

Mass Spectrometry-Based Proteomic Profiling of a Silvaner White Wine

 ,
,  , , , and
, , , and
Abstract

1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Silvaner Wine
2.3. Technical Scale Isolation and Analysis of Silvaner Wine Colloids
2.4. Protein Content and Visualization
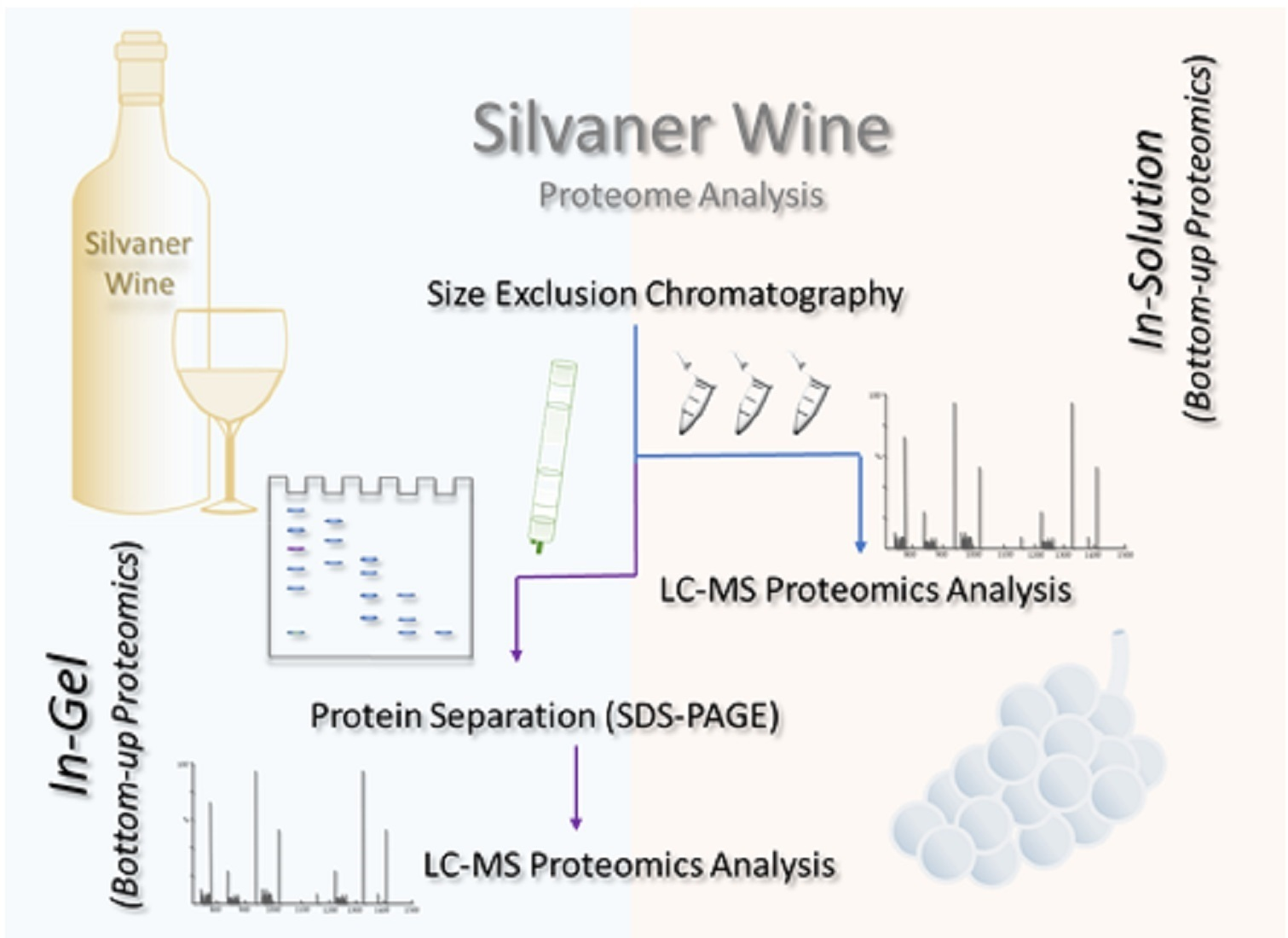
2.5. MS-Based Proteomics Analysis of Proteins from a Silvaner Wine
2.5.1. In-solution Digestion: Protein Fractionation by SEC Chromatography
2.5.2. In-gel Digestion: Proteins Fractionated by Gel Electrophoresis
2.5.3. Liquid Chromatography Mass Spectrometry (LC-MS) Analysis
2.5.4. MS Data Analysis
3. Results
3.1. Protein Fractionation and Visualization
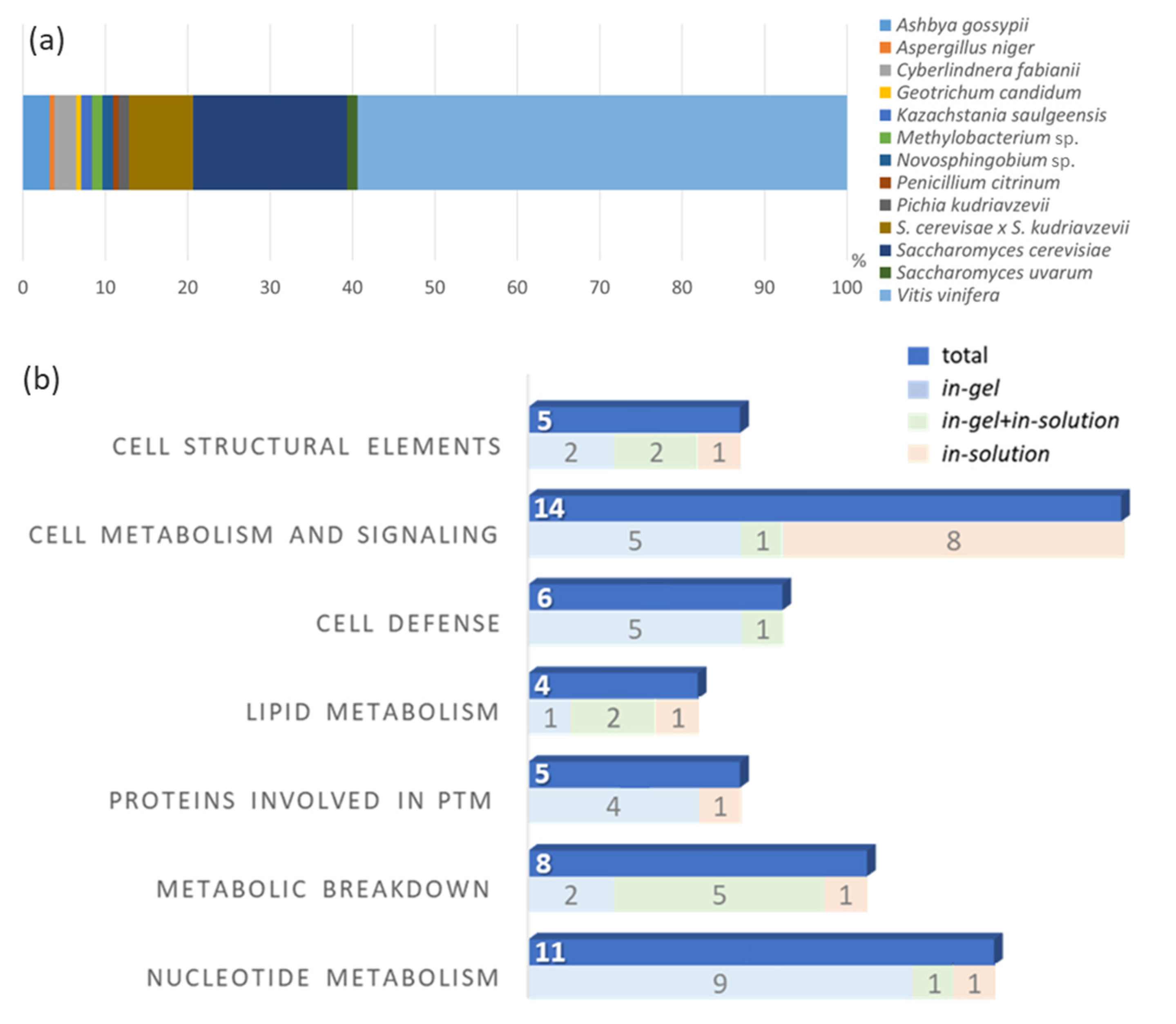
3.2. MS-Based Proteomics Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sefc, K.M.; Steinkellner, H.; Glössl, J.; Kampfer, S.; Regner, F. Reconstruction of a grapevine pedigree by microsatellite analysis. Theor. Appl. Genet. 1998, 97, 227–231. [Google Scholar] [CrossRef]
- Fröhlich, R. Studien zur Geschichte, Kunst und Kultur der Zisterzienser. In Die Zisterzienser und ihre Weinberge in Brandenburg Studien zur Geschichte; Lukas Verlag: Brandenburg, Germany, 2010; Volume 30. [Google Scholar]
- Deutsches Weininstitut (2022). Deutscher Wein Statistik 2022/2023. Available online: https://www.deutscheweine.de/fileadmin/user_upload/Website/Service/Downloads/PDF/Statistik_2022-2023.pdf (accessed on 1 December 2022).
- Bock, A.; Sparks, T.; Estrella, N.; Menzel, A. Changes in the phenology and composition of wine from Franconia, Germany. Clim. Res. 2011, 50, 69–81. [Google Scholar] [CrossRef]
- Pasquier, G.; Feilhes, C.; Dufourcq, T.; Geffroy, O. Potential contribution of climate change to the protein haze of white wines from the french southwest region. Foods 2021, 10, 1355. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.R.; Gawel, R.; Francis, I.L.; Waters, E.J. The influence of interactions between major white wine components on the aroma, flavour and texture of model white wine. Food Qual. Prefer. 2008, 19, 596–607. [Google Scholar] [CrossRef]
- Jones-Moore, H.R.; Jelley, R.E.; Marangon, M.; Fedrizzi, B. The interactions of wine polysaccharides with aroma compounds, tannins, and proteins, and their importance to winemaking. Food Hydrocoll. 2022, 123, 107150. [Google Scholar] [CrossRef]
- Van Sluyter, S.C.; McRae, J.M.; Falconer, R.J.; Smith, P.A.; Bacic, A.; Waters, E.J.; Marangon, M. Wine protein haze: Mechanisms of formation and advances in prevention. J. Agric. Food Chem. 2015, 63, 4020–4030. [Google Scholar] [CrossRef]
- Cilindre, C.; Castro, A.J.; Clément, C.; Jeandet, P.; Marchal, R. Influence of Botrytis cinerea infection on Champagne wine proteins (characterized by two-dimensional electrophoresis/immunodetection) and wine foaming properties. Food Chem. 2007, 103, 139–149. [Google Scholar] [CrossRef]
- Heinonen, M.; Rein, D.; Satué-Gracia, M.T.; Huang, S.W.; German, J.B.; Frankel, E.N. Effect of protein on the antioxidant activity of phenolic compounds in a lecithin-liposome oxidation system. J. Agric. Food Chem. 1998, 46, 917–922. [Google Scholar] [CrossRef]
- Peñas, E.; di Lorenzo, C.; Uberti, F.; Restani, P. Allergenic proteins in enology: A review on technological applications and safety aspects. Molecules 2015, 20, 13144–13164. [Google Scholar] [CrossRef]
- D’Amato, A.; Fasoli, E.; Kravchuk, A.V.; Righetti, P.G. Mehercules, adhuc Bacchus! The debate on wine proteomics continues. J. Proteome Res. 2011, 10, 3789–3801. [Google Scholar] [CrossRef]
- Giribaldi, M.; Giuffrida, M.G. Heard it through the grapevine: Proteomic perspective on grape and wine. J. Proteom. 2010, 73, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.C.; Pérez-Álvarez, E.P.; Peregrina, F.; Martínez de Toda, F. Relationships between grape composition of Tempranillo variety and available soil water and water stress under different weather conditions. Sci. Hortic. 2020, 262, 109063. [Google Scholar] [CrossRef]
- Wigand, P.; Tenzer, S.; Schild, H.; Decker, H. Analysis of protein composition of red wine in comparison with rosé and white wines by electrophoresis and high-pressure liquid chromatography−mass spectrometry (HPLC-MS). J. Agric. Food Chem. 2009, 57, 4328–4333. [Google Scholar] [CrossRef]
- Cosme, F.; Fernandes, C.; Ribeiro, T.; Filipe-Ribeiro, L.; Nunes, F.M. White wine protein instability: Mechanism, quality control and technological alternatives for wine stabilization—An overview. Beverages 2020, 6, 19. [Google Scholar] [CrossRef]
- Ferreira, R.B.; Piçarra-Pereira, M.A.; Monteiro, S.; Loureiro, V.B.; Teixeira, A.R. The wine proteins. Trends Food Sci. Technol. 2001, 12, 230–239. [Google Scholar] [CrossRef]
- Pocock, K.F.; Hayasaka, Y.; Peng, Z.; Williams, P.J.; Waters, E.J. The effect of mechanical harvesting and long-distance transport on the concentration of haze-forming proteins in grape juice. Aust. J. Grape Wine Res. 1998, 4, 23–29. [Google Scholar] [CrossRef]
- Tian, B.; Harrison, R.; Morton, J.; Jaspers, M.; Hodge, S.; Grose, C.; Trought, M. Extraction of pathogenesis-related proteins and phenolics in Sauvignon blanc as affected by grape harvesting and processing conditions. Molecules 2017, 22, 1164. [Google Scholar] [CrossRef]
- Seidel, L.; Albuquerque, W.; Happel, K.; Ghezellou, P.; Gand, M.; Spengler, B.; Zorn, H.; Will, F.; Schweiggert, R. Composition, zeta potential and molar mass distribution of 20 must and wine colloids from five different cultivars obtained during four consecutive vintages. J. Agric. Food Chem. 2023. [Google Scholar] [CrossRef]
- Dupin, I.V.S.; McKinnon, B.M.; Ryan, C.; Boulay, M.; Markides, A.J.; Jones, G.P.; Williams, P.J.; Waters, E.J. Saccharomyces cerevisiae mannoproteins that protect wine from protein haze: Their release during fermentation and lees contact and a proposal for their mechanism of action. J. Agric. Food Chem. 2000, 48, 3098–3105. [Google Scholar] [CrossRef]
- Cilindre, C.; Jégou, S.; Hovasse, A.; Schaeffer, C.; Castro, A.J.; Clément, C.; van Dorsselaer, A.; Jeandet, P.; Marchal, R. Proteomic approach to identify champagne wine proteins as modified by Botrytis cinerea infection. J. Proteome Res. 2008, 7, 1199–1208. [Google Scholar] [CrossRef]
- Weber, P.; Steinhart, H.; Paschke, A. Investigation of the allergenic potential of wines fined with various proteinogenic fining agents by ELISA. J. Agric. Food Chem. 2007, 55, 3127–3133. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, W.; Seidel, L.; Zorn, H.; Will, F.; Gand, M. Haze formation and the challenges for peptidases in wine protein fining. J. Agric. Food Chem. 2021, 69, 14402–14414. [Google Scholar] [CrossRef]
- Albuquerque, W.; Sturm, P.; Schneider, Q.; Ghezellou, P.; Seidel, L.; Bakonyi, D.; Will, F.; Spengler, B.; Zorn, H.; Gand, M. Recombinant thaumatin-like protein (rTLP) and chitinase (rCHI) from Vitis vinifera as models for wine haze formation. Molecules 2022, 27, 6409. [Google Scholar] [CrossRef]
- Flamini, R.; de Rosso, M. Mass spectrometry in the analysis of grape and wine proteins. Expert Rev. Proteom. 2006, 3, 321–331. [Google Scholar] [CrossRef] [PubMed]
- Hines, K.M.; Ford, G.C.; Klaus, K.A.; Irving, B.A.; Ford, B.L.; Johnson, K.L.; Lanza, I.R.; Nair, K.S. Application of high-resolution mass spectrometry to measure low abundance isotope enrichment in individual muscle proteins. Anal. Bioanal. Chem. 2015, 407, 4045–4052. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Kerr, E.D.; Pegg, C.L.; Schulz, B.L. Proteomics and glycoproteomics of beer and wine. Proteomics 2022, 22, e2100329. [Google Scholar] [CrossRef]
- Aebersold, R.; Mann, M. Mass-spectrometric exploration of proteome structure and function. Nature 2016, 537, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.W. Profiling of soluble proteins in wine by nano-high-performance liquid chromatography/tandem mass spectrometry. J. Agric. Food Chem. 2004, 52, 7258–7263. [Google Scholar] [CrossRef]
- Choksawangkarn, W.; Edwards, N.; Wang, Y.; Gutierrez, P.; Fenselau, C. Comparative study of workflows optimized for in-gel, in-solution, and on-filter proteolysis in the analysis of plasma membrane proteins. J. Proteome Res. 2012, 11, 3030–3034. [Google Scholar] [CrossRef]
- Albuquerque, W.; Ghezellou, P.; Li, B.; Spengler, B.; Will, F.; Zorn, H.; Gand, M. Identification of intact peptides by top-down peptidomics reveals cleavage spots in thermolabile wine proteins. Food Chem. 2021, 363, 130437. [Google Scholar] [CrossRef]
- Ahlborn, J.; Stephan, A.; Meckel, T.; Maheshwari, G.; Rühl, M.; Zorn, H. Upcycling of food industry side streams by basidiomycetes for production of a vegan protein source. Int. J. Recycl. Org. Waste Agric. 2019, 8, 447–455. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Martin, M.-J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. mBio 2016, 7, e00631-16. [Google Scholar] [CrossRef]
- Marangon, M.; van Sluyter, S.C.; Haynes, P.A.; Waters, E.J. Grape and wine proteins: Their fractionation by hydrophobic interaction chromatography and identification by chromatographic and proteomic analysis. J. Agric. Food Chem. 2009, 57, 4415–4425. [Google Scholar] [CrossRef]
- D’Amato, A.; Kravchuk, A.V.; Bachi, A.; Righetti, P.G. Noah’s nectar: The proteome content of a glass of red wine. J. Proteom. 2010, 73, 2370–2377. [Google Scholar] [CrossRef]
- Oliva, E.; Mir-Cerdà, A.; Sergi, M.; Sentellas, S.; Saurina, J. Characterization of sparkling wine based on polyphenolic profiling by liquid chromatography coupled to mass spectrometry. Fermentation 2023, 9, 223. [Google Scholar] [CrossRef]
- Burns, R.L.; Alexander, R.; Snaychuk, L.; Edwards, J.C.; Fitzgerald, N.; Gao, P.; Quan, D.; Douvris, C.; Vaughan, T.; Bussan, D.D. A fast, straightforward and inexpensive method for the authentication of Baijiu spirit samples by fluorescence spectroscopy. Beverages 2021, 7, 65. [Google Scholar] [CrossRef]
- Lorenzini, M.; Mainente, F.; Zapparoli, G.; Cecconi, D.; Simonato, B. Post-harvest proteomics of grapes infected by Penicillium during withering to produce Amarone wine. Food Chem. 2016, 199, 639–647. [Google Scholar] [CrossRef]
- Swiegers, J.; Bartowsky, E.; Henschke, P.; Pretorius, I. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Meier, M.; Jaeckels, N.; Tenzer, S.; Stoll, M.; Decker, H.; Fronk, P.; Dietrich, H.; Will, F. Impact of drought stress on concentration and composition of wine proteins in Riesling. Eur. Food Res. Technol. 2016, 242, 1883–1891. [Google Scholar] [CrossRef]
- Pocock, K.F.; Hayasaka, Y.; McCarthy, M.G.; Waters, E.J. Thaumatin-like proteins and chitinases, the haze-forming proteins of wine, accumulate during ripening of grape (Vitis vinifera) berries and drought stress does not affect the final levels per berry at maturity. J. Agric. Food Chem. 2000, 48, 1637–1643. [Google Scholar] [CrossRef]
- Pilolli, R.; Visconti, A.; Monaci, L. Rapid and label-free detection of egg allergen traces in wines by surface plasmon resonance biosensor. Anal. Bioanal. Chem. 2015, 407, 3787–3797. [Google Scholar] [CrossRef]
- Jakubowski, N.; Brandt, R.; Stuewer, D.; Eschnauer, H.R.; Görtges, S. Analysis of wines by ICP-MS: Is the pattern of the rare earth elements a reliable fingerprint for the provenance? Fresenius J. Anal Chem. 1999, 364, 424–428. [Google Scholar] [CrossRef]
- Righetti, P.G.; D’Amato, A.; Fasoli, E.; Boschetti, E. In taberna quando sumus: A Drunkard’s Cakewalk through wine proteomics. Food Technol. Biotechnol. 2012, 50, 253–260. [Google Scholar]
- Cereda, A.; Kravchuk, A.V.; D’Amato, A.; Bachi, A.; Righetti, P.G. Proteomics of wine additives: Mining for the invisible via combinatorial peptide ligand libraries. J. Proteomics 2010, 73, 1732–1739. [Google Scholar] [CrossRef]
- Ortea, I.; O’Connor, G.; Maquet, A. Review on proteomics for food authentication. J. Proteom. 2016, 147, 212–225. [Google Scholar] [CrossRef]
- Kuang, L.; Chen, S.; Guo, Y.; Ma, H. Quantitative proteome analysis reveals changes in the protein landscape during grape berry development with a focus on vacuolar transport proteins. Front. Plant Sci. 2019, 10, 641. [Google Scholar] [CrossRef]
- Marsoni, M.; Vannini, C.; Campa, M.; Cucchi, U.; Espen, L.; Bracale, M. Protein extraction from grape tissues by two-dimensional electrophoresis. Vitis J. Grapevine Res. 2005, 44, 181–186. [Google Scholar]
- Sarry, J.E.; Sommerer, N.; Sauvage, F.-X.; Bergoin, A.; Rossignol, M.; Albagnac, G.; Romieu, C. Grape berry biochemistry revisited upon proteomic analysis of the mesocarp. Proteomics 2004, 4, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Enoki, S.; Suzuki, S. Pathogenesis-Related Proteins in Grape. In Grape and Wine Biotechnology; Morata, A., Loira, I., Eds.; IntechOpen: London, UK, 2016. [Google Scholar] [CrossRef]
- Marangon, M.; van Sluyter, S.C.; Neilson, K.A.; Chan, C.; Haynes, P.A.; Waters, E.J.; Falconer, R.J. Roles of grape thaumatin-like protein and chitinase in white wine haze formation. J. Agric. Food Chem. 2011, 59, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Dufrechou, M.; Poncet-Legrand, C.; Sauvage, F.X.; Vernhet, A. Stability of white wine proteins: Combined effect of pH, ionic strength, and temperature on their aggregation. J. Agric. Food Chem. 2012, 60, 1308–1319. [Google Scholar] [CrossRef] [PubMed]
- Okuda, T.; Fukui, M.; Takayanagi, T.; Yokotsuka, K. Characterization of major stable proteins in Chardonnay wine. Food Sci. Technol. Res. 2006, 12, 131–136. [Google Scholar] [CrossRef]
- Rešetar, D.; Marchetti-Deschmann, M.; Allmaier, G.; Katalinić, J.P.; Kraljević Pavelić, S. Matrix assisted laser desorption ionization mass spectrometry linear time-of-flight method for white wine fingerprinting and classification. Food Control. 2016, 64, 157–164. [Google Scholar] [CrossRef]
- Chambery, A.; del Monaco, G.; di Maro, A.; Parente, A. Peptide fingerprint of high quality Campania white wines by MALDI-TOF mass spectrometry. Food Chem. 2009, 113, 1283–1289. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IN-GEL (Exclusively Identified by in-gel Digestion) | ||||||
|---|---|---|---|---|---|---|
| Accession | Gel Band | Description | Organism | MW (kDa) | Reported by (Ref *) | |
| 1 | C8ZG69 | 1 | Ygp1p | Saccharomyces cerevisiae | 37.3 | 5 |
| 2 | G2WD47 | 1 | K7_Spt2p | Saccharomyces cerevisiae | 38.5 | - |
| 3 | H0GMG3 | 1 | Ygp1p | S. cerevisiae x S. kudriavzevii | 37.3 | 5 |
| 4 | A0A438HVN1 | 1 and 12 | Endochitinase EP3 | Vitis vinifera | 27.2 | 1,2,3,4,6 |
| 5 | A0A438ENJ7 | 2 and 6 | Retrovirus-related Pol polyprotein from transposon TNT 1-94 | Vitis vinifera | 33.7 | - |
| 6 | C8Z7L9 | 3 | EC1118_1F14_0100p | Saccharomyces cerevisiae | 53.7 | - |
| 7 | G2WEU0 | 3 | K7_Zpr1p | Saccharomyces cerevisiae | 55.1 | - |
| 8 | A0A061ASV5 | 3 | CYFA0S02e01574g1_1 | Cyberlindnera fabianii | 34.6 | - |
| 9 | A0A1V2L9U0 | 3 | Cytokinesis protein sepH | Cyberlindnera fabianii | 116.3 | - |
| 10 | I9C1P4 | 3 | Aminopeptidase | Novosphingobium sp. | 72 | - |
| 11 | A0A1V2LS96 | 3 | Putative lipase ATG15 | Pichia kudriavzevii | 56.8 | - |
| 12 | A6ZPP5 | 5 | Pathogen-related protein | Saccharomyces cerevisiae | 30.6 | - |
| 13 | C8ZFH3 | 5 | EC1118_1M3_5204p | Saccharomyces cerevisiae | 12.8 | - |
| 14 | A0A438EI04 | 5 and 13 | IAA-amino acid hydrolase ILR1-like 4 | Vitis vinifera | 72.7 | - |
| 15 | A0A438F5Y0 | 5 | Retrovirus-related Pol polyprotein from transposon TNT 1-94 | Vitis vinifera | 10.1 | - |
| 16 | A0A438HFW8 | 5 | UDP-glycosyltransferase 85A8 | Vitis vinifera | 20.5 | - |
| 17 | A0A438HSQ5 | 6 | Rust resistance kinase Lr10 | Vitis vinifera | 68.4 | - |
| 18 | I9WYJ6 | 6 | 6-carboxy-5,6,7,8-tetrahydropterin synthase | Methylobacterium sp. | 13.5 | - |
| 19 | A0A438JNK9 | 7 | WAT1-related protein | Vitis vinifera | 40.3 | - |
| 20 | A6ZLG3 | 7 | Tyrosine-DNA phosphodiesterase | Saccharomyces cerevisiae | 62.2 | - |
| 21 | A6ZMC5 | 7 | Conserved protein | Saccharomyces cerevisiae | 104.7 | - |
| 22 | A0A438C3D6 | 8 | LysM domain-containing GPI-anchored protein 1 | Vitis vinifera | 43.7 | - |
| 23 | A0A0M3M4Y7 | 8 and 9 | Pectin lyase A | Aspergillus niger | 39.7 | 5 |
| 24 | O24531 | 8 and 11 | Class IV endochitinase (fragment) | Vitis vinifera | 27 | 1,2,3,4,6 |
| 25 | A6ZQF9 | 9 | Killer toxin resistant protein | Saccharomyces cerevisiae | 30 | - |
| 26 | A0A1X7QY33 | 9 | Similar to Saccharomyces cerevisiae YHR098C SFB3 component of the Sec23p-Sfb3p heterodimer of the COPII vesicle coat | Kazachstania saulgeensis | 106.6 | - |
| 27 | A0A1X7R1P0 | 9 | Similar to Saccharomyces cerevisiae YJL170C ASG7 protein that regulates signaling from a G protein β-subunit Ste4p | Kazachstania saulgeensis | 25.7 | - |
| 28 | A0A438F8T9 | 10 | Ethylene-overproduction protein 1 | Vitis vinifera | 113.4 | - |
| 29 | A0A1V2LQA7 | 10 and 11 | Nuclear GTP-binding protein NUG1 | Pichia kudriavzevii | 58.7 | - |
| 30 | A0A438F497 | 11 | Protein HUA2-like 3 | Vitis vinifera | 187.4 | - |
| 31 | H0GDF3 | 11 | Non-specific serine/threonine protein kinase | S. cerevisiae x S. kudriavzevii | 120 | - |
| 32 | A6ZWD3 | 12 | ATP-dependent RNA helicase DBP1 | Saccharomyces cerevisiae | 67.9 | - |
| 33 | A0A438FBU5 | 12 | Cytochrome P450 81E8 | Vitis vinifera | 16.9 | - |
| 34 | A3QRB5 | 12, 13 and 14 | Thaumatin-like protein | Vitis vinifera | 23.9 | 1,2,3,4,5 |
| 35 | Q75E94 | 13 | AAR186Wp | Ashbya gossypii | 25.8 | - |
| 36 | H0GH06 | 13 | Yor1p | S. cerevisiae x S. kudriavzevii | 166.7 | - |
| 37 | H0GRW5 | 13 | Mak32p | S. cerevisiae x S. kudriavzevii | 36.3 | - |
| 38 | A0A438CAI5 | 13 | Retrovirus-related Pol polyprotein from transposon RE1 | Vitis vinifera | 73.7 | - |
| 39 | A0A438F753 | 13 | 5′-nucleotidase SurE | Vitis vinifera | 39.7 | - |
| 40 | A0A438KCF4 | 13 | α-Crystallin domain-containing protein 22.3 | Vitis vinifera | 18.1 | - |
| 41 | A0A438KHH5 | 13 | RNA exonuclease 4 | Vitis vinifera | 44.2 | - |
| 42 | A0A0J9X743 | 13 | Similar to Saccharomyces cerevisiae YGL131C SNT2 DNA binding protein with similarity to the S. pombe Snt2 protein | Geotrichum candidum | 153.2 | - |
| 43 | I9C4L4 | 13 | Protein ImuA | Novosphingobium sp. | 29.1 | - |
| 44 | A0A438FPT4 | 13 | Retrovirus-related Pol polyprotein from transposon 17.6 | Vitis vinifera | 98.5 | - |
| 45 | A0A1V2L627 | 15 | Sensitive to high expression protein 9, mitochondrial | Cyberlindnera fabianii | 42.6 | - |
| 46 | H0GZX2 | 15 | Prm1p | S. cerevisiae x S. kudriavzevii | 73.2 | - |
| 47 | A0A438IBY2 | 15 | Retrovirus-related Pol polyprotein from transposon opus | Vitis vinifera | 144.6 | - |
| 48 | A0A438IP20 | 15 | Putative ribonuclease H protein | Vitis vinifera | 16.6 | - |
| IN-SOLUTION (exclusively identified by the in-solution digestion method) | ||||||
| Accession | SEC Fraction | Description | Organism | MW (kDa) | Reported by Ref * | |
| 49 | A6ZL40 | A | Acid phosphatase | Saccharomyces cerevisiae | 52.9 | 1 |
| 50 | B3LP15 | A | Protein YGP1 | Saccharomyces cerevisiae | 37.3 | 5 |
| 51 | A6ZM69 | A | Lysophospholipase | Saccharomyces cerevisiae | 71.6 | - |
| 52 | F8KAD2 | A | Exo-(1,3)-β-glucanase of the cell wall | Saccharomyces uvarum | 51.2 | 1 |
| 53 | A6ZQA6 | A | Cell wall mannoprotein | Saccharomyces cerevisiae | 29.6 | - |
| 54 | A0A438EWP8 | A | Plasma membrane ATPase | Vitis vinifera | 105.8 | - |
| 55 | H0GZ48 | A | Lysophospholipase | S. cerevisiae x S. kudriavzevii | 75.4 | - |
| 56 | A0A438F6R5 | A | Pentatricopeptide repeat-containing protein | Vitis vinifera | 104.7 | - |
| 57 | A0A438JSE9 | A | Ubiquitin-60S ribosomal protein L40 | Vitis vinifera | 80.1 | - |
| 58 | C7GRZ8 | A | YJL171C-like protein | Saccharomyces cerevisiae | 42.9 | - |
| 59 | C8Z9T5 | A | Sps100p | Saccharomyces cerevisiae | 34.2 | - |
| 60 | H0GRF2 | A | Tos1p | S. cerevisiae x S. kudriavzevii | 48.2 | 4 |
| 61 | G2WLU7 | A | K7_Ygp1p | Saccharomyces cerevisiae | 37.3 | 5 |
| 62 | H0GVA1 | A | Glycosidase | S. cerevisiae x S. kudriavzevii | 54.8 | 4,5 |
| 63 | A0A438CXL6 | A | Transposon Ty3-I Gag-Pol polyprotein | Vitis vinifera | 59.1 | - |
| 64 | Q753A2 | A | AFR422Wp | Ashbya gossypii | 39.2 | - |
| 65 | Q758V6 | A | AEL320Wp | Ashbya gossypii | 112.9 | - |
| 66 | A5ANX3 | A and B | Cysteine proteinase inhibitor | Vitis vinifera | 11.2 | - |
| 67 | A0A438HVZ7 | A and C | Endochitinase EP3 | Vitis vinifera | 28.6 | 1,2,3,4,6 |
| 68 | A6ZVW2 | A, C and D | Seripauperin | Saccharomyces cerevisiae | 17.7 | 5 |
| 69 | A0A438DZR8 | A, C and D | Non-specific lipid-transfer protein | Vitis vinifera | 11.7 | 3,4,6 |
| 70 | A7A1R6 | B | Cell wall mannoprotein | Saccharomyces cerevisiae | 23.3 | - |
| 71 | G2WE85 | B | Plasma membrane ATPase | Saccharomyces cerevisiae | 99.6 | - |
| 72 | Q9P963 | B | ACC synthase | Penicillium citrinum | 48.2 | - |
| 73 | A0A438J3Y1 | B | Retrovirus-related Pol polyprotein from transposon TNT 1-94 | Vitis vinifera | 135.4 | - |
| 74 | H0GGT5 | B | Glycosidase | S. cerevisiae x S. kudriavzevii | 53.7 | 1,4,5 |
| 75 | C8ZED9 | B | Sma2p | Saccharomyces cerevisiae | 40.8 | - |
| 76 | A6ZLA4 | B and C | Target of Sbf | Saccharomyces cerevisiae | 47.9 | 1 |
| 77 | H0GYP4 | C | Ccw14p | S. cerevisiae x S. kudriavzevii | 25 | - |
| 78 | A6ZPT3 | C | GTPase-activating protein | Saccharomyces cerevisiae | 53.9 | - |
| 79 | A6ZVC9 | C | Histidine kinase osmosensor that regulates an osmosensing MAP kinase cascade | Saccharomyces cerevisiae | 134.5 | - |
| 80 | H0GWM4 | C | Cis3p | S. cerevisiae x S. kudriavzevii | 23.3 | - |
| 81 | H0GL37 | C | Asi1p | S. cerevisiae x S. kudriavzevii | 71.4 | - |
| 82 | A0A438DEP9 | C | Retrovirus-related Pol polyprotein from transposon TNT 1-94 | Vitis vinifera | 169.1 | - |
| 83 | G2WJP1 | C | K7_Sen1p | Saccharomyces cerevisiae | 252.5 | - |
| 84 | Q2QCI7 | D | Non-specific lipid-transfer protein | Vitis vinifera | 11.8 | 3,4,6 |
| 85 | I9WWM7 | D | PAS domain-containing protein | Methylobacterium sp. | 21.3 | - |
| 86 | Q752D0 | D | AFR645Wp | Ashbya gossypii | 44.7 | - |
| IN-GEL/IN-SOLUTION (identified by in-gel and in-solution digestion) | ||||||
| Accession | Gel Band/SEC fraction | Description | Organism | MW (kDa) | Reported by Ref * | |
| 87 | A6ZSE1 | 1/A | Daughter-specific expression-related protein | Saccharomyces cerevisiae | 121.1 | 1 |
| 88 | C7GQJ1 | 1 and 2/A, B | Cell wall protein ECM33 | Saccharomyces cerevisiae | 43.8 | 1 |
| 89 | A0A438I656 | 1, 2, 4, 5, 6, 8, 9 and 10/A, B, C | Glucan endo-(1,3)-β-glucosidase | Vitis vinifera | 36.8 | - |
| 90 | Q9S944 | 1, 3 and 8/D | Vacuolar invertase 1 | Vitis vinifera | 71.5 | 1,2,3,4,6 |
| 91 | Q7XAU6 | 1, 4, 5, 6, 8, 9, 10, 11, 12 and 13/A, B, C, D | Class IV chitinase | Vitis vinifera | 27.5 | 2,3,4,6 |
| 92 | A6ZVQ6 | 2/A, B | Cell wall mannoprotein | Saccharomyces cerevisiae | 26.6 | - |
| 93 | A0A438I659 | 1, 2, 4, 5, 6, 8, 9 and 10/A, B, C | Glucan endo-(1,3)-β-glucosidase | Vitis vinifera | 23.9 | - |
| 94 | A0A438DX78 | 4 and 5/A, B | β-Fructofuranosidase, soluble isoenzyme I | Vitis vinifera | 23.9 | - |
| 95 | A0A438JJ75 | 4, 5, 6, 8, 9, 10, 11, 12, 14 and 16/A, B, C, D | Thaumatin-like protein | Vitis vinifera | 23.9 | 1,2,3,4,5,6 |
| 96 | A0A438BZP1 | 6, 8, 9, 10, 11, 12, 13, 14 and 15/B, C, D | Thaumatin-like protein | Vitis vinifera | 36.8 | 1,2,3,4,5,6 |
| 97 | Q756G2 | 8, 9 and 14/C | Probable E3 ubiquitin-protein ligase TOM1 | Ashbya gossypii | 372.2 | - |
| 98 | A0A438JJ53 | 8, 9, 12, 13 and 14/C, D | Thaumatin-like protein | Vitis vinifera | 23.9 | 1,2,3,4,5,6 |
| 99 | F8KAD7 | 9/ B | Endo-(1,3)-β-glucanase | Vitis vinifera | 34 | 1,2,6 |
| 100 | F8KAD8 | 10 and 11/C | Endo-(1,3)-β-glucanase | Vitis vinifera | 63.5 | 1,2,6 |
| 101 | A0A438GZ57 | 16/ D | Putative non-specific lipid-transfer protein AKCS9 | Vitis vinifera | 9.8 | 3,4,6 |
| 102 | Q850K5 | 16/C, D | Non-specific lipid-transfer protein | Vitis vinifera | 11.7 | 3,4,6 |
| IN-GEL (Exclusively Identified by in-gel Digestion) | |||||
|---|---|---|---|---|---|
| Accession | Gel Band | Description | Organism | MW (kDa) | |
| 1 | A0A438J4X9 | 1 | Uncharacterized protein | Vitis vinifera | 67.3 |
| 2 | F6HUG6 | 1, 4 and 5 | Uncharacterized protein | Vitis vinifera | 25.3 |
| 3 | A0A438HSP1 | 2 and 9 | Uncharacterized protein | Vitis vinifera | 32.6 |
| 4 | A0A438J6G3 | 2 | Uncharacterized protein | Vitis vinifera | 77.5 |
| 5 | A5AP16 | 2 | Uncharacterized protein | Vitis vinifera | 61.5 |
| 6 | A0A438HTJ6 | 3 | Uncharacterized protein | Vitis vinifera | 26.6 |
| 7 | A5B108 | 3 | Uncharacterized protein | Vitis vinifera | 101.2 |
| 8 | A5BPD3 | 3 | Uncharacterized protein | Vitis vinifera | 93.1 |
| 9 | A5BUH4 | 3 and 6 | Uncharacterized protein | Vitis vinifera | 73.7 |
| 10 | D7SRI7 | 3 | Uncharacterized protein | Vitis vinifera | 44.4 |
| 11 | A5BGP0 | 4 | Uncharacterized protein | Vitis vinifera | 42.1 |
| 12 | A5BD73 | 4 | Uncharacterized protein | Vitis vinifera | 73.2 |
| 13 | A5BWA5 | 4 | Uncharacterized protein | Vitis vinifera | 28.7 |
| 14 | A5AD63 | 4, 9 and 13 | Uncharacterized protein | Vitis vinifera | 71.8 |
| 15 | F6GZ16 | 5 | Uncharacterized protein | Vitis vinifera | 98.2 |
| 16 | A0A438IVS9 | 7 | Uncharacterized protein | Vitis vinifera | 88.7 |
| 17 | A5AYX1 | 7 | Uncharacterized protein | Vitis vinifera | 73.9 |
| 18 | A5B6K0 | 9 | Uncharacterized protein | Vitis vinifera | 91.9 |
| 19 | A5BKS0 | 9 | Uncharacterized protein | Vitis vinifera | 71.5 |
| 20 | A5BW59 | 9 | Uncharacterized protein | Vitis vinifera | 91.8 |
| 21 | A5BX40 | 9 | Uncharacterized protein | Vitis vinifera | 147.5 |
| 22 | A0A1V2L6J1 | 9 | Uncharacterized protein | Cyberlindnera fabianii | 105.9 |
| 23 | A0A438JPS2 | 9 | Uncharacterized protein | Vitis vinifera | 76.1 |
| 24 | A5BRN8 | 9 | Uncharacterized protein | Vitis vinifera | 38.3 |
| 25 | D7SL13 | 9 | Uncharacterized protein | Vitis vinifera | 6.4 |
| 26 | A5AVZ0 | 9 | Uncharacterized protein | Vitis vinifera | 168.4 |
| 27 | A5BVR4 | 10 | Uncharacterized protein | Vitis vinifera | 38.6 |
| 28 | F6HAW3 | 11 | Uncharacterized protein | Vitis vinifera | 32 |
| 29 | A5B6N1 | 11 | Uncharacterized protein | Vitis vinifera | 54.9 |
| 30 | D7SVF8 | 12 | Uncharacterized protein | Vitis vinifera | 16.8 |
| 31 | A0A438I1U6 | 13 | Uncharacterized protein | Vitis vinifera | 10.8 |
| 32 | F6I094 | 13 | Uncharacterized protein | Vitis vinifera | 58.7 |
| 33 | A5AK33 | 14 | Uncharacterized protein | Vitis vinifera | 36.1 |
| 34 | A5B9R1 | 14 | Uncharacterized protein | Vitis vinifera | 248.6 |
| 35 | A5B1A9 | 15 | Uncharacterized protein | Vitis vinifera | 69.3 |
| 36 | A0A438JBK9 | 15 | Uncharacterized protein | Vitis vinifera | 24.9 |
| 37 | A5BEX7 | 15 | Uncharacterized protein | Vitis vinifera | 118.5 |
| 38 | A5BUI9 | 15 | Uncharacterized protein | Vitis vinifera | 40.2 |
| 39 | A5CAU1 | 15 | Uncharacterized protein | Vitis vinifera | 84.7 |
| 40 | A5AT89 | 16 | Uncharacterized protein | Vitis vinifera | 65.6 |
| IN-SOLUTION (exclusively identified by in-solution digestion) | |||||
| Accession | SEC Fraction | Description | Organism | MW (kDa) | |
| 41 | F6H9W6 | A | Uncharacterized protein | Vitis vinifera | 133.1 |
| 42 | A5BP85 | B | Uncharacterized protein | Vitis vinifera | 113.1 |
| 43 | A5BY31 | C | Uncharacterized protein | Vitis vinifera | 125.3 |
| 44 | D7TT81 | C | Uncharacterized protein | Vitis vinifera | 47 |
| 45 | F6H4B3 | C | Uncharacterized protein | Vitis vinifera | 58.1 |
| 46 | A5BYL8 | D | Uncharacterized protein | Vitis vinifera | 103.5 |
| 47 | A0A438FVB3 | D | Uncharacterized protein | Vitis vinifera | 22.2 |
| IN-GEL/IN-SOLUTION (identified by in-gel and in-solution digestion) | |||||
| Accession | Gel Band/SEC fraction | Description | Organism | MW (kDa) | |
| 48 | F6HMA2 | 1/ A, B | Uncharacterized protein | Vitis vinifera | 60.7 |
| 49 | F6HAU0 | 4, 5, 6, 9, 10, 11 and 12/A, B, C | Uncharacterized protein | Vitis vinifera | 60 |
| 50 | F6HUH1 | 4, 5, 6, 8, 9, 10, 11, 12, 13 and 14 /B, C, D | Uncharacterized protein | Vitis vinifera | 24 |
| 51 | A5C9F1 | 10, 11 and 16/A, B | Uncharacterized protein | Vitis vinifera | 23.8 |
| 52 | D7TXF5 | 10, 11 and 16/D | Uncharacterized protein | Vitis vinifera | 15.1 |
| Wine | Protein Extraction | Protein Separation | MS Analysis | Digestion Method | Identified Proteins (n) | % of Grape + Yeast Proteins | Reference |
|---|---|---|---|---|---|---|---|
| Sauvignon blanc | Cellulose acetate membrane (MWCO—5 kDa) Precipitation [(NH4)2SO4] | SDS-PAGE | Nano-LC-MS Ion trap MS | In-gel | Total: 20 5 (grape) 12 (yeast) 1 (fungi) 2 (bacteria) | 85% | Kwon [30] |
| Chardonnay | Polysulfone membrane (MWCO—10 kDa) Precipitation (85%—C2H6O + 15% C2HCl3O2) | Isoelectric Focusing (IEF) SDS-PAGE | Nano-LC-MS/MS Ion trap MS | In-gel | Total: 13 10 (grape) 1 (yeast) 2 (fungi) | 84.6% | Cilindre et al. [22] |
| Semillon | Precipitation [(NH4)2SO4] | Hydrophobic interaction chromatography (HIC) Reversed phase HPLC SDS-PAGE | Nano-LC-MS/MS TOF-MS | In-gel In-solution | Total: 10 10 (grape) | 100% | Marangon et al. [38] |
| German Portugieser | Cellulose membrane (MWCO—3.5 kDa) | SDS- PAGE | LC-MS TOF-MS | In-gel | Total: 18 12 (grape) 6 (yeast) | 100% | Wigand et al. [15] |
| Valpolicella | Protein adsorption (ProteoMiner beads) Protein desorption (Laemmli buffer) | SDS-PAGE | LC-MS TOF-MS | In-gel | Total: 23 1 (grape) 4 (yeast) 13 (fungi) 2 (bacteria) 3 (bovine) | 17.3% | D’Amato et al. [39] |
| Recioto | Protein adsorption (ProteoMiner beads) | SDS-PAGE | Nano-LC-MS/MS | In-gel | Total: 106 95 (grape) 11 (yeast) | 100% | D’Amato et al. [12] |
| Silvaner | Ultrafiltration Cellulose membrane (MWCO—10 kDa) | Size exclusion chromatography (SEC) SDS-PAGE | LC-MS Quadrupole Orbitrap | In-gel In-solution | Total: 154 91 (grape) 47 (yeast) 12 (fungi) 4 (bacteria) | 89.6% | Present study |
| n° | Protein Description | Organism | MW (kDa) | Digestion Method |
|---|---|---|---|---|
| Gene expression and nucleotide metabolism | ||||
| 1 | DNA binding protein | Geotrichum candidum | 153.2 | In-gel |
| 2 | 6-carboxy-5,6,7,8-tetrahydropterin synthase | Methylobacterium sp. | 13.5 | In-gel |
| 3 | Nuclear GTP-binding protein NUG1 | Pichia kudriavzevii | 58.7 | In-gel |
| 4 | ATP-dependent RNA helicase DBP1 | Saccharomyces cerevisiae | 67.9 | In-gel |
| 5 | Tyrosine-DNA phosphodiesterase | Saccharomyces cerevisiae | 62.2 | In-gel |
| 6 | Daughter-specific expression-related protein | Saccharomyces cerevisiae | 121.1 | In-gel+In-solution |
| 7 | Putative ribonuclease H protein | Vitis vinifera | 16.6 | In-gel |
| 8 | Retrovirus-related Pol polyprotein from transposon RE1 | Vitis vinifera | 73.7 | In-gel |
| 9 | RNA exonuclease 4 | Vitis vinifera | 44.2 | In-gel |
| 10 | Transposon Ty3-I Gag-Pol polyprotein | Vitis vinifera | 59.1 | In-solution |
| 11 | 5′-nucleotidase SurE | Vitis vinifera | 39.7 | In-gel |
| Metabolic breakdown and formation of carbohydrates | ||||
| 12 | Pectin lyase A | Aspergillus niger | 39.7 | In-gel |
| 13 | Glycosidase | S. cerevisiae x S. kudriavzevii | 53.7 | In-solution |
| 14 | Endo-(1,3)-β-glucanase | Saccharomyces uvarum | 34 | In-gel+In-solution |
| 15 | Exo-(1,3)-β-glucanase of the cell wall | Saccharomyces uvarum | 51.2 | In-gel+In-solution |
| 16 | Glucan endo-(1,3)-β-glucosidase | Vitis vinifera | 36.8 | In-gel+In-solution |
| 17 | UDP-glycosyltransferase 85A8 | Vitis vinifera | 20.5 | In-gel |
| 18 | Vacuolar invertase 1 | Vitis vinifera | 71.5 | In-gel+In-solution |
| 19 | β-fructofuranosidase, soluble isoenzyme I | Vitis vinifera | 63.5 | In-gel+In-solution |
| Proteins involved in post-translational modifications | ||||
| 20 | Aminopeptidase | Novosphingobium sp. | 72 | In-gel |
| 21 | Non-specific serine/threonine protein kinase | S. cerevisiae x S. kudriavzevii | 120 | In-gel |
| 22 | Cysteine proteinase inhibitor | Vitis vinifera | 11.2 | In-solution |
| 23 | IAA-amino acid hydrolase ILR1-like 4 | Vitis vinifera | 72.7 | In-gel |
| 24 | α-Crystallin domain-containing protein 22.3 | Vitis vinifera | 18.1 | In-gel |
| Lipid metabolism | ||||
| 25 | Putative lipase ATG15 | Pichia kudriavzevii | 56.8 | In-gel |
| 26 | Lysophospholipase | Saccharomyces cerevisiae | 71.6 | In-solution |
| 27 | Putative non-specific lipid-transfer protein AKCS9 | Vitis vinifera | 9.8 | In-gel+In-solution |
| 28 | Non-specific lipid-transfer protein | Vitis vinifera | 11.7 | In-gel+In-solution |
| Cell defense | ||||
| 29 | Killer toxin resistant protein | Saccharomyces cerevisiae | 30 | In-gel |
| 30 | Pathogen-related protein | Saccharomyces cerevisiae | 30.6 | In-gel |
| 31 | Class IV endochitinase (Fragment) | Vitis vinifera | 27 | In-gel |
| 32 | Endochitinase EP3 | Vitis vinifera | 27.2 | In-gel |
| 33 | LysM domain-containing GPI-anchored protein 1 | Vitis vinifera | 43.7 | In-gel |
| 34 | Thaumatin-like protein | Vitis vinifera | 23.9 | In-gel+In-solution |
| Cell metabolism and signaling | ||||
| 35 | Probable E3 ubiquitin-protein ligase TOM1 | Ashbya gossypii | 372.2 | In-gel+In-solution |
| 36 | Cytokinesis protein sepH | Cyberlindnera fabianii | 116.3 | In-gel |
| 37 | Protein that regulates signaling from a G protein β subunit Ste4p | Kazachstania saulgeensis | 25.7 | In-gel |
| 38 | ACC synthase | Penicillium citrinum | 48.2 | In-solution |
| 39 | PAS domain-containing protein | Methylobacterium sp. | 21.3 | In-solution |
| 40 | Acid phosphatase | Saccharomyces cerevisiae | 52.9 | In-solution |
| 39 | GTPase-activating protein | Saccharomyces cerevisiae | 53.9 | In-solution |
| 40 | Histidine kinase osmosensor that regulates an osmosensing MAP kinase cascade | Saccharomyces cerevisiae | 134.5 | In-solution |
| 41 | Cytochrome P450 81E8 | Vitis vinifera | 16.9 | In-gel |
| 42 | Ethylene-overproduction protein 1 | Vitis vinifera | 113.4 | In-gel |
| 43 | Pentatricopeptide repeat-containing protein | Vitis vinifera | 104.7 | In-solution |
| 44 | Plasma membrane ATPase | Vitis vinifera | 105.8 | In-solution |
| 45 | Rust resistance kinase Lr10 | Vitis vinifera | 68.4 | In-gel |
| 46 | Ubiquitin-60S ribosomal protein L40 | Vitis vinifera | 80.1 | In-solution |
| Cell structural elements | ||||
| 47 | Sensitive to high expression protein 9, mitochondrial | Cyberlindnera fabianii | 42.6 | In-gel |
| 48 | Component of the Sec23p-Sfb3p heterodimer of the COPII vesicle coat | Kazachstania saulgeensis | 106.6 | In-gel |
| 49 | Cell wall mannoprotein | Saccharomyces cerevisiae | 29.6 | In-gel+In-solution |
| 50 | Cell wall protein ECM33 | Saccharomyces cerevisiae | 43.8 | In-gel+In-solution |
| 51 | Seripauperin | Saccharomyces cerevisiae | 17.7 | In-solution |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albuquerque, W.; Ghezellou, P.; Seidel, L.; Burkert, J.; Will, F.; Schweiggert, R.; Spengler, B.; Zorn, H.; Gand, M. Mass Spectrometry-Based Proteomic Profiling of a Silvaner White Wine. Biomolecules 2023, 13, 650. https://doi.org/10.3390/biom13040650
Albuquerque W, Ghezellou P, Seidel L, Burkert J, Will F, Schweiggert R, Spengler B, Zorn H, Gand M. Mass Spectrometry-Based Proteomic Profiling of a Silvaner White Wine. Biomolecules. 2023; 13(4):650. https://doi.org/10.3390/biom13040650
Chicago/Turabian StyleAlbuquerque, Wendell, Parviz Ghezellou, Leif Seidel, Johannes Burkert, Frank Will, Ralf Schweiggert, Bernhard Spengler, Holger Zorn, and Martin Gand. 2023. "Mass Spectrometry-Based Proteomic Profiling of a Silvaner White Wine" Biomolecules 13, no. 4: 650. https://doi.org/10.3390/biom13040650
APA StyleAlbuquerque, W., Ghezellou, P., Seidel, L., Burkert, J., Will, F., Schweiggert, R., Spengler, B., Zorn, H., & Gand, M. (2023). Mass Spectrometry-Based Proteomic Profiling of a Silvaner White Wine. Biomolecules, 13(4), 650. https://doi.org/10.3390/biom13040650