
Melatonin in Health and Disease: A Perspective for Livestock Production


Abstract
1. Introduction, Photoperiod, and Synthesis Pathway

{kind=link}
| Specie | Sample | Daytime (pg/mL) | Nighttime (pg/mL) | Method | Reference |
|---|---|---|---|---|---|
| Human (Homo sapiens) | Plasma | 1.5–4.9 | 19.4–42.6 | GC-MS | [32] |
| Swine (Sus domesticus) | Plasma | 30–40 | 35–65 | RIA | [33] |
| Swine (Sus domesticus) | Serum | 42.9 | 38.4 | RIA | [34] |
| Chicken (Gallus domesticus) | Plasma | 53 | 201 | RIA | [35] |
| Chicken (Gallus domesticus) | Plasma | 67.4–117.9 | 325–379 | RIA | [36] |
| Sheep (Ovis aries) | Serum | 10–30 | 100–300 | RIA | [37] |
| Sheep (Ovis aries) | Serum | 15.7–36.9 | 60.7–239.2 | ELISA | [38] |
| Horse (Equus caballus) | Plasma | 0–10 | 10–45 | RIA | [39] |
| Horse (Cycling mare in March) | Serum | 11.6 | 13.2 | RIA | [40] |
| Horse (Noncycling mare in March) | Serum | 8.9 | 8.2 | RIA | [40] |
| Cow (Bos taurus) | Plasma | 5 | 90 | RIA | [41] |
| Cow (Fall) | Plasma | 13.9 | 27.8 | ELISA | [42] |
| Cow (Summer) | Plasma | 12.8 | 24.73 | ELISA | [42] |
2. Circadian Rhythms, Environmental Factors, and Body Temperature
3. Immune System
3.1. Innate Immune Response
3.2. Adaptive Immune Response
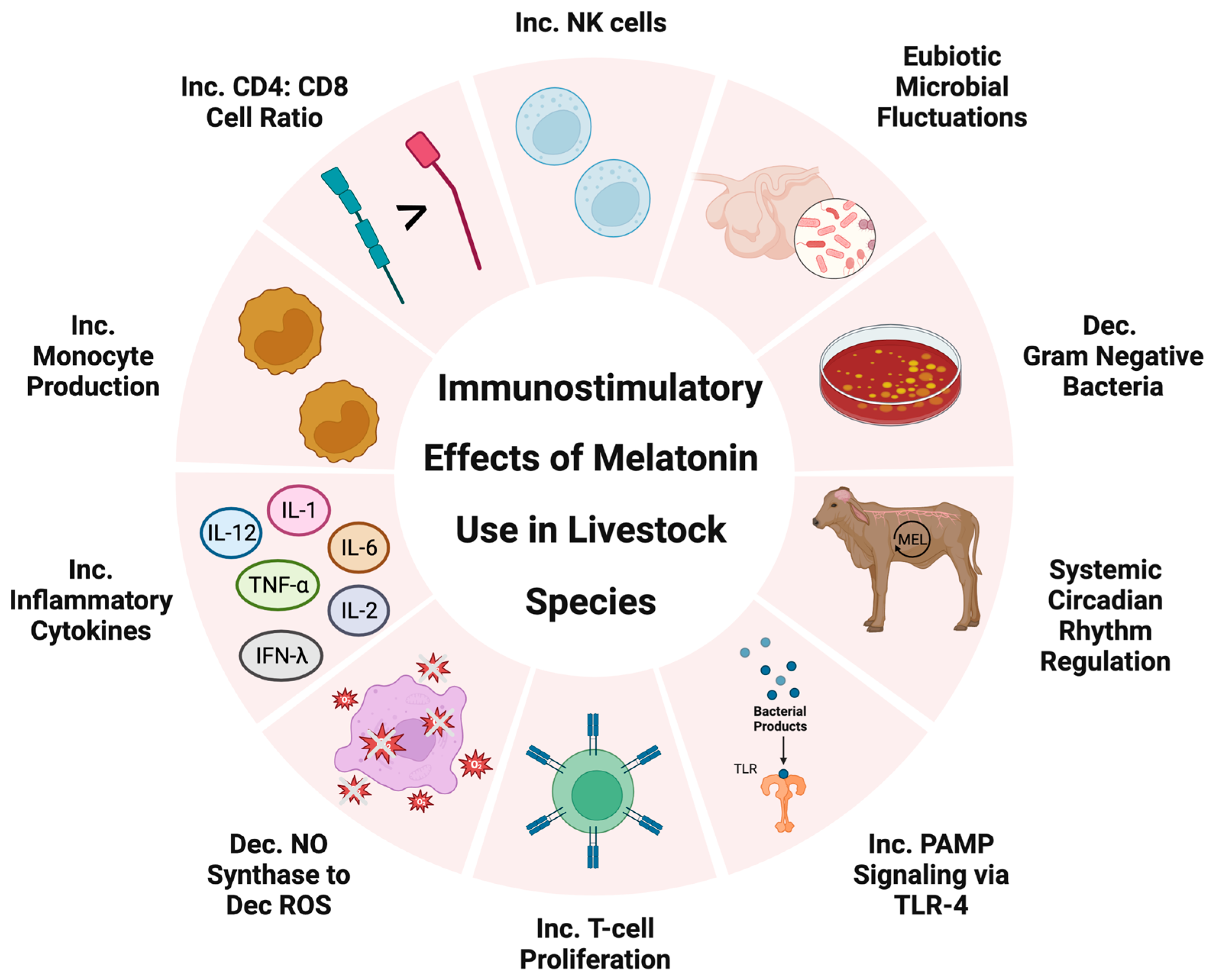
3.3. Immunostimulatory Melatonin in Livestock Production
4. Microbiome
4.1. Gut Microbiome
4.2. Reproductive Tract Microbiome
4.3. Melatonin in Livestock Microbiomes
5. Skeletal Muscle and Growth and Development
6. Amino Acids in Livestock Maternal Blood
7. Endocrine and Receptor Pathways
8. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lerner, A.B.; Case, J.D.; Takahashi, Y.; Lee, T.H.; Mori, W. Isolation of melatonin, the pineal gland factor that lightens melanocytes1. J. Am. Chem. Soc. 1958, 80, 2587. [Google Scholar] [CrossRef]
- Reiter, R.J.; Hester, R.J. Interrelationships of the Pineal Gland, the Superior Cervical Ganglia and the Photoperiod in the Regulation of the Endocrine Systems of Hamsters. Endocrinology 1966, 79, 1168–1170. [Google Scholar] [CrossRef]
- Chemineau, P.; Normant, E.; Ravault, J.P.; Thimonier, J. Induction and persistence of pituitary and ovarian activity in the out-of-season lactating dairy goat after a treatment combining a skeleton photoperiod, melatonin and the male effect. J. Reprod. Fertil. 1986, 78, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Ronayne, E.; Jordan, B.; Quirke, J.; Roche, J. The effect of frequency of administration of melatonin on the time of onset of the breeding season in anoestrous ewes. Anim. Reprod. Sci. 1989, 18, 13–24. [Google Scholar] [CrossRef]
- Malpaux, B.; Viguié, C.; Skinner, D.; Thiéry, J.; Pelletier, J.; Chemineau, P. Seasonal breeding in sheep: Mechanism of action of melatonin. Anim. Reprod. Sci. 1996, 42, 109–117. [Google Scholar] [CrossRef]
- Chemineau, P.; Malpaux, B.; Delgadillo, J.; Guérin, Y.; Ravault, J.; Thimonier, J.; Pelletier, J. Control of sheep and goat reproduction: Use of light and melatonin. Anim. Reprod. Sci. 1992, 30, 157–184. [Google Scholar] [CrossRef]
- Haresign, W.; Peters, A.R.; Staples, L.D. The effect of melatonin implants on breeding activity and litter size in commercial sheep flocks in the UK. Anim. Sci. 1990, 50, 111–121. [Google Scholar] [CrossRef]
- Guillaume, D.; Palmer, E. Effect of oral melatonin on the date of the first ovulation after ovarian inactivity in mares under artificial photoperiod. J. Reprod. Fertil. Suppl. 1991, 44, 249–257. [Google Scholar]
- Walsh, C.; Prendergast, R.; Sheridan, J.; Murphy, B. Blue light from light-emitting diodes directed at a single eye elicits a dose-dependent suppression of melatonin in horses. Veter. J. 2013, 196, 231–235. [Google Scholar] [CrossRef]
- Ganguly, S.; Coon, S.L.; Klein, D.C. Control of melatonin synthesis in the mammalian pineal gland: The critical role of serotonin acetylation. Cell Tissue Res. 2002, 309, 127–137. [Google Scholar] [CrossRef]
- Arendt, J. Melatonin and the pineal gland: Influence on mammalian seasonal and circadian physiology. Rev. Reprod. 1998, 3, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.-X.; Manchester, L.C.; Paredes, S.D.; Mayo, J.C.; Sainz, R.M. Melatonin and Reproduction Revisited. Biol. Reprod. 2009, 81, 445–456. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Sharma, R. Central and peripheral actions of melatonin on reproduction in seasonal and continuous breeding mammals. Gen. Comp. Endocrinol. 2021, 300, 113620. [Google Scholar] [CrossRef] [PubMed]
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Lovenberg, W.; Jequier, E.; Sjoerdsma, A. Tryptophan Hydroxylation: Measurement in Pineal Gland, Brainstem, and Carcinoid Tumor. Science 1967, 155, 217–219. [Google Scholar] [CrossRef]
- Klein, D.C.; Weller, J.L. Rapid Light-Induced Decrease in Pineal Serotonin N-Acetyltransferase Activity. Science 1972, 177, 532–533. [Google Scholar] [CrossRef]
- Wurtman, R.J.; Axelrod, J.; Phillips, L.S. Melatonin Synthesis in the Pineal Gland: Control by Light. Science 1963, 142, 1071–1073. [Google Scholar] [CrossRef] [PubMed]
- Brzezinski, A.; Seibel, M.M.; Lynch, H.J.; Deng, M.-H.; Wurtman, R.J. Melatonin in Human Preovulatory Follicular Fluid. J. Clin. Endocrinol. Metab. 1987, 64, 865–867. [Google Scholar] [CrossRef]
- Bubenik, G.; Ayles, H.; Friendship, R.; Brown, G.; Ball, R. Relationship between melatonin levels in plasma and gastrointestinal tissues and the incidence and severity of gastric ulcers in pigs. J. Pineal Res. 1998, 24, 62–66. [Google Scholar] [CrossRef]
- Chong, N.W.; Chaurasia, S.S.; Haque, R.; Klein, D.C.; Iuvone, P.M. Temporal-spatial characterization of chicken clock genes: Circadian expression in retina, pineal gland, and peripheral tissues. J. Neurochem. 2003, 85, 851–860. [Google Scholar] [CrossRef]
- Reiter, R.J.; Mayo, J.C.; Tan, D.-X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Manchester, L.C.; Liu, X.; Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Reiter, R.J. Mitochondria and chloroplasts as the original sites of melatonin synthesis: A hypothesis related to melatonin’s primary function and evolution in eukaryotes. J. Pineal Res. 2013, 54, 127–138. [Google Scholar] [CrossRef]
- Reiter, R.J. Melatonin: Clinical relevance. Best Pract. Res. Clin. Endocrinol. Metab. 2003, 17, 273–285. [Google Scholar] [CrossRef] [PubMed]
- Paulis, L.; Šimko, F. Blood pressure modulation and cardiovascular protection by melatonin: Potential mechanisms behind. Physiol. Res. 2007, 56, 671–684. [Google Scholar] [CrossRef]
- Pogan, L.; Bissonnette, P.; Parent, L.; Sauvé, R. The effects of melatonin on Ca2+ homeostasis in endothelial cells. J. Pineal Res. 2002, 33, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Garg, U.C.; Hassid, A. Nitric oxide-generating vasodilators and 8-bromo-cyclic guanosine monophosphate inhibit mitogenesis and proliferation of cultured rat vascular smooth muscle cells. J. Clin. Investig. 1989, 83, 1774–1777. [Google Scholar] [CrossRef]
- Mahle, C.D.; Goggins, G.D.; Agarwal, P.; Ryan, E.; Watson, A.J. Melatonin modulates vascular smooth muscle tone. J. Biol. Rhythm. 1997, 12, 690–696. [Google Scholar] [CrossRef]
- Shibata, S.; Satake, N.; Takagi, T.; Usui, H. Vasorelaxing action of melatonin in rabbit basilar artery. Gen. Pharmacol. Vasc. Syst. 1989, 20, 677–680. [Google Scholar] [CrossRef]
- Viswanathan, M.; Laitinen, J.T.; Saavedra, J.M. Expression of melatonin receptors in arteries involved in thermoregulation. Proc. Natl. Acad. Sci. USA 1990, 87, 6200–6203. [Google Scholar] [CrossRef]
- Weekley, L.B. Effects of melatonin on pulmonary and coronary vessels are exerted through perivascular nerves. Clin. Auton. Res. 1993, 3, 45–47. [Google Scholar] [CrossRef]
- Shukla, P.; Sun, C.; O’Rourke, S.T. Melatonin inhibits nitric oxide signaling by increasing PDE5 phosphorylation in coronary arteries. Am. J. Physiol. Circ. Physiol. 2012, 303, H1418–H1425. [Google Scholar] [CrossRef] [PubMed]
- Lewy, A.J.; Markey, S.P. Analysis of Melatonin in Human Plasma by Gas Chromatography Negative Chemical Ionization Mass Spectrometry. Science 1978, 201, 741–743. [Google Scholar] [CrossRef] [PubMed]
- Bubenik, G.A.; Pang, S.F.; Cockshut, J.R.; Smith, P.S.; Grovum, L.W.; Friendship, R.M.; Hacker, R.R. Circadian variation of portal, arterial and venous blood levels of melatonin in pigs and its relationship to food intake and sleep. J. Pineal Res. 2000, 28, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Griffith, M.K.; Minton, J.E. Effect of light intensity on circadian profiles of melatonin, prolactin, ACTH, and cortisol in pigs. J. Anim. Sci. 1992, 70, 492–498. [Google Scholar] [CrossRef]
- Liou, S.S.; Cogburn, L.; Biellier, H.V. Photoperiodic regulation of plasma melatonin levels in the laying chicken (Gallus domesticus). Gen. Comp. Endocrinol. 1987, 67, 221–226. [Google Scholar] [CrossRef]
- Doi, O.; Iwasawa, A.; Nakamura, T.; Tanabe, Y. Effects of Different Photoperiods on Plasma Melatonin Rhythm of the Chicken. Anim. Sci. Technol. 1995, 66, 16–26. [Google Scholar] [CrossRef]
- Rollag, M.; Niswender, G. Radioimmunoassay of Serum Concentrations of Melatonin in Sheep Exposed to Different Lighting Regimens. Endocrinology 1976, 98, 482–489. [Google Scholar] [CrossRef]
- Lemley, C.O.; Meyer, A.M.; Camacho, L.E.; Neville, T.L.; Newman, D.J.; Caton, J.S.; Vonnahme, K.A. Melatonin supplementation alters uteroplacental hemodynamics and fetal development in an ovine model of intrauterine growth restriction. Am. J. Physiol. Integr. Comp. Physiol. 2012, 302, R454–R467. [Google Scholar] [CrossRef]
- Haritou, S.J.A.; Zylstra, R.; Ralli, C.; Turner, S.; Tortonese, D.J. Seasonal Changes in Circadian Peripheral Plasma Concentrations of Melatonin, Serotonin, Dopamine and Cortisol in Aged Horses with Cushing’s Disease under Natural Photoperiod. J. Neuroendocr. 2008, 20, 988–996. [Google Scholar] [CrossRef]
- Diekman, M.A.; Braun, W.; Peter, D.; Cook, D. Seasonal serum concentrations of melatonin in cycling and noncycling mares. J. Anim. Sci. 2002, 80, 2949–2952. [Google Scholar] [CrossRef]
- Berthelot, X.; Laurentie, M.; Ravault, J.; Ferney, J.; Toutain, P. Circadian profile and production rate of melatonin in the cow. Domest. Anim. Endocrinol. 1990, 7, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Correa, E.Z.; Messman, R.D.; Sidelinger, D.R.; King, E.H.; Sánchez-Rodríguez, H.L.; Burnett, D.D.; Lemley, O.C. Melatonin alters bovine uterine artery hemodynamics, vaginal temperatures, and fetal morphometrics during late gestational nutrient restriction in a season-dependent manner. J. Anim. Sci. 2021, 99, skab242. [Google Scholar] [CrossRef] [PubMed]
- Weaver, D. The Suprachiasmatic Nucleus: A 25-Year Retrospective. J. Biol. Rhythm. 1998, 13, 100–112. [Google Scholar] [CrossRef]
- Dolatshad, H.; Cary, A.J.; Davis, F.C. Differential Expression of the Circadian Clock in Maternal and Embryonic Tissues of Mice. PLoS ONE 2010, 5, e9855. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.; Antoch, M.P.; Miller, B.H.; Su, A.I.; Schook, A.B.; Straume, M.; Schultz, P.G.; Kay, S.A.; Takahashi, J.S.; Hogenesch, J.B. Coordinated Transcription of Key Pathways in the Mouse by the Circadian Clock. Cell 2002, 109, 307–320. [Google Scholar] [CrossRef]
- Lowrey, P.L.; Takahashi, J.S. Mammalian Circadian Biology: Elucidating Genome-Wide Levels of Temporal Organization. Annu. Rev. Genom. Hum. Genet. 2004, 5, 407–441. [Google Scholar] [CrossRef]
- Chong, N.W.; Bernard, M.; Klein, D.C. Characterization of the Chicken SerotoninN-Acetyltransferase Gene: Activation via clock gene heterodimer/e box interaction. J. Biol. Chem. 2000, 275, 32991–32998. [Google Scholar] [CrossRef]
- Ibraheem, M.; Galbraith, H.; Scaife, J.; Ewen, S. Growth of secondary hair follicles of the Cashmere goat in vitro and their response to prolactin and melatonin. J. Anat. 1994, 185, 135–142. [Google Scholar]
- Santiago-Moreno, J.; López-Sebastián, A.; del Campo, A.; González-Bulnes, A.; Picazo, R.; Gómez-Brunet, A. Effect of constant-release melatonin implants and prolonged exposure to a long day photoperiod on prolactin secretion and hair growth in mouflon (Ovis gmelini musimon). Domest. Anim. Endocrinol. 2004, 26, 303–314. [Google Scholar] [CrossRef]
- Aiken, G.; Klotz, J.; Looper, M.; Tabler, S.; Schrick, F. Disrupted hair follicle activity in cattle grazing endophyte-infected tall fescue in the summer insulates core body temperatures. Prof. Anim. Sci. 2011, 27, 336–343. [Google Scholar] [CrossRef]
- Kendall, P.; Webster, J. Season and physiological status affects the circadian body temperature rhythm of dairy cows. Livest. Sci. 2009, 125, 155–160. [Google Scholar] [CrossRef]
- Suarez-Trujillo, A.; Hoang, N.; Robinson, L.; McCabe, C.J.; Conklin, D.; Minor, R.C.; Townsend, J.; Plaut, K.; George, U.Z.; Boerman, J.; et al. Effect of circadian system disruption on the concentration and daily oscillations of cortisol, progesterone, melatonin, serotonin, growth hormone, and core body temperature in periparturient dairy cattle. J. Dairy Sci. 2022, 105, 2651–2668. [Google Scholar] [CrossRef]
- Cagnacci, A.; Kräuchi, K.; Wirz-Justice, A.; Volpe, A. Homeostatic versus Circadian Effects of Melatonin on Core Body Temperature in Humans. J. Biol. Rhythm. 1997, 12, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Dawson, D.; Gibbon, S.; Singh, P. The hypothermic effect of melatonin on core body temperature: Is more better? J. Pineal Res. 1996, 20, 192–197. [Google Scholar] [CrossRef]
- Samuel, V.T.; Shulman, G.I. The pathogenesis of insulin resistance: Integrating signaling pathways and substrate flux. J. Clin. Investig. 2016, 126, 12–22. [Google Scholar] [CrossRef]
- West, J.W. Effects of Heat-Stress on Production in Dairy Cattle. J. Dairy Sci. 2003, 86, 2131–2144. [Google Scholar] [CrossRef]
- Matsuzuka, T.; Sakamoto, N.; Ozawa, M.; Ushitani, A.; Hirabayashi, M.; Kanai, Y. Alleviation of maternal hyperthermia-induced early embryonic death by administration of melatonin to mice. J. Pineal Res. 2005, 39, 217–223. [Google Scholar] [CrossRef]
- Porter, J.K.; Stuedemann, J.A.; Thompson, F.N.; Buchanan, B.A.; Tucker, H.A. Melatonin and pineal neurochemicals in steers grazed on endophyte-infected tall fescue: Effects of metoclopramide. J. Anim. Sci. 1993, 71, 1526–1531. [Google Scholar] [CrossRef] [PubMed]
- Mote, R.S.; Hill, N.S.; Uppal, K.; Tran, V.T.; Jones, D.P.; Filipov, N.M. Metabolomics of fescue toxicosis in grazing beef steers. Food Chem. Toxicol. 2017, 105, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Porter, J.K.; Thompson, J.F.N. Effects of fescue toxicosis on reproduction in livestock. J. Anim. Sci. 1992, 70, 1594–1603. [Google Scholar] [CrossRef]
- Esquifino, A.I.; Pandi-Perumal, S.; Cardinali, D.P. Circadian organization of the immune response: A role for melatonin. Clin. Appl. Immunol. Rev. 2004, 4, 423–433. [Google Scholar] [CrossRef]
- Maestroni, G.J. The immunotherapeutic potential of melatonin. Expert Opin. Investig. Drugs 2001, 10, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Pioli, C.; Caroleo, M.; Nistico, G.; Doriac, G. Melatonin increases antigen presentation and amplifies specific and non specific signals for T-cell proliferation. Int. J. Immunopharmacol. 1993, 15, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Del Gobbo, V.; Libri, V.; Villani, N.; Caliō, R.; Nisticō, G. Pinealectomy inhibits interleukin-2 production and natural killer activity in mice. Int. J. Immunopharmacol. 1989, 11, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Castrillón, P.O.; Esquifino, A.I.; Varas, A.; Zapata, A.; Cutrera, R.A.; Cardinali, D.P. Effect of Melatonin Treatment on 24-h Variations in Responses to Mitogens and Lymphocyte Subset Populations in Rat Submaxillary Lymph Nodes. J. Neuroendocr. 2000, 12, 758–765. [Google Scholar] [CrossRef]
- Maestroni, G.J.M. Is hematopoiesis under the influence of neural and neuroendocrine mechanisms? Histol. Histopathol. 1998, 13, 271–274. [Google Scholar] [CrossRef]
- Wichmann, M.W.; Zellweger, R.; DeMaso, C.M.; Ayala, A.; Chaudry, I.H. Melatonin Administration Attenuates Depressed Immune Functions after Trauma-Hemorrhage. J. Surg. Res. 1996, 63, 256–262. [Google Scholar] [CrossRef]
- Maestroni, G.J.; Conti, A.; Pierpaoli, W. Role of the pineal gland in immunity: Circadian synthesis and release of melatonin modulates the antibody response and antagonizes the immunosuppressive effect of corticosterone. J. Neuroimmunol. 1986, 13, 19–30. [Google Scholar] [CrossRef]
- Srinivasan, V.; Maestroni, G.J.M.; Cardinali, D.P.; Esquifino, A.I.; Pandi-Perumal, S.R.; Miller, S.C. Melatonin, immune function and aging. Immun. Ageing 2005, 2, 17. [Google Scholar] [CrossRef]
- IQWiG, Institute for Quality and Efficiency in Health Care. Innate and Adaptive Immune Systems; National Library of Medicine: Bethesda, MD, USA, 2020.
- García-Mauriño, S.; Pozo, D.; Carrillo-Vico, A.; Calvo, J.R.; Guerrero, J.M. Melatonin activates Th1 lymphocytes by increasing IL-12 production. Life Sci. 1999, 65, 2143–2150. [Google Scholar] [CrossRef]
- Currier, N.L.; Sun, L.Z.-Y.; Miller, S.C. Exogenous melatonin: Quantitative enhancement in vivo of cells mediating non-specific immunity. J. Neuroimmunol. 2000, 104, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Barriga, C.; Martín, M.I.; Ortega, E.; Rodriguez, A.B. Physiological Concentrations of Melatonin and Corticosterone in Stress and their Relationship with Phagocytic Activity. J. Neuroendocr. 2002, 14, 691–695. [Google Scholar] [CrossRef] [PubMed]
- Minagar, A.; Shapshak, P.; Fujimura, R.; Ownby, R.; Heyes, M.; Eisdorfer, C. The role of macrophage/microglia and astrocytes in the pathogenesis of three neurologic disorders: HIV-associated dementia, Alzheimer disease, and multiple sclerosis. J. Neurol. Sci. 2002, 202, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Li, W.; Gao, Q.; Wei, T. Effect of melatonin on the generation of nitric oxide in murine macrophages. Eur. J. Pharmacol. 2004, 501, 25–30. [Google Scholar] [CrossRef]
- Garcia-Mauriño, S.; Gonzalez-Haba, M.G.; Calvo, J.R.; Rafii-El-Idrissi, M.; Sanchez-Margalet, V.; Goberna, R.; Guerrero, J.M. Melatonin enhances IL-2, IL-6, and IFN-gamma production by human circulating CD4+ cells: A possible nuclear receptor-mediated mechanism involving T helper type 1 lymphocytes and monocytes. J. Immunol. 1997, 159, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Morrey, K.M.; McLachlan, A.J.; Serkin, C.D.; Bakouche, O. Activation of human monocytes by the pineal hormone melatonin. J. Immunol. 1994, 153, 2671–2680. [Google Scholar] [CrossRef] [PubMed]
- Berger, A. Th1 and Th2 responses: What are they? BMJ 2000, 321, 424. [Google Scholar] [CrossRef]
- Nelson, R.J.; Drazen, D.L. Melatonin Mediates Seasonal Changes in Immune Function. Ann. N. Y. Acad. Sci. 2000, 917, 404–415. [Google Scholar] [CrossRef]
- Nunnari, G.; Nigro, L.L.; Palermo, F.; Leto, D.; Pomerantz, R.J.; Cacopardo, B. Reduction of Serum Melatonin Levels in HIV-1-Infected Individuals’ Parallel Disease Progression: Correlation with Serum Interleukin-12 Levels. Infection 2003, 31, 379–382. [Google Scholar] [CrossRef]
- Regodón, S.; Ramos, A.; Míguez, M.P.; Carrillo-Vico, A.; Rosado, J.A.; Jardín, I. Vaccination prepartum enhances the beneficial effects of melatonin on the immune response and reduces platelet responsiveness in sheep. BMC Veter. Res. 2012, 8, 84. [Google Scholar] [CrossRef]
- Canto, F.; González, E.; Abecia, J.A. Effects of Implanting Exogenous Melatonin 40 Days before Lambing on Milk and Colostrum Quality. Animals 2022, 12, 1257. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ispierto, I.; Abdelfatah, A.; López-Gatius, F. Melatonin Treatment at Dry-off Improves Reproductive Performance Postpartum in High-producing Dairy Cows under Heat Stress Conditions. Reprod. Domest. Anim. 2013, 48, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Abdelnaby, E.A.; El-Maaty, A.M.A. Melatonin and CIDR improved the follicular and luteal haemodynamics, uterine and ovarian arteries vascular perfusion, ovarian hormones and nitric oxide in cyclic cows. Reprod. Domest. Anim. 2021, 56, 498–510. [Google Scholar] [CrossRef]
- Kim, S.W.; Kim, S.; Son, M.; Cheon, J.H.; Park, Y.S. Melatonin controls microbiota in colitis by goblet cell differentiation and antimicrobial peptide production through Toll-like receptor 4 signalling. Sci. Rep. 2020, 10, 2232. [Google Scholar] [CrossRef]
- Meylan, E.; Tschopp, J.; Karin, M. Intracellular pattern recognition receptors in the host response. Nature 2006, 442, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Stebelova, K.; Herichová, I.; Zeman, M. Diabetes induces changes in melatonin concentrations in peripheral tissues of rat. Neuro Endocrinol. Lett. 2007, 28, 159–165. [Google Scholar]
- Bubenik, G.A.; Brown, G.M. Pinealectomy Reduces Melatonin Levels in the Serum but Not in the Gastrointestinal Tract of Rats. Neurosignals 1997, 6, 40–44. [Google Scholar] [CrossRef]
- Leone, V.; Gibbons, S.M.; Martinez, K.; Hutchison, A.L.; Huang, E.Y.; Cham, C.M.; Pierre, J.F.; Heneghan, A.F.; Nadimpalli, A.; Hubert, N.; et al. Effects of Diurnal Variation of Gut Microbes and High-Fat Feeding on Host Circadian Clock Function and Metabolism. Cell Host Microbe 2015, 17, 681–689. [Google Scholar] [CrossRef]
- Tsang, A.; Astiz, M.; Friedrichs, M.; Oster, H. Endocrine regulation of circadian physiology. J. Endocrinol. 2016, 230, R1–R11. [Google Scholar] [CrossRef]
- Gröschl, M. The physiological role of hormones in saliva. Bioessays 2009, 31, 843–852. [Google Scholar] [CrossRef]
- Ouyang, J.; Wang, M.; Bu, D.; Ma, L.; Liu, F.; Xue, C.; Du, C.; Aboragah, A.; Loor, J.J. Ruminal Microbes Exhibit a Robust Circadian Rhythm and Are Sensitive to Melatonin. Front. Nutr. 2021, 8, 760578. [Google Scholar] [CrossRef] [PubMed]
- Tekbas, O.F.; Ogur, R.; Korkmaz, A.; Kilic, A.; Reiter, R.J. Melatonin as an antibiotic: New insights into the actions of this ubiquitous molecule. J. Pineal Res. 2008, 44, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Messman, R.D.; Contreras-Correa, E.Z.; Paz, A.H.; Lemley, O.C. Melatonin-induced changes in the bovine vaginal microbiota during maternal nutrient restriction. J. Anim. Sci. 2021, 99, skab098. [Google Scholar] [CrossRef] [PubMed]
- Iesanu, M.I.; Zahiu, C.D.M.; Dogaru, I.-A.; Chitimus, D.M.; Pircalabioru, G.G.; Voiculescu, S.E.; Isac, S.; Galos, F.; Pavel, B.; O’Mahony, S.M.; et al. Melatonin–Microbiome Two-Sided Interaction in Dysbiosis-Associated Conditions. Antioxidants 2022, 11, 2244. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; You, W.; Shan, T. Myomaker, and Myomixer-Myomerger-Minion modulate the efficiency of skeletal muscle development with melatonin supplementation through Wnt/β-catenin pathway. Exp. Cell Res. 2019, 385, 111705. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Chen, X.; Qian, Y.; Tang, H.; Song, J.; Qu, X.; Yue, B.; Yuan, W. Melatonin-Based and Biomimetic Scaffold as Muscle–ECM Implant for Guiding Myogenic Differentiation of Volumetric Muscle Loss. Adv. Funct. Mater. 2020, 30, 2002378. [Google Scholar] [CrossRef]
- Sayed, R.K.A.; Fernández-Ortiz, M.; Diaz-Casado, M.E.; Rusanova, I.; Rahim, I.; Escames, G.; López, L.C.; Mokhtar, D.M.; Acuña-Castroviejo, D. The Protective Effect of Melatonin Against Age-Associated, Sarcopenia-Dependent Tubular Aggregate Formation, Lactate Depletion, and Mitochondrial Changes. J. Gerontol. Ser. A 2018, 73, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Sartori, C.; Dessen, P.; Mathieu, C.; Monney, A.; Bloch, J.; Nicod, P.; Scherrer, U.; Duplain, H. Melatonin Improves Glucose Homeostasis and Endothelial Vascular Function in High-Fat Diet-Fed Insulin-Resistant Mice. Endocrinology 2009, 150, 5311–5317. [Google Scholar] [CrossRef]
- Zanuto, R.; Siqueira-Filho, M.A.; Caperuto, L.C.; Bacurau, R.F.P.; Hirata, E.; Peliciari-Garcia, R.A.; Amaral, F.G.D.; Marçal, A.C.; Ribeiro, L.M.; Camporez, J.P.G.; et al. Melatonin improves insulin sensitivity independently of weight loss in old obese rats. J. Pineal Res. 2013, 55, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Owino, S.; Contreras-Alcantara, S.; Baba, K.; Tosini, G. Melatonin Signaling Controls the Daily Rhythm in Blood Glucose Levels Independent of Peripheral Clocks. PLoS ONE 2016, 11, e0148214. [Google Scholar] [CrossRef]
- Duan, T.; Wu, Z.; Zhang, H.; Liu, Y.; Li, Y.; Zhang, W. Effects of melatonin implantation on carcass characteristics, meat quality and tissue levels of melatonin and prolactin in Inner Mongolian cashmere goats. J. Anim. Sci. Biotechnol. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Lemley, C.; Camacho, L.; Meyer, A.; Kapphahn, M.; Caton, J.; Vonnahme, K. Dietary melatonin supplementation alters uteroplacental amino acid flux during intrauterine growth restriction in ewes. Animal 2013, 7, 1500–1507. [Google Scholar] [CrossRef] [PubMed]
- Swanson, R.; Contreras-Correa, Z.; Dinh, T.; King, H.; Sidelinger, D.; Burnett, D.; Lemley, C. Melatonin Supplementation Alters Maternal and Fetal Amino Acid Concentrations and Placental Nutrient Transporters in a Nutrient Restriction Bovine Model. Metabolites 2022, 12, 1208. [Google Scholar] [CrossRef] [PubMed]
- Junior, R.P.; Chuffa, L.G.D.A.; Simão, V.A.; Sonehara, N.M.; Chammas, R.; Reiter, R.J.; Zuccari, D.A.P.D.C. Melatonin Regulates the Daily Levels of Plasma Amino Acids, Acylcarnitines, Biogenic Amines, Sphingomyelins, and Hexoses in a Xenograft Model of Triple Negative Breast Cancer. Int. J. Mol. Sci. 2022, 23, 9105. [Google Scholar] [CrossRef]
- Wallace, J.M.; Robinson, J.J.; Wigzell, S.; Aitken, R.P. Effect of melatonin on the peripheral concentrations of LH and progesterone after oestrus, and on conception rate in ewes. J. Endocrinol. 1988, 119, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Abecia, J.; Forcada, F.; Lozano, J. A preliminary report on the effect of dietary energy on prostaglandin f2α production in vitro, interferon-tau synthesis by the conceptus, endometrial progesterone concentration on days 9 and 15 of pregnancy and associated rates of embryo wastage in ewes. Theriogenology 1999, 52, 1203–1213. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, D.; Ritta, M.; Fuentes, A.; Gimeno, M.; Gimeno, A. Prostaglandin E release by rat medial basal hypothalamus in vitro. Inhibition by melatonin at submicromolar concentrations. Eur. J. Pharmacol. 1980, 67, 151–153. [Google Scholar] [CrossRef] [PubMed]
- Abd-Allah, A. Effect of melatonin on estrogen and progesterone receptors in relation to uterine contraction in rats. Pharmacol. Res. 2003, 47, 349–354. [Google Scholar] [CrossRef]
- Cos, S.; Gonzalez, A.; Martinez-Campa, C.; Mediavilla, M.; Alonso-Gonzalez, C.; Sanchez-Barcelo, E. Melatonin as a Selective Estrogen Enzyme Modulator. Curr. Cancer Drug Targets 2008, 8, 691–702. [Google Scholar] [CrossRef]
- Brockus, K.; Hart, C.; Fleming, B.; Smith, T.; Ward, S.; Lemley, C. Effects of Supplementing Holstein Heifers with Dietary Melatonin during Late Gestation on Growth and Cardiovascular Measurements of their Offspring. Reprod. Domest. Anim. 2016, 51, 240–247. [Google Scholar] [CrossRef]
- Zhu, B.T. Functional role of estrogen metabolism in target cells: Review and perspectives. Carcinogenesis 1998, 19, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Männistö, P.T.; Kaakkola, S. Catechol-O-methyltransferase (COMT): Biochemistry, molecular biology, pharmacology, and clinical efficacy of the new selective COMT inhibitors. Pharmacol. Rev. 1999, 51, 593–628. [Google Scholar] [PubMed]
- Kanasaki, K.; Palmsten, K.; Sugimoto, H.; Ahmad, S.; Hamano, Y.; Xie, L.; Parry, S.; Augustin, H.G.; Gattone, V.H.; Folkman, J.; et al. Deficiency in catechol-O-methyltransferase and 2-methoxyoestradiol is associated with pre-eclampsia. Nature 2008, 453, 1117–1121. [Google Scholar] [CrossRef]
- Denison, M.S.; Nagy, S.R. Activation of the Aryl Hydrocarbon Receptor by Structurally Diverse Exogenous and Endogenous Chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef] [PubMed]
- Resnik, R.; Brink, G.W. Effects of prostaglandins E1, E2, and F2alpha on uterine blood flow in nonpregnant sheep. Am. J. Physiol. Circ. Physiol. 1978, 234, H557–H561. [Google Scholar] [CrossRef]
- Fowden, A.L.; Ward, J.W.; Wooding, F.P.B.; Forhead, A.; Constancia, M. Programming placental nutrient transport capacity. J. Physiol. 2006, 572, 5–15. [Google Scholar] [CrossRef]
- Ford, S.P. Control of blood flow to the gravid uterus of domestic livestock species. J. Anim. Sci. 1995, 73, 1852–1860. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Contreras-Correa, Z.E.; Messman, R.D.; Swanson, R.M.; Lemley, C.O. Melatonin in Health and Disease: A Perspective for Livestock Production. Biomolecules 2023, 13, 490. https://doi.org/10.3390/biom13030490
Contreras-Correa ZE, Messman RD, Swanson RM, Lemley CO. Melatonin in Health and Disease: A Perspective for Livestock Production. Biomolecules. 2023; 13(3):490. https://doi.org/10.3390/biom13030490
Chicago/Turabian StyleContreras-Correa, Zully E., Riley D. Messman, Rebecca M. Swanson, and Caleb O. Lemley. 2023. "Melatonin in Health and Disease: A Perspective for Livestock Production" Biomolecules 13, no. 3: 490. https://doi.org/10.3390/biom13030490
APA StyleContreras-Correa, Z. E., Messman, R. D., Swanson, R. M., & Lemley, C. O. (2023). Melatonin in Health and Disease: A Perspective for Livestock Production. Biomolecules, 13(3), 490. https://doi.org/10.3390/biom13030490