
Effects of 1,25-Vitamin D3 and 24,25-Vitamin D3 on Corneal Nerve Regeneration in Diabetic Mice



Abstract
:1. Introduction
2. Material and Methods
2.1. Materials
2.2. Animal Experiments
2.3. Corneal Injury
2.4. Immunostaining for CD 45+ Cells and Macrophages
2.5. Vitamin D Administration and Treatment
2.6. Nerve Immunofluorescence Labeling and Imaging
2.7. Protein Extraction from Culture Medium
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
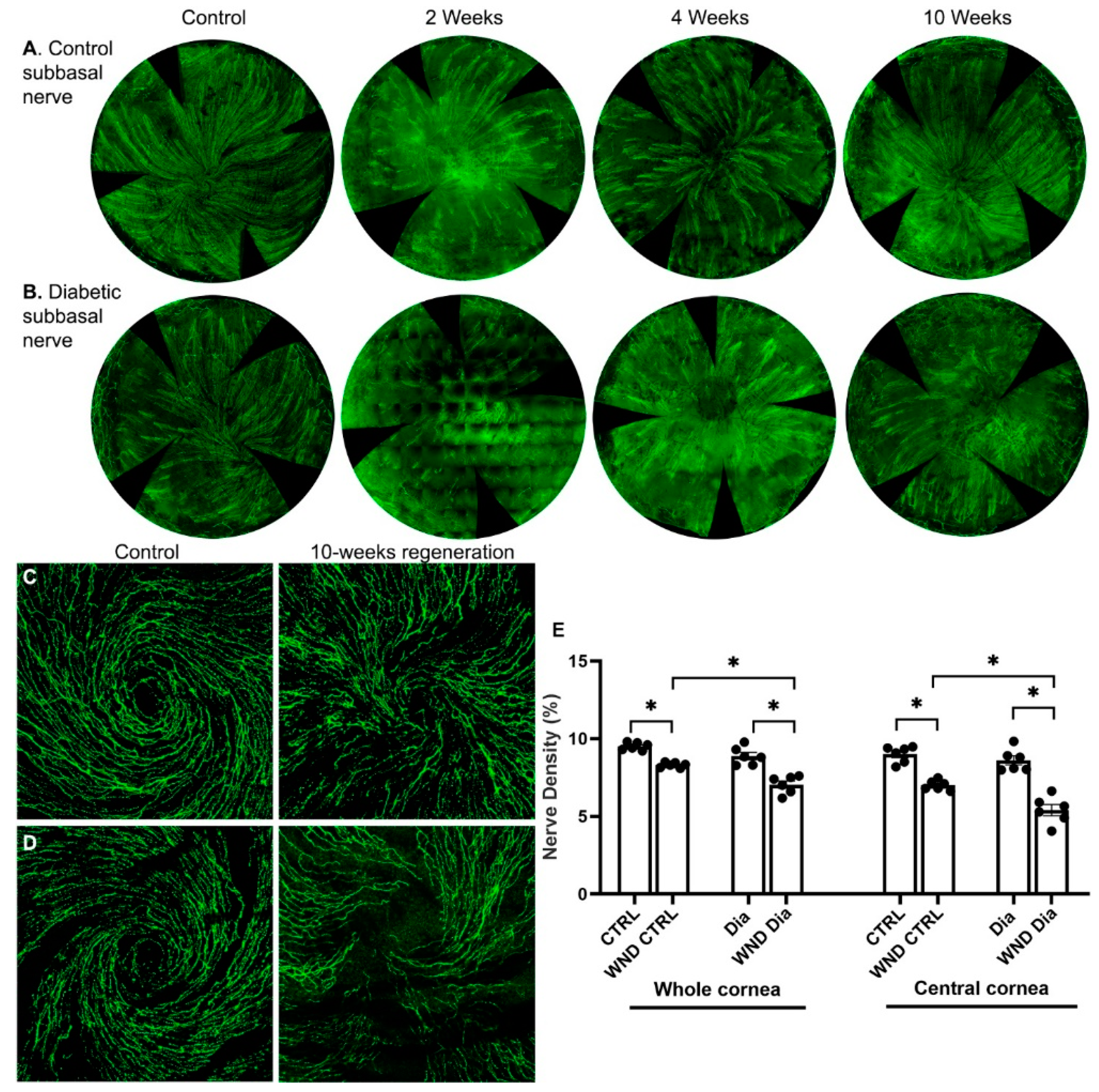
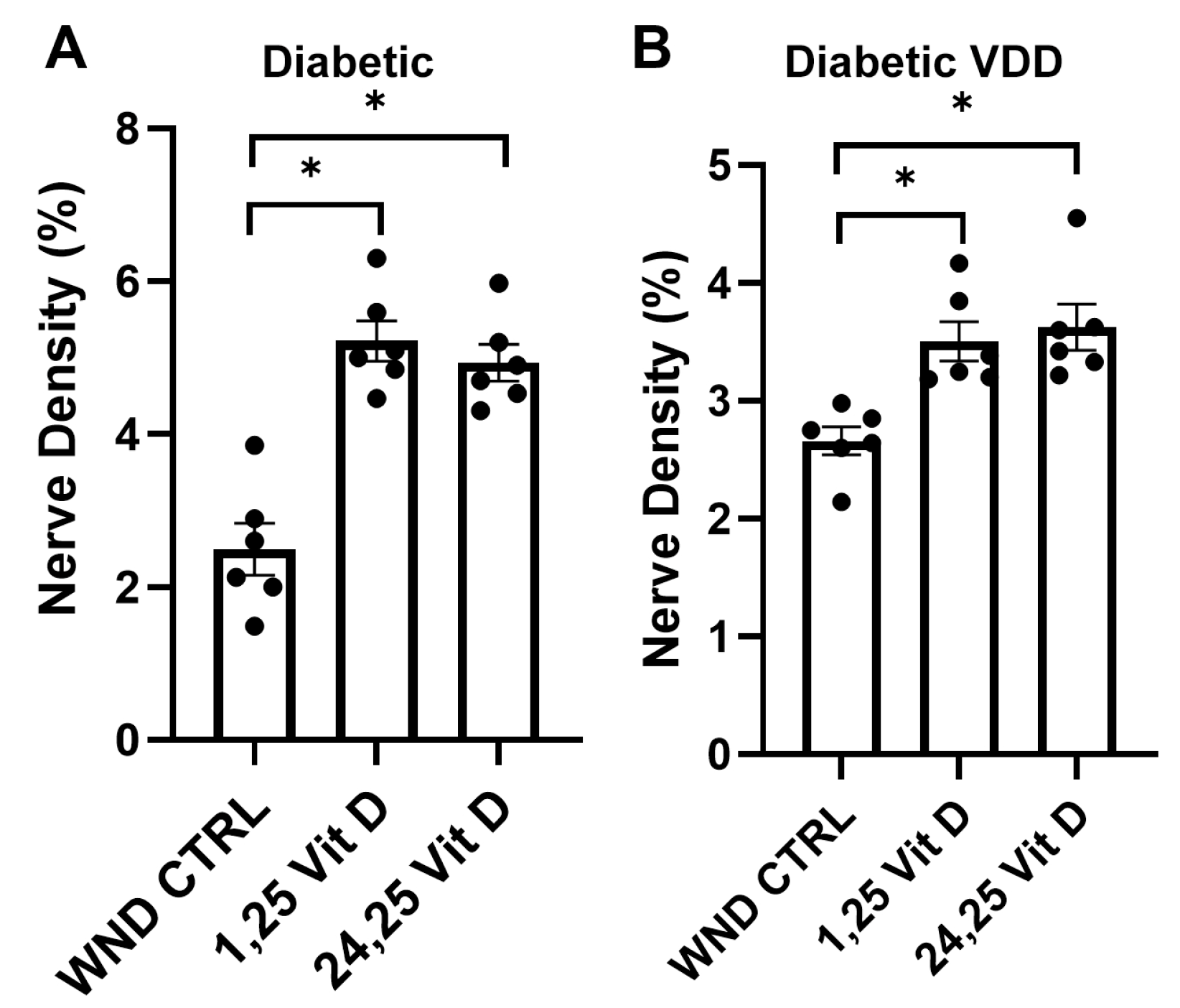
3.1. Diabetic Nerve Regeneration
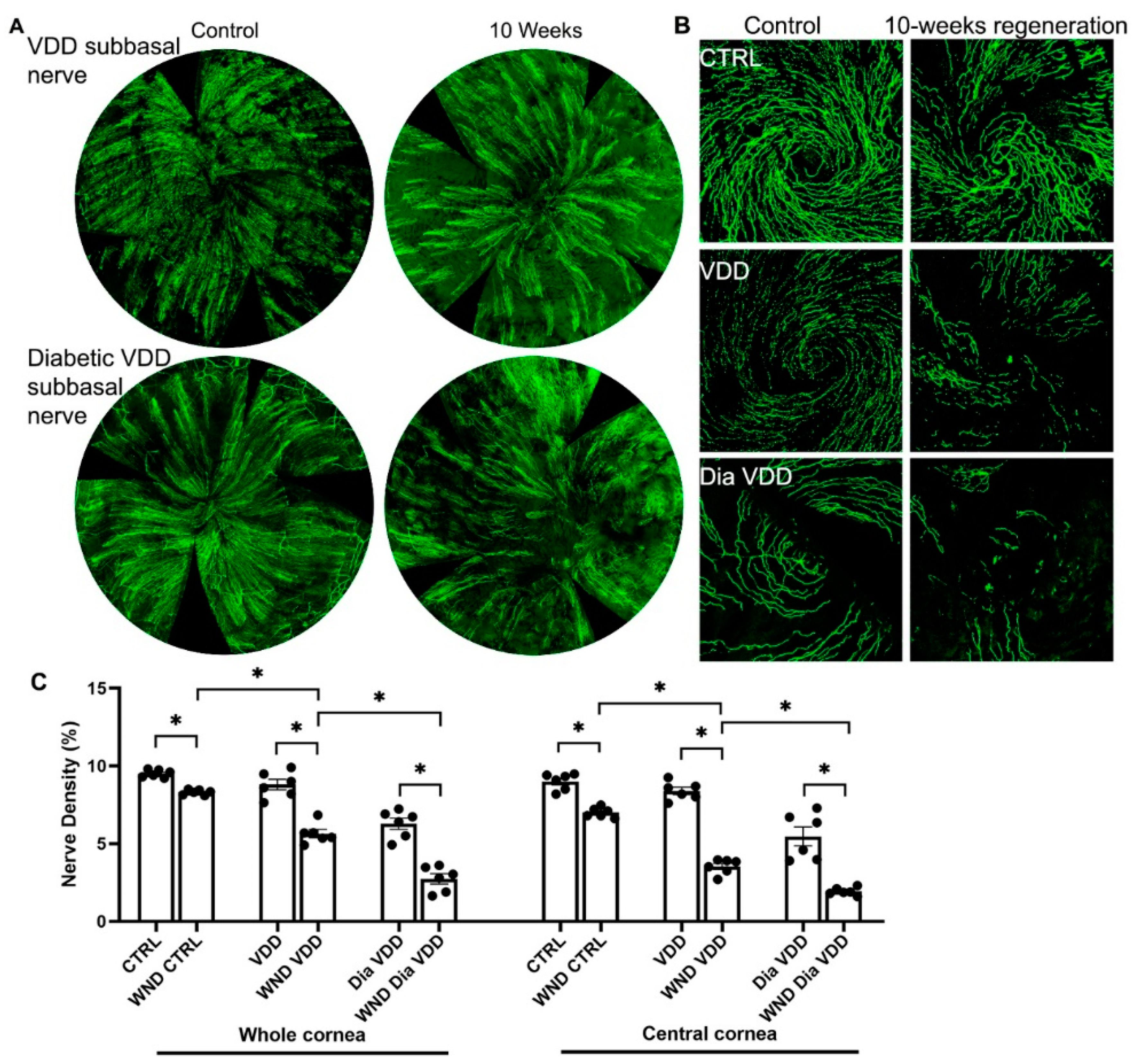
3.2. Diabetic Vitamin D Deficiency Nerve Regeneration
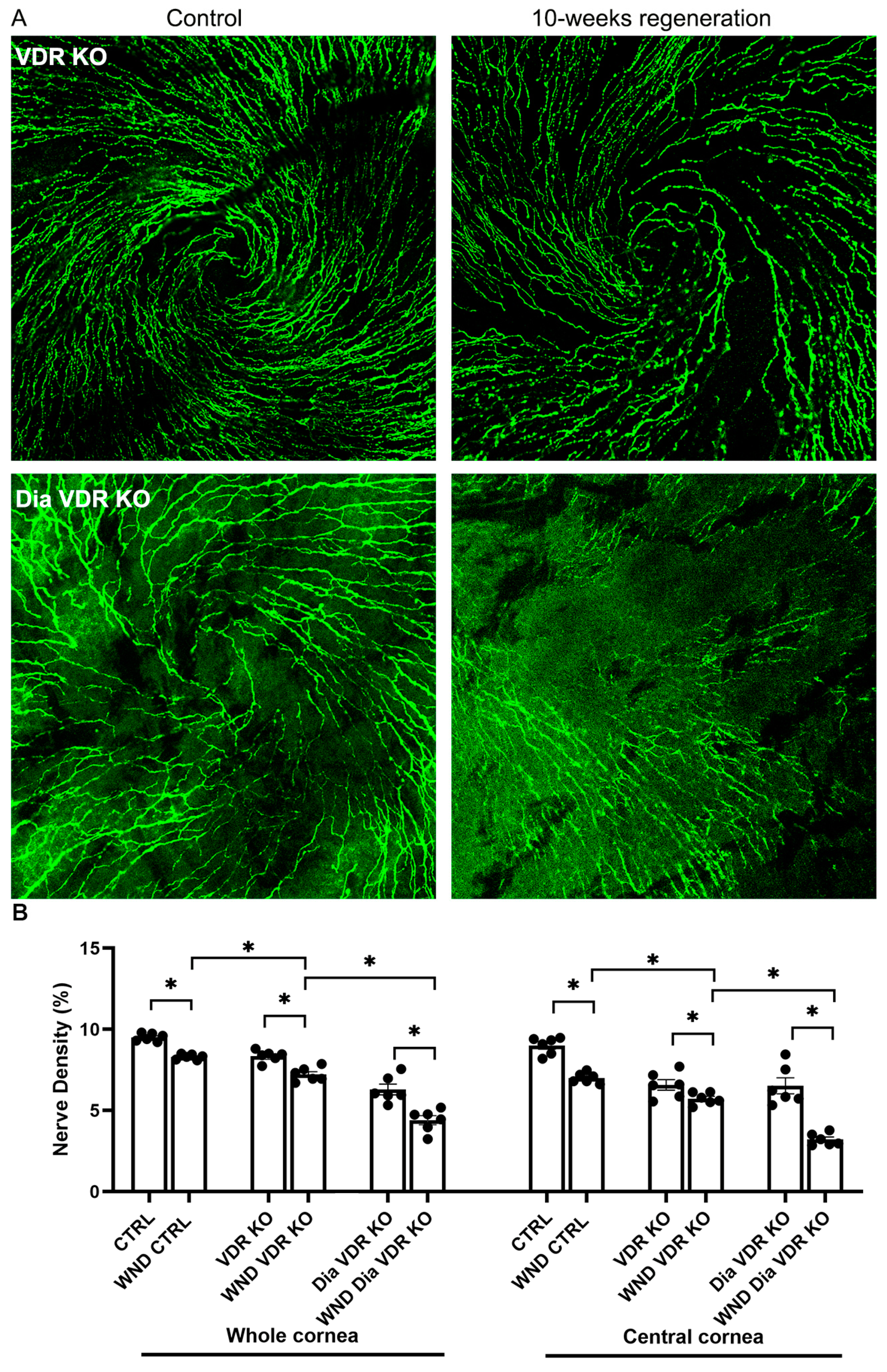
3.3. Diabetic VDR KO and VDD Nerve Regeneration
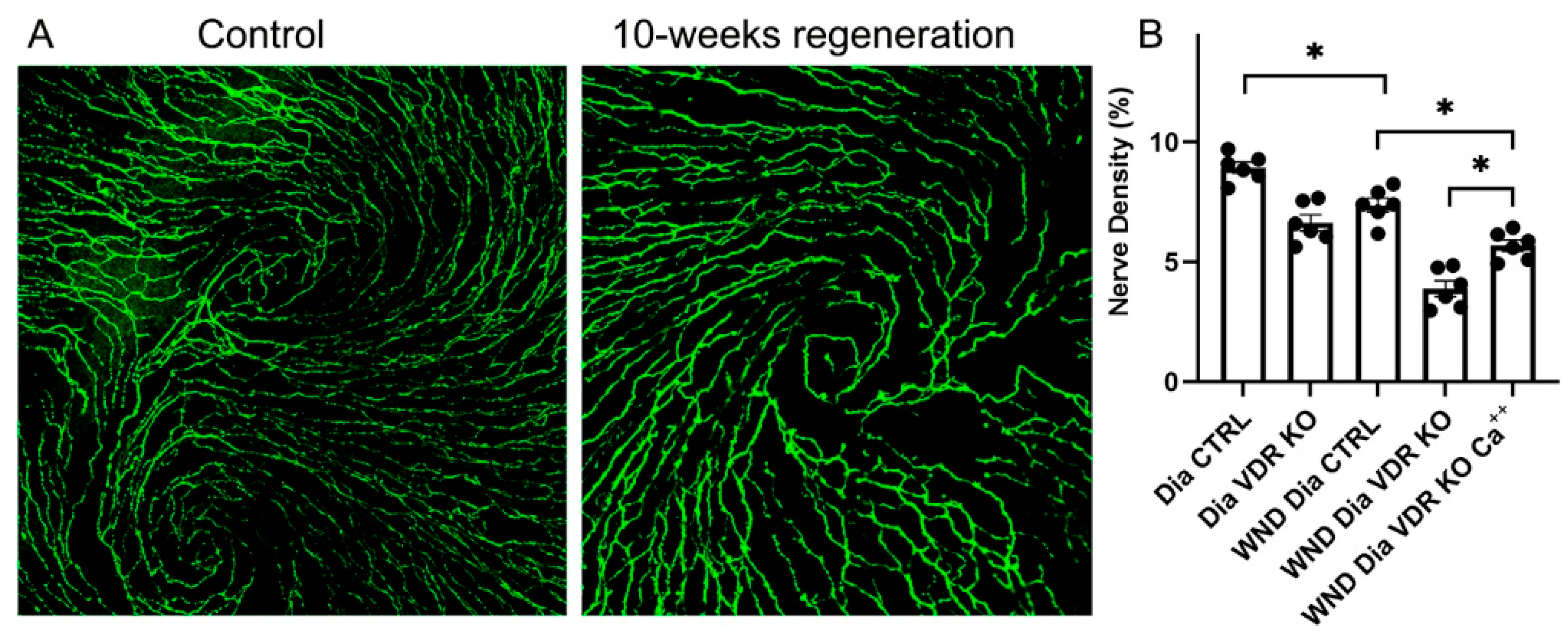
3.4. Nerve Regeneration in Dia VDR KO Mice Fed the Supplemental Diet
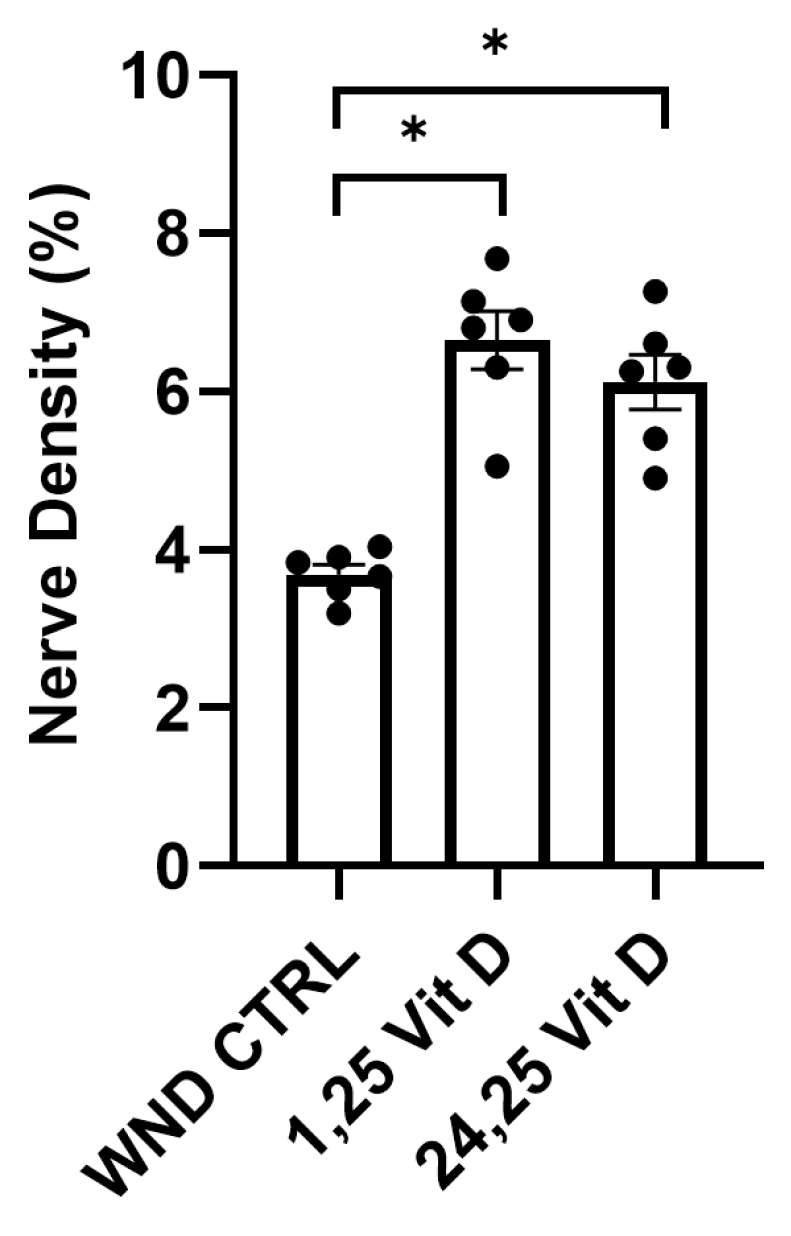
3.5. Nerve Regeneration in Wounded Normoglycemic Mouse Corneas Following Topical Vit D Treatment
3.6. Nerve Regeneration in Diabetic and Dia VDD Mouse Corneas Following Topical Vit D Treatment
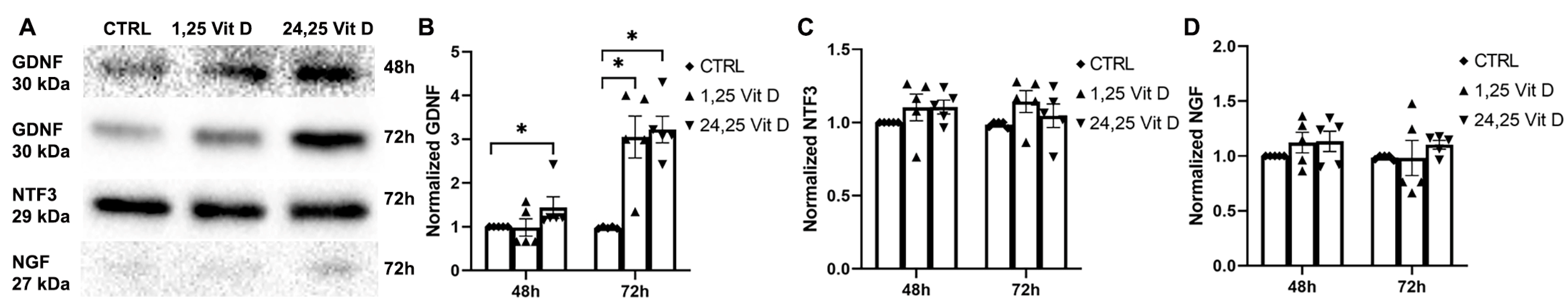
3.7. Effects of 1,25 Vit D and 24,25 Vit D on NGF Protein Expression
3.8. Effects of 1,25 Vit D and 24,25 Vit D on NTF3 Protein Expression
3.9. Effects of 1,25 Vit D and 24,25 Vit D on GDNF Protein Expression
3.10. Effects of 1,25 Vit D and 24,25 Vit D on HCEC NGF, NTF3, and GDNF Protein Secretion
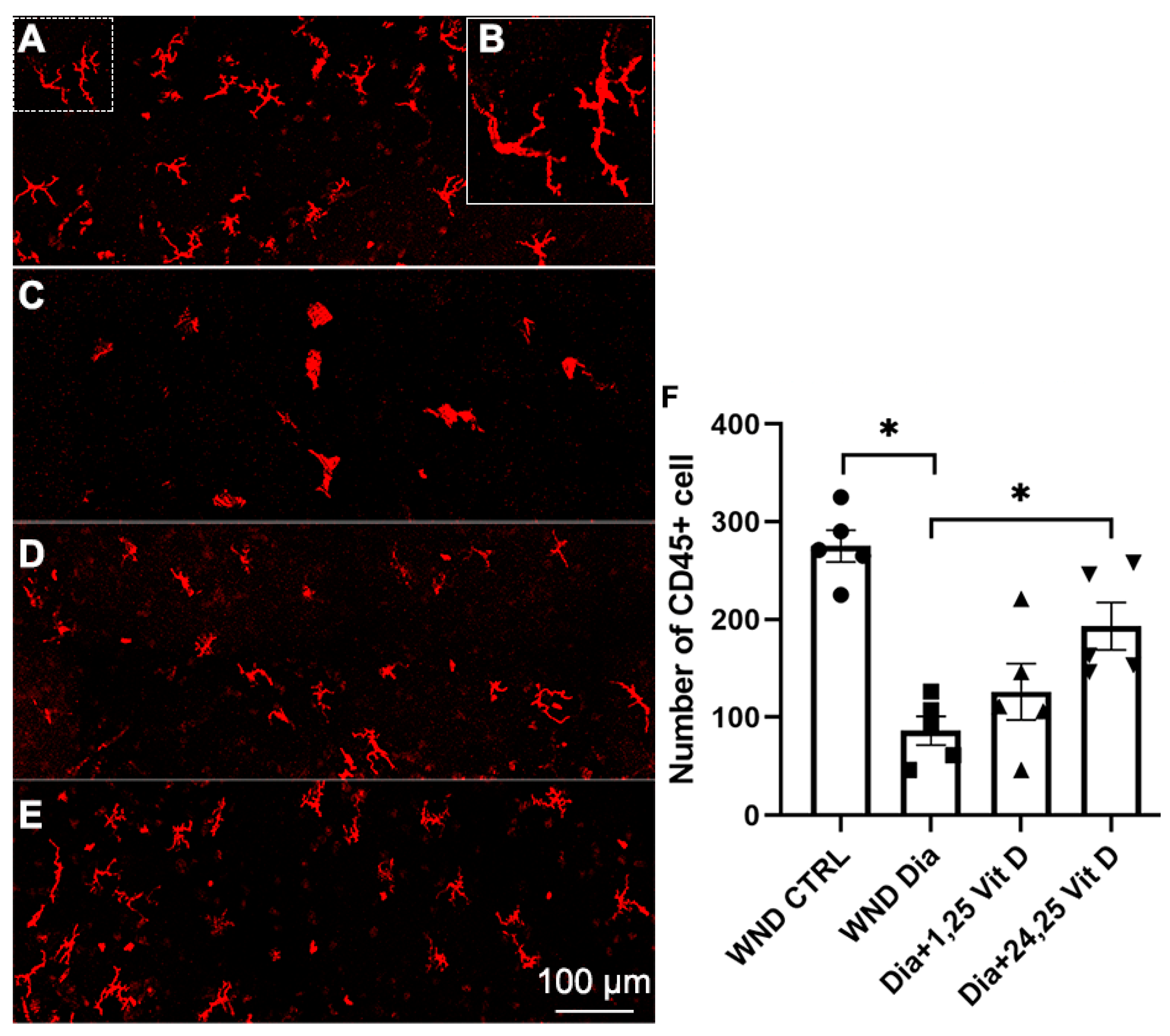
3.11. CD45+ Cell Recruitment Following 1,25 Vit D and 24,25 Vit D Treatment
3.12. Macrophage Recruitment Following 1,25 Vit D and 24,25 Vit D Treatment
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muller, L.J.; Marfurt, C.F.; Kruse, F.; Tervo, T.M. Corneal nerves: Structure, contents and function. Exp. Eye Res. 2003, 76, 521–542. [Google Scholar] [CrossRef] [PubMed]
- Beuerman, R.W.; Schimmelpfennig, B. Sensory denervation of the rabbit cornea affects epithelial properties. Exp. Neurol. 1980, 69, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, I.N.; Alam, U.; Fadavi, H.; Asghar, O.; Green, P.; Ponirakis, G.; Marshall, A.; Boulton, A.J.; Tavakoli, M.; Malik, R.A. Corneal nerve loss detected with corneal confocal microscopy is symmetrical and related to the severity of diabetic polyneuropathy. Diabetes Care 2013, 36, 3646–3651. [Google Scholar] [CrossRef] [PubMed]
- Bikbova, G.; Oshitari, T.; Baba, T.; Bikbov, M.; Yamamoto, S. Diabetic corneal neuropathy: Clinical perspectives. Clin. Ophthalmol. 2018, 12, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Petropoulos, I.N.; Ponirakis, G.; Ferdousi, M.; Azmi, S.; Kalteniece, A.; Khan, A.; Gad, H.; Bashir, B.; Marshall, A.; Boulton, A.J.M.; et al. Corneal Confocal Microscopy: A Biomarker for Diabetic Peripheral Neuropathy. Clin. Ther. 2021, 43, 1457–1475. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.S.; Yin, J.; Lee, P.; Hwang, F.S.; McDermott, M. Sensory nerve regeneration after epithelium wounding in normal and diabetic cornea. Expert Rev. Ophthalmol. 2015, 10, 383–392. [Google Scholar] [CrossRef] [PubMed]
- Edwards, K.; Pritchard, N.; Poole, C.; Dehghani, C.; Al Rashah, K.; Russell, A.; Malik, R.A.; Efron, N. Development of a Novel Technique to Measure Corneal Nerve Migration Rate. Cornea 2016, 35, 700–705. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Pham, T.L.; Kakazu, A.; Bazan, H.E.P. Recovery of Corneal Sensitivity and Increase in Nerve Density and Wound Healing in Diabetic Mice after PEDF Plus DHA Treatment. Diabetes 2017, 66, 2511–2520. [Google Scholar] [CrossRef]
- Gao, N.; Yan, C.; Lee, P.; Sun, H.; Yu, F.S. Dendritic cell dysfunction and diabetic sensory neuropathy in the cornea. J. Clin. Investig. 2016, 126, 1998–2011. [Google Scholar] [CrossRef]
- Cousen, P.; Cackett, P.; Bennett, H.; Swa, K.; Dhillon, B. Tear production and corneal sensitivity in diabetes. J. Diabetes Complicat. 2007, 21, 371–373. [Google Scholar] [CrossRef]
- Rosenberg, M.E.; Tervo, T.M.; Immonen, I.J.; Muller, L.J.; Gronhagen-Riska, C.; Vesaluoma, M.H. Corneal structure and sensitivity in type 1 diabetes mellitus. Investig. Ophthalmol. Vis. Sci. 2000, 41, 2915–2921. [Google Scholar]
- Hamrah, P.; Seyed-Razavi, Y.; Yamaguchi, T. Translational Immunoimaging and Neuroimaging Demonstrate Corneal Neuroimmune Crosstalk. Cornea 2016, 35 (Suppl. S1), S20–S24. [Google Scholar] [CrossRef]
- Yu, F.X.; Lee, P.S.Y.; Yang, L.; Gao, N.; Zhang, Y.; Ljubimov, A.V.; Yang, E.; Zhou, Q.; Xie, L. The impact of sensory neuropathy and inflammation on epithelial wound healing in diabetic corneas. Prog. Retin. Eye Res. 2022, 89, 101039. [Google Scholar] [CrossRef]
- Liu, J.; Li, Z. Resident Innate Immune Cells in the Cornea. Front. Immunol. 2021, 12, 620284. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Hill, L.J.; Downie, L.E.; Chinnery, H.R. Neuroimmune crosstalk in the cornea: The role of immune cells in corneal nerve maintenance during homeostasis and inflammation. Prog. Retin. Eye Res. 2022, 91, 101105. [Google Scholar] [CrossRef] [PubMed]
- Wilson, S.E.; Sampaio, L.P.; Shiju, T.M.; de Oliveira, R.C. Fibroblastic and bone marrow-derived cellularity in the corneal stroma. Exp. Eye Res. 2021, 202, 108303. [Google Scholar] [CrossRef] [PubMed]
- Brissette-Storkus, C.S.; Reynolds, S.M.; Lepisto, A.J.; Hendricks, R.L. Identification of a novel macrophage population in the normal mouse corneal stroma. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2264–2271. [Google Scholar]
- Lassance, L.; Marino, G.K.; Medeiros, C.S.; Thangavadivel, S.; Wilson, S.E. Fibrocyte migration, differentiation and apoptosis during the corneal wound healing response to injury. Exp. Eye Res. 2018, 170, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chen, Z.; Lu, J.; Watsky, M.A. Effects of Topical 1,25 and 24,25 Vitamin D on Diabetic, Vitamin D Deficient and Vitamin D Receptor Knockout Mouse Corneal Wound Healing. Biomolecules 2023, 13, 1065. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Xue, Y.; Dong, D.; Xiao, C.; Lin, C.; Wang, H.; Song, F.; Fu, T.; Wang, Z.; Chen, J.; et al. CCR2(-) and CCR2(+) corneal macrophages exhibit distinct characteristics and balance inflammatory responses after epithelial abrasion. Mucosal Immunol. 2017, 10, 1145–1159. [Google Scholar] [CrossRef]
- Chinnery, H.R.; McMenamin, P.G.; Dando, S.J. Macrophage physiology in the eye. Pflugers Arch. 2017, 469, 501–515. [Google Scholar] [CrossRef]
- Lin, Y.; Ubels, J.L.; Schotanus, M.P.; Yin, Z.; Pintea, V.; Hammock, B.D.; Watsky, M.A. Enhancement of vitamin D metabolites in the eye following vitamin D3 supplementation and UV-B irradiation. Curr. Eye Res. 2012, 37, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, P.P.; Gao, X.H.; Tan, J.C.; Evdokiou, A.; Ferrante, A.; Morris, H.A.; May, B.K.; Hii, C.S. A role for the phosphatidylinositol 3-kinase–protein kinase C zeta–Sp1 pathway in the 1,25-dihydroxyvitamin D3 induction of the 25-hydroxyvitamin D3 24-hydroxylase gene in human kidney cells. Cell. Signal. 2010, 22, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.; Strugnell, S.A.; DeLuca, H.F. Current understanding of the molecular actions of vitamin D. Physiol. Rev. 1998, 78, 1193–1231. [Google Scholar] [CrossRef] [PubMed]
- Grammatiki, M.; Rapti, E.; Karras, S.; Ajjan, R.A.; Kotsa, K. Vitamin D and diabetes mellitus: Causal or casual association? Rev. Endocr. Metab. Disord. 2017, 18, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Norris, J.M.; Lee, H.S.; Frederiksen, B.; Erlund, I.; Uusitalo, U.; Yang, J.; Lernmark, A.; Simell, O.; Toppari, J.; Rewers, M.; et al. Plasma 25-Hydroxyvitamin D Concentration and Risk of Islet Autoimmunity. Diabetes 2018, 67, 146–154. [Google Scholar] [CrossRef]
- Van Belle, T.L.; Gysemans, C.; Mathieu, C. Vitamin D and diabetes: The odd couple. Trends Endocrinol. Metab. 2013, 24, 561–568. [Google Scholar] [CrossRef]
- Angellotti, E.; Pittas, A.G. The Role of Vitamin D in the Prevention of Type 2 Diabetes: To D or Not to D? Endocrinology 2017, 158, 2013–2021. [Google Scholar] [CrossRef]
- Lu, X.; Vick, S.; Chen, Z.; Chen, J.; Watsky, M.A. Effects of Vitamin D Receptor Knockout and Vitamin D Deficiency on Corneal Epithelial Wound Healing and Nerve Density in Diabetic Mice. Diabetes 2020, 69, 1042–1051. [Google Scholar] [CrossRef]
- Calik Basaran, N.; Kiragi, D.; Tan, C.; Ozisik, L.; Cagdas Ayvaz, N.D.; Kocabeyoglu, S.; Oz, S.G.; Irkec, M.; Tezcan, F.I. Ocular Changes and Tear Cytokines in Individuals with Low Serum Vitamin D Levels: A Cross-Sectional, Controlled Study. Ocul. Immunol. Inflamm. online ahead of print. 2023. [Google Scholar] [CrossRef]
- Garcion, E.; Wion-Barbot, N.; Montero-Menei, C.N.; Berger, F.; Wion, D. New clues about vitamin D functions in the nervous system. Trends Endocrinol. Metab. 2002, 13, 100–105. [Google Scholar] [CrossRef]
- Landel, V.; Annweiler, C.; Millet, P.; Morello, M.; Feron, F. Vitamin D, Cognition and Alzheimer’s Disease: The Therapeutic Benefit is in the D-Tails. J. Alzheimer’s Dis. 2016, 53, 419–444. [Google Scholar] [CrossRef]
- Lu, X.; Watsky, M.A. Influence of Vitamin D on Corneal Epithelial Cell Desmosomes and Hemidesmosomes. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4074–4083. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Chen, Z.; Watsky, M.A. Effects of 1,25 and 24,25 Vitamin D on Corneal Fibroblast VDR and Vitamin D Metabolizing and Catabolizing Enzymes. Curr. Eye Res. 2021, 46, 1271–1282. [Google Scholar] [CrossRef]
- Yamakawa, I.; Kojima, H.; Terashima, T.; Katagi, M.; Oi, J.; Urabe, H.; Sanada, M.; Kawai, H.; Chan, L.; Yasuda, H.; et al. Inactivation of TNF-alpha ameliorates diabetic neuropathy in mice. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E844–E852. [Google Scholar] [CrossRef] [PubMed]
- Kollenkirchen, U.; Fox, J.; Walters, M.R. Normocalcemia without Hyperparathyroidism in Vitamin-D-Deficient Rats. J. Bone Miner. Res. 1991, 6, 273–278. [Google Scholar] [CrossRef]
- Lu, X.; Chen, Z.; Vick, S.; Watsky, M.A. Vitamin D receptor and metabolite effects on corneal epithelial cell gap junction proteins. Exp. Eye Res. 2019, 187, 107776. [Google Scholar] [CrossRef]
- Pham, T.L.; Kakazu, A.; He, J.; Bazan, H.E.P. Mouse strains and sexual divergence in corneal innervation and nerve regeneration. FASEB J. 2019, 33, 4598–4609. [Google Scholar] [CrossRef]
- Jiao, H.; Naranjo Golborne, C.; Dando, S.J.; McMenamin, P.G.; Downie, L.E.; Chinnery, H.R. Topographical and Morphological Differences of Corneal Dendritic Cells during Steady State and Inflammation. Ocul. Immunol. Inflamm. 2020, 28, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Yamagami, S.; Yokoo, S.; Usui, T.; Yamagami, H.; Amano, S.; Ebihara, N. Distinct populations of dendritic cells in the normal human donor corneal epithelium. Investig. Ophthalmol. Vis. Sci. 2005, 46, 4489–4494. [Google Scholar] [CrossRef]
- Lu, X.; Chen, Z.; Mylarapu, N.; Watsky, M.A. Effects of 1,25 and 24,25 Vitamin D on Corneal Epithelial Proliferation, Migration and Vitamin D Metabolizing and Catabolizing Enzymes. Sci. Rep. 2017, 7, 16951. [Google Scholar] [CrossRef]
- Yin, Z.; Pintea, V.; Lin, Y.; Hammock, B.D.; Watsky, M.A. Vitamin D enhances corneal epithelial barrier function. Investig. Ophthalmol. Vis. Sci. 2011, 52, 7359–7364. [Google Scholar] [CrossRef]
- Reinstein, D.Z.; Archer, T.J.; Gobbe, M.; Bartoli, E. Corneal sensitivity after small-incision lenticule extraction and laser in situ keratomileusis. J. Cataract. Refract. Surg. 2015, 41, 1580–1587. [Google Scholar] [CrossRef]
- Medeiros, C.S.; Santhiago, M.R. Corneal nerves anatomy, function, injury and regeneration. Exp. Eye Res. 2020, 200, 108243. [Google Scholar] [CrossRef] [PubMed]
- Ishii, R.; Shimizu, K.; Igarashi, A.; Kobashi, H.; Kamiya, K. Influence of femtosecond lenticule extraction and small incision lenticule extraction on corneal nerve density and ocular surface: A 1-year prospective, confocal, microscopic study. J. Refract. Surg. 2015, 31, 10–15. [Google Scholar] [CrossRef]
- Filipovic, N.; Ferhatovic, L.; Marelja, I.; Puljak, L.; Grkovic, I. Increased vitamin D receptor expression in dorsal root ganglia neurons of diabetic rats. Neurosci. Lett. 2013, 549, 140–145. [Google Scholar] [CrossRef]
- Naveilhan, P.; Neveu, I.; Wion, D.; Brachet, P. 1,25-Dihydroxyvitamin D3, an inducer of glial cell line-derived neurotrophic factor. Neuroreport 1996, 7, 2171–2175. [Google Scholar] [CrossRef]
- Wion, D.; MacGrogan, D.; Neveu, I.; Jehan, F.; Houlgatte, R.; Brachet, P. 1,25-Dihydroxyvitamin D3 is a potent inducer of nerve growth factor synthesis. J. Neurosci. Res. 1991, 28, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Eyles, D.; Brown, J.; Mackay-Sim, A.; McGrath, J.; Feron, F. Vitamin D3 and brain development. Neuroscience 2003, 118, 641–653. [Google Scholar] [CrossRef]
- Bouillon, R.; Carmeliet, G.; Verlinden, L.; van Etten, E.; Verstuyf, A.; Luderer, H.F.; Lieben, L.; Mathieu, C.; Demay, M. Vitamin D and human health: Lessons from vitamin D receptor null mice. Endocr. Rev. 2008, 29, 726–776. [Google Scholar] [CrossRef]
- Feron, F.; Burne, T.H.; Brown, J.; Smith, E.; McGrath, J.J.; Mackay-Sim, A.; Eyles, D.W. Developmental Vitamin D3 deficiency alters the adult rat brain. Brain Res. Bull. 2005, 65, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Hellweg, R.; Hartung, H.D. Endogenous levels of nerve growth factor (NGF) are altered in experimental diabetes mellitus: A possible role for NGF in the pathogenesis of diabetic neuropathy. J. Neurosci. Res. 1990, 26, 258–267. [Google Scholar] [CrossRef]
- Fernyhough, P.; Diemel, L.T.; Brewster, W.J.; Tomlinson, D.R. Altered neurotrophin mRNA levels in peripheral nerve and skeletal muscle of experimentally diabetic rats. J. Neurochem. 1995, 64, 1231–1237. [Google Scholar] [CrossRef]
- Riaz, S.; Malcangio, M.; Miller, M.; Tomlinson, D.R. A vitamin D(3) derivative (CB1093) induces nerve growth factor and prevents neurotrophic deficits in streptozotocin-diabetic rats. Diabetologia 1999, 42, 1308–1313. [Google Scholar] [CrossRef]
- Charoenngam, N.; Holick, M.F. Immunologic Effects of Vitamin D on Human Health and Disease. Nutrients 2020, 12, 2097. [Google Scholar] [CrossRef]
- Chen, P.; Piao, X.; Bonaldo, P. Role of macrophages in Wallerian degeneration and axonal regeneration after peripheral nerve injury. Acta Neuropathol. 2015, 130, 605–618. [Google Scholar] [CrossRef]
- Sosnova, M.; Bradl, M.; Forrester, J.V. CD34+ corneal stromal cells are bone marrow-derived and express hemopoietic stem cell markers. Stem Cells 2005, 23, 507–515. [Google Scholar] [CrossRef]
- Chinnery, H.R.; Ruitenberg, M.J.; Plant, G.W.; Pearlman, E.; Jung, S.; McMenamin, P.G. The chemokine receptor CX3CR1 mediates homing of MHC class II-positive cells to the normal mouse corneal epithelium. Investig. Ophthalmol. Vis. Sci. 2007, 48, 1568–1574. [Google Scholar] [CrossRef]
- Liu, J.; Wu, M.; He, J.; Xiao, C.; Xue, Y.; Fu, T.; Lin, C.; Dong, D.; Li, Z. Antibiotic-Induced Dysbiosis of Gut Microbiota Impairs Corneal Nerve Regeneration by Affecting CCR2-Negative Macrophage Distribution. Am. J. Pathol. 2018, 188, 2786–2799. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Du, S.; Yu, J.; Yang, X.; Yang, C.; Zhou, D.; Wang, Q.; Qin, S.; Yan, X.; He, L.; et al. Common housekeeping proteins are upregulated in colorectal adenocarcinoma and hepatocellular carcinoma, making the total protein a better “housekeeper”. Oncotarget 2016, 7, 66679–66688. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Groups | Abbreviation | |
|---|---|---|
| Normoglycemic | Wild type | CTRL |
| Vit D receptor knockout | VDR KO | |
| Vit D receptor knockout on supplemental diet | VDR KO Ca++ | |
| Vitamin D deficient | VDD | |
| Diabetic | Wild type | Dia CTRL |
| Vit D receptor knockout | Dia VDR KO | |
| Vit D receptor knockout on supplemental diet | Dia VDR KO Ca++ | |
| Vitamin D deficient | Dia VDD | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Chen, Z.; Lu, J.; Watsky, M.A. Effects of 1,25-Vitamin D3 and 24,25-Vitamin D3 on Corneal Nerve Regeneration in Diabetic Mice. Biomolecules 2023, 13, 1754. https://doi.org/10.3390/biom13121754
Lu X, Chen Z, Lu J, Watsky MA. Effects of 1,25-Vitamin D3 and 24,25-Vitamin D3 on Corneal Nerve Regeneration in Diabetic Mice. Biomolecules. 2023; 13(12):1754. https://doi.org/10.3390/biom13121754
Chicago/Turabian StyleLu, Xiaowen, Zhong Chen, Jerry Lu, and Mitchell A. Watsky. 2023. "Effects of 1,25-Vitamin D3 and 24,25-Vitamin D3 on Corneal Nerve Regeneration in Diabetic Mice" Biomolecules 13, no. 12: 1754. https://doi.org/10.3390/biom13121754
APA StyleLu, X., Chen, Z., Lu, J., & Watsky, M. A. (2023). Effects of 1,25-Vitamin D3 and 24,25-Vitamin D3 on Corneal Nerve Regeneration in Diabetic Mice. Biomolecules, 13(12), 1754. https://doi.org/10.3390/biom13121754