
β3 Adrenoceptor Agonism Prevents Hyperoxia-Induced Colonic Alterations

 , , , , ,
, , , , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Oxygen Exposure and Drug Administration
2.2. Flow Cytometric Analysis
2.3. Histological Examinations
2.4. Immunofluorescence Analysis
2.5. Statistical Analysis
3. Results
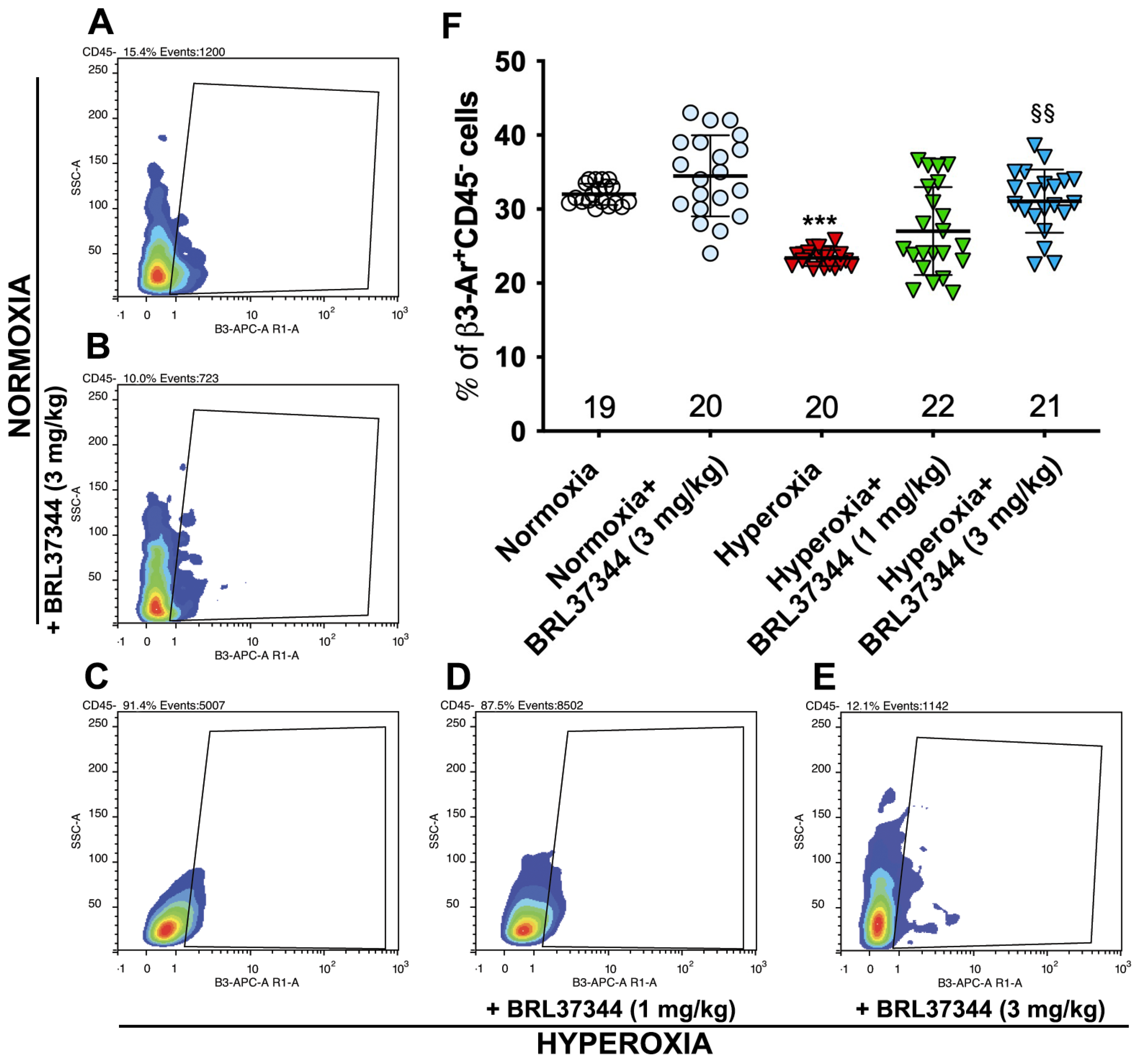
3.1. β3-AR Expression on Colonic Cells
3.2. Body Weight
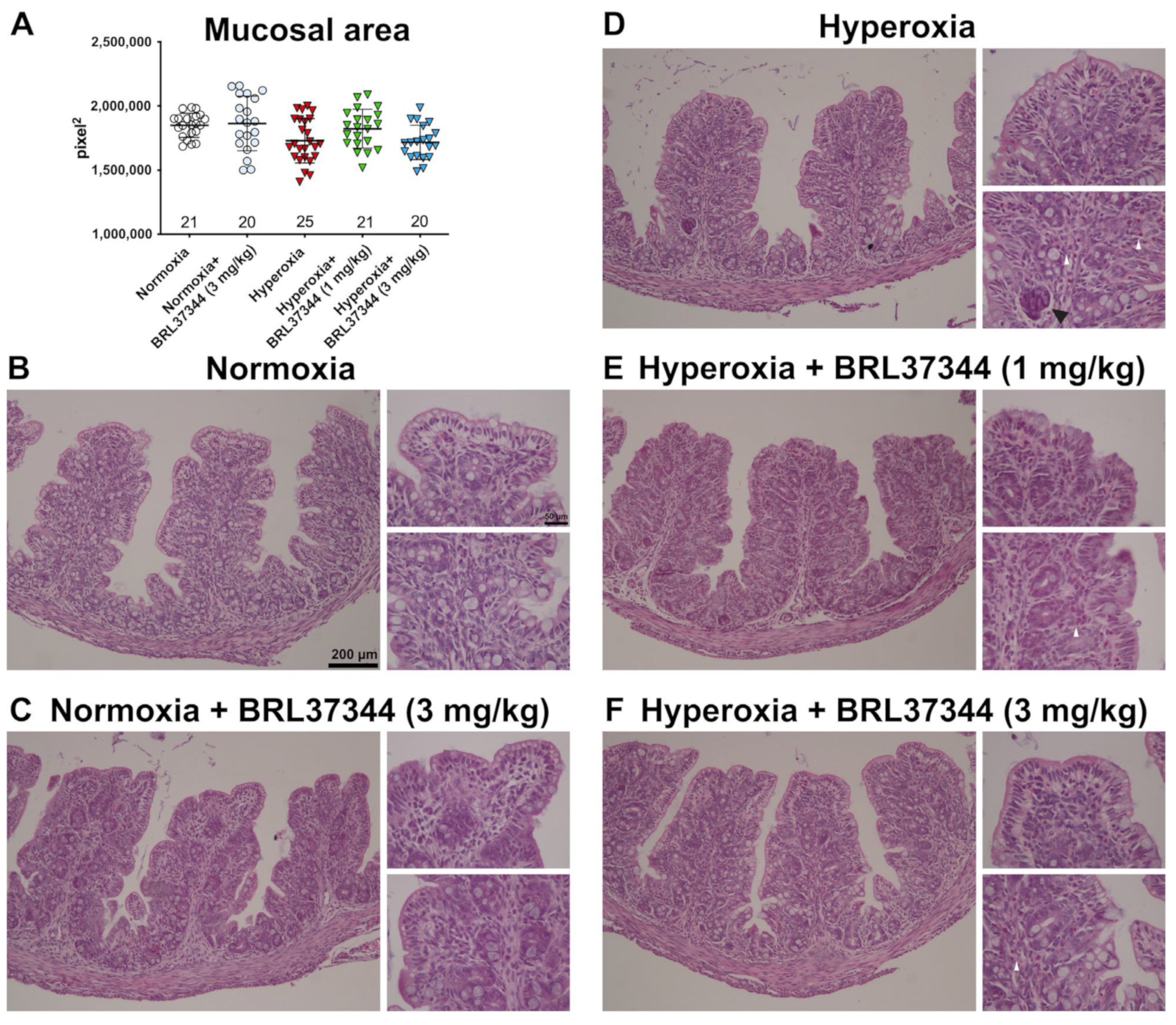
3.3. Colon Length and Mucosal Morphology
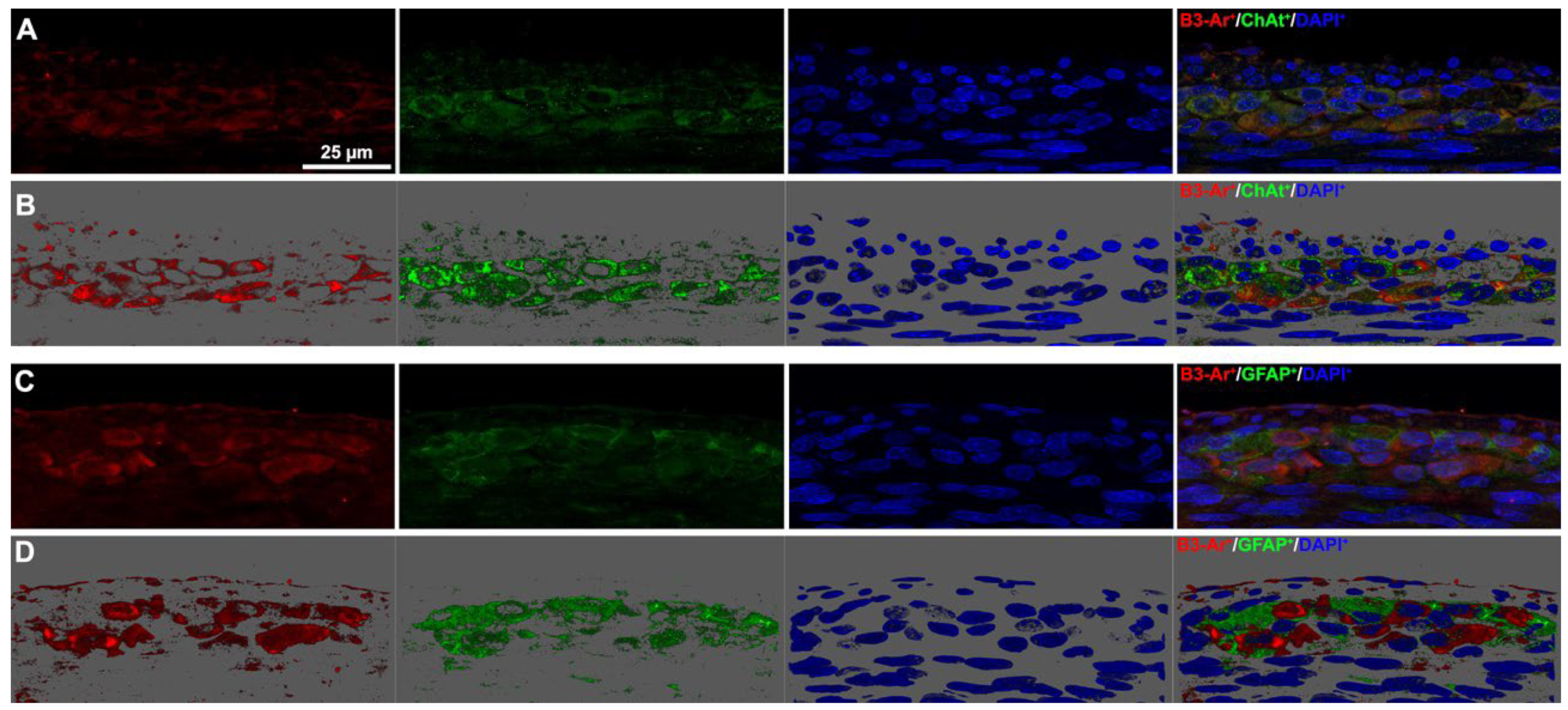
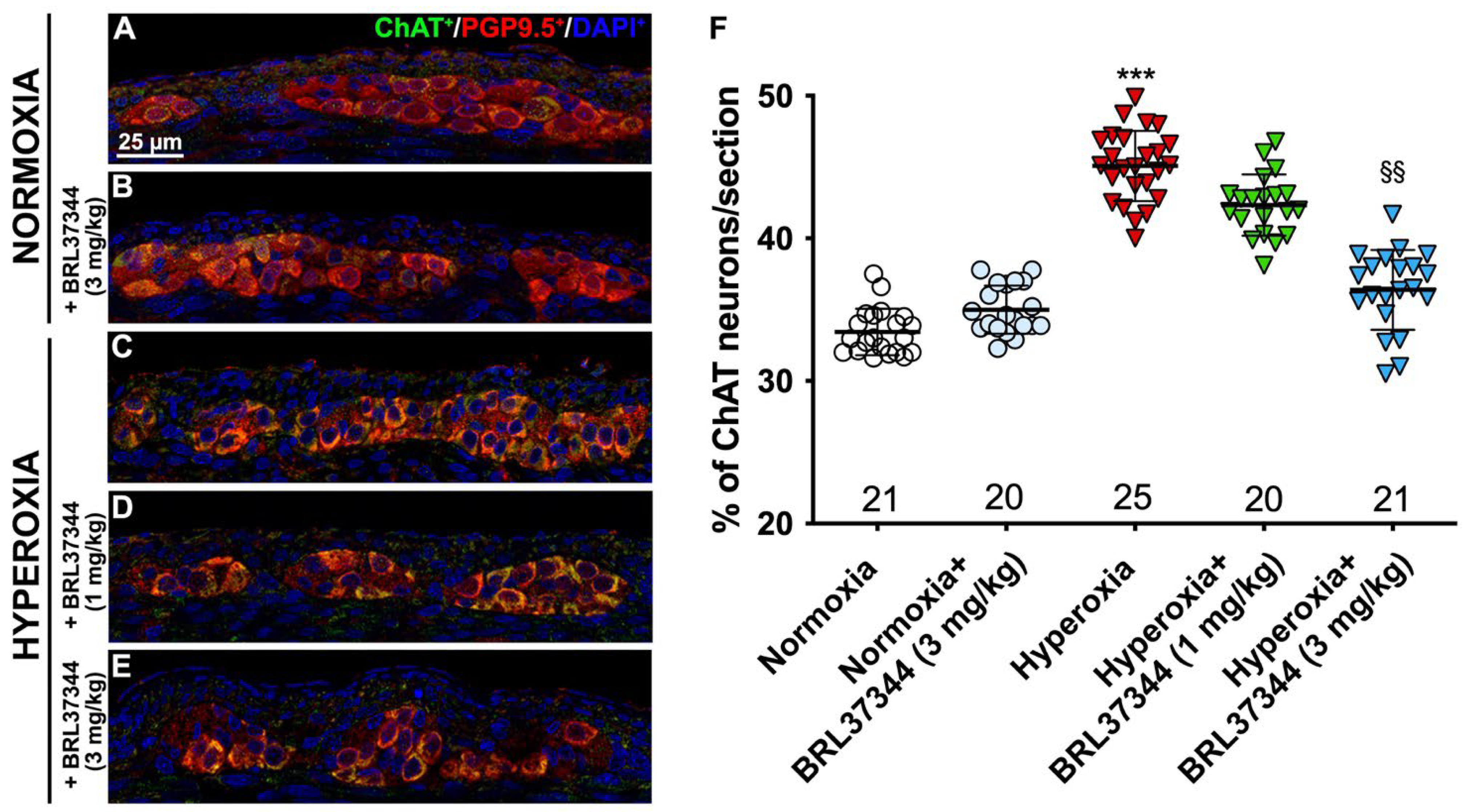
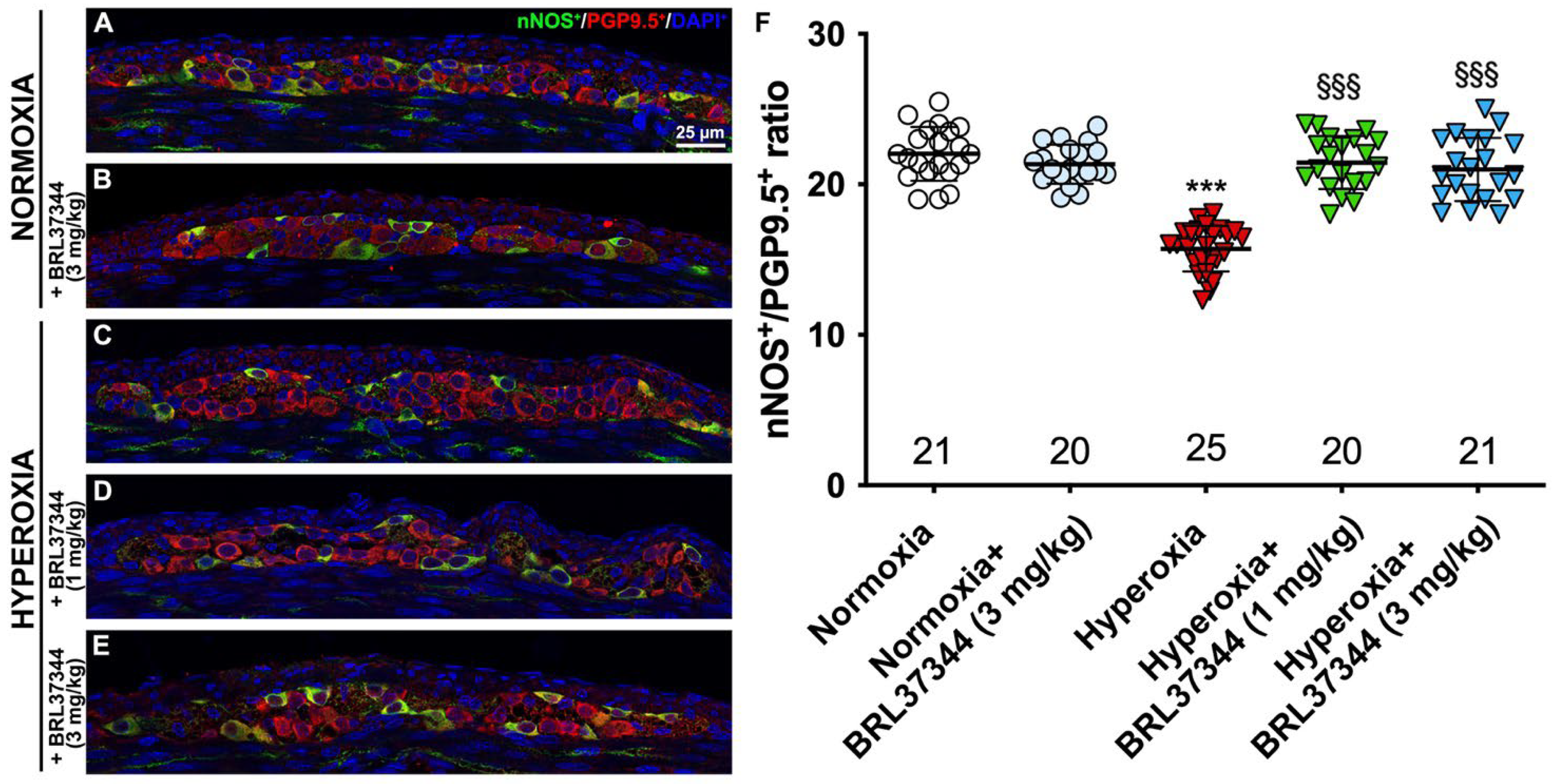
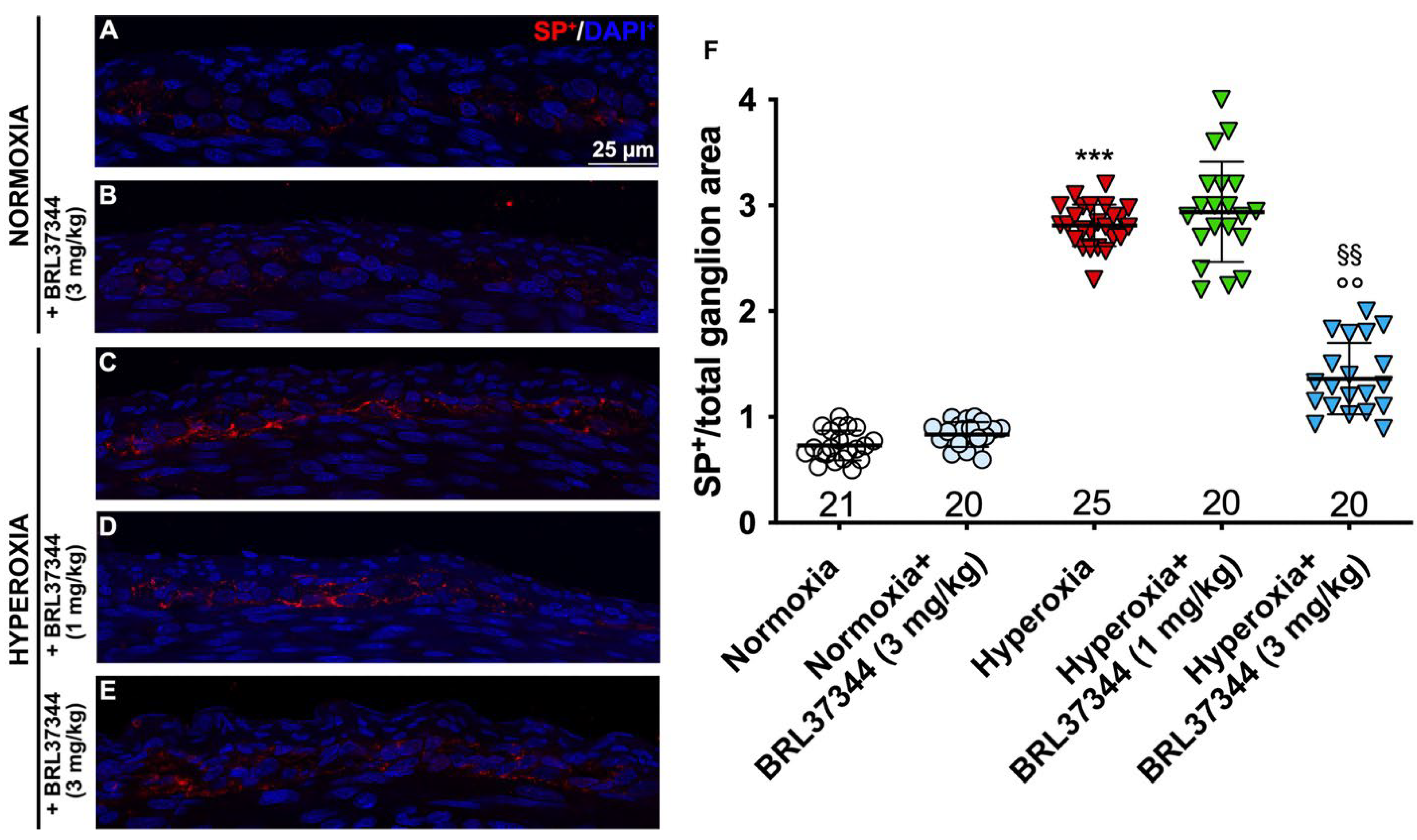
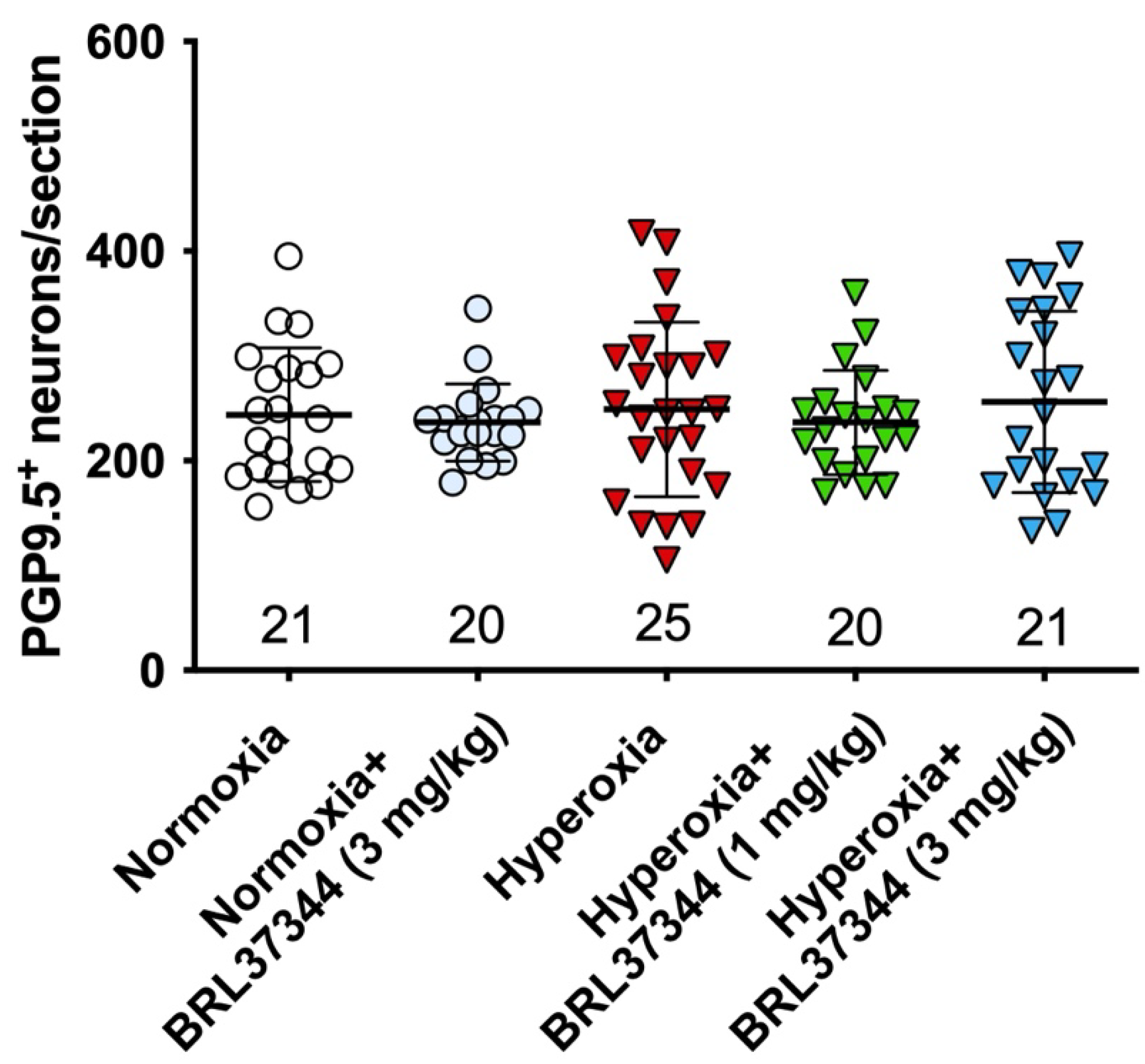
3.4. Neuronal Chemical Coding in the Myenteric Plexus
4. Discussion
5. Conclusions
6. Patent
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Imanirad, P.; Dzierzak, E. Hypoxia and HIFs in Regulating the Development of the Hematopoietic System. Blood Cells Mol. Dis. 2013, 51, 256–263. [Google Scholar] [CrossRef]
- Fathollahipour, S.; Patil, P.S.; Leipzig, N.D. Oxygen Regulation in Development: Lessons from Embryogenesis towards Tissue Engineering. Cells Tissues Organs 2018, 205, 350–371. [Google Scholar] [CrossRef] [PubMed]
- Coelho-Santos, V.; Shih, A.Y. Postnatal Development of Cerebrovascular Structure and the Neurogliovascular Unit. Wiley Interdiscip. Rev. Dev. Biol. 2020, 9, e363. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, M.; Thébaud, B. Animal Models of Bronchopulmonary Dysplasia. The Term Rat Models. Am. J. Physiol. Lung Cell Mol. Physiol. 2014, 307, L948–L958. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-M.; Chou, H.-C. Hyperoxia Disrupts the Intestinal Barrier in Newborn Rats. Exp. Mol. Pathol. 2016, 101, 44–49. [Google Scholar] [CrossRef]
- Filippi, L.; Pini, A.; Cammalleri, M.; Bagnoli, P.; Dal Monte, M. Β3-Adrenoceptor, a Novel Player in the Round-Trip from Neonatal Diseases to Cancer: Suggestive Clues from Embryo. Med. Res. Rev. 2022, 42, 1179–1201. [Google Scholar] [CrossRef]
- Giusto, K.; Wanczyk, H.; Jensen, T.; Finck, C. Hyperoxia-Induced Bronchopulmonary Dysplasia: Better Models for Better Therapies. Dis. Model. Mech. 2021, 14, dmm047753. [Google Scholar] [CrossRef] [PubMed]
- Giannone, P.J.; Bauer, J.A.; Schanbacher, B.L.; Reber, K.M. Effects of Hyperoxia on Postnatal Intestinal Development. Biotech. Histochem. 2007, 82, 17–22. [Google Scholar] [CrossRef]
- Liu, D.Y.; Lou, W.J.; Zhang, D.Y.; Sun, S.Y. ROS Plays a Role in the Neonatal Rat Intestinal Barrier Damages Induced by Hyperoxia. BioMed Res. Int. 2020, 2020, 8819195. [Google Scholar] [CrossRef]
- Filippi, L.; Scaramuzzo, R.T.; Pascarella, F.; Pini, A.; Morganti, R.; Cammalleri, M.; Bagnoli, P.; Ciantelli, M. Fetal Oxygenation in the Last Weeks of Pregnancy Evaluated through the Umbilical Cord Blood Gas Analysis. Front. Pediatr. 2023, 11, 1140021. [Google Scholar] [CrossRef]
- Emorine, L.J.; Marullo, S.; Briend-Sutren, M.M.; Patey, G.; Tate, K.; Delavier-Klutchko, C.; Strosberg, A.D. Molecular Characterization of the Human Beta 3-Adrenergic Receptor. Science 1989, 245, 1118–1121. [Google Scholar] [CrossRef] [PubMed]
- Dal Monte, M.; Filippi, L.; Bagnoli, P. Beta3-Adrenergic Receptors Modulate Vascular Endothelial Growth Factor Release in Response to Hypoxia through the Nitric Oxide Pathway in Mouse Retinal Explants. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2013, 386, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Semenza, G.L.; Wang, G.L. A Nuclear Factor Induced by Hypoxia via de Novo Protein Synthesis Binds to the Human Erythropoietin Gene Enhancer at a Site Required for Transcriptional Activation. Mol. Cell. Biol. 1992, 12, 5447–5454. [Google Scholar] [CrossRef] [PubMed]
- Amato, R.; Pisani, F.; Laudadio, E.; Cammalleri, M.; Lucchesi, M.; Marracci, S.; Filippi, L.; Galeazzi, R.; Svelto, M.; Dal Monte, M.; et al. HIF-1-Dependent Induction of Β3 Adrenoceptor: Evidence from the Mouse Retina. Cells 2022, 11, 1271. [Google Scholar] [CrossRef] [PubMed]
- Fujinaga, M.; Scott, J.C. Gene Expression of Catecholamine Synthesizing Enzymes and Beta Adrenoceptor Subtypes during Rat Embryogenesis. Neurosci. Lett. 1997, 231, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Resch, B.E.; Ducza, E.; Gáspár, R.; Falkay, G. Role of Adrenergic Receptor Subtypes in the Control of Human Placental Blood Vessels. Mol. Reprod. Dev. 2003, 66, 166–171. [Google Scholar] [CrossRef]
- Hynes, P.G.; Friel, A.M.; Smith, T.J.; Morrison, J.J. Beta-Adrenoceptor Subtype Expression in Human Placenta and Umbilical Arteries in Normal and Preeclamptic Pregnancies. Hypertens. Pregnancy 2008, 27, 169–181. [Google Scholar] [CrossRef]
- Pini, A.; Fazi, C.; Nardini, P.; Calvani, M.; Fabbri, S.; Guerrini, A.; Forni, G.; La Marca, G.; Rosa, A.C.; Filippi, L. Effect of Beta 3 Adrenoreceptor Modulation on Patency of the Ductus Arteriosus. Cells 2020, 9, 2625. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Collins, J.M.; Hyland, N.P.; Clarke, G.; Fitzgerald, P.; Julio-Pieper, M.; Bulmer, D.C.; Dinan, T.G.; Cryan, J.F.; O’Mahony, S.M. Beta 3-Adrenoceptor Agonism Ameliorates Early-Life Stress-Induced Visceral Hypersensitivity in Male Rats. J. Neurochem. 2023, 00, 1–14. [Google Scholar] [CrossRef]
- Calvani, M.; Bruno, G.; Dal Monte, M. Beta3 -Adrenoceptor as a Potential Immuno-Suppressor Agent in Melanoma. Br. J. Pharmacol. 2019, 176, 2509–2524. [Google Scholar] [CrossRef]
- Bruno, G.; Nastasi, N.; Subbiani, A.; Boaretto, A.; Ciullini Mannurita, S.; Mattei, G.; Nardini, P.; Della Bella, C.; Magi, A.; Pini, A.; et al. Β3-Adrenergic Receptor on Tumor-Infiltrating Lymphocytes Sustains IFN-γ-Dependent PD-L1 Expression and Impairs Anti-Tumor Immunity in Neuroblastoma. Cancer Gene Ther. 2023, 30, 890–904. [Google Scholar] [CrossRef]
- Misharin, A.V.; Morales-Nebreda, L.; Mutlu, G.M.; Budinger, G.R.S.; Perlman, H. Flow Cytometric Analysis of Macrophages and Dendritic Cell Subsets in the Mouse Lung. Am. J. Respir. Cell Mol. Biol. 2013, 49, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Erben, U.; Loddenkemper, C.; Doerfel, K.; Spieckermann, S.; Haller, D.; Heimesaat, M.M.; Zeitz, M.; Siegmund, B.; Kühl, A.A. A Guide to Histomorphological Evaluation of Intestinal Inflammation in Mouse Models. Int. J. Clin. Exp. Pathol. 2014, 7, 4557–4576. [Google Scholar]
- Rosa, A.C.; Nardini, P.; Sgambellone, S.; Gurrieri, M.; Spampinato, S.F.; Dell’Accio, A.; Chazot, P.L.; Obara, I.; Liu, W.L.; Pini, A. CNS-Sparing Histamine H3 Receptor Antagonist as a Candidate to Prevent the Diabetes-Associated Gastrointestinal Symptoms. Biomolecules 2022, 12, 184. [Google Scholar] [CrossRef] [PubMed]
- Cellek, S.; Thangiah, R.; Bassil, A.K.; Campbell, C.A.; Gray, K.M.; Stretton, J.L.; Lalude, O.; Vivekanandan, S.; Wheeldon, A.; Winchester, W.J.; et al. Demonstration of Functional Neuronal Beta3-Adrenoceptors within the Enteric Nervous System. Gastroenterology 2007, 133, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Chou, H.-C.; Chen, C.-M. Neonatal Hyperoxia Disrupts the Intestinal Barrier and Impairs Intestinal Function in Rats. Exp. Mol. Pathol. 2017, 102, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, C.; Leitz, M.R.; Oberdorf-Maass, S.; Lohse, M.J.; Klotz, K.-N. Comparative Pharmacology of Human Beta-Adrenergic Receptor Subtypes-Characterization of Stably Transfected Receptors in CHO Cells. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 369, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Schena, G.; Caplan, M.J. Everything You Always Wanted to Know about β3-AR * (* But Were Afraid to Ask). Cells 2019, 8, 357. [Google Scholar] [CrossRef]
- Ngala, R.A.; O’Dowd, J.; Wang, S.J.; Stocker, C.; Cawthorne, M.A.; Arch, J.R.S. Beta2-Adrenoceptors and Non-Beta-Adrenoceptors Mediate Effects of BRL37344 and Clenbuterol on Glucose Uptake in Soleus Muscle: Studies Using Knockout Mice. Br. J. Pharmacol. 2009, 158, 1676–1682. [Google Scholar] [CrossRef]
- Barbier, M.; Attoub, S.; Joubert, M.; Bado, A.; Laboisse, C.; Cherbut, C.; Galmiche, J.P. Proinflammatory Role of Leptin in Experimental Colitis in Rats Benefit of Cholecystokinin-B Antagonist and Beta3-Agonist. Life Sci. 2001, 69, 567–580. [Google Scholar] [CrossRef] [PubMed]
- Filippi, L.; Cammalleri, M.; Amato, R.; Ciantelli, M.; Pini, A.; Bagnoli, P.; Dal Monte, M. Decoupling Oxygen Tension from Retinal Vascularization as a New Perspective for Management of Retinopathy of Prematurity. New Opportunities from β-Adrenoceptors. Front. Pharmacol. 2022, 13, 835771. [Google Scholar] [CrossRef] [PubMed]
- Henning, S.J. Postnatal Development: Coordination of Feeding, Digestion, and Metabolism. Am. J. Physiol. 1981, 241, G199–G214. [Google Scholar] [CrossRef] [PubMed]
- Kanzler, S.A.; Januario, A.C.; Paschoalini, M.A. Involvement of Β3-Adrenergic Receptors in the Control of Food Intake in Rats. Braz. J. Med. Biol. Res. 2011, 44, 1141–1147. [Google Scholar] [CrossRef]
- Richard, J.E.; López-Ferreras, L.; Chanclón, B.; Eerola, K.; Micallef, P.; Skibicka, K.P.; Wernstedt Asterholm, I. CNS β3-Adrenergic Receptor Activation Regulates Feeding Behavior, White Fat Browning, and Body Weight. Am. J. Physiol. Endocrinol. Metab. 2017, 313, E344–E358. [Google Scholar] [CrossRef] [PubMed]
- Nardini, P.; Pini, A.; Bessard, A.; Duchalais, E.; Niccolai, E.; Neunlist, M.; Vannucchi, M.G. GLP-2 Prevents Neuronal and Glial Changes in the Distal Colon of Mice Chronically Treated with Cisplatin. Int. J. Mol. Sci. 2020, 21, 8875. [Google Scholar] [CrossRef]
- Domènech, A.; Pasquinelli, G.; De Giorgio, R.; Gori, A.; Bosch, F.; Pumarola, M.; Jiménez, M. Morphofunctional Changes Underlying Intestinal Dysmotility in Diabetic RIP-I/hIFNβ Transgenic Mice. Int. J. Exp. Pathol. 2011, 92, 400–412. [Google Scholar] [CrossRef]
- Kang, Y.; Park, H.; Choe, B.-H.; Kang, B. The Role and Function of Mucins and Its Relationship to Inflammatory Bowel Disease. Front. Med. 2022, 9, 848344. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Faussone-Pellegrini, M.S. Differentiation of Cholinergic Cells in the Rat Gut during Pre- and Postnatal Life. Neurosci. Lett. 1996, 206, 105–108. [Google Scholar] [CrossRef]
- De Vries, P.; Soret, R.; Suply, E.; Heloury, Y.; Neunlist, M. Postnatal Development of Myenteric Neurochemical Phenotype and Impact on Neuromuscular Transmission in the Rat Colon. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G539–G547. [Google Scholar] [CrossRef]
- Parathan, P.; Wang, Y.; Leembruggen, A.J.; Bornstein, J.C.; Foong, J.P. The Enteric Nervous System Undergoes Significant Chemical and Synaptic Maturation during Adolescence in Mice. Dev. Biol. 2020, 458, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Winston, J.H.; Li, Q.; Sarna, S.K. Paradoxical Regulation of ChAT and nNOS Expression in Animal Models of Crohn’s Colitis and Ulcerative Colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G295–G302. [Google Scholar] [CrossRef]
- Caetano, M.A.F.; Magalhães, H.I.R.; Duarte, J.R.L.; Conceição, L.B.; Castelucci, P. Butyrate Protects Myenteric Neurons Loss in Mice Following Experimental Ulcerative Colitis. Cells 2023, 12, 1672. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, F.; Nakahara, T.; Akanuma, K.; Ueda, K.; Mori, A.; Sakamoto, K.; Ishii, K. Protective Effects of the Β3-Adrenoceptor Agonist CL316243 against N-Methyl-D-Aspartate-Induced Retinal Neurotoxicity. Naunyn Schmiedebergs Arch. Pharmacol. 2012, 385, 1077–1081. [Google Scholar] [CrossRef] [PubMed]
- Tournissac, M.; Vu, T.-M.; Vrabic, N.; Hozer, C.; Tremblay, C.; Mélançon, K.; Planel, E.; Pifferi, F.; Calon, F. Repurposing Beta-3 Adrenergic Receptor Agonists for Alzheimer’s Disease: Beneficial Effects in a Mouse Model. Alzheimers Res. Ther. 2021, 13, 103. [Google Scholar] [CrossRef]
- Nasser, Y.; Ho, W.; Sharkey, K.A. Distribution of Adrenergic Receptors in the Enteric Nervous System of the Guinea Pig, Mouse, and Rat. J. Comp. Neurol. 2006, 495, 529–553. [Google Scholar] [CrossRef]
- Anthony, A.; Schepelmann, S.; Guillaume, J.L.; Strosberg, A.D.; Dhillon, A.P.; Pounder, R.E.; Wakefield, A.J. Localization of the β3-Adrenoceptor in the Human Gastrointestinal Tract: An Immunohistochemical Study. Aliment. Pharmacol. Ther. 1998, 12, 519–525. [Google Scholar] [CrossRef]
- Pawolski, V.; Schmidt, M.H.H. Neuron-Glia Interaction in the Developing and Adult Enteric Nervous System. Cells 2020, 10, 47. [Google Scholar] [CrossRef]
- Li, T.-M.; Liu, D.-Y. Mechanism of Neonatal Intestinal Injury Induced by Hyperoxia Therapy. J. Immunol. Res. 2022, 2022, 2316368. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antigen | Species | Source | Catalog Number | RRID | Concentration |
|---|---|---|---|---|---|
| Primary Antisera | |||||
| β3-AR | Rabbit | GeneTex (Irvine, CA, USA) | GTX70685 | AB_375242 | 1:300 |
| ChAT | Chicken | GeneTex (Irvine, CA, USA) | GTX85450 | AB_10622340 | 1:200 |
| GFAP | Chicken | Abcam (Cambridge, UK) | ab4674 | AB_304558 | 1:1000 |
| nNOS | Rabbit | GeneTex (Irvine, CA, USA) | GTX133403 | AB_2886968 | 1:500 |
| PGP 9.5 | Mouse | GeneTex (Irvine, CA, USA) | GTX634797 | AB_2888478 | 1:300 |
| SP | Goat | Santa Cruz Biotech (Santa Cruz, CA, USA) | AB_661439 | AB_661439 | 1:100 |
| Secondary Antisera | |||||
| Alexa Fluor 488 | Donkey | Jackson ImmunoResearch (Ely, UK) | 711-545-152 | AB_2313584 | 1:175 |
| Alexa Fluor 488 | Donkey | Jackson ImmunoResearch (Ely, UK) | 703-545-155 | AB_2340375 | 1:175 |
| Alexa Fluor 594 | Donkey | Jackson ImmunoResearch (Ely, UK) | 711-585-152 | AB_2340621 | 1:175 |
| Alexa Fluor 594 | Bovine | Jackson ImmunoResearch (Ely, UK) | 805-585-180 | AB_2340884 | 1:175 |
| Alexa Fluor 594 | Sheep | Jackson Immunoresearch (Ely, UK) | 515-585-062 | AB_2340337 | 1:175 |
| Experimental Group | Pups/Group | Age (d) | Body Weight (g) | Age (d) | Body Weight (g) |
|---|---|---|---|---|---|
| Normoxia | 21 | 0 | 6.5 ± 0.6 | 14 | 33.5 ± 1.7 |
| Normoxia + BRL37344 (3 mg/kg) | 20 | 0 | 6.5 ± 0.7 | 14 | 32.1 ± 1.9 |
| Hyperoxia | 25 | 0 | 6.4 ± 0.7 | 14 | 21.4 ± 1.7 *** |
| Hyperoxia + BRL37344 (1 mg/kg) | 22 | 0 | 6.5 ± 0.9 | 14 | 21.8 ± 1.6 °°° |
| Hyperoxia + BRL37344 (3 mg/kg) | 21 | 0 | 6.6 ± 0.9 | 14 | 23.5 ± 1.7 °°° |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippi, L.; Nardini, P.; Zizi, V.; Molino, M.; Fazi, C.; Calvani, M.; Carrozzo, F.; Cavallaro, G.; Giuseppetti, G.; Calosi, L.; et al. β3 Adrenoceptor Agonism Prevents Hyperoxia-Induced Colonic Alterations. Biomolecules 2023, 13, 1755. https://doi.org/10.3390/biom13121755
Filippi L, Nardini P, Zizi V, Molino M, Fazi C, Calvani M, Carrozzo F, Cavallaro G, Giuseppetti G, Calosi L, et al. β3 Adrenoceptor Agonism Prevents Hyperoxia-Induced Colonic Alterations. Biomolecules. 2023; 13(12):1755. https://doi.org/10.3390/biom13121755
Chicago/Turabian StyleFilippi, Luca, Patrizia Nardini, Virginia Zizi, Marta Molino, Camilla Fazi, Maura Calvani, Francesco Carrozzo, Giacomo Cavallaro, Giorgia Giuseppetti, Laura Calosi, and et al. 2023. "β3 Adrenoceptor Agonism Prevents Hyperoxia-Induced Colonic Alterations" Biomolecules 13, no. 12: 1755. https://doi.org/10.3390/biom13121755
APA StyleFilippi, L., Nardini, P., Zizi, V., Molino, M., Fazi, C., Calvani, M., Carrozzo, F., Cavallaro, G., Giuseppetti, G., Calosi, L., Crociani, O., & Pini, A. (2023). β3 Adrenoceptor Agonism Prevents Hyperoxia-Induced Colonic Alterations. Biomolecules, 13(12), 1755. https://doi.org/10.3390/biom13121755