

The Role of Spermidine and Its Key Metabolites in Important, Pathogenic Human Viruses and in Parasitic Infections Caused by Plasmodium falciparum and Trypanosoma brucei


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
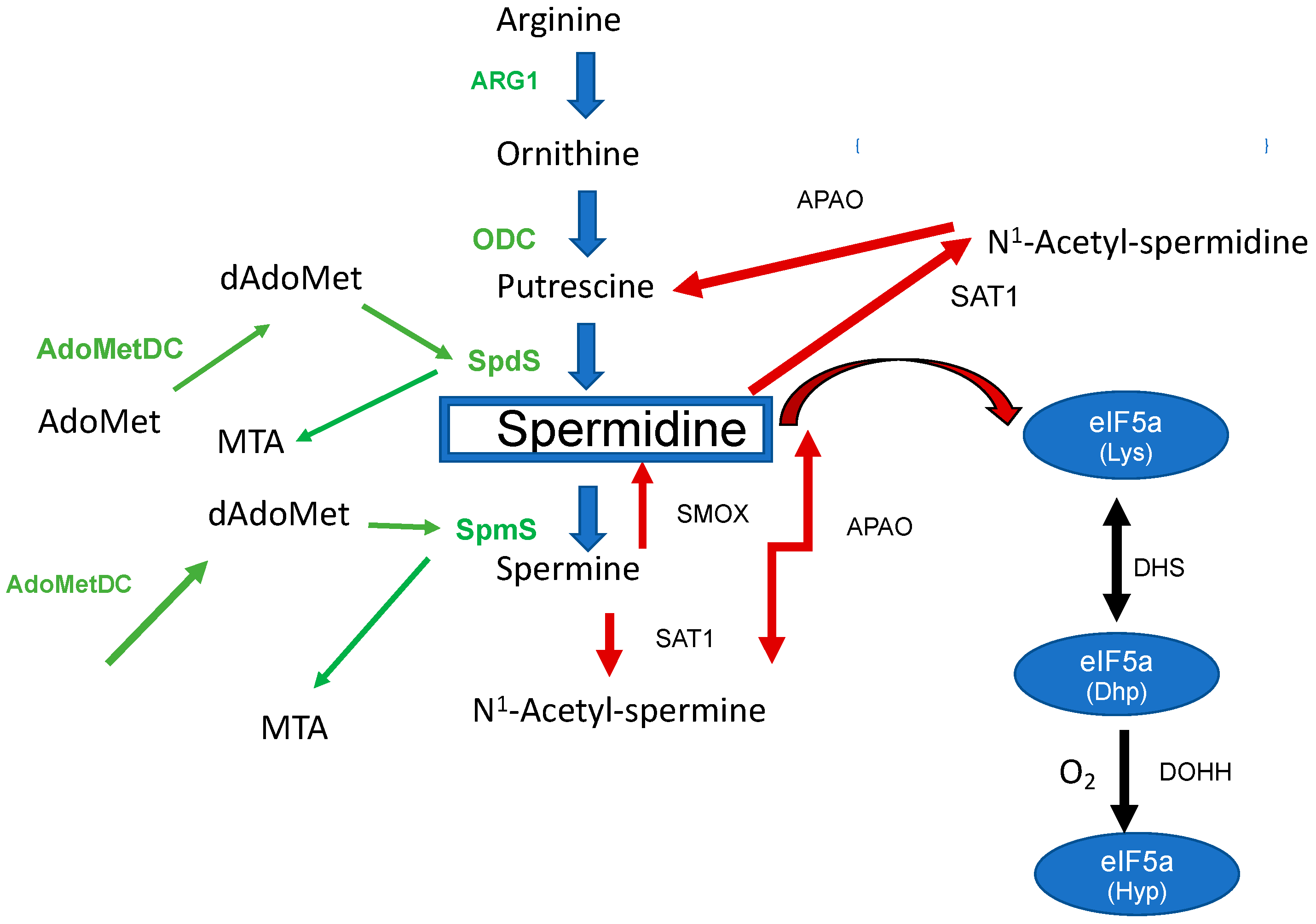
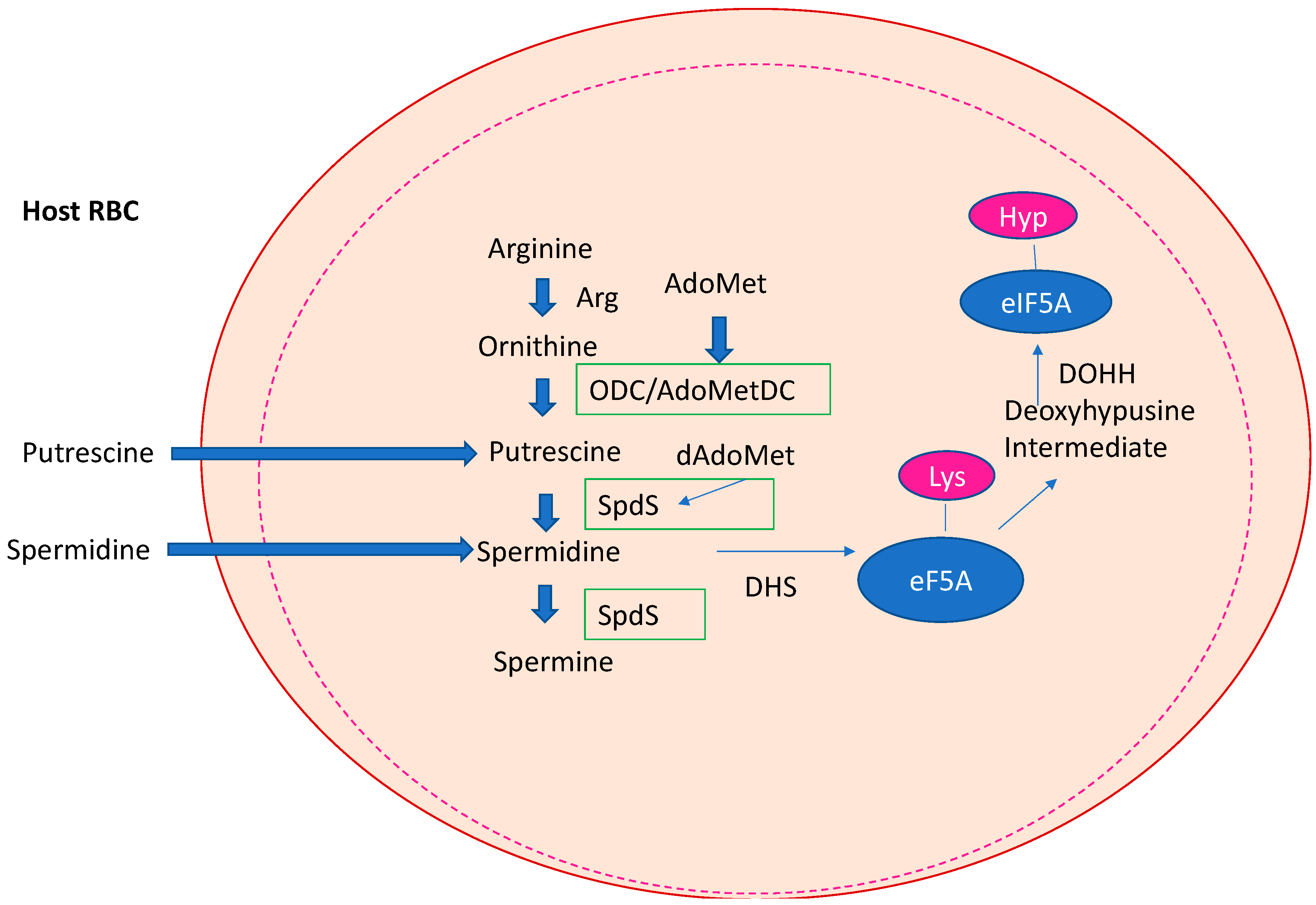
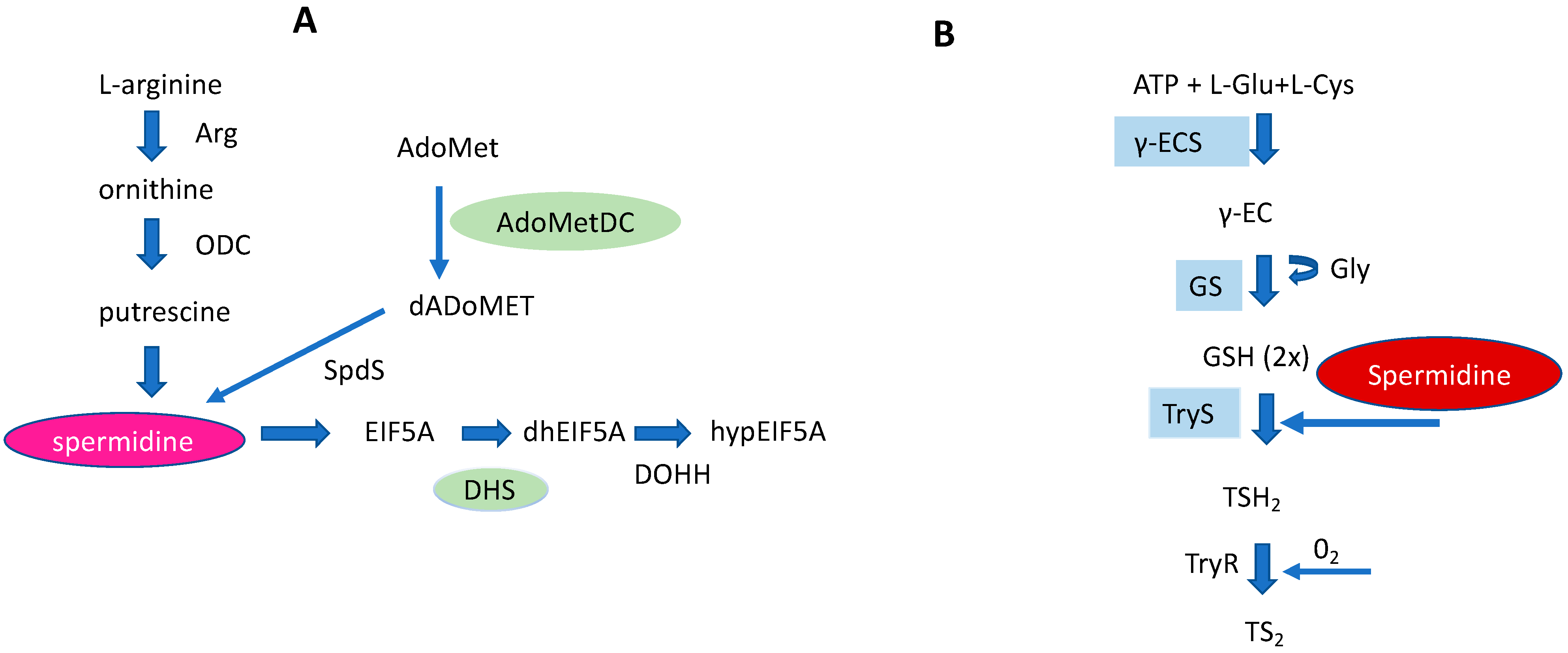
The Biosynthesis and Metabolism of the Triamine Spermidine in the Human Host Cell and Its Regulation
2. The Impact of Spermidine and Its Metabolites in Infection with Human Pathogenic Viruses
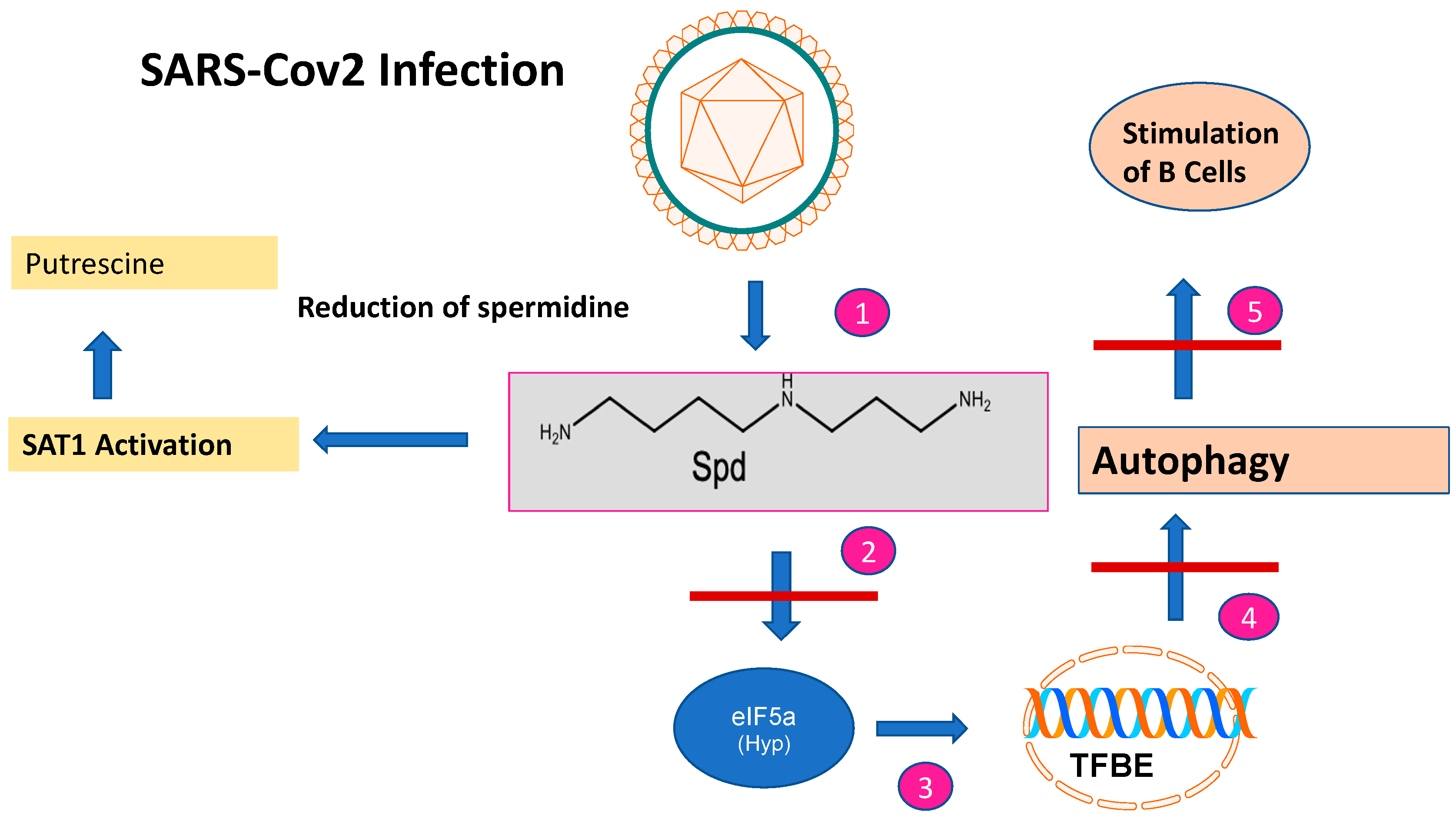
2.1. Dysregulation of Spermidine Metabolism Determines SARS-CoV-2 Infection
2.2. The Role of Hypusinated EIF-5A and PAs in Ebola Propagation
2.3. Hypusinated eIF5A Is a Cofactor of HIV-1 Rev Regulatory Protein
3. Spermidine as a Hallmark in the Apicomplexan Parasite P. falciparum
4. Spermidine Biosynthesis and Its Metabolizing Pathways in the Trypanosomatids Are Unique
5. The Impact to Develop Inhibitors against Enzymes Involved in Spermidine Biosynthesis and Metabolism Treating Human African Trypanomiasis (HAT)
6. Conclusions and Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pegg, A.E. Mammalian polyamine metabolism and function. IUBMB Life 2010, 61, 880–894. [Google Scholar] [CrossRef] [PubMed]
- Zahedi, K.; Barone, S.; Soleimani, M. Polyamines and Their Metabolism: From the Maintenance of Physiological Homeostasis to the Mediation of Disease. Med. Sci. 2022, 15, 38. [Google Scholar] [CrossRef]
- Bello-Fernandez, C.; Packham, G.; Cleveland, J.L. The ornithine decarboxylase gene is a transcriptional target of c-Myc. Proc. Natl. Acad. Sci. USA 1993, 90, 7804–7808. [Google Scholar] [CrossRef] [PubMed]
- Murakami, Y.; Matsufuji, S.; Tanaka, K.; Ichihara, A.; Hayashi, S. Involvement of the proteasome and antizyme in ornithine decarboxylase degradation by a reticulocyte lysate. Biochem. J. 1993, 295, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Devereux, W.; Stewart, T.M.; Casero, R.A., Jr. Cloning and characterization of human polyamine-modulated factor-1, a transcriptional cofactor that regulates the transcription of the spermidine/spermine N(1)-acetyltransferase gene. J. Biol. Chem. 1999, 274, 22095–22101. [Google Scholar] [CrossRef]
- Tomitori, H.; Nenoi, M.; Mita, K.; Daino, K.; Igarashi, K.; Ichimura, S. Functional characterization of the human spermidine/spermine N(1)-acetyltransferase gene promoter. Biochim. Biophys. Acta 2002, 1579, 180–184. [Google Scholar] [CrossRef]
- Bewley, M.C.; Graziano, V.; Jiang, J.; Matz, E.; Studier, F.W.; Pegg, A.E.; Coleman, C.S.; Flanagan, J.M. Structures of wild-type and mutant human spermidine/spermine N1-acetyltransferase, a potential therapeutic drug target. Proc. Natl. Acad. Sci. USA 2006, 103, 2063–2068. [Google Scholar] [CrossRef]
- Murray-Stewart, T.; Sierra, J.C.; Piazuelo, M.B.; Mera, R.M.; Chaturvedi, R.; Bravo, L.E.; Correa, P.; Schneider, B.G.; Wilson, K.T.; Casero, R.A. Epigenetic silencing of miR-124 prevents spermine oxidase regulation: Implications for Helicobacter pylori-induced gastric cancer. Oncogene 2016, 35, 5480–5488. [Google Scholar] [CrossRef]
- Kashiwagi, K.; Igarashi, K. Molecular characteristics of toxicity of acrolein produced from spermidine. Biomolecules 2023, 13, 298. [Google Scholar] [CrossRef]
- Zahedi, K.; Huttinger, F.; Morrison, R.; Murray-Stewart, T.; Casero, R.A.; Strauss, K.I. Polyamine catabolism is enhanced after traumatic brain injury. J. Neurotrauma 2010, 27, 515–525. [Google Scholar] [CrossRef]
- Wang, Y.; Hacker, A.; Murray-Stewart, T.; Fleischer, J.G.; Woster, P.M.; Casero, R.A., Jr. Induction of human spermine oxidase SMO (PAOh1) is regulated at the levels of new mRNA synthesis, mRNA stabilization and newly synthesized protein. Biochem. J. 2005, 386, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Cooper, H.L.; Folk, J.E. Identification of hypusine, an unusual amino acid, in a protein from human lymphocytes and of spermidine as its biosynthetic precursor. Proc. Natl. Acad. Sci. USA 1981, 78, 2869–2873. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Wolff, E.C. Hypusine, a polyamine-derived amino acid critical for eukaryotic translation. J. Biol. Chem. 2018, 293, 18710–18718. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; Sakai, N.; Böttger, L.H.; Klinke, S.; Hauber, J.; Trautwein, A.; Hilgenfeld, R. Crystal structure of the peroxo-diiron(III) intermediate of deoxyhypusine hydroxylase, an oxygenase involved in hypusination. Structure 2015, 23, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, E.; Shin, B.S.; Woolstenhulme, C.J.; Kim, J.R.; Saini, P.; Buskirk, A.R.; Dever, T.E. eIF5A promotes translation of polyproline motifs. Mol. Cell 2013, 51, 35–45. [Google Scholar] [CrossRef]
- Maier, B.; Ogihara, T.; Trace, A.P.; Tersey, S.A.; Robbins, R.D.; Chakrabarti, S.K.; Nunemaker, C.S.; Stull, N.D.; Taylor, C.A.; Thompson, J.E.; et al. The unique hypusine modification of eIF5A promotes islet beta cell inflammation and dysfunction in mice. J. Clin. Investig. 2010, 120, 2156–2170. [Google Scholar] [CrossRef]
- Anderson-Baucum, E.; Piñeros, A.R.; Kulkarni, A.; Webb-Robertson, B.J.; Maier, B.; Anderson, R.M.; Wu, W.; Tersey, S.A.; Mastracci, T.L.; Casimiro, I.; et al. Deoxyhypusine synthase promotes a pro-inflammatory macrophage phenotype. Cell Metab. 2021, 33, 1883–1893. [Google Scholar] [CrossRef]
- Ganapathi, M.; Padgett, L.R.; Yamada, K.; Devinsky, O.; Willaert, R.; Person, R.; Au, P.B.; Tagoe, J.; McDonald, M.; Karlowicz, D.; et al. Recessive Rare Variants in Deoxyhypusine Synthase, an Enzyme Involved in the Synthesis of Hypusine, Are Associated with a Neurodevelopmental Disorder. Am. J. Hum. 2019, 104, 287–298. [Google Scholar] [CrossRef]
- Ziegler, A.; Steindl, K.; Hanner, A.S.; Kumar Kar, R.; Prouteau, C.; Boland, A.; Deleuze, J.F.; Coubes, C.; Bézieau, S.; Küry, S.; et al. Bi-allelic variants in DOHH, catalyzing the last step of hypusine biosynthesis, are associated with a neurodevelopmental disorder. Am. J. Hum. Genet. 2022, 109, 1549–1558. [Google Scholar] [CrossRef]
- Gassen, N.C.; Papies, J.; Bajaj, T.; Emanuel, J.; Dethloff, F.; Chua, R.L.; Trimpert, J.; Heinemann, N.; Niemeyer, C.; Weege, F.; et al. SARS-CoV-2-mediated dysregulation of metabolism and autophagy uncovers host-targeting antivirals. Nat. Commun. 2021, 12, 3818. [Google Scholar] [CrossRef]
- Sridhar, S.; Nicholls, J. Pathophysiology of infection with SARS-CoV-2-What is known and what remains a mystery. Respirology 2021, 26, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of autophagyby spermidine promotes longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Mounce, B.C.; Poirier, E.Z.; Passoni, G.; Simon-Loriere, E.; Cesaro, T.; Prot, M.; Stapleford, K.A.; Moratorio, G.; Sakuntabhai, A.; Levraud, J.P.; et al. Interferon-induced spermidine-spermine acetyltransferase and polyamine depletion restrict Zika and Chikungunya viruses. Cell Host Microbe 2016, 20, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Alsaleh, G.; Feltham, J.; Sun, Y.; Napolitano, G.; Riffelmacher, T.; Charles, P.; Frau, L.; Hublitz, P.; Yu, Z.; et al. Polyamines control eIF5A hypusination, TFEB translation, and autophagy to reverse B cell senescence. Mol. Cell 2019, 76, 110–125. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Crozier, I.; Fischer, W.A., 2nd; Hewlett, A.; Kraft, C.S.; Vega, M.A.; Soka, M.J.; Wahl, V.; Griffiths, A.; Bollinger, L.; et al. Ebola virus disease. Nat. Rev. Dis. Primers 2020, 20, 13. [Google Scholar] [CrossRef]
- European Commission. Vaccine against Ebola: Commission Grants First-Ever Market Authorisation. Market Authorization in Europe for Ebola Virus Vaccine. 2019. Available online: https://ec.europa.eu/commission/presscorner/detail/en/IP_19_6246 (accessed on 11 November 2019).
- Santoro, M.G.; Carafoli, E. From Ebola to COVID-19. Biochem. Biophys. Res. Commun. 2021, 29, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Tzitzi, E.; Pyrpasopoulou, A.; Kalmoukos, P.; Kefas, A.; Kyrana, Z.; Doukelis, P.; Varouktsi, A.; Imprialos, K.; Doumas, M. Casivirimab/Imdevimab effect on COVID-19 outcome and reinfection in a real-world SARS-CoV-2 variant transition period setting. Monoclon. Antib. Immundiagn. Immunotherap. 2023, 42, 48–50. [Google Scholar] [CrossRef]
- Firpo, M.R.; Mounce, B.C. Diverse functions of polyamines in virus infections. Biomolecules 2020, 10, 628. [Google Scholar] [CrossRef]
- Kaiser, A.; Vollmert, M.; Tholl, D.; Graves, M.V.; Gurnon, J.R.; Xing, W.; Lisec, A.D.; Nickerson, K.W.; Van Etten, J.L. Chlorella virus PBCV-1 encodes a functional homospermidine synthase. Virology 1999, 263, 254–262. [Google Scholar] [CrossRef]
- Olsen, M.E.; Cressey, T.N.; Mühlberger, E.; Connor, J.H. Differential mechanisms for the involvement of polyamines and hypusinated eIF5A in Ebola Virus gene expression. J. Virol. 2018, 26, e01260-18. [Google Scholar] [CrossRef]
- UNAIDS Global HIV & AIDS Statistics—2021 Fact Sheet. Available online: www.unaids.org (accessed on 27 July 2022).
- Bevec, D.; Hauber, J. Eukaryotic initiation factor 5A activity and HIV-1 Rev function. Biol. Signals 1997, 6, 124–133. [Google Scholar] [CrossRef]
- Hoffmann, D.; Schwarck, D.; Banning, C.; Brenner, M.; Mariyanna, L.; Krepstakies, M.; Schindler, M.; Millar, D.P.; Hauber, J. Formation of trans-activation competent HIV-1 Rev:RRE complexes requires the recruitment of multiple protein activation domains. PLoS ONE 2012, 7, e38305. [Google Scholar] [CrossRef] [PubMed]
- Hauber, I.; Bevec, D.; Heukeshoven, J.; Krätzer, F.; Horn, F.; Choidas, A.; Harrer, T.; Hauber, J. Identification of cellular deoxyhypusine synthase as a novel target for antiretroviral therapy. J. Clin. Investig. 2005, 115, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Schäfer, B.; Hauber, I.; Bunk, A.; Heukeshoven, J.; Düsedau, A.; Bevec, D.; Hauber, J. Inhibition of multidrug-resistant HIV-1 by interference with cellular S-adenosylmethionine decarboxylase activity. J. Infect. Dis. 2006, 194, 740–750. [Google Scholar] [CrossRef][Green Version]
- Capone, C.; Cervelli, M.; Angelucci, E.; Colasanti, M.; Macone, A.; Mariottini, P.; Persichini, T. A role for spermine oxidase as a mediator of reactive oxygen species production in HIV-Tat-induced neuronal toxicity. Free Rad. Biol. Med. 2013, 63, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Arisue, N.; Hashimoto, T. Phylogeny and evolution of apicoplasts and apicomplexan parasites. Parasitol Int. 2015, 64, 254–259. [Google Scholar] [CrossRef]
- Ghebreyesus, T.A. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021; ISBN 978-92-4-004050-2.
- Zavala, F. RTS,S: The first malaria vaccine. J. Clin. Investig. 2022, 132, e156588. [Google Scholar] [CrossRef]
- Nkumama, I.N.; Osier, F.H.A. Malaria vaccine roller coaster. Nat. Microbiol. 2021, 6, 1345–1346. [Google Scholar] [CrossRef]
- Maier, A.G.; Matuschewski, K.; Zhang, M.; Rug, M. Plasmodium falciparum. Trends Parasitol. 2019, 35, 481–482. [Google Scholar] [CrossRef]
- Ramya, T.N.; Surolia, N.; Surolia, A. Polyamine synthesis and salvage pathways in the malaria parasite Plasmodium falciparum. Biochem. Biophys. Res. Commun. 2006, 348, 579–584. [Google Scholar] [CrossRef]
- Niemand, J.; Louw, A.I.; Birkholtz, L.; Kirk, K. Polyamine uptake by the intraerythrocytic malaria parasite, Plasmodium falciparum. Int. J. Parasitol. 2012, 42, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Da’dara, A.; Lüersen, K.; Wrenger, C.; Das Gupta, R.; Madhubala, R.; Walter, R.D. In the human malaria parasite Plasmodium falciparum, polyamines are synthesized by a bifunctional ornithine decarboxylase, S-adenosylmethionine decarboxylase. J. Biol. Chem. 2000, 275, 8097–8102. [Google Scholar] [CrossRef] [PubMed]
- Haider, N.; Eschbach, M.L.; Dias Sde, S.; Gilberger, T.W.; Walter, R.D.; Lüersen, K. The spermidine synthase of the malaria parasite Plasmodium falciparum: Molecular and biochemical characterisation of the polyamine synthesis enzyme. Mol. Biochem. Parasitol. 2005, 142, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Frommholz, D.; Kusch, P.; Blavid, R.; Scheer, H.; Tu, J.M.; Marcus, K.; Zhao, K.H.; Atemnkeng, V.; Marciniak, J.; Kaiser, A.E. Completing the hypusine pathway in Plasmodium. FEBS J. 2009, 276, 5881–5891. [Google Scholar] [CrossRef]
- Kersting, D.; Krüger, M.; Sattler, J.M.; Mueller, A.K.; Kaiser, A. A suggested vital function for eIF-5A and dhs genes during murine malaria blood-stage infection. FEBS Open Bio 2016, 23, 860–872. [Google Scholar] [CrossRef]
- Kamil, M.; Kina, U.Y.; Deveci, G.; Akyuz, S.N.; Yilmaz, I.; Aly, A.S.I. Mitochondrial Spermidine Synthase is essential for Blood-stage growth of the Malaria Parasite. Microbiol. Res. 2022, 265, 20127181. [Google Scholar] [CrossRef]
- Sprenger, J.; Carey, J.; Svensson, B.; Wengel, V.; Persson, L. Binding and inhibition of Spermidine Synthase from Plasmodium falciparum and implications for in vitro inhibitor testing. PLoS ONE 2016, 23, e0163442. [Google Scholar] [CrossRef]
- Njuguna, J.T.; Nassar, M.; Hoerauf, A.; Kaiser, A.E. Cloning, expression and functional activity of deoxyhypusine synthase from Plasmodium vivax. BMC Microbiol. 2006, 16, 91. [Google Scholar] [CrossRef]
- Atemnkeng, V.A.; Pink, M.; Schmitz-Spanke, S.; Wu, X.J.; Dong, L.L.; Zhao, M.C.; Laufer, S.; Langer, B.; Kaiser, A. Deoxyhypusine Hydroxylase from Plasmodium vivax, the Neglected Human Malaria Parasite: Molecular Cloning, Expression and Specific Inhibition by the 5-LOX Inhibitor Zileuton. PLoS ONE 2013, 8, e58318. [Google Scholar] [CrossRef]
- Tanaka, Y.; Kurasawa, O.; Yokota, A.; Klein, M.G.; Ono, K.; Saito, B.; Matsumoto, S.; Okaniwa, M.; Ambrus-Aikelin, G.; Morishita, D.; et al. Discovery of Novel Allosteric Inhibitors of Deoxyhypusine Synthase. J. Med. Chem. 2021, 26, 3215–3226. [Google Scholar] [CrossRef]
- Factsheet on Trypanosomes; WHO: Geneva, Switzerland, 2022.
- Ilari, A.; Fiorillo, A.; Genovese, I.; Colotti, G. Polyamine-trypanothione pathway: An update. Fut. Med. Chem. 2017, 9, 61–67. [Google Scholar] [CrossRef]
- Phillips, M.A. Polyamines in protozoan pathogens. J. Biol. Chem. 2018, 293, 18746–18756. [Google Scholar] [CrossRef] [PubMed]
- Willert, E.K.; Phillips, M.A. Regulated expression of an essential allosteric activator of polyamine biosynthesis in African trypanosomes. PLoS Pathog. 2008, 4, e1000183. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.C.; Kaur, H.; Blessington, B.; Kelly, J.M.; Wilkinson, S.R. Validation of spermidine synthase as a drug target in African trypanosomes. Biochem. J. 2008, 409, 563–569. [Google Scholar] [CrossRef]
- Xiao, Y.; McCloskey, D.E.; Phillips, M.A. RNA interference-mediated silencing of ornithine decarboxylase and spermidine synthase genes in Trypanosoma brucei provides insight into regulation of polyamine biosynthesis. Eukaryot. Cell 2009, 8, 747–755. [Google Scholar] [CrossRef]
- Creek, D.J.; Mazet, M.; Achcar, F.; Anderson, J.; Kim, D.H.; Kamour, R.; Morand, P.; Millerioux, Y.; Biran, M.; Kerkhoven, E.J.; et al. Probing the metabolic network in bloodstream-form Trypanosoma brucei using untargeted metabolomics with stable isotope labelled glucose. PLoS Pathog. 2015, 11, e1004689. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, S.; Jones, D.C.; Wyllie, S.; Fairlamb, A.H.; Phillips, M.A. Allosteric activation of trypanosomatid deoxyhypusine synthase by a catalytically dead paralog. J. Biol. Chem. 2013, 288, 15256–15267. [Google Scholar] [CrossRef] [PubMed]
- Ariyanayagam, M.R.; Fairlamb, A.H. Diamine auxotrophy may be a universal feature of Trypanosoma cruzi epimastigotes. Mol. Biochem. Parasitol. 1997, 84, 111–121. [Google Scholar] [CrossRef]
- Willert, E.K.; Phillips, M.A. Cross-species activation of trypanosome S-adenosylmethionine decarboxylase by the regulatory subunit prozyme. Mol. Biochem. Parasitol. 2009, 168, 1–6. [Google Scholar] [CrossRef][Green Version]
- Yamasaki, K.; Tani, O.; Tateishi, Y.; Tanabe, E.; Namatame, I.; Niimi, T.; Furukawa, K.; Sakashita, H. An NMR biochemical assay for fragment-based drug discovery: Evaluation of an inhibitor activity on spermidine synthase of Trypanosoma cruzi. J. Med. Chem. 2016, 59, 2261–2266. [Google Scholar] [CrossRef]
- Aslett, M.; Aurrecoechea, C.; Berriman, M.; Brestelli, J.; Brunk, B.P.; Carrington, M.; Depledge, D.P.; Fischer, S.; Gajria, B.; Gao, X.; et al. Tri- TrypDB. A functional genomic resource for the Trypanosomatidae. Nucleic. Acids Res. 2010, 38, 457–462. [Google Scholar] [CrossRef]
- Wéry, M. Drug used in the treatment of sleeping sickness (human African trypanosomiasis: HAT). Int. J. Antimicrob. Agents 1994, 4, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Phillips, M.A.; Coffino, P.; Wang, C.C. Cloning and sequencing of the ornithine decarboxylase gene from Trypanosoma brucei. Implications for enzyme turnover and selective α-difluoromethylornithine inhibition. J. Biol. Chem. 1987, 262, 8721–8727. [Google Scholar] [CrossRef] [PubMed]
- Afanador, G.A.; Tomchick, D.R.; Phillips, M.A. Trypanosomatid deoxyhypusine synthase activity is dependent on shared active site complementation between pseudoenzyme paralogs. Structure 2018, 26, 1499–1512. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaiser, A. The Role of Spermidine and Its Key Metabolites in Important, Pathogenic Human Viruses and in Parasitic Infections Caused by Plasmodium falciparum and Trypanosoma brucei. Biomolecules 2023, 13, 803. https://doi.org/10.3390/biom13050803
Kaiser A. The Role of Spermidine and Its Key Metabolites in Important, Pathogenic Human Viruses and in Parasitic Infections Caused by Plasmodium falciparum and Trypanosoma brucei. Biomolecules. 2023; 13(5):803. https://doi.org/10.3390/biom13050803
Chicago/Turabian StyleKaiser, Annette. 2023. "The Role of Spermidine and Its Key Metabolites in Important, Pathogenic Human Viruses and in Parasitic Infections Caused by Plasmodium falciparum and Trypanosoma brucei" Biomolecules 13, no. 5: 803. https://doi.org/10.3390/biom13050803
APA StyleKaiser, A. (2023). The Role of Spermidine and Its Key Metabolites in Important, Pathogenic Human Viruses and in Parasitic Infections Caused by Plasmodium falciparum and Trypanosoma brucei. Biomolecules, 13(5), 803. https://doi.org/10.3390/biom13050803