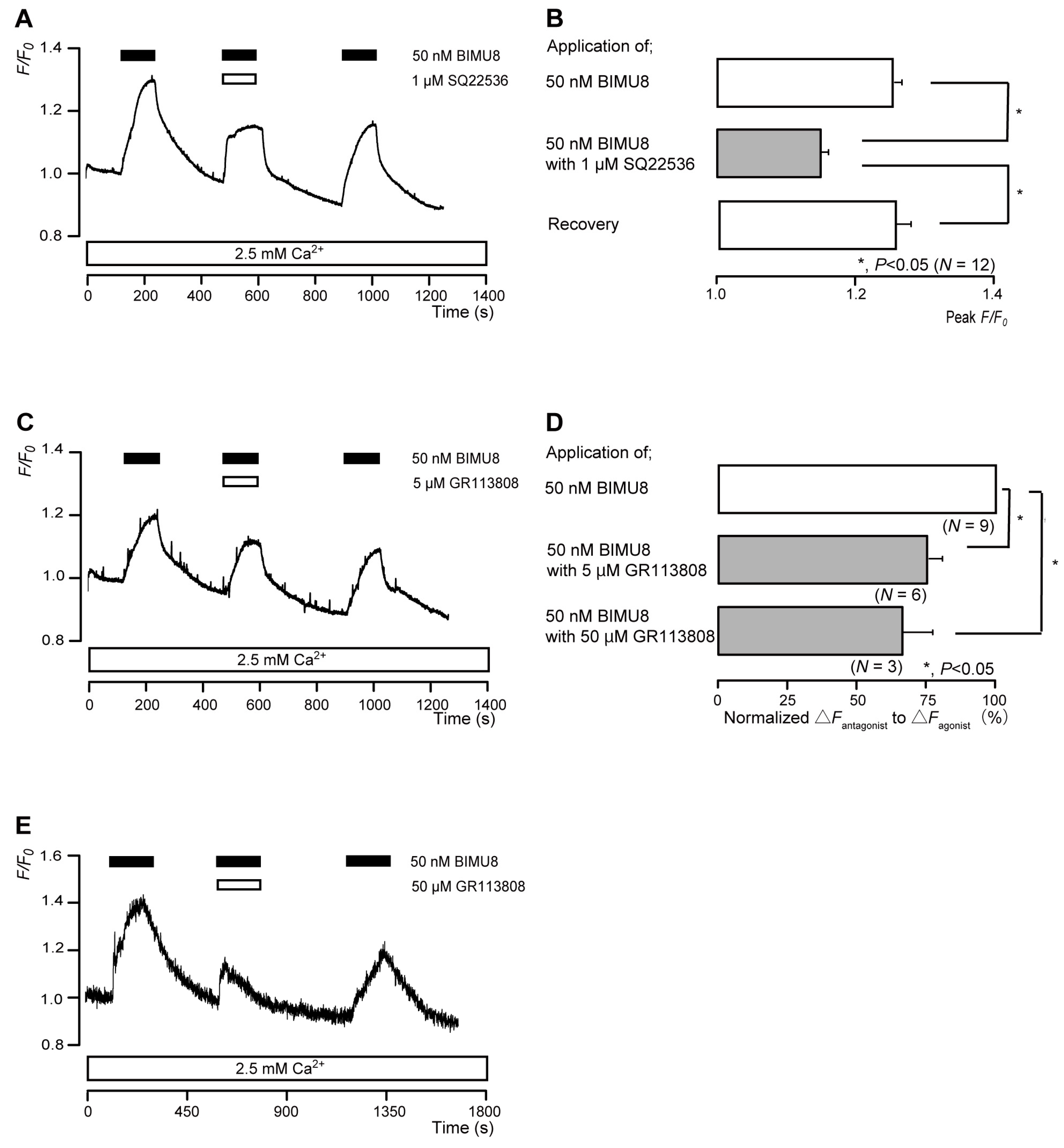
3.5. The D1 Receptor Agonist Increased Intracellular cAMP Levels in Odontoblasts
3.6. The A2A Receptor Agonist Increased Intracellular cAMP Levels in Odontoblasts
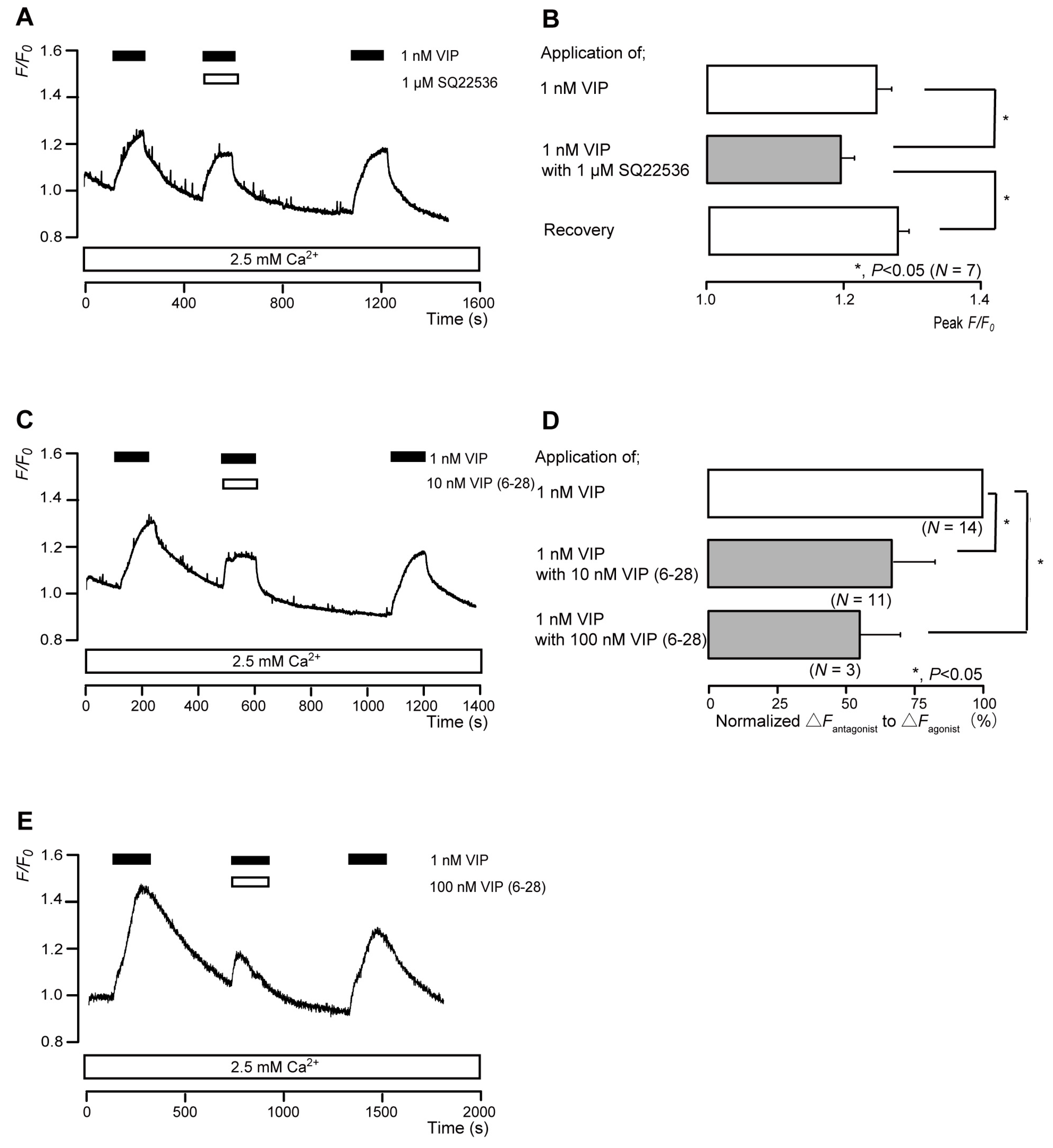
3.7. The Nonselective VIP Receptor Agonist Increased Intracellular cAMP Levels in Odontoblasts
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Duc, N.M.; Kim, H.R.; Chung, K.Y. Structural Mechanism of G Protein Activation by G Protein-Coupled Receptor. Eur. J. Pharmacol. 2015, 763, 214–222. [Google Scholar] [CrossRef]
- Syrovatkina, V.; Alegre, K.O.; Dey, R.; Huang, X.-Y. Regulation, Signaling, and Physiological Functions of G-Proteins. J. Mol. Biol. 2016, 428, 3850–3868. [Google Scholar] [CrossRef]
- Chaudhary, P.K.; Kim, S. An Insight into GPCR and G-Proteins as Cancer Drivers. Cells 2021, 10, 3288. [Google Scholar] [CrossRef]
- Aslam, M.; Ladilov, Y. Emerging Role of CAMP/AMPK Signaling. Cells 2022, 11, 308. [Google Scholar] [CrossRef]
- Sato, M.; Sobhan, U.; Tsumura, M.; Kuroda, H.; Soya, M.; Masamura, A.; Nishiyama, A.; Katakura, A.; Ichinohe, T.; Tazaki, M.; et al. Hypotonic-Induced Stretching of Plasma Membrane Activates Transient Receptor Potential Vanilloid Channels and Sodium-Calcium Exchangers in Mouse Odontoblasts. J. Endod. 2013, 39, 779–787. [Google Scholar] [CrossRef]
- Shibukawa, Y.; Sato, M.; Kimura, M.; Sobhan, U.; Shimada, M.; Nishiyama, A.; Kawaguchi, A.; Soya, M.; Kuroda, H.; Katakura, A.; et al. Odontoblasts as Sensory Receptors: Transient Receptor Potential Channels, Pannexin-1, and Ionotropic ATP Receptors Mediate Intercellular Odontoblast-Neuron Signal Transduction. Pflugers Arch. 2015, 467, 843–863. [Google Scholar] [CrossRef]
- Ohyama, S.; Ouchi, T.; Kimura, M.; Kurashima, R.; Yasumatsu, K.; Nishida, D.; Hitomi, S.; Ubaidus, S.; Kuroda, H.; Ito, S.; et al. Piezo1-Pannexin-1-P2X 3 Axis in Odontoblasts and Neurons Mediates Sensory Transduction in Dentinal Sensitivity. Front. Physiol. 2022, 13, 891759. [Google Scholar] [CrossRef]
- Tsumura, M.; Sobhan, U.; Muramatsu, T.; Sato, M.; Ichikawa, H.; Sahara, Y.; Tazaki, M.; Shibukawa, Y. TRPV1-Mediated Calcium Signal Couples with Cannabinoid Receptors and Sodium-Calcium Exchangers in Rat Odontoblasts. Cell Calcium 2012, 52, 124–136. [Google Scholar] [CrossRef]
- Nishiyama, A.; Sato, M.; Kimura, M.; Katakura, A.; Tazaki, M.; Shibukawa, Y. Intercellular Signal Communication among Odontoblasts and Trigeminal Ganglion Neurons via Glutamate. Cell Calcium 2016, 60, 341–355. [Google Scholar] [CrossRef]
- Sato, M.; Ogura, K.; Kimura, M.; Nishi, K.; Ando, M.; Tazaki, M.; Shibukawa, Y. Activation of Mechanosensitive Transient Receptor Potential/Piezo Channels in Odontoblasts Generates Action Potentials in Cocultured Isolectin B4-Negative Medium-Sized Trigeminal Ganglion Neurons. J. Endod. 2018, 44, 984–991.e2. [Google Scholar] [CrossRef]
- Glass, M.; Felder, C.C. Concurrent Stimulation of Cannabinoid CB1 and Dopamine D2 Receptors Augments CAMP Accumulation in Striatal Neurons: Evidence for a Gs Linkage to the CB1 Receptor. J. Neurosci. 1997, 17, 5327–5333. [Google Scholar] [CrossRef]
- Gu, J.; Ikeda, H.; Suda, H. Sympathetic Regulation of Tertiary Dentinogenesis via Beta-2 Adrenergic Receptor on Rat Odontoblasts. J. Endod. 2015, 41, 1056–1060. [Google Scholar] [CrossRef]
- Saito, N.; Kimura, M.; Ouchi, T.; Ichinohe, T.; Shibukawa, Y. Gαs-Coupled CGRP Receptor Signaling Axis from the Trigeminal Ganglion Neuron to Odontoblast Negatively Regulates Dentin Mineralization. Biomolecules 2022, 12, 1747. [Google Scholar] [CrossRef]
- Calvi, L.M.; Shin, H.I.; Knight, M.C.; Weber, J.M.; Young, M.F.; Giovannetti, A.; Schipani, E. Constitutively Active PTH/PTHrP Receptor in Odontoblasts Alters Odontoblast and Ameloblast Function and Maturation. Mech. Dev. 2004, 121, 397–408. [Google Scholar] [CrossRef]
- Ohkura, M.; Ohkura, N.; Yoshiba, N.; Yoshiba, K.; Ida-Yonemochi, H.; Ohshima, H.; Saito, I.; Okiji, T. Orthodontic Force Application Upregulated Pain-Associated Prostaglandin-I2/PGI2-Receptor/TRPV1 Pathway-Related Gene Expression in Rat Molars. Odontology 2018, 106, 2–10. [Google Scholar] [CrossRef]
- Fujino, S.; Hamano, S.; Tomokiyo, A.; Itoyama, T.; Hasegawa, D.; Sugii, H.; Yoshida, S.; Washio, A.; Nozu, A.; Ono, T.; et al. Expression and Function of Dopamine in Odontoblasts. J. Cell. Physiol. 2020, 235, 4376–4387. [Google Scholar] [CrossRef]
- Yi, X.; Wang, W.; Xie, Q. Adenosine Receptors Enhance the ATP-Induced Odontoblastic Differentiation of Human Dental Pulp Cells. Biochem. Biophys. Res. Commun. 2018, 497, 850–856. [Google Scholar] [CrossRef]
- Luthman, J.; Luthman, D.; Hökfelt, T. Occurrence and Distribution of Different Neurochemical Markers in the Human Dental Pulp. Arch. Oral Biol. 1992, 37, 193–208. [Google Scholar] [CrossRef]
- Dimitrova-Nakov, S.; Baudry, A.; Harichane, Y.; Collet, C.; Marchadier, A.; Kellermann, O.; Goldberg, M. Deletion of Serotonin 2B Receptor Provokes Structural Alterations of Mouse Dental Tissues. Calcif. Tissue Int. 2014, 94, 293–300. [Google Scholar] [CrossRef]
- Kitagawa, M.; Ueda, H.; Iizuka, S.; Sakamoto, K.; Oka, H.; Kudo, Y.; Ogawa, I.; Miyauchi, M.; Tahara, H.; Takata, T. Immortalization and Characterization of Human Dental Pulp Cells with Odontoblastic Differentiation. Arch. Oral Biol. 2007, 52, 727–731. [Google Scholar] [CrossRef]
- Ichikawa, H.; Kim, H.-J.; Shuprisha, A.; Shikano, T.; Tsumura, M.; Shibukawa, Y.; Tazaki, M. Voltage-Dependent Sodium Channels and Calcium-Activated Potassium Channels in Human Odontoblasts in Vitro. J. Endod. 2012, 38, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M.; Sase, T.; Higashikawa, A.; Sato, M.; Sato, T.; Tazaki, M.; Shibukawa, Y. High PH-Sensitive TRPA1 Activation in Odontoblasts Regulates Mineralization. J. Dent. Res. 2016, 95, 1057–1064. [Google Scholar] [CrossRef]
- Alcolado, N.G.; Conrad, D.J.; Poroca, D.; Li, M.; Alshafie, W.; Chappe, F.G.; Pelis, R.M.; Anini, Y.; Xu, Z.; Hamidi, S.; et al. Cystic Fibrosis Transmembrane Conductance Regulator Dysfunction in VIP Knockout Mice. Am. J. Physiol. Cell. Physiol. 2014, 307, C195–C207. [Google Scholar] [CrossRef] [PubMed]
- Rafferty, S.; Alcolado, N.; Norez, C.; Chappe, F.; Pelzer, S.; Becq, F.; Chappe, V. Rescue of Functional F508del Cystic Fibrosis Transmembrane Conductance Regulator by Vasoactive Intestinal Peptide in the Human Nasal Epithelial Cell Line JME/CF15. J. Pharmacol. Exp. Ther. 2009, 331, 2–13. [Google Scholar] [CrossRef]
- Yan, L.; Sun, S.; Qu, L. Insulin-like Growth Factor-1 Promotes the Proliferation and Odontoblastic Differentiation of Human Dental Pulp Cells under High Glucose Conditions. Int. J. Mol. Med. 2017, 40, 1253–1260. [Google Scholar] [CrossRef] [PubMed]
- Narumiya, S.; Sugimoto, Y.; Ushikubi, F. Prostanoid Receptors: Structures, Properties, and Functions. Physiol. Rev. 1999, 79, 1193–1226. [Google Scholar] [CrossRef]
- Shen, L.; Patel, J.A.; Norel, X.; Moledina, S.; Whittle, B.J.; von Kessler, K.; Sista, P.; Clapp, L.H. Pharmacology of the Single Isomer, Esuberaprost (Beraprost-314d) on Pulmonary Vascular Tone, IP Receptors and Human Smooth Muscle Proliferation in Pulmonary Hypertension. Biochem. Pharmacol. 2019, 166, 242–252. [Google Scholar] [CrossRef]
- Harris, D.N.; Asaad, M.M.; Phillips, M.B.; Goldenberg, H.J.; Antonaccio, M.J. Inhibition of Adenylate Cyclase in Human Blood Platelets by 9-Substituted Adenine Derivatives. J. Cycl. Nucleotide Res. 1979, 5, 125–134. [Google Scholar]
- Bley, K.R.; Bhattacharya, A.; Daniels, D.V.; Gever, J.; Jahangir, A.; O’Yang, C.; Smith, S.; Srinivasan, D.; Ford, A.P.D.W.; Jett, M.-F. RO1138452 and RO3244794: Characterization of Structurally Distinct, Potent and Selective IP (Prostacyclin) Receptor Antagonists. Br. J. Pharmacol. 2006, 147, 335–345. [Google Scholar] [CrossRef]
- Falcetti, E.; Flavell, D.M.; Staels, B.; Tinker, A.; Haworth, S.G.; Clapp, L.H. IP Receptor-Dependent Activation of PPARgamma by Stable Prostacyclin Analogues. Biochem. Biophys. Res. Commun. 2007, 360, 821–827. [Google Scholar] [CrossRef]
- Osawa, T.; Ohga, N.; Hida, Y.; Kitayama, K.; Akiyama, K.; Onodera, Y.; Fujie, M.; Shinohara, N.; Shindoh, M.; Nonomura, K.; et al. Prostacyclin Receptor in Tumor Endothelial Cells Promotes Angiogenesis in an Autocrine Manner. Cancer Sci. 2012, 103, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Okiji, T.; Morita, I.; Kawashima, N.; Kosaka, T.; Suda, H.; Murota, S. Immunohistochemical Detection of Prostaglandin I2 Synthase in Various Calcified Tissue-Forming Cells in Rat. Arch. Oral Biol. 1993, 38, 31–36. [Google Scholar] [CrossRef] [PubMed]
- Moriyama, T.; Higashi, T.; Togashi, K.; Iida, T.; Segi, E.; Sugimoto, Y.; Tominaga, T.; Narumiya, S.; Tominaga, M. Sensitization of TRPV1 by EP1 and IP Reveals Peripheral Nociceptive Mechanism of Prostaglandins. Mol. Pain 2005, 1, 3. [Google Scholar] [CrossRef]
- Limjeerajarus, C.N.; Chanarattanubol, T.; Trongkij, P.; Rujiwanichkul, M.; Pavasant, P. Iloprost Induces Tertiary Dentin Formation. J. Endod. 2014, 40, 1784–1790. [Google Scholar] [CrossRef] [PubMed]
- Neumeyer, J.L.; Kula, N.S.; Bergman, J.; Baldessarini, R.J. Receptor Affinities of Dopamine D1 Receptor-Selective Novel Phenylbenzazepines. Eur. J. Pharmacol. 2003, 474, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Kassack, M.U.; Höfgen, B.; Decker, M.; Eckstein, N.; Lehmann, J. Pharmacological Characterization of the Benz[d]Indolo[2,3-g]Azecine LE300, a Novel Type of a Nanomolar Dopamine Receptor Antagonist. Naunyn Schmiedebergs Arch. Pharmacol. 2002, 366, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Robaa, D.; Kretschmer, R.; Siol, O.; AbulAzm, S.E.; ElKhawass, E.; Lehmann, J.; Enzensperger, C. Residues at the Indole-NH of LE300 Modulate Affinities and Selectivities for Dopamine Receptors. Arch. Pharm. 2011, 344, 28–36. [Google Scholar] [CrossRef]
- Baudry, A.; Schneider, B.; Launay, J.-M.; Kellermann, O. Serotonin in Stem Cell Based-Dental Repair and Bone Formation: A Review. Biochimie 2019, 161, 65–72. [Google Scholar] [CrossRef]
- Sato, M.; Furuya, T.; Kimura, M.; Kojima, Y.; Tazaki, M.; Sato, T.; Shibukawa, Y. Intercellular Odontoblast Communication via ATP Mediated by Pannexin-1 Channel and Phospholipase C-Coupled Receptor Activation. Front. Physiol. 2015, 6, 326. [Google Scholar] [CrossRef]
- Dunwiddie, T.V.; Masino, S.A. The Role and Regulation of Adenosine in the Central Nervous System. Annu. Rev. Neurosci. 2001, 24, 31–55. [Google Scholar] [CrossRef]
- El-Tayeb, A.; Michael, S.; Abdelrahman, A.; Behrenswerth, A.; Gollos, S.; Nieber, K.; Müller, C.E. Development of Polar Adenosine A2A Receptor Agonists for Inflammatory Bowel Disease: Synergism with A2B Antagonists. ACS Med. Chem. Lett. 2011, 2, 890–895. [Google Scholar] [CrossRef]
- Navarro, G.; Gonzalez, A.; Campanacci, S.; Rivas-Santisteban, R.; Reyes-Resina, I.; Casajuana-Martin, N.; Cordomí, A.; Pardo, L.; Franco, R. Experimental and Computational Analysis of Biased Agonism on Full-Length and a C-Terminally Truncated Adenosine A2A Receptor. Comput. Struct. Biotechnol. J. 2020, 18, 2723–2732. [Google Scholar] [CrossRef] [PubMed]
- Rau, A.R.; Ariwodola, O.J.; Weiner, J.L. Postsynaptic Adenosine A2A Receptors Modulate Intrinsic Excitability of Pyramidal Cells in the Rat Basolateral Amygdala. Int. J. Neuropsychopharmacol. 2015, 18, pyv017. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, A.M.; Traini, C.; Cipriani, S.; Gianfriddo, M.; Mello, T.; Giovannini, M.G.; Galli, A.; Pedata, F. The Adenosine A2A Receptor Antagonist ZM241385 Enhances Neuronal Survival after Oxygen-Glucose Deprivation in Rat CA1 Hippocampal Slices. Br. J. Pharmacol. 2009, 157, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Yu, L.; Wang, Q.; Pelletier, J.; Fausther, M.; Sévigny, J.; Malmström, H.S.; Dirksen, R.T.; Ren, Y.F. Expression of Ecto-ATPase NTPDase2 in Human Dental Pulp. J. Dent. Res. 2012, 91, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Couvineau, A.; Laburthe, M. VPAC Receptors: Structure, Molecular Pharmacology and Interaction with Accessory Proteins. Br. J. Pharmacol. 2012, 166, 42–50. [Google Scholar] [CrossRef]
- Couvineau, A.; Rouyer-Fessard, C.; Maoret, J.J.; Gaudin, P.; Nicole, P.; Laburthe, M. Vasoactive Intestinal Peptide (VIP)1 Receptor. Three Nonadjacent Amino Acids Are Responsible for Species Selectivity with Respect to Recognition of Peptide Histidine Isoleucineamide. J. Biol. Chem. 1996, 271, 12795–12800. [Google Scholar] [CrossRef]
- Piet, R.; Dunckley, H.; Lee, K.; Herbison, A.E. Vasoactive Intestinal Peptide Excites GnRH Neurons in Male and Female Mice. Endocrinology 2016, 157, 3621–3630. [Google Scholar] [CrossRef]
- Schuelert, N.; McDougall, J.J. Electrophysiological Evidence That the Vasoactive Intestinal Peptide Receptor Antagonist VIP6-28 Reduces Nociception in an Animal Model of Osteoarthritis. Osteoarthr. Cartil. 2006, 14, 1155–1162. [Google Scholar] [CrossRef]
- Hogan, K.; Markos, F. Vasoactive Intestinal Polypeptide Receptor Antagonism Enhances the Vagally Induced Increase in Cardiac Interval of the Rat Atrium in Vitro. Exp. Physiol. 2006, 91, 641–646. [Google Scholar] [CrossRef]
- Casasco, A.; Calligaro, A.; Casasco, M.; Springall, D.R.; Polak, J.M.; Poggi, P.; Marchetti, C. Peptidergic Nerves in Human Dental Pulp. An Immunocytochemical Study. Histochemistry 1990, 95, 115–121. [Google Scholar] [CrossRef] [PubMed]
- El Karim, I.A.; Lamey, P.-J.; Ardill, J.; Linden, G.J.; Lundy, F.T. Vasoactive Intestinal Polypeptide (VIP) and VPAC1 Receptor in Adult Human Dental Pulp in Relation to Caries. Arch. Oral Biol. 2006, 51, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Jonnakuty, C.; Gragnoli, C. What Do We Know about Serotonin? J. Cell. Physiol. 2008, 217, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.E.; Nichols, C.D. Serotonin Receptors. Chem. Rev. 2008, 108, 1614–1641. [Google Scholar] [CrossRef] [PubMed]
- Pineda-Farias, J.B.; Barragán-Iglesias, P.; Valdivieso-Sánchez, A.; Rodríguez-Silverio, J.; Flores-Murrieta, F.J.; Granados-Soto, V.; Rocha-González, H.I. Spinal 5-HT4 and 5-HT6 Receptors Contribute to the Maintenance of Neuropathic Pain in Rats. Pharmacol. Rep. PR 2017, 69, 916–923. [Google Scholar] [CrossRef]
- Grossman, C.J.; Kilpatrick, G.J.; Bunce, K.T. Development of a Radioligand Binding Assay for 5-HT4 Receptors in Guinea-Pig and Rat Brain. Br. J. Pharmacol. 1993, 109, 618–624. [Google Scholar] [CrossRef]
- Ponimaskin, E.G.; Schmidt, M.F.; Heine, M.; Bickmeyer, U.; Richter, D.W. 5-Hydroxytryptamine 4(a) Receptor Expressed in Sf9 Cells Is Palmitoylated in an Agonist-Dependent Manner. Biochem. J. 2001, 353, 627–634. [Google Scholar] [CrossRef]
- Bickmeyer, U.; Heine, M.; Manzke, T.; Richter, D.W. Differential Modulation of I(h) by 5-HT Receptors in Mouse CA1 Hippocampal Neurons. Eur. J. Neurosci. 2002, 16, 209–218. [Google Scholar] [CrossRef]
- Ho, S.S.N.; Chow, B.K.C.; Yung, W.-H. Serotonin Increases the Excitability of the Hypothalamic Paraventricular Nucleus Magnocellular Neurons. Eur. J. Neurosci. 2007, 25, 2991–3000. [Google Scholar] [CrossRef]
- Baudry, A.; Alleaume-Butaux, A.; Dimitrova-Nakov, S.; Goldberg, M.; Schneider, B.; Launay, J.-M.; Kellermann, O. Essential Roles of Dopamine and Serotonin in Tooth Repair: Functional Interplay Between Odontogenic Stem Cells and Platelets. Stem Cells 2015, 33, 2586–2595. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitayama, E.; Kimura, M.; Ouchi, T.; Furusawa, M.; Shibukawa, Y. Functional Expression of IP, 5-HT4, D1, A2A, and VIP Receptors in Human Odontoblast Cell Line. Biomolecules 2023, 13, 879. https://doi.org/10.3390/biom13060879
Kitayama E, Kimura M, Ouchi T, Furusawa M, Shibukawa Y. Functional Expression of IP, 5-HT4, D1, A2A, and VIP Receptors in Human Odontoblast Cell Line. Biomolecules. 2023; 13(6):879. https://doi.org/10.3390/biom13060879
Chicago/Turabian StyleKitayama, Eri, Maki Kimura, Takehito Ouchi, Masahiro Furusawa, and Yoshiyuki Shibukawa. 2023. "Functional Expression of IP, 5-HT4, D1, A2A, and VIP Receptors in Human Odontoblast Cell Line" Biomolecules 13, no. 6: 879. https://doi.org/10.3390/biom13060879
APA StyleKitayama, E., Kimura, M., Ouchi, T., Furusawa, M., & Shibukawa, Y. (2023). Functional Expression of IP, 5-HT4, D1, A2A, and VIP Receptors in Human Odontoblast Cell Line. Biomolecules, 13(6), 879. https://doi.org/10.3390/biom13060879