


Anthocyanins: Molecular Aspects on Their Neuroprotective Activity


 and
and 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Anthocyanins
2.1. Dietary Sources
2.2. Chemistry and Its Relationship with Bioavailability and Biodistribution
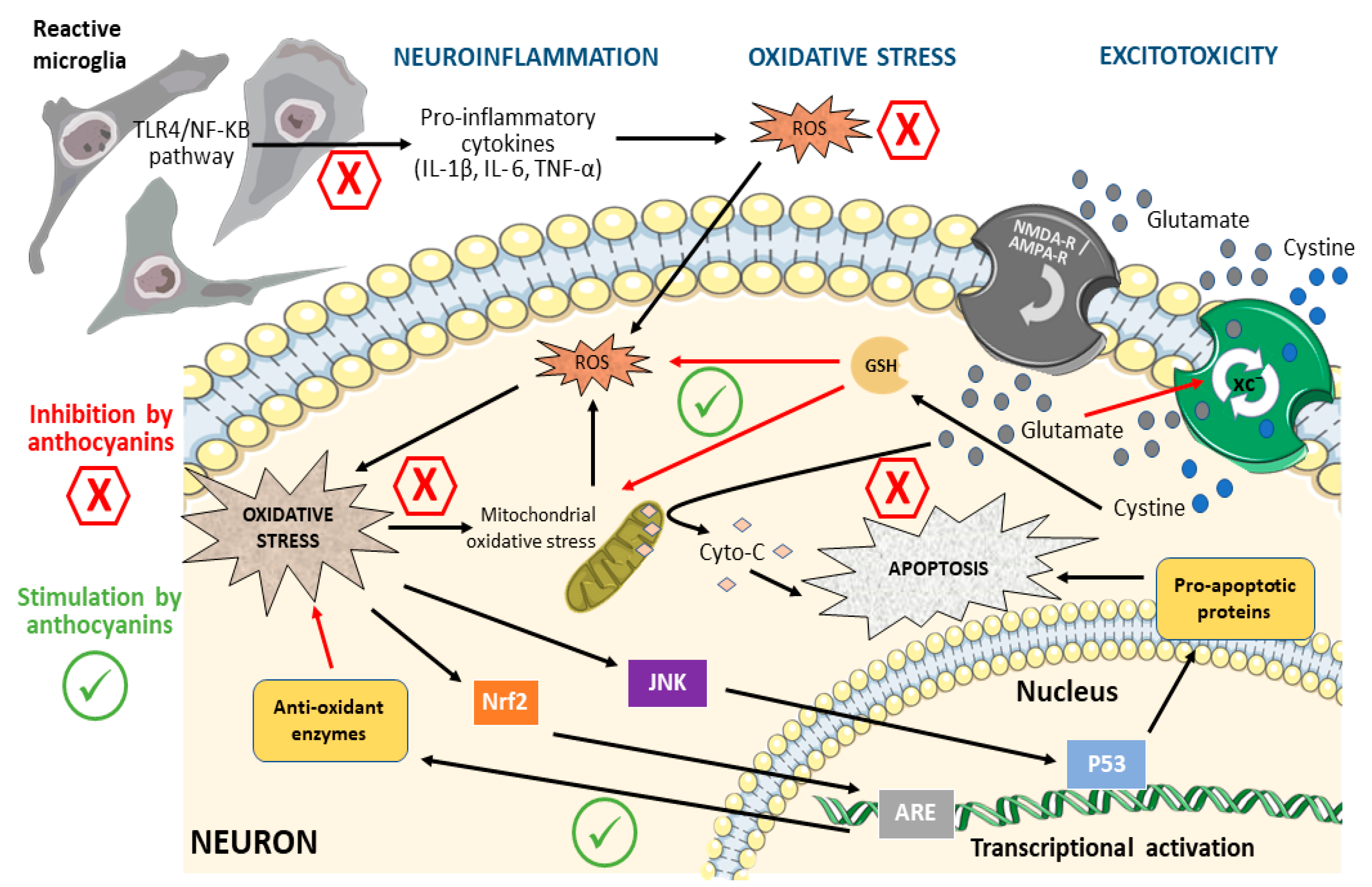
3. Mechanisms of Neuronal Damage Affected by Anthocyanins
3.1. Oxidative Stress
3.2. Excitotoxicity
3.3. Neuroinflammation
3.4. Altered Cholinergic Transmission

4. Effect of Anthocyanins on Neurological Disorders
4.1. Alzheimer’s Disease
4.2. Parkinson’s Disease (PD)
4.3. Hypoxia/Cerebral Ischemia
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Naomi, R.; Yazid, M.D.; Teoh, S.H.; Balan, S.S.; Shariff, H.; Kumar, J.; Bahari, H.; Embong, H. Dietary Polyphenols as a Protection against Cognitive Decline: Evidence from Animal Experiments; Mechanisms and Limitations. Antioxidants 2023, 12, 1054. [Google Scholar] [CrossRef] [PubMed]
- Graf, B.A.; Milbury, P.E.; Blumberg, J.B. Flavonols, Flavones, Flavanones, and Human Health: Epidemiological Evidence. J. Med. Food 2005, 8, 281–290. [Google Scholar] [CrossRef]
- van Westen, G.J.P.; Gaulton, A.; Overington, J.P. Chemical, Target, and Bioactive Properties of Allosteric Modulation. PLoS Comput. Biol. 2014, 10, e1003559. [Google Scholar] [CrossRef] [PubMed]
- Middleton, E.; Kandaswami, C.; Theoharides, T.C. The Effects of Plant Flavonoids on Mammalian Cells: Implications for Inflammation, Heart Disease, and Cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar]
- Cheynier, V. Polyphenols in Foods Are More Complex than Often Thought. Am. J. Clin. Nutr. 2005, 81, 223S–229S. [Google Scholar] [CrossRef]
- Tsao, R. Chemistry and Biochemistry of Dietary Polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef]
- Hardman, W.E. Diet Components Can Suppress Inflammation and Reduce Cancer Risk. Nutr. Res. Pract. 2014, 8, 233. [Google Scholar] [CrossRef]
- Truzzi, F.; Tibaldi, C.; Zhang, Y.; Dinelli, G.; D’Amen, E. An Overview on Dietary Polyphenols and Their Biopharmaceutical Classification System (BCS). Int. J. Mol. Sci. 2021, 22, 5514. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef] [PubMed]
- Riaz, M.; Zia-Ul-Haq, M.; Saad, B. Diet and Herbal-Derived Medicines; Springer: Berlin/Heidelberg, Germany, 2016; pp. 1–19. [Google Scholar]
- Veitch, N.C.; Grayer, R.J. Flavonoids and Their Glycosides, Including Anthocyanins. Nat. Prod. Rep. 2008, 25, 555. [Google Scholar] [CrossRef] [PubMed]
- Prior, R.L.; Wu, X. Anthocyanins: Structural Characteristics That Result in Unique Metabolic Patterns and Biological Activities. Free Radic. Res. 2006, 40, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Choi, M.; Lee, M. Effects of Anthocyanin Supplementation on Reduction of Obesity Criteria: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2021, 13, 2121. [Google Scholar] [CrossRef] [PubMed]
- Fallah, A.A.; Sarmast, E.; Fatehi, P.; Jafari, T. Impact of Dietary Anthocyanins on Systemic and Vascular Inflammation: Systematic Review and Meta-Analysis on Randomised Clinical Trials. Food Chem. Toxicol. 2020, 135, 110922. [Google Scholar] [CrossRef] [PubMed]
- Nikbakht, E.; Singh, I.; Vider, J.; Williams, L.T.; Vugic, L.; Gaiz, A.; Kundur, A.R.; Colson, N. Potential of Anthocyanin as an Anti-Inflammatory Agent: A Human Clinical Trial on Type 2 Diabetic, Diabetic at-Risk and Healthy Adults. Inflamm. Res. 2021, 70, 275–284. [Google Scholar] [CrossRef]
- Whyte, A.; Cheng, N.; Fromentin, E.; Williams, C. A Randomized, Double-Blinded, Placebo-Controlled Study to Compare the Safety and Efficacy of Low Dose Enhanced Wild Blueberry Powder and Wild Blueberry Extract (ThinkBlueTM) in Maintenance of Episodic and Working Memory in Older Adults. Nutrients 2018, 10, 660. [Google Scholar] [CrossRef]
- Boespflug, E.L.; Eliassen, J.C.; Dudley, J.A.; Shidler, M.D.; Kalt, W.; Summer, S.S.; Stein, A.L.; Stover, A.N.; Krikorian, R. Enhanced Neural Activation with Blueberry Supplementation in Mild Cognitive Impairment. Nutr. Neurosci. 2018, 21, 297–305. [Google Scholar] [CrossRef]
- Krikorian, R.; Skelton, M.R.; Summer, S.S.; Shidler, M.D.; Sullivan, P.G. Blueberry Supplementation in Midlife for Dementia Risk Reduction. Nutrients 2022, 14, 1619. [Google Scholar] [CrossRef]
- Feng, R.C.; Dong, Y.H.; Hong, X.L.; Su, Y.; Wu, X.V. Effects of Anthocyanin-Rich Supplementation on Cognition of the Cognitively Healthy Middle-Aged and Older Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutr. Rev. 2023, 81, 287–303. [Google Scholar] [CrossRef]
- Lopresti, A.L.; Smith, S.J.; Pouchieu, C.; Pourtau, L.; Gaudout, D.; Pallet, V.; Drummond, P.D. Effects of a Polyphenol-Rich Grape and Blueberry Extract (MemophenolTM) on Cognitive Function in Older Adults with Mild Cognitive Impairment: A Randomized, Double-Blind, Placebo-Controlled Study. Front. Psychol. 2023, 14, 1144231. [Google Scholar] [CrossRef] [PubMed]
- Wood, E.; Hein, S.; Mesnage, R.; Fernandes, F.; Abhayaratne, N.; Xu, Y.; Zhang, Z.; Bell, L.; Williams, C.; Rodriguez-Mateos, A. Wild Blueberry (Poly)Phenols Can Improve Vascular Function and Cognitive Performance in Healthy Older Individuals: A Double-Blind Randomized Controlled Trial. Am. J. Clin. Nutr. 2023, 117, 1306–1319. [Google Scholar] [CrossRef]
- Barfoot, K.L.; May, G.; Lamport, D.J.; Ricketts, J.; Riddell, P.M.; Williams, C.M. The Effects of Acute Wild Blueberry Supplementation on the Cognition of 7–10-Year-Old Schoolchildren. Eur. J. Nutr. 2019, 58, 2911–2920. [Google Scholar] [CrossRef] [PubMed]
- Bowtell, J.L.; Aboo-Bakkar, Z.; Conway, M.E.; Adlam, A.-L.R.; Fulford, J. Enhanced Task-Related Brain Activation and Resting Perfusion in Healthy Older Adults after Chronic Blueberry Supplementation. Appl. Physiol. Nutr. Metab. 2017, 42, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Ahles, S.; Joris, P.J.; Plat, J. Effects of Berry Anthocyanins on Cognitive Performance, Vascular Function and Cardiometabolic Risk Markers: A Systematic Review of Randomized Placebo-Controlled Intervention Studies in Humans. Int. J. Mol. Sci. 2021, 22, 6482. [Google Scholar] [CrossRef] [PubMed]
- Ullah, R.; Khan, M.; Shah, S.A.; Saeed, K.; Kim, M.O. Natural Antioxidant Anthocyanins—A Hidden Therapeutic Candidate in Metabolic Disorders with Major Focus in Neurodegeneration. Nutrients 2019, 11, 1195. [Google Scholar] [CrossRef]
- Andersen, Ø.M.; Jordheim, M. Anthocyanins. In Encyclopedia of Life Sciences; Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar]
- Shipp, J.; Abdel-Aal, E.-S.M. Food Applications and Physiological Effects of Anthocyanins as Functional Food Ingredients. Open Food Sci. J. 2010, 4, 7–22. [Google Scholar] [CrossRef]
- Valls, J.; Millán, S.; Martí, M.P.; Borràs, E.; Arola, L. Advanced Separation Methods of Food Anthocyanins, Isoflavones and Flavanols. J. Chromatogr. A 2009, 1216, 7143–7172. [Google Scholar] [CrossRef]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A Comprehensive Review of Their Chemical Properties and Health Effects on Cardiovascular and Neurodegenerative Diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and Anthocyanins: Colored Pigments as Food, Pharmaceutical Ingredients, and the Potential Health Benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Giuliani, A.; Cerretani, L.; Cichelli, A. Colors: Properties and Determination of Natural Pigments. In Encyclopedia of Food and Health; Elsevier: Amsterdam, The Netherlands, 2016; pp. 273–283. [Google Scholar]
- Welch, C.R.; Wu, Q.; Simon, J.E. Recent Advances in Anthocyanin Analysis and Characterization. Curr. Anal. Chem. 2008, 4, 75–101. [Google Scholar] [CrossRef] [PubMed]
- Kamonpatana, K.; Failla, M.L.; Kumar, P.S.; Giusti, M.M. Anthocyanin Structure Determines Susceptibility to Microbial Degradation and Bioavailability to the Buccal Mucosa. J. Agric. Food Chem. 2014, 62, 6903–6910. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Tan, Y.; Chen, G.; Wang, G.; Sun, J.; Ou, S.; Chen, W.; Bai, W. Metabolism of Anthocyanins and Consequent Effects on the Gut Microbiota. Crit. Rev. Food Sci. Nutr. 2019, 59, 982–991. [Google Scholar] [CrossRef] [PubMed]
- Hahm, T.H.; Tanaka, M.; Matsui, T. Current Knowledge on Intestinal Absorption of Anthocyanins. J. Agric. Food Chem. 2022, 70, 2501–2509. [Google Scholar] [CrossRef]
- Kalt, W. Anthocyanins and Their C6-C3-C6 Metabolites in Humans and Animals. Molecules 2019, 24, 4024. [Google Scholar] [CrossRef]
- Milbury, P.E.; Cao, G.; Prior, R.L.; Blumberg, J. Bioavailablility of Elderberry Anthocyanins. Mech. Ageing Dev. 2002, 123, 997–1006. [Google Scholar] [CrossRef]
- Felgines, C.; Texier, O.; Besson, C.; Fraisse, D.; Lamaison, J.-L.; Rémésy, C. Blackberry Anthocyanins Are Slightly Bioavailable in Rats. J. Nutr. 2002, 132, 1249–1253. [Google Scholar] [CrossRef][Green Version]
- Passamonti, S.; Vrhovsek, U.; Vanzo, A.; Mattivi, F. The Stomach as a Site for Anthocyanins Absorption from Food. FEBS Lett. 2003, 544, 210–213. [Google Scholar] [CrossRef]
- Talavéra, S.; Felgines, C.; Texier, O.; Besson, C.; Manach, C.; Lamaison, J.-L.; Rémésy, C. Anthocyanins Are Efficiently Absorbed from the Small Intestine in Rats. J. Nutr. 2004, 134, 2275–2279. [Google Scholar] [CrossRef]
- Miyazawa, T.; Nakagawa, K.; Kudo, M.; Muraishi, K.; Someya, K. Direct Intestinal Absorption of Red Fruit Anthocyanins, Cyanidin-3-Glucoside and Cyanidin-3,5-Diglucoside, into Rats and Humans. J. Agric. Food Chem. 1999, 47, 1083–1091. [Google Scholar] [CrossRef]
- Milbury, P.E.; Kalt, W. Xenobiotic Metabolism and Berry Flavonoid Transport across the Blood-Brain Barrier. J. Agric. Food Chem. 2010, 58, 3950–3956. [Google Scholar] [CrossRef] [PubMed]
- McGhie, T.K.; Ainge, G.D.; Barnett, L.E.; Cooney, J.M.; Jensen, D.J. Anthocyanin Glycosides from Berry Fruit Are Absorbed and Excreted Unmetabolized by Both Humans and Rats. J. Agric. Food Chem. 2003, 51, 4539–4548. [Google Scholar] [CrossRef] [PubMed]
- El Mohsen, M.A.; Marks, J.; Kuhnle, G.; Moore, K.; Debnam, E.; Srai, S.K.; Rice-Evans, C.; Spencer, J.P.E. Absorption, Tissue Distribution and Excretion of Pelargonidin and Its Metabolites Following Oral Administration to Rats. Br. J. Nutr. 2006, 95, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.M.; El Mohsen, M.A.; Vauzour, D.; Rendeiro, C.; Butler, L.T.; Ellis, J.A.; Whiteman, M.; Spencer, J.P.E. Blueberry-Induced Changes in Spatial Working Memory Correlate with Changes in Hippocampal CREB Phosphorylation and Brain-Derived Neurotrophic Factor (BDNF) Levels. Free Radic. Biol. Med. 2008, 45, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Youdim, K.A.; Dobbie, M.S.; Kuhnle, G.; Proteggente, A.R.; Abbott, N.J.; Rice-Evans, C. Interaction between Flavonoids and the Blood-Brain Barrier: In Vitro Studies. J. Neurochem. 2003, 85, 180–192. [Google Scholar] [CrossRef]
- Fornasaro, S.; Ziberna, L.; Gasperotti, M.; Tramer, F.; Vrhovšek, U.; Mattivi, F.; Passamonti, S. Determination of Cyanidin 3-Glucoside in Rat Brain, Liver and Kidneys by UPLC/MS-MS and Its Application to a Short-Term Pharmacokinetic Study. Sci. Rep. 2016, 6, 22815. [Google Scholar] [CrossRef]
- Andres-Lacueva, C.; Shukitt-Hale, B.; Galli, R.L.; Jauregui, O.; Lamuela-Raventos, R.M.; Joseph, J.A. Anthocyanins in Aged Blueberry-Fed Rats Are Found Centrally and May Enhance Memory. Nutr. Neurosci. 2005, 8, 111–120. [Google Scholar] [CrossRef]
- Muddapu, V.R.; Dharshini, S.A.P.; Chakravarthy, V.S.; Gromiha, M.M. Neurodegenerative Diseases–Is Metabolic Deficiency the Root Cause? Front. Neurosci. 2020, 14, 213. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Orlacchio, A.; Maccarrone, M. Is Modulation of Oxidative Stress an Answer? The State of the Art of Redox Therapeutic Actions in Neurodegenerative Diseases. Oxid. Med. Cell Longev. 2016, 2016, 7909380. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Barber, A.J.; Spagnuolo, C.; Russo, G.L.; Daglia, M.; Nabavi, S.M.; Sobarzo-Sánchez, E. Nrf2 as Molecular Target for Polyphenols: A Novel Therapeutic Strategy in Diabetic Retinopathy. Crit. Rev. Clin. Lab. Sci. 2016, 53, 293–312. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary Intake and Bioavailability of Polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [PubMed]
- Brigelius-Flohé, R. Commentary: Oxidative Stress Reconsidered. Genes. Nutr. 2009, 4, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef]
- Basli, A.; Soulet, S.; Chaher, N.; Mérillon, J.-M.; Chibane, M.; Monti, J.-P.; Richard, T. Wine Polyphenols: Potential Agents in Neuroprotection. Oxid. Med. Cell Longev. 2012, 2012, 805762. [Google Scholar] [CrossRef]
- Barnham, K.J.; Masters, C.L.; Bush, A.I. Neurodegenerative Diseases and Oxidative Stress. Nat. Rev. Drug Discov. 2004, 3, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Caspersen, C.; Wang, N.; Yao, J.; Sosunov, A.; Chen, X.; Lustbader, J.W.; Xu, H.W.; Stern, D.; McKhann, G.; Yan, S. Du Mitochondrial Abeta: A Potential Focal Point for Neuronal Metabolic Dysfunction in Alzheimer’s Disease. FASEB J. 2005, 19, 2040–2041. [Google Scholar] [CrossRef]
- Casley, C.S.; Canevari, L.; Land, J.M.; Clark, J.B.; Sharpe, M.A. Beta-Amyloid Inhibits Integrated Mitochondrial Respiration and Key Enzyme Activities. J. Neurochem. 2002, 80, 91–100. [Google Scholar] [CrossRef]
- Canevari, L.; Clark, J.B.; Bates, T.E. Beta-Amyloid Fragment 25-35 Selectively Decreases Complex IV Activity in Isolated Mitochondria. FEBS Lett. 1999, 457, 131–134. [Google Scholar] [CrossRef]
- Abramov, A.Y.; Smulders-Srinivasan, T.K.; Kirby, D.M.; Acin-Perez, R.; Enriquez, J.A.; Lightowlers, R.N.; Duchen, M.R.; Turnbull, D.M. Mechanism of Neurodegeneration of Neurons with Mitochondrial DNA Mutations. Brain 2010, 133, 797–807. [Google Scholar] [CrossRef]
- Solleiro-Villavicencio, H.; Rivas-Arancibia, S. Effect of Chronic Oxidative Stress on Neuroinflammatory Response Mediated by CD4+T Cells in Neurodegenerative Diseases. Front. Cell Neurosci. 2018, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wuliji, O.; Li, W.; Jiang, Z.-G.; Ghanbari, H.A. Oxidative Stress and Neurodegenerative Disorders. Int. J. Mol. Sci. 2013, 14, 24438–24475. [Google Scholar] [CrossRef] [PubMed]
- Kishida, K.T.; Klann, E. Sources and Targets of Reactive Oxygen Species in Synaptic Plasticity and Memory. Antioxid. Redox Signal 2006, 9, 233–244. [Google Scholar] [CrossRef]
- Rahman, M.H.; Bajgai, J.; Fadriquela, A.; Sharma, S.; Trinh, T.T.; Akter, R.; Jeong, Y.J.; Goh, S.H.; Kim, C.-S.; Lee, K.-J. Therapeutic Potential of Natural Products in Treating Neurodegenerative Disorders and Their Future Prospects and Challenges. Molecules 2021, 26, 5327. [Google Scholar] [CrossRef] [PubMed]
- Stote, K.S.; Burns, G.; Mears, K.; Sweeney, M.; Blanton, C. The Effect of Berry Consumption on Oxidative Stress Biomarkers: A Systematic Review of Randomized Controlled Trials in Humans. Antioxidants 2023, 12, 1443. [Google Scholar] [CrossRef]
- Winter, A.N.; Bickford, P.C. Anthocyanins and Their Metabolites as Therapeutic Agents for Neurodegenerative Disease. Antioxidants 2019, 8, 333. [Google Scholar] [CrossRef]
- Tarozzi, A.; Morroni, F.; Hrelia, S.; Angeloni, C.; Marchesi, A.; Cantelli-Forti, G.; Hrelia, P. Neuroprotective Effects of Anthocyanins and Their in Vivo Metabolites in SH-SY5Y Cells. Neurosci. Lett. 2007, 424, 36–40. [Google Scholar] [CrossRef]
- Shah, S.A.; Yoon, G.H.; Kim, M.O. Protection of the Developing Brain with Anthocyanins Against Ethanol-Induced Oxidative Stress and Neurodegeneration. Mol. Neurobiol. 2015, 51, 1278–1291. [Google Scholar] [CrossRef]
- Bhuiyan, M.I.H.; Kim, H.-B.; Kim, S.Y.; Cho, K.-O. The Neuroprotective Potential of Cyanidin-3-Glucoside Fraction Extracted from Mulberry Following Oxygen-Glucose Deprivation. Korean J. Physiol. Pharmacol. 2011, 15, 353–361. [Google Scholar] [CrossRef]
- Ye, J.; Meng, X.; Yan, C.; Wang, C. Effect of Purple Sweet Potato Anthocyanins on β-Amyloid-Mediated PC-12 Cells Death by Inhibition of Oxidative Stress. Neurochem. Res. 2010, 35, 357–365. [Google Scholar] [CrossRef]
- Kelsey, N.; Hulick, W.; Winter, A.; Ross, E.; Linseman, D. Neuroprotective Effects of Anthocyanins on Apoptosis Induced by Mitochondrial Oxidative Stress. Nutr. Neurosci. 2011, 14, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Kensler, T.W.; Wakabayashi, N.; Biswal, S. Cell Survival Responses to Environmental Stresses Via the Keap1-Nrf2-ARE Pathway. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 89–116. [Google Scholar] [CrossRef] [PubMed]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The Keap1-Nrf2 Pathway: Mechanisms of Activation and Dysregulation in Cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef]
- Zgorzynska, E.; Dziedzic, B.; Walczewska, A. An Overview of the Nrf2/ARE Pathway and Its Role in Neurodegenerative Diseases. Int. J. Mol. Sci. 2021, 22, 9592. [Google Scholar] [CrossRef]
- Thummayot, S.; Tocharus, C.; Jumnongprakhon, P.; Suksamrarn, A.; Tocharus, J. Cyanidin Attenuates Aβ25-35-Induced Neuroinflammation by Suppressing NF-ΚB Activity Downstream of TLR4/NOX4 in Human Neuroblastoma Cells. Acta Pharmacol. Sin. 2018, 39, 1439–1452. [Google Scholar] [CrossRef]
- Kropat, C.; Mueller, D.; Boettler, U.; Zimmermann, K.; Heiss, E.H.; Dirsch, V.M.; Rogoll, D.; Melcher, R.; Richling, E.; Marko, D. Modulation of Nrf2-Dependent Gene Transcription by Bilberry Anthocyanins in Vivo. Mol. Nutr. Food Res. 2013, 57, 545–550. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Bielinski, D.F.; Carey, A.; Schauss, A.G.; Shukitt-Hale, B. Modulation of Oxidative Stress, Inflammation, Autophagy and Expression of Nrf2 in Hippocampus and Frontal Cortex of Rats Fed with Açaí-Enriched Diets. Nutr. Neurosci. 2017, 20, 305–315. [Google Scholar] [CrossRef]
- Sukprasansap, M.; Chanvorachote, P.; Tencomnao, T. Cyanidin-3-Glucoside Activates Nrf2-Antioxidant Response Element and Protects against Glutamate-Induced Oxidative and Endoplasmic Reticulum Stress in HT22 Hippocampal Neuronal Cells. BMC Complement. Med. Ther. 2020, 20, 46. [Google Scholar] [CrossRef]
- Zhou, L.; Chang, J.; Zhao, W.; Gao, Y. Proanthocyanidins Regulate the Nrf2/ARE Signaling Pathway and Protect Neurons from Cypermethrin-Induced Oxidative Stress and Apoptosis. Pestic. Biochem. Physiol. 2021, 177, 104898. [Google Scholar] [CrossRef]
- Dong, X.; Wang, Y.; Qin, Z. Molecular Mechanisms of Excitotoxicity and Their Relevance to Pathogenesis of Neurodegenerative Diseases. Acta Pharmacol. Sin. 2009, 30, 379–387. [Google Scholar] [CrossRef]
- Ren, J.-X.; Li, C.; Yan, X.-L.; Qu, Y.; Yang, Y.; Guo, Z.-N. Crosstalk between Oxidative Stress and Ferroptosis/Oxytosis in Ischemic Stroke: Possible Targets and Molecular Mechanisms. Oxid. Med. Cell Longev. 2021, 2021, 6643382. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Schubert, D.; Maher, P. Oxytosis: A Novel Form of Programmed Cell Death. Curr. Top. Med. Chem. 2001, 1, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Lewerenz, J.; Ates, G.; Methner, A.; Conrad, M.; Maher, P. Oxytosis/Ferroptosis-(Re-) Emerging Roles for Oxidative Stress-Dependent Non-Apoptotic Cell Death in Diseases of the Central Nervous System. Front. Neurosci. 2018, 12, 214. [Google Scholar] [CrossRef] [PubMed]
- Murphy, T.H.; Miyamoto, M.; Sastre, A.; Schnaar, R.L.; Coyle, J.T. Glutamate Toxicity in a Neuronal Cell Line Involves Inhibition of Cystine Transport Leading to Oxidative Stress. Neuron 1989, 2, 1547–1558. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Tiziani, S.; Park, G.; Kaul, M.; Paternostro, G. Cellular Protection Using Flt3 and PI3Kα Inhibitors Demonstrates Multiple Mechanisms of Oxidative Glutamate Toxicity. Nat. Commun. 2014, 5, 3672. [Google Scholar] [CrossRef]
- Yang, W.S.; Stockwell, B.R. Ferroptosis: Death by Lipid Peroxidation. Trends Cell Biol. 2016, 26, 165–176. [Google Scholar] [CrossRef]
- Berliocchi, L.; Bano, D.; Nicotera, P. Ca 2+ Signals and Death Programmes in Neurons. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 2255–2258. [Google Scholar] [CrossRef]
- Froissard, P.; Duval, D. Cytotoxic Effects of Glutamic Acid on PC12 Cells. Neurochem. Int. 1994, 24, 485–493. [Google Scholar] [CrossRef]
- Zhang, Y.; Bhavnani, B.R. Glutamate-Induced Apoptosis in Neuronal Cells Is Mediated via Caspase-Dependent and Independent Mechanisms Involving Calpain and Caspase-3 Proteases as Well as Apoptosis Inducing Factor (AIF) and This Process Is Inhibited by Equine Estrogens. BMC Neurosci. 2006, 7, 49. [Google Scholar] [CrossRef]
- Lewerenz, J.; Klein, M.; Methner, A. Cooperative Action of Glutamate Transporters and Cystine/Glutamate Antiporter System Xc- Protects from Oxidative Glutamate Toxicity. J. Neurochem. 2006, 98, 916–925. [Google Scholar] [CrossRef]
- Bleakman, D.; Lodge, D. Neuropharmacology of AMPA and Kainate Receptors. Neuropharmacology 1998, 37, 1187–1204. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Park, H.Y.; Kim, M.O. Anthocyanins Protect against Kainic Acid-Induced Excitotoxicity and Apoptosis via ROS-Activated AMPK Pathway in Hippocampal Neurons. CNS Neurosci. Ther. 2014, 20, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Vignes, M.; Clarke, V.R.J.; Parry, M.J.; Bleakman, D.; Lodge, D.; Ornstein, P.L.; Collingridge, G.L. The GluR5 Subtype of Kainate Receptor Regulates Excitatory Synaptic Transmission in Areas CA1 and CA3 of the Rat Hippocampus. Neuropharmacology 1998, 37, 1269–1277. [Google Scholar] [CrossRef] [PubMed]
- Duangjan, C.; Rangsinth, P.; Zhang, S.; Gu, X.; Wink, M.; Tencomnao, T. Vitis Vinifera Leaf Extract Protects Against Glutamate-Induced Oxidative Toxicity in HT22 Hippocampal Neuronal Cells and Increases Stress Resistance Properties in Caenorhabditis Elegans. Front. Nutr. 2021, 8, 634100. [Google Scholar] [CrossRef]
- Lee, H.Y.; Weon, J.B.; Ryu, G.; Yang, W.S.; Kim, N.Y.; Kim, M.K.; Ma, C.J. Neuroprotective Effect of Aronia Melanocarpa Extract against Glutamate-Induced Oxidative Stress in HT22 Cells. BMC Complement. Altern. Med. 2017, 17, 207. [Google Scholar] [CrossRef]
- Shah, S.A.; Amin, F.U.; Khan, M.; Abid, M.N.; Rehman, S.U.; Kim, T.H.; Kim, M.W.; Kim, M.O. Anthocyanins Abrogate Glutamate-Induced AMPK Activation, Oxidative Stress, Neuroinflammation, and Neurodegeneration in Postnatal Rat Brain. J. Neuroinflamm. 2016, 13, 286. [Google Scholar] [CrossRef]
- Wen, X.; Huang, A.; Hu, J.; Zhong, Z.; Liu, Y.; Li, Z.; Pan, X.; Liu, Z. Neuroprotective Effect of Astaxanthin against Glutamate-Induced Cytotoxicity in HT22 Cells: Involvement of the Akt/GSK-3β Pathway. Neuroscience 2015, 303, 558–568. [Google Scholar] [CrossRef]
- Muzio, L.; Viotti, A.; Martino, G. Microglia in Neuroinflammation and Neurodegeneration: From Understanding to Therapy. Front. Neurosci. 2021, 15, 742065. [Google Scholar] [CrossRef]
- Shao, F.; Wang, X.; Wu, H.; Wu, Q.; Zhang, J. Microglia and Neuroinflammation: Crucial Pathological Mechanisms in Traumatic Brain Injury-Induced Neurodegeneration. Front. Aging Neurosci. 2022, 14, 825086. [Google Scholar] [CrossRef]
- Mango, D.; Saidi, A.; Cisale, G.Y.; Feligioni, M.; Corbo, M.; Nisticò, R. Targeting Synaptic Plasticity in Experimental Models of Alzheimer’s Disease. Front. Pharmacol. 2019, 10, 778. [Google Scholar] [CrossRef]
- Stephenson, J.; Nutma, E.; van der Valk, P.; Amor, S. Inflammation in CNS Neurodegenerative Diseases. Immunology 2018, 154, 204–219. [Google Scholar] [CrossRef] [PubMed]
- Kettenmann, H.; Hanisch, U.-K.; Noda, M.; Verkhratsky, A. Physiology of Microglia. Physiol. Rev. 2011, 91, 461–553. [Google Scholar] [CrossRef] [PubMed]
- Kacimi, R.; Giffard, R.G.; Yenari, M.A. Endotoxin-Activated Microglia Injure Brain Derived Endothelial Cells via NF-ΚB, JAK-STAT and JNK Stress Kinase Pathways. J. Inflamm. 2011, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Shabab, T.; Khanabdali, R.; Moghadamtousi, S.Z.; Kadir, H.A.; Mohan, G. Neuroinflammation Pathways: A General Review. Int. J. Neurosci. 2017, 127, 624–633. [Google Scholar] [CrossRef]
- Frakes, A.E.; Ferraiuolo, L.; Haidet-Phillips, A.M.; Schmelzer, L.; Braun, L.; Miranda, C.J.; Ladner, K.J.; Bevan, A.K.; Foust, K.D.; Godbout, J.P.; et al. Microglia Induce Motor Neuron Death via the Classical NF-ΚB Pathway in Amyotrophic Lateral Sclerosis. Neuron 2014, 81, 1009–1023. [Google Scholar] [CrossRef] [PubMed]
- Clausen, B.H.; Lambertsen, K.L.; Babcock, A.A.; Holm, T.H.; Dagnaes-Hansen, F.; Finsen, B. Interleukin-1beta and Tumor Necrosis Factor-Alpha Are Expressed by Different Subsets of Microglia and Macrophages after Ischemic Stroke in Mice. J. Neuroinflamm. 2008, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Garaschuk, O.; Verkhratsky, A. Physiology of Microglia. Methods Mol. Biol. 2019, 2034, 27–40. [Google Scholar]
- Poulose, S.M.; Fisher, D.R.; Larson, J.; Bielinski, D.F.; Rimando, A.M.; Carey, A.N.; Schauss, A.G.; Shukitt-Hale, B. Anthocyanin-Rich Açai (Euterpe Oleracea Mart.) Fruit Pulp Fractions Attenuate Inflammatory Stress Signaling in Mouse Brain BV-2 Microglial Cells. J. Agric. Food Chem. 2012, 60, 1084–1093. [Google Scholar] [CrossRef]
- Jeong, J.-W.; Lee, W.; Shin, S.; Kim, G.-Y.; Choi, B.; Choi, Y. Anthocyanins Downregulate Lipopolysaccharide-Induced Inflammatory Responses in BV2 Microglial Cells by Suppressing the NF-ΚB and Akt/MAPKs Signaling Pathways. Int. J. Mol. Sci. 2013, 14, 1502–1515. [Google Scholar] [CrossRef]
- Ali, T.; Kim, M.J.; Rehman, S.U.; Ahmad, A.; Kim, M.O. Anthocyanin-Loaded PEG-Gold Nanoparticles Enhanced the Neuroprotection of Anthocyanins in an Aβ1–42 Mouse Model of Alzheimer’s Disease. Mol. Neurobiol. 2017, 54, 6490–6506. [Google Scholar] [CrossRef]
- Carvalho, F.B.; Gutierres, J.M.; Bohnert, C.; Zago, A.M.; Abdalla, F.H.; Vieira, J.M.; Palma, H.E.; Oliveira, S.M.; Spanevello, R.M.; Duarte, M.M.; et al. Anthocyanins Suppress the Secretion of Proinflammatory Mediators and Oxidative Stress, and Restore Ion Pump Activities in Demyelination. J. Nutr. Biochem. 2015, 26, 378–390. [Google Scholar] [CrossRef]
- Moreira, N.C.D.S.; Lima, J.E.B.D.F.; Marchiori, M.F.; Carvalho, I.; Sakamoto-Hojo, E.T. Neuroprotective Effects of Cholinesterase Inhibitors: Current Scenario in Therapies for Alzheimer’s Disease and Future Perspectives. J. Alzheimers Dis. Rep. 2022, 6, 177–193. [Google Scholar] [CrossRef]
- Moss, D.E. Improving Anti-Neurodegenerative Benefits of Acetylcholinesterase Inhibitors in Alzheimer’s Disease: Are Irreversible Inhibitors the Future? Int. J. Mol. Sci. 2020, 21, 3438. [Google Scholar] [CrossRef] [PubMed]
- Pervin, M.; Hasnat, M.A.; Lee, Y.M.; Kim, D.H.; Jo, J.E.; Lim, B.O. Antioxidant Activity and Acetylcholinesterase Inhibition of Grape Skin Anthocyanin (GSA). Molecules 2014, 19, 9403–9418. [Google Scholar] [CrossRef] [PubMed]
- Siniawska, M.; Wojdyło, A. Polyphenol Profiling by LC QTOF/ESI-MS and Biological Activity of Purple Passion Fruit Epicarp Extract. Molecules 2023, 28, 6711. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Li, F.; Zhang, J.; Yang, B.; Ji, Z.; Chen, W. Phytochemical Compositions of Extract from Peel of Hawthorn Fruit, and Its Antioxidant Capacity, Cell Growth Inhibition, and Acetylcholinesterase Inhibitory Activity. BMC Complement. Altern. Med. 2017, 17, 151. [Google Scholar] [CrossRef] [PubMed]
- Temviriyanukul, P.; Sritalahareuthai, V.; Jom, K.N.; Jongruaysup, B.; Tabtimsri, S.; Pruesapan, K.; Thangsiri, S.; Inthachat, W.; Siriwan, D.; Charoenkiatkul, S.; et al. Comparison of Phytochemicals, Antioxidant, and In Vitro Anti-Alzheimer Properties of Twenty-Seven Morus Spp. Cultivated in Thailand. Molecules 2020, 25, 2600. [Google Scholar] [CrossRef]
- Afzal, M.; Redha, A.; AlHasan, R. Anthocyanins Potentially Contribute to Defense against Alzheimer’s Disease. Molecules 2019, 24, 4255. [Google Scholar] [CrossRef]
- Yousof Ali, M.; Zaib, S.; Jannat, S.; Khan, I. Discovery of Potent and Selective Dual Cholinesterases and β-Secretase Inhibitors in Pomegranate as a Treatment for Alzheimer’s Disease. Bioorg Chem. 2022, 129, 106137. [Google Scholar] [CrossRef]
- Koly, H.K.; Sutradhar, K.; Rahman, M.S. Acetylcholinesterase Inhibition of Alzheimer’s Disease: Identification of Potential Phytochemicals and Designing More Effective Derivatives to Manage Disease Condition. J. Biomol. Struct. Dyn. 2023, 1–13. [Google Scholar] [CrossRef]
- Kakutani, S.; Watanabe, H.; Murayama, N. Green Tea Intake and Risks for Dementia, Alzheimer’s Disease, Mild Cognitive Impairment, and Cognitive Impairment: A Systematic Review. Nutrients 2019, 11, 1165. [Google Scholar] [CrossRef] [PubMed]
- Silveira, I.A.; Mullis, A.S.; Cairns, D.M.; Shevzov-Zebrun, A.; Whalen, J.; Galuppo, A.; Walsh, K.G.; Kaplan, D.L. Screening Neuroprotective Compounds in Herpes-Induced Alzheimer’s Disease Cell and 3D Tissue Models. Free Radic. Biol. Med. 2022, 186, 76–92. [Google Scholar] [CrossRef] [PubMed]
- Zandi, P.P. Reduced Risk of Alzheimer Disease in Users of Antioxidant Vitamin Supplements. Arch. Neurol. 2004, 61, 82. [Google Scholar] [CrossRef] [PubMed]
- Vepsäläinen, S.; Koivisto, H.; Pekkarinen, E.; Mäkinen, P.; Dobson, G.; McDougall, G.J.; Stewart, D.; Haapasalo, A.; Karjalainen, R.O.; Tanila, H.; et al. Anthocyanin-Enriched Bilberry and Blackcurrant Extracts Modulate Amyloid Precursor Protein Processing and Alleviate Behavioral Abnormalities in the APP/PS1 Mouse Model of Alzheimer’s Disease. J. Nutr. Biochem. 2013, 24, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Gutierres, J.M.; Carvalho, F.B.; Schetinger, M.R.C.; Marisco, P.; Agostinho, P.; Rodrigues, M.; Rubin, M.A.; Schmatz, R.; da Silva, C.R.; Cognato, G.D.P.; et al. Anthocyanins Restore Behavioral and Biochemical Changes Caused by Streptozotocin-Induced Sporadic Dementia of Alzheimer’s Type. Life Sci. 2014, 96, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Zhang, J.; Qin, M. Protective Effect of Cyanidin 3-O-Glucoside on Beta-Amyloid Peptide-Induced Cognitive Impairment in Rats. Neurosci. Lett. 2013, 534, 285–288. [Google Scholar] [CrossRef]
- Belkacemi, A.; Ramassamy, C. Innovative Anthocyanin/Anthocyanidin Formulation Protects SK-N-SH Cells Against the Amyloid-β Peptide-Induced Toxicity: Relevance to Alzheimer’s Disease. Cent. Nerv. Syst. Agents Med. Chem. 2015, 16, 37–49. [Google Scholar] [CrossRef]
- Yamakawa, M.Y.; Uchino, K.; Watanabe, Y.; Adachi, T.; Nakanishi, M.; Ichino, H.; Hongo, K.; Mizobata, T.; Kobayashi, S.; Nakashima, K.; et al. Anthocyanin Suppresses the Toxicity of Aβ Deposits through Diversion of Molecular Forms in in Vitro and in Vivo Models of Alzheimer’s Disease. Nutr. Neurosci. 2016, 19, 32–42. [Google Scholar] [CrossRef]
- Tarozzi, A.; Morroni, F.; Merlicco, A.; Bolondi, C.; Teti, G.; Falconi, M.; Cantelli-Forti, G.; Hrelia, P. Neuroprotective Effects of Cyanidin 3-O-Glucopyranoside on Amyloid Beta (25–35) Oligomer-Induced Toxicity. Neurosci. Lett. 2010, 473, 72–76. [Google Scholar] [CrossRef]
- Zhu, Y.; Hou, H.; Nikolic, W.V.; Ehrhart, J.; Rrapo, E.; Bickford, P.; Giunta, B.; Tan, J. CD45RB Is a Novel Molecular Therapeutic Target to Inhibit Aβ Peptide-Induced Microglial MAPK Activation. PLoS ONE 2008, 3, e2135. [Google Scholar] [CrossRef]
- Pacheco, S.M.; Soares, M.S.P.; Gutierres, J.M.; Gerzson, M.F.B.; Carvalho, F.B.; Azambuja, J.H.; Schetinger, M.R.C.; Stefanello, F.M.; Spanevello, R.M. Anthocyanins as a Potential Pharmacological Agent to Manage Memory Deficit, Oxidative Stress and Alterations in Ion Pump Activity Induced by Experimental Sporadic Dementia of Alzheimer’s Type. J. Nutr. Biochem. 2018, 56, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Badshah, H.; Kim, T.H.; Kim, M.O. Protective Effects of Anthocyanins against Amyloid Beta-Induced Neurotoxicity in Vivo and in Vitro. Neurochem. Int. 2015, 80, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Ali, T.; Kim, T.; Rehman, S.U.; Khan, M.S.; Amin, F.U.; Khan, M.; Ikram, M.; Kim, M.O. Natural Dietary Supplementation of Anthocyanins via PI3K/Akt/Nrf2/HO-1 Pathways Mitigate Oxidative Stress, Neurodegeneration, and Memory Impairment in a Mouse Model of Alzheimer’s Disease. Mol. Neurobiol. 2018, 55, 6076–6093. [Google Scholar] [CrossRef] [PubMed]
- Gutierres, J.M.; Carvalho, F.B.; Schetinger, M.R.C.; Agostinho, P.; Marisco, P.C.; Vieira, J.M.; Rosa, M.M.; Bohnert, C.; Rubin, M.A.; Morsch, V.M.; et al. Neuroprotective Effect of Anthocyanins on Acetylcholinesterase Activity and Attenuation of Scopolamine-Induced Amnesia in Rats. Int. J. Dev. Neurosci. 2014, 33, 88–97. [Google Scholar] [CrossRef]
- de Mello, J.E.; Luduvico, K.P.; Dos Santos, A.; Teixeira, F.C.; de Souza Cardoso, J.; de Aguiar, M.S.S.; Cunico, W.; Vizzotto, M.; Stefanello, F.; Spanevello, R. Therapeutic Potential of Blackberry Extract in the Preventing Memory Deficits and Neurochemical Alterations in the Cerebral Cortex, Hippocampus and Cerebellum of a Rat Model with Amnesia. Metab. Brain Dis. 2023, 38, 1261–1272. [Google Scholar] [CrossRef]
- Papandreou, M.A.; Dimakopoulou, A.; Linardaki, Z.I.; Cordopatis, P.; Klimis-Zacas, D.; Margarity, M.; Lamari, F.N. Effect of a Polyphenol-Rich Wild Blueberry Extract on Cognitive Performance of Mice, Brain Antioxidant Markers and Acetylcholinesterase Activity. Behav. Brain Res. 2009, 198, 352–358. [Google Scholar] [CrossRef]
- Heysieattalab, S.; Sadeghi, L. Effects of Delphinidin on Pathophysiological Signs of Nucleus Basalis of Meynert Lesioned Rats as Animal Model of Alzheimer Disease. Neurochem. Res. 2020, 45, 1636–1646. [Google Scholar] [CrossRef]
- Gao, X.; Cassidy, A.; Schwarzschild, M.A.; Rimm, E.B.; Ascherio, A. Habitual Intake of Dietary Flavonoids and Risk of Parkinson Disease. Neurology 2012, 78, 1138–1145. [Google Scholar] [CrossRef]
- Fan, D.; Alamri, Y.; Liu, K.; MacAskill, M.; Harris, P.; Brimble, M.; Dalrymple-Alford, J.; Prickett, T.; Menzies, O.; Laurenson, A.; et al. Supplementation of Blackcurrant Anthocyanins Increased Cyclic Glycine-Proline in the Cerebrospinal Fluid of Parkinson Patients: Potential Treatment to Improve Insulin-Like Growth Factor-1 Function. Nutrients 2018, 10, 714. [Google Scholar] [CrossRef]
- Kim, H.G.; Ju, M.S.; Shim, J.S.; Kim, M.C.; Lee, S.-H.; Huh, Y.; Kim, S.Y.; Oh, M.S. Mulberry Fruit Protects Dopaminergic Neurons in Toxin-Induced Parkinson’s Disease Models. Br. J. Nutr. 2010, 104, 8–16. [Google Scholar] [CrossRef]
- Strathearn, K.E.; Yousef, G.G.; Grace, M.H.; Roy, S.L.; Tambe, M.A.; Ferruzzi, M.G.; Wu, Q.-L.; Simon, J.E.; Lila, M.A.; Rochet, J.-C. Neuroprotective Effects of Anthocyanin- and Proanthocyanidin-Rich Extracts in Cellular Models of Parkinson’s Disease. Brain Res. 2014, 1555, 60–77. [Google Scholar] [CrossRef]
- Long, J.; Gao, H.; Sun, L.; Liu, J.; Zhao-Wilson, X. Grape Extract Protects Mitochondria from Oxidative Damage and Improves Locomotor Dysfunction and Extends Lifespan in a Drosophila Parkinson’s Disease Model. Rejuvenation Res. 2009, 12, 321–331. [Google Scholar] [CrossRef]
- Roghani, M.; Niknam, A.; Jalali-Nadoushan, M.-R.; Kiasalari, Z.; Khalili, M.; Baluchnejadmojarad, T. Oral Pelargonidin Exerts Dose-Dependent Neuroprotection in 6-Hydroxydopamine Rat Model of Hemi-Parkinsonism. Brain Res. Bull. 2010, 82, 279–283. [Google Scholar] [CrossRef]
- Chen, H.; Yoshioka, H.; Kim, G.S.; Jung, J.E.; Okami, N.; Sakata, H.; Maier, C.M.; Narasimhan, P.; Goeders, C.E.; Chan, P.H. Oxidative Stress in Ischemic Brain Damage: Mechanisms of Cell Death and Potential Molecular Targets for Neuroprotection. Antioxid. Redox Signal 2011, 14, 1505–1517. [Google Scholar] [CrossRef] [PubMed]
- Shin, W.-H.; Park, S.-J.; Kim, E.-J. Protective Effect of Anthocyanins in Middle Cerebral Artery Occlusion and Reperfusion Model of Cerebral Ischemia in Rats. Life Sci. 2006, 79, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Cui, M.; Dai, G.; Yuan, T.; Li, Y.; Ji, T.; Pan, Y. Protective Effect of Anthocyanin on Neurovascular Unit in Cerebral Ischemia/Reperfusion Injury in Rats. Front. Neurosci. 2018, 12, 947. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.-X.; Chen, J.-H.; Li, J.-W.; Cheng, F.-R.; Yuan, K. Protection of Anthocyanin from Myrica Rubra against Cerebral Ischemia-Reperfusion Injury via Modulation of the TLR4/NF-ΚB and NLRP3 Pathways. Molecules 2018, 23, 1788. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.H.; Hur, J.Y.; Kim, H.B.; Ryu, J.H.; Kim, S.Y. Neuroprotective Effects of the Cyanidin-3-O-β-d-Glucopyranoside Isolated from Mulberry Fruit against Cerebral Ischemia. Neurosci. Lett. 2006, 391, 122–126. [Google Scholar] [CrossRef]
- Hwang, S.-N.; Kim, J.-C.; Bhuiyan, M.I.H.; Kim, J.Y.; Yang, J.S.; Yoon, S.H.; Yoon, K.D.; Kim, S.Y. Black Rice (Oryza Sativa L., Poaceae) Extract Reduces Hippocampal Neuronal Cell Death Induced by Transient Global Cerebral Ischemia in Mice. Exp. Neurobiol. 2018, 27, 129–138. [Google Scholar] [CrossRef]
- Adnyana, I.M.O.; Sudewi, A.R.; Samatra, D.P.; Suprapta, D.N. Neuroprotective Effects of Purple Sweet Potato Balinese Cultivar in Wistar Rats with Ischemic Stroke. Open Access Maced. J. Med. Sci. 2018, 6, 1959–1964. [Google Scholar] [CrossRef]
- Skemiene, K.; Pampuscenko, K.; Rekuviene, E.; Borutaite, V. Protective Effects of Anthocyanins against Brain Ischemic Damage. J. Bioenerg. Biomembr. 2020, 52, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Skemiene, K.; Rakauskaite, G.; Trumbeckaite, S.; Liobikas, J.; Brown, G.C.; Borutaite, V. Anthocyanins Block Ischemia-Induced Apoptosis in the Perfused Heart and Support Mitochondrial Respiration Potentially by Reducing Cytosolic Cytochrome c. Int. J. Biochem. Cell Biol. 2013, 45, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, J.; Liu, F.; Tong, L.; Chen, Z.; Chen, J.; He, H.; Xu, R.; Ma, Y.; Huang, C. Neuroprotective Effects of Anthocyanins and Its Major Component Cyanidin-3-O-Glucoside (C3G) in the Central Nervous System: An Outlined Review. Eur. J. Pharmacol. 2019, 858, 172500. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.I.H.; Kim, J.Y.; Ha, T.J.; Kim, S.Y.; Cho, K.-O. Anthocyanins Extracted from Black Soybean Seed Coat Protect Primary Cortical Neurons against in Vitro Ischemia. Biol. Pharm. Bull. 2012, 35, 999–1008. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaa, C.A.; Marcelo, Á.J.; An, Z.; Medina-Franco, J.L.; Velasco-Velázquez, M.A. Anthocyanins: Molecular Aspects on Their Neuroprotective Activity. Biomolecules 2023, 13, 1598. https://doi.org/10.3390/biom13111598
Zaa CA, Marcelo ÁJ, An Z, Medina-Franco JL, Velasco-Velázquez MA. Anthocyanins: Molecular Aspects on Their Neuroprotective Activity. Biomolecules. 2023; 13(11):1598. https://doi.org/10.3390/biom13111598
Chicago/Turabian StyleZaa, César A., Álvaro J. Marcelo, Zhiqiang An, José L. Medina-Franco, and Marco A. Velasco-Velázquez. 2023. "Anthocyanins: Molecular Aspects on Their Neuroprotective Activity" Biomolecules 13, no. 11: 1598. https://doi.org/10.3390/biom13111598
APA StyleZaa, C. A., Marcelo, Á. J., An, Z., Medina-Franco, J. L., & Velasco-Velázquez, M. A. (2023). Anthocyanins: Molecular Aspects on Their Neuroprotective Activity. Biomolecules, 13(11), 1598. https://doi.org/10.3390/biom13111598