
Th17-Dependent Nasal Hyperresponsiveness Is Mitigated by Steroid Treatment

 ,
, 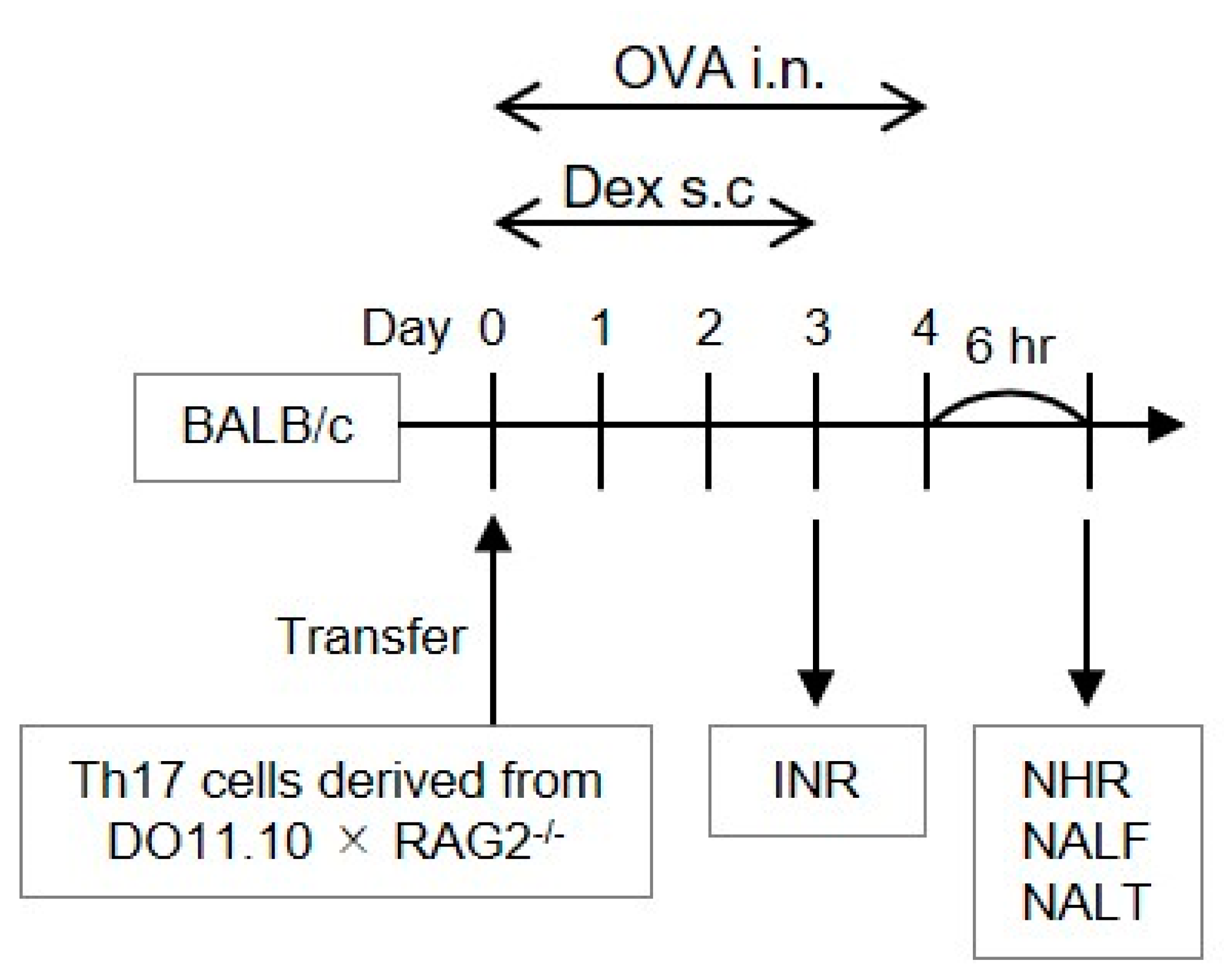{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
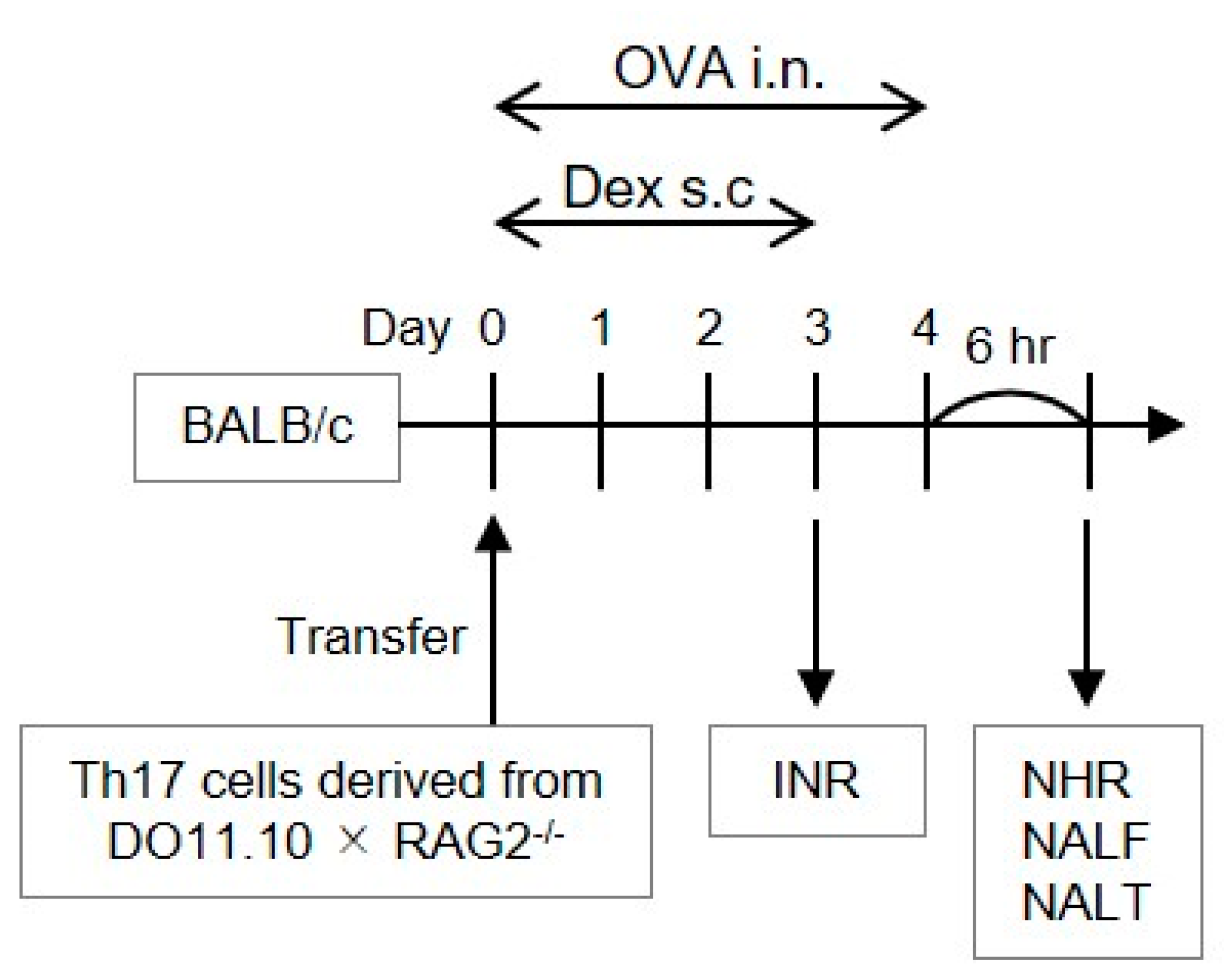
2. Materials and Methods
2.1. In Vitro Development of Allergen-Reactive Th17 Cells
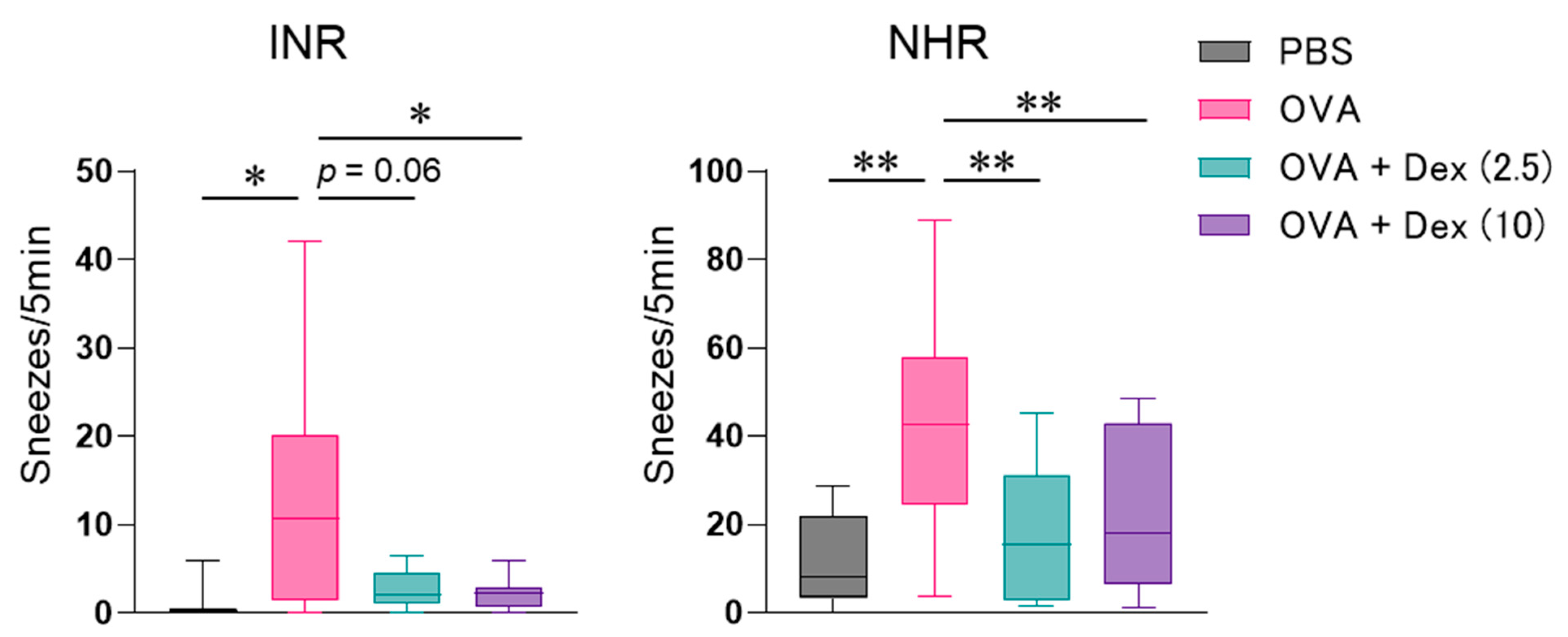
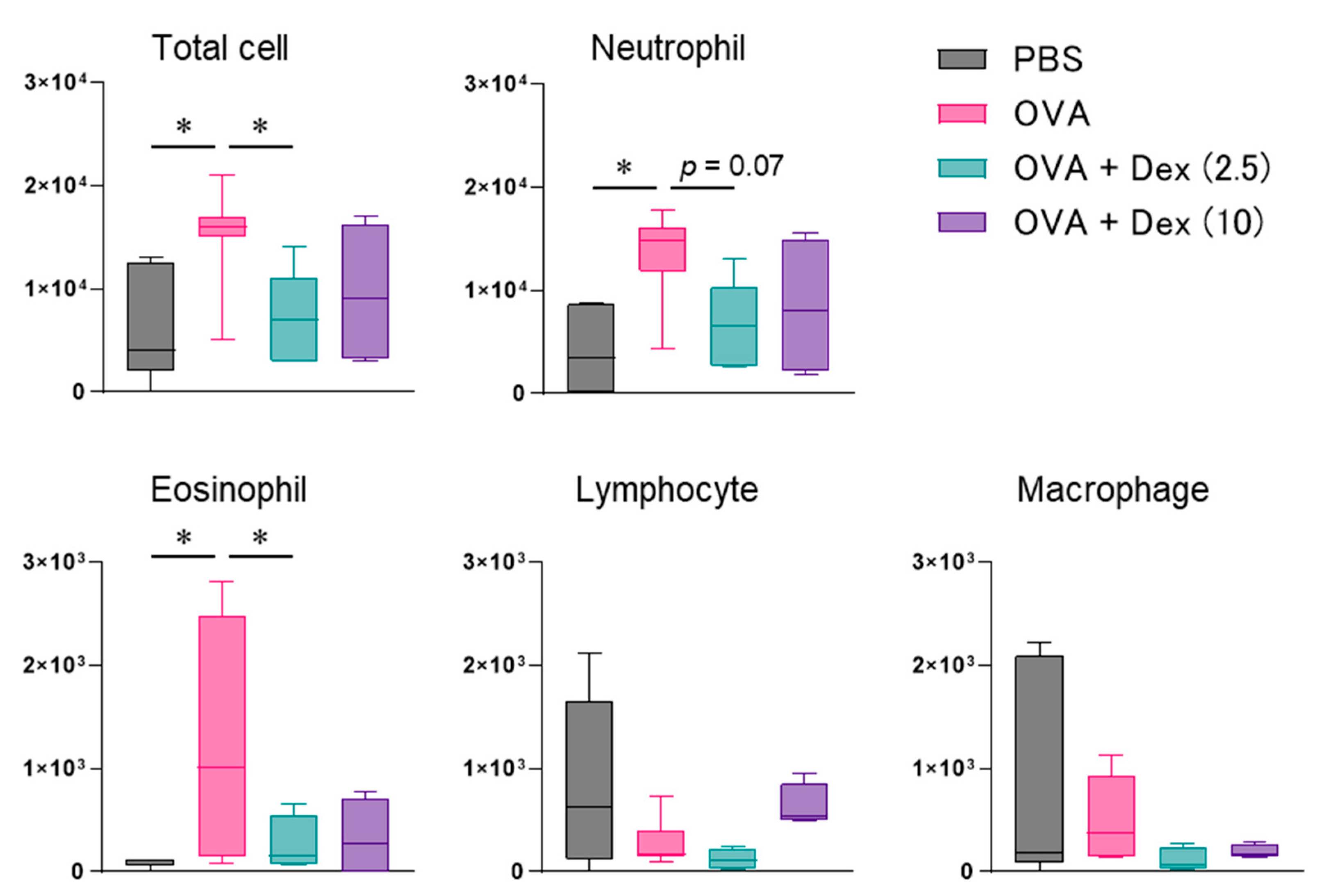
2.2. Allergen-Induced Nasal Responses
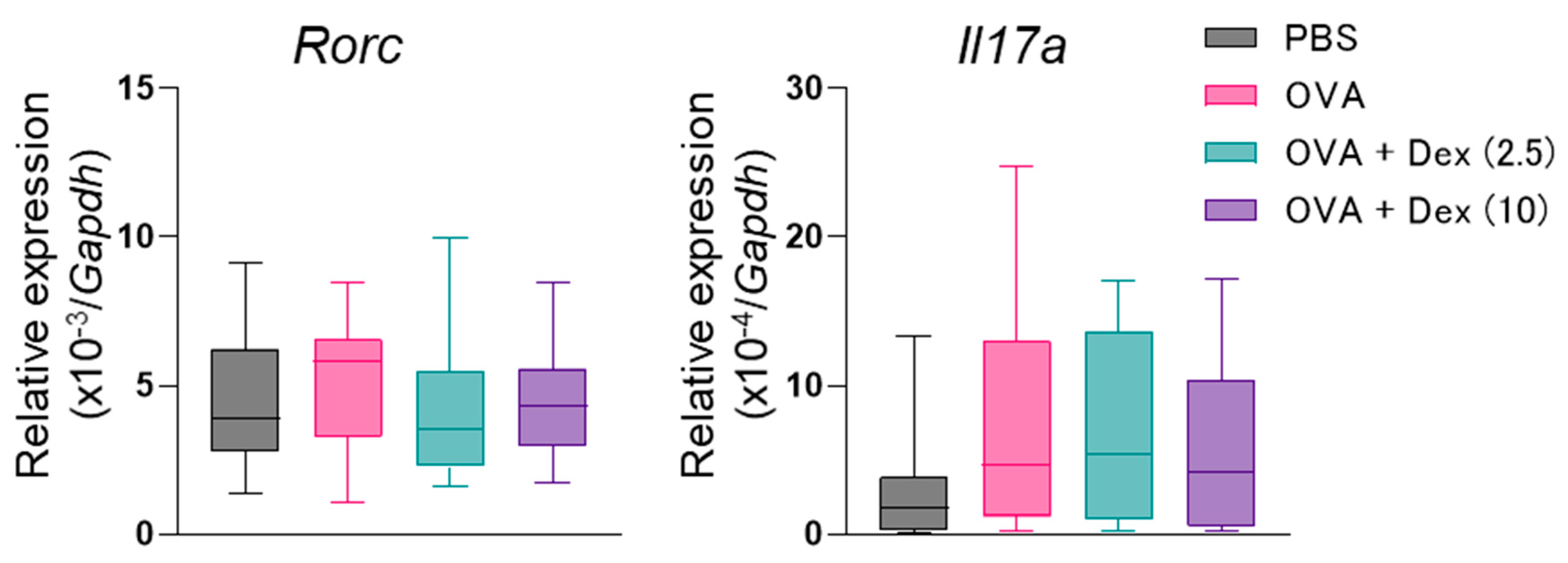
2.3. Gene Expression Assessment
2.4. Statistical Analysis
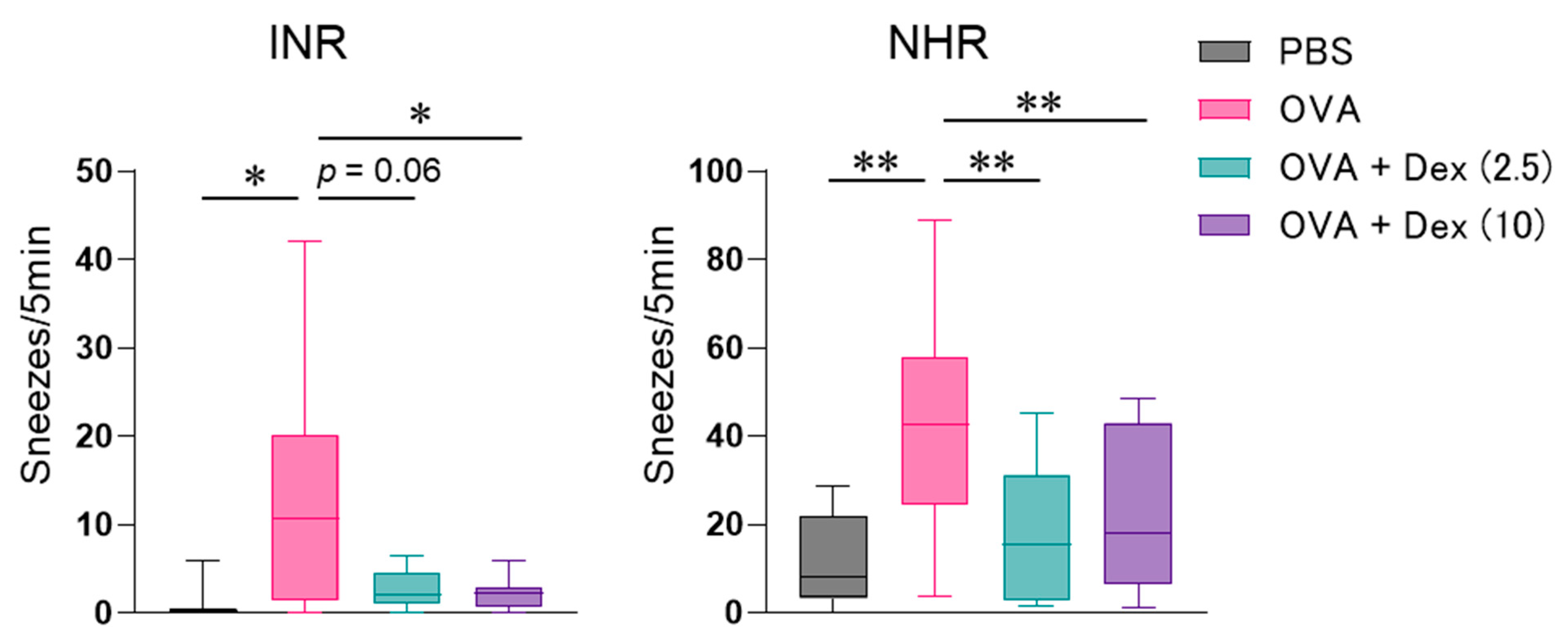
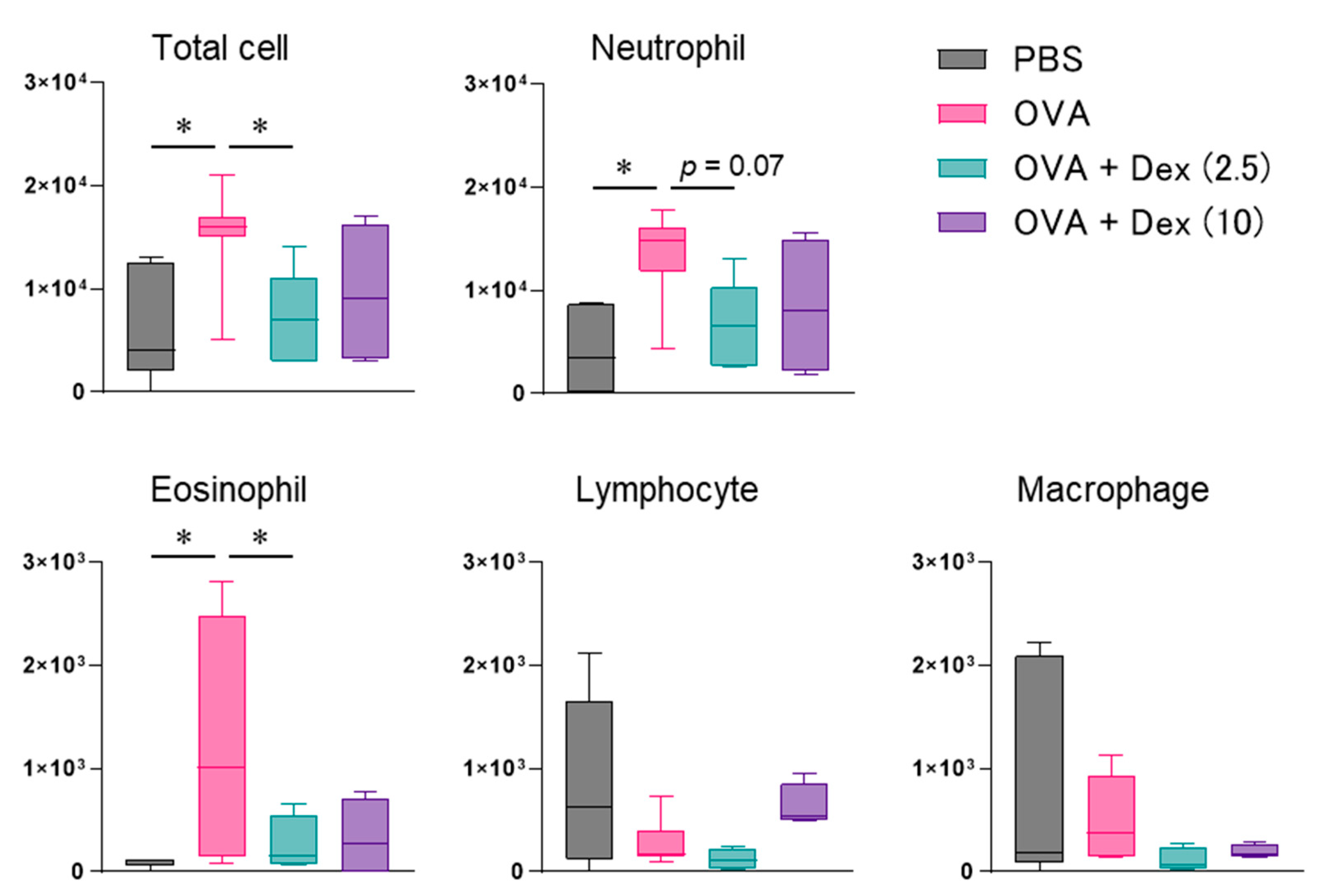
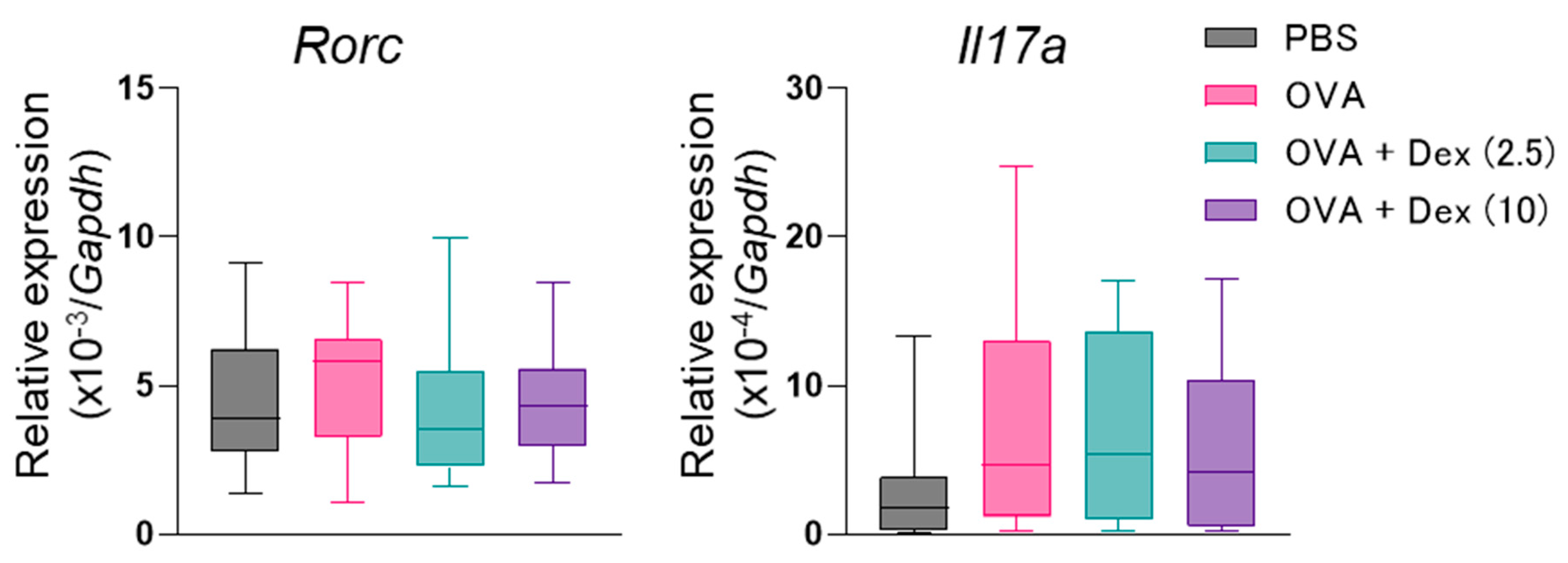
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Sin, B.; Togias, A. Pathophysiology of allergic and nonallergic rhinitis. Proc. Am. Thorac. Soc. 2011, 8, 106–114. [Google Scholar] [CrossRef] [PubMed]
- De Graaf-in t Veld, C.; Garrelds, I.M.; Koenders, S.; Gerth van Wijk, R. Relationship between nasal hyperreactivity, mediators and eosinophils in patients with perennial allergic rhinitis and controls. Clin. Exp. Allergy 1996, 26, 903–908. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.M.; Cripps, A.W.; West, N.P.; Cox, A.J. Modulation of Allergic Inflammation in the Nasal Mucosa of Allergic Rhinitis Sufferers with Topical Pharmaceutical Agents. Front. Pharmacol. 2019, 10, 294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishimura, T.; Kaminuma, O.; Saeki, M.; Kitamura, N.; Matsuoka, K.; Yonekawa, H.; Mori, A.; Hiroi, T. Essential Contribution of CD4+ T Cells to Antigen-Induced Nasal Hyperresponsiveness in Experimental Allergic Rhinitis. PLoS ONE 2016, 11, e0146686. [Google Scholar] [CrossRef]
- Koyama, T.; Miura, K.; Yamasaki, N.; Ogata, S.; Ito, D.; Saeki, M.; Hiroi, T.; Mori, A.; Kaminuma, O. Suppressive effect of dexamethasone on murine Th9 cell-mediated nasal eosinophilic inflammation. Asia Pac. Allergy 2021, 11, e25. [Google Scholar] [CrossRef]
- Bousquet, P.J.; Bachert, C.; Canonica, G.W.; Casale, T.B.; Mullol, J.; Klossek, J.M.; Zuberbier, T.; Bousquet, J. Uncontrolled allergic rhinitis during treatment and its impact on quality of life: A cluster randomized trial. J. Allergy Clin. Immunol. 2010, 126, 666–668. [Google Scholar] [CrossRef]
- McKinley, L.; Alcorn, J.F.; Peterson, A.; Dupont, R.B.; Kapadia, S.; Logar, A.; Henry, A.; Irvin, C.G.; Piganelli, J.D.; Ray, A.; et al. TH17 cells mediate steroid-resistant airway inflammation and airway hyperresponsiveness in mice. J. Immunol. 2008, 181, 4089–4097. [Google Scholar] [CrossRef] [Green Version]
- Kaminuma, O.; Ohtomo, T.; Mori, A.; Nagakubo, D.; Hieshima, K.; Ohmachi, Y.; Noda, Y.; Katayama, K.; Suzuki, K.; Motoi, Y.; et al. Selective down-regulation of Th2 cell-mediated airway inflammation in mice by pharmacological intervention of CCR4. Clin. Exp. Allergy 2012, 42, 315–325. [Google Scholar] [CrossRef]
- Saeki, M.; Nishimura, T.; Mori, A.; Kaminuma, O.; Hiroi, T. Antigen-induced mixed and separated inflammation in murine upper and lower airways. Allergol. Int. 2014, 63, 59–61. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, T.; Kaminuma, O.; Saeki, M.; Kitamura, N.; Gotoh, M.; Mori, A.; Hiroi, T. Effects of anti-allergic drugs on T cell-mediated nasal hyperresponsiveness in a murine model of allergic rhinitis. Allergol. Int. 2018, 67S, S25–S31. [Google Scholar] [CrossRef]
- Saeki, M.; Kaminuma, O.; Nishimura, T.; Kitamura, N.; Mori, A.; Hiroi, T. Th9 cells induce steroid-resistant bronchial hyperresponsiveness in mice. Allergol. Int. 2017, 66S, S35–S40. [Google Scholar] [CrossRef] [PubMed]
- Koch, S.; Sopel, N.; Finotto, S. Th9 and other IL-9-producing cells in allergic asthma. Semin. Immunopathol. 2017, 39, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Nabe, T. Steroid-Resistant Asthma and Neutrophils. Biol. Pharm. Bull. 2020, 43, 31–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villey, I.; de Chasseval, R.; de Villartay, J.P. RORgammaT, a thymus-specific isoform of the orphan nuclear receptor RORgamma/TOR, is up-regulated by signaling through the pre-T cell receptor and binds to the TEA promoter. Eur. J. Immunol. 1999, 29, 4072–4080. [Google Scholar] [CrossRef]
- Ivanov, I.I.; McKenzie, B.S.; Zhou, L.; Tadokoro, C.E.; Lepelley, A.; Lafaille, J.J.; Cua, D.J.; Littman, D.R. The orphan nuclear receptor RORgammat directs the differentiation program of proinflammatory IL-17+ T helper cells. Cell 2006, 126, 1121–1133. [Google Scholar] [CrossRef] [Green Version]
- Murdaca, G.; Colombo, B.M.; Puppo, F. The role of Th17 lymphocytes in the autoimmune and chronic inflammatory diseases. Intern. Emerg. Med. 2011, 6, 487–495. [Google Scholar] [CrossRef]
- Bullens, D.M.; Truyen, E.; Coteur, L.; Dilissen, E.; Hellings, P.W.; Dupont, L.J.; Ceuppens, J.L. IL-17 mRNA in sputum of asthmatic patients: Linking T cell driven inflammation and granulocytic influx? Respir. Res. 2006, 7, 135. [Google Scholar] [CrossRef] [Green Version]
- Al-Ramli, W.; Prefontaine, D.; Chouiali, F.; Martin, J.G.; Olivenstein, R.; Lemiere, C.; Hamid, Q. T(H)17-associated cytokines (IL-17A and IL-17F) in severe asthma. J. Allergy Clin. Immunol. 2009, 123, 1185–1187. [Google Scholar] [CrossRef]
- Kudo, M.; Melton, A.C.; Chen, C.; Engler, M.B.; Huang, K.E.; Ren, X.; Wang, Y.; Bernstein, X.; Li, J.T.; Atabai, K.; et al. IL-17A produced by alphabeta T cells drives airway hyper-responsiveness in mice and enhances mouse and human airway smooth muscle contraction. Nat. Med. 2012, 18, 547–554. [Google Scholar] [CrossRef] [Green Version]
- Chenuet, P.; Fauconnier, L.; Madouri, F.; Marchiol, T.; Rouxel, N.; Ledru, A.; Mauny, P.; Lory, R.; Uttenhove, C.; van Snick, J.; et al. Neutralization of either IL-17A or IL-17F is sufficient to inhibit house dust mite induced allergic asthma in mice. Clin. Sci. 2017, 131, 2533–2548. [Google Scholar] [CrossRef]
- Gupta, A.; Dimeloe, S.; Richards, D.F.; Chambers, E.S.; Black, C.; Urry, Z.; Ryanna, K.; Xystrakis, E.; Bush, A.; Saglani, S.; et al. Defective IL-10 expression and in vitro steroid-induced IL-17A in paediatric severe therapy-resistant asthma. Thorax 2014, 69, 508–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansbro, P.M.; Kim, R.Y.; Starkey, M.R.; Donovan, C.; Dua, K.; Mayall, J.R.; Liu, G.; Hansbro, N.G.; Simpson, J.L.; Wood, L.G.; et al. Mechanisms and treatments for severe, steroid-resistant allergic airway disease and asthma. Immunol. Rev. 2017, 278, 41–62. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ueda, S.; Miura, K.; Kawasaki, H.; Ogata, S.; Yamasaki, N.; Miura, S.; Mori, A.; Kaminuma, O. Th17-Dependent Nasal Hyperresponsiveness Is Mitigated by Steroid Treatment. Biomolecules 2022, 12, 674. https://doi.org/10.3390/biom12050674
Ueda S, Miura K, Kawasaki H, Ogata S, Yamasaki N, Miura S, Mori A, Kaminuma O. Th17-Dependent Nasal Hyperresponsiveness Is Mitigated by Steroid Treatment. Biomolecules. 2022; 12(5):674. https://doi.org/10.3390/biom12050674
Chicago/Turabian StyleUeda, Shusaku, Kento Miura, Hideki Kawasaki, Sawako Ogata, Norimasa Yamasaki, Shuka Miura, Akio Mori, and Osamu Kaminuma. 2022. "Th17-Dependent Nasal Hyperresponsiveness Is Mitigated by Steroid Treatment" Biomolecules 12, no. 5: 674. https://doi.org/10.3390/biom12050674
APA StyleUeda, S., Miura, K., Kawasaki, H., Ogata, S., Yamasaki, N., Miura, S., Mori, A., & Kaminuma, O. (2022). Th17-Dependent Nasal Hyperresponsiveness Is Mitigated by Steroid Treatment. Biomolecules, 12(5), 674. https://doi.org/10.3390/biom12050674