
Fungal Guttation, a Source of Bioactive Compounds, and Its Ecological Role—A Review


Abstract
1. Introduction
2. Conditions and Factors Promoting Guttation
3. Ecological Role of Fungal Exudates
4. Secondary Metabolites Present in Fungal Exudates
4.1. Antimicrobials
4.1.1. Antibacterial and Antiviral Compounds
4.1.2. Antifungal Compounds
4.2. Bioinsecticides, Bioherbicides
4.3. Mycotoxins
4.4. Anticancer Substances
5. Proteins Excreted by Guttation
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bills, G.F.; Gloer, J.B. Biologically active secondary metabolites from the fungi. Microbiol. Spectr. 2016, 4, 4–6. [Google Scholar] [CrossRef]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.H.; Stone, M.J.; Hauck, P.R.; Rahman, S.K. Why are secondary metabolites (natural products) biosynthesized? J. Nat. Prod. 1989, 52, 1189–1208. [Google Scholar] [CrossRef] [PubMed]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and role of fungal secondary metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef]
- Gareis, M.; Gottschalk, C. Stachybotrys spp. and the guttation phenomenon. Mycotoxin Res. 2014, 30, 151–159. [Google Scholar] [CrossRef]
- Bayram, Ö. Fungal Jewels: Secondary Metabolites; Frontiers Media SA: Lausanne, Switzerland, 2017; ISBN 978-2-88945-136-4. [Google Scholar]
- Ivanoff, S.S. Guttation injuries of plants. Bot. Rev. 1963, 29, 202–229. [Google Scholar] [CrossRef]
- Sun, Y.-P.; Unestam, T.; Lucas, S.D.; Johanson, K.J.; Kenne, L.; Finlay, R. Exudation-reabsorption in a mycorrhizal fungus, the dynamic interface for interaction with soil and soil microorganisms. Mycorrhiza 1999, 9, 137–144. [Google Scholar] [CrossRef]
- Thom, C. The Penicillia; The Williams & Wilkins Company: Baltimore, MD, USA, 1930. [Google Scholar]
- Colotelo, N. Fungal exudates. Can. J. Microbiol. 1978, 24, 1173–1181. [Google Scholar] [CrossRef]
- Gareis, M.; Gareis, E.-M. Guttation droplets of Penicillium nordicum and Penicillium verrucosum contain high concentrations of the mycotoxins ochratoxin A and B. Mycopathologia 2007, 163, 207–214. [Google Scholar] [CrossRef]
- McPhee, W.; Colotelo, N. Fungal exudates. I. Characteristics of hyphal exudates in Fusarium culmorum. Can. J. Bot. 1977, 55, 358–365. [Google Scholar] [CrossRef]
- Georgiou, C.D.; Patsoukis, N.; Papapostolou, I.; Zervoudakis, G. Sclerotial metamorphosis in filamentous fungi is induced by oxidative stress. Integr. Comp. Biol. 2006, 46, 691–712. [Google Scholar] [CrossRef]
- Jennings, D.H. The role of droplets in helping to maintain a constant growth rate of aerial hyphae. Mycol. Res. 1991, 95, 883–884. [Google Scholar] [CrossRef]
- Hutwimmer, S.; Wang, H.; Strasser, H.; Burgstaller, W. Formation of exudate droplets by Metarhizium anisopliae and the presence of destruxins. Mycologia 2010, 102, 1–10. [Google Scholar] [CrossRef]
- Wang, D.; Fu, J.F.; Zhou, R.J.; Li, Z.B.; Xie, Y.J. Proteomics research and related functional classification of liquid sclerotial exudates of Sclerotinia ginseng. PeerJ 2017, 5, e3979. [Google Scholar] [CrossRef][Green Version]
- Aliferis, K.A.; Jabaji, S. Metabolite composition and bioactivity of Rhizoctonia solani sclerotial exudates. J. Agric. Food Chem. 2010, 58, 7604–7615. [Google Scholar] [CrossRef]
- Figueroa, M.; Jarmusch, A.K.; Raja, H.A.; El-Elimat, T.; Kavanaugh, J.S.; Horswill, A.R.; Cooks, R.G.; Cech, N.B.; Oberlies, N.H. Polyhydroxyanthraquinones as quorum sensing inhibitors from the guttates of Penicillium restrictum and their analysis by desorption electrospray ionization mass spectrometry. J. Nat. Prod. 2014, 77, 1351–1358. [Google Scholar] [CrossRef]
- Castagnoli, E.; Marik, T.; Mikkola, R.; Kredics, L.; Andersson, M.A.; Salonen, H.; Kurnitski, J. Indoor Trichoderma strains emitting peptaibols in guttation droplets. J. Appl. Microbiol. 2018, 125, 1408–1422. [Google Scholar] [CrossRef]
- Salo, M.J.; Marik, T.; Mikkola, R.; Andersson, M.A.; Kredics, L.; Salonen, H.; Kurnitski, J. Penicillium expansum strain isolated from indoor building material was able to grow on gypsum board and emitted guttation droplets containing chaetoglobosins and communesins A, B and D. J. Appl. Microbiol. 2019, 127, 1135–1147. [Google Scholar] [CrossRef]
- Wang, H.; Yang, X.; Wei, S.; Wang, Y. Proteomic analysis of mycelial exudates of Ustilaginoidea virens. Pathogens 2021, 10, 364. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Chen, C.; Sun, H.; Wang, Z.; Steinkellner, S.; Feng, J.; Liang, Y. Proteomic analysis reveals the importance of exudates on sclerotial development in Sclerotinia sclerotiorum. J. Agric. Food Chem. 2021, 69, 1430–1440. [Google Scholar] [CrossRef]
- Andersson, M.A.; Salo, J.; Kedves, O.; Kredics, L.; Druzhinina, I.; Kurnitski, J.; Salonen, H. Bioreactivity, guttation and agents influencing surface tension of water emitted by actively growing indoor mould isolates. Microorganisms 2020, 8, 1940. [Google Scholar] [CrossRef] [PubMed]
- Holzknecht, J.; Kühbacher, A.; Papp, C.; Farkas, A.; Váradi, G.; Marcos, J.F.; Manzanares, P.; Tóth, G.K.; Galgóczy, L.; Marx, F. The Penicillium chrysogenum Q176 antimicrobial protein PAFC effectively inhibits the growth of the opportunistic human pathogen Candida albicans. J. Fungi 2020, 6, 141. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, S.; Yang, X.; Liu, W.; Zhu, L. Proteomic analysis of exudate of Cercospora armoraciae from Armoracia rusticana. PeerJ 2020, 8, e9592. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Miao, Y.; Rahimi, M.J.; Zhu, H.; Steindorff, A.; Schiessler, S.; Cai, F.; Pang, G.; Chenthamara, K.; Xu, Y.; et al. Guttation capsules containing hydrogen peroxide: An evolutionarily conserved NADPH oxidase gains a role in wars between related fungi. Environ. Microbiol. 2019, 21, 2644–2658. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Kreuzenbeck, N.B.; Otani, S.; Garcia-Altares, M.; Dahse, H.-M.; Weigel, C.; Aanen, D.K.; Hertweck, C.; Poulsen, M.; Beemelmanns, C. Pseudoxylallemycins A-F, cyclic tetrapeptides with rare allenyl modifications isolated from Pseudoxylaria sp. X802: A competitor of fungus-growing termite cultivars. Org. Lett. 2016, 18, 3338–3341. [Google Scholar] [CrossRef] [PubMed]
- Sica, V.P.; Rees, E.R.; Tchegnon, E.; Bardsley, R.H.; Raja, H.A.; Oberlies, N.H. Spatial and temporal profiling of griseofulvin production in Xylaria cubensis using mass spectrometry mapping. Front. Microbiol. 2016, 7, 544. [Google Scholar] [CrossRef] [PubMed]
- Caraballo-Rodríguez, A.M.; Mayor, C.A.; Chagas, F.O.; Pupo, M.T. Amphotericin B as an inducer of griseofulvin-containing guttate in the endophytic fungus Xylaria cubensis FLe9. Chemoecology 2017, 27, 177–185. [Google Scholar] [CrossRef]
- Sica, V.P.; Figueroa, M.; Raja, H.A.; El-Elimat, T.; Darveaux, B.A.; Pearce, C.J.; Oberlies, N.H. Optimizing production and evaluating biosynthesis in situ of a herbicidal compound, mevalocidin, from Coniolariella sp. J. Ind. Microbiol. Biotechnol. 2016, 43, 1149–1157. [Google Scholar] [CrossRef]
- Sica, V.P.; Raja, H.A.; El-Elimat, T.; Kertesz, V.; Van Berkel, G.J.; Pearce, C.J.; Oberlies, N.H. Dereplicating and spatial mapping of secondary metabolites from fungal cultures in situ. J. Nat. Prod. 2015, 78, 1926–1936. [Google Scholar] [CrossRef]
- Calder, C.; Ford, S.; Selwood, A.I.; Ginkel, R.V.; Wilkins, A.L. Anti-Microbial Compositions. WO Patent WO 2012/023865, 23 February 2012. [Google Scholar]
- Lee, A.J.; Cadelis, M.M.; Kim, S.H.; Swift, S.; Copp, B.R.; Villas-Boas, S.G. Epipyrone A, a broad-spectrum antifungal compound produced by Epicoccum nigrum ICMP 19927. Molecules 2020, 25, 5997. [Google Scholar] [CrossRef]
- Muñoz, K.; Vega, M.; Rios, G.; Geisen, R.; Degen, G.H. Mycotoxin production by different ochratoxigenic Aspergillus and Penicillium species on coffee- and wheat-based media. Mycotoxin Res. 2011, 27, 239–247. [Google Scholar] [CrossRef]
- Liang, Y.; Strelkov, S.E.; Kav, N.N.V. The proteome of liquid sclerotial exudates from Sclerotinia sclerotiorum. J. Proteome Res. 2010, 9, 3290–3298. [Google Scholar] [CrossRef]
- Wang, X.; Sena Filho, J.G.; Hoover, A.R.; King, J.B.; Ellis, T.K.; Powell, D.R.; Cichewicz, R.H. Chemical epigenetics alters the secondary metabolite composition of guttate excreted by an atlantic-forest-soil-derived Penicillium citreonigrum. J. Nat. Prod. 2010, 73, 942–948. [Google Scholar] [CrossRef]
- Pandey, M.K.; Sarma, B.K.; Singh, D.P.; Singh, U.P. Biochemical investigations of sclerotial exudates of Sclerotium rolfsii and their antifungal activity. J. Phytopathol. 2007, 155, 84–89. [Google Scholar] [CrossRef]
- Olsen, J.G.; Flensburg, C.; Olsen, O.; Bricogne, G.; Henriksen, A. Solving the structure of the bubble protein using the anomalous sulfur signal from single-crystal in-house Cu Kα diffraction data only. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Seibold, M.; Wolschann, P.; Bodevin, S.; Olsen, O. Properties of the bubble protein, a defensin and an abundant component of a fungal exudate. Peptides 2011, 32, 1989–1995. [Google Scholar] [CrossRef]
- Grovel, O.; Pouchus, Y.F.; Verbist, J.-F. Accumulation of gliotoxin, a cytotoxic mycotoxin from Aspergillus fumigatus, in blue mussel (Mytilus edulis). Toxicon 2003, 42, 297–300. [Google Scholar] [CrossRef]
- Kerzaon, I.; Grovel, O.; Robiou Du Pont, T.; Le Pape, P.; Pouchus, Y.-F. Effects of seawater on growth and gliotoxin excretion of marine strains of Aspergillus fumigatus Fres. Toxicon Off. J. Int. Soc. Toxinol. 2008, 51, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Colotelo, N.; Sumner, J.L.; Voegelin, W.S. Chemical studies on the exudate and developing sclerotia of Sclerotinia sclerotiorum (Lib.) DeBary. Can. J. Microbiol. 1971, 17, 1189–1194. [Google Scholar] [CrossRef]
- Döll, K.; Chatterjee, S.; Scheu, S.; Karlovsky, P.; Rohlfs, M. Fungal metabolic plasticity and sexual development mediate induced resistance to arthropod fungivory. Proc. Biol. Sci. 2013, 280, 20131219. [Google Scholar] [CrossRef]
- Colotelo, N. Physiological and biochemical properties of the exudate associated with developing sclerotia of Sclerotinia sclerotiorum (Lib.) DeBary. Can. J. Microbiol. 1973, 19, 73–79. [Google Scholar] [CrossRef]
- Reddy, M.N.; Anandaraj, G.V.D.; Devi, P.S. Chemical composition of sclerotial exudates of Macrophomina phaseolina pathogenic to groundnut. Soil Biol. Biochem. 1989, 21, 931–933. [Google Scholar] [CrossRef]
- Sarma, B.K.; Singh, U.P.; Singh, K.P. Variability in Indian isolates of Sclerotium rolfsii. Mycologia 2002, 94, 1051–1058. [Google Scholar] [CrossRef]
- Coggins, C.R.; Jennings, D.H.; Clarke, R.W. Tear or drop formation by mycelium of Serpula lacrimans. Trans. Br. Mycol. Soc. 1980, 75, 63–67. [Google Scholar] [CrossRef]
- Cooke, R.C. Changes in soluble carbohydrates during sclerotium formation by Sclerotinia sclerotiorum and S. trifoliorum. Trans. Br. Mycol. Soc. 1969, 53, 77–86. [Google Scholar] [CrossRef]
- Ríos-Moreno, A.; Garrido-Jurado, I.; Raya-Ortega, M.C.; Quesada-Moraga, E. Quantification of fungal growth and destruxin A during infection of Galleria mellonella larvae by Metarhizium brunneum. J. Invertebr. Pathol. 2017, 149, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Rohlfs, M. Fungal secondary metabolite dynamics in fungus-grazer interactions: Novel insights and unanswered questions. Front. Microbiol. 2014, 5, 788. [Google Scholar] [CrossRef][Green Version]
- Zhu, H.; Zhang, J.; Gao, Q.; Pang, G.; Sun, T.; Li, R.; Yu, Z.; Shen, Q. A new atypical short-chain dehydrogenase is required for interfungal combat and conidiation in Trichoderma guizhouense. Environ. Microbiol. 2021. [Google Scholar] [CrossRef]
- Karwehl, S.; Stadler, M. Exploitation of fungal biodiversity for discovery of novel antibiotics. Curr. Top. Microbiol. Immunol. 2016, 398, 303–338. [Google Scholar] [CrossRef]
- Durand, G.A.; Raoult, D.; Dubourg, G. Antibiotic discovery: History, methods and perspectives. Int. J. Antimicrob. Agents 2019, 53, 371–382. [Google Scholar] [CrossRef]
- Van Duin, D.; Paterson, D.L. Multidrug-resistant bacteria in the community: An update. Infect. Dis. Clin. N. Am. 2020, 34, 709–722. [Google Scholar] [CrossRef]
- Terreni, M.; Taccani, M.; Pregnolato, M. New antibiotics for multidrug-resistant bacterial strains: Latest research developments and future perspectives. Molecules 2021, 26, 2671. [Google Scholar] [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schäberle, T.F.; Hughes, D.E.; Epstein, S.; et al. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef]
- Duclohier, H. Antimicrobial peptides and peptaibols, substitutes for conventional antibiotics. Curr. Pharm. Des. 2010, 16, 3212–3223. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.F.S.; Filho, E.R. Peptaibols of Trichoderma. Nat. Prod. Rep. 2007, 24, 1128–1141. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Sun, L.; Huang, S.; Zhu, C.; Li, P.; He, J.; Mackey, V.; Coy, D.H.; He, Q. The antimicrobial peptides and their potential clinical applications. Am. J. Transl. Res. 2019, 11, 3919–3931. [Google Scholar] [PubMed]
- Rivera-Chávez, J.; Raja, H.A.; Graf, T.N.; Gallagher, J.M.; Metri, P.; Xue, D.; Pearce, C.J.; Oberlies, N.H. Prealamethicin F50 and related peptaibols from Trichoderma arundinaceum: Validation of their authenticity via in situ chemical analysis. RSC Adv. 2017, 7, 45733–45751. [Google Scholar] [CrossRef] [PubMed]
- Dickey, S.W.; Cheung, G.Y.C.; Otto, M. Different drugs for bad bugs: Antivirulence strategies in the age of antibiotic resistance. Nat. Rev. Drug Discov. 2017, 16, 457–471. [Google Scholar] [CrossRef]
- Defoirdt, T. Quorum-sensing systems as targets for antivirulence therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef]
- Daly, S.M.; Elmore, B.O.; Kavanaugh, J.S.; Triplett, K.D.; Figueroa, M.; Raja, H.A.; El-Elimat, T.; Crosby, H.A.; Femling, J.K.; Cech, N.B.; et al. ω-Hydroxyemodin limits Staphylococcus aureus quorum sensing-mediated pathogenesis and inflammation. Antimicrob. Agents Chemother. 2015, 59, 2223–2235. [Google Scholar] [CrossRef]
- Pedras, M.S.C.; Irina Zaharia, L.; Ward, D.E. The destruxins: Synthesis, biosynthesis, biotransformation, and biological activity. Phytochemistry 2002, 59, 579–596. [Google Scholar] [CrossRef]
- Liu, B.-L.; Tzeng, Y.-M. Development and applications of destruxins: A review. Biotechnol. Adv. 2012, 30, 1242–1254. [Google Scholar] [CrossRef]
- Yeh, S.F.; Pan, W.; Ong, G.T.; Chiou, A.J.; Chuang, C.C.; Chiou, S.H.; Wu, S.H. Study of structure-activity correlation in destruxins, a class of cyclodepsipeptides possessing suppressive effect on the generation of hepatitis B virus surface antigen in human hepatoma cells. Biochem. Biophys. Res. Commun. 1996, 229, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.C.; Chou, C.K.; Sun, C.M.; Yeh, S.F. Suppressive effects of destruxin B on hepatitis B virus surface antigen gene expression in human hepatoma cells. Antivir. Res. 1997, 34, 137–144. [Google Scholar] [CrossRef]
- Yuan, B.; Wu, Z.; Ji, W.; Liu, D.; Guo, X.; Yang, D.; Fan, A.; Jia, H.; Ma, M.; Lin, W. Discovery of cyclohexadepsipeptides with anti-Zika virus activities and biosynthesis of the nonproteinogenic building block (3S)-methyl-l-proline. J. Biol. Chem. 2021, 297, 100822. [Google Scholar] [CrossRef] [PubMed]
- Che, Y.; Swenson, D.C.; Gloer, J.B.; Koster, B.; Malloch, D. Pseudodestruxins A and B: New cyclic depsipeptides from the coprophilous fungus Nigrosabulum globosum. J. Nat. Prod. 2001, 64, 555–558. [Google Scholar] [CrossRef]
- Dornetshuber-Fleiss, R.; Heffeter, P.; Mohr, T.; Hazemi, P.; Kryeziu, K.; Seger, C.; Berger, W.; Lemmens-Gruber, R. Destruxins: Fungal-derived cyclohexadepsipeptides with multifaceted anticancer and antiangiogenic activities. Biochem. Pharmacol. 2013, 86, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, B.C.; Brewster, W.K.; Deboer, G.J.; Fields, S.C.; Graupner, P.R.; Hahn, D.R.; Pearce, C.J.; Schmitzer, P.R.; Webster, J.D. Mevalocidin: A novel, phloem mobile phytotoxin from Fusarium DA056446 and Rosellinia DA092917. J. Chem. Ecol. 2013, 39, 253–261. [Google Scholar] [CrossRef]
- Park, J.H.; Choi, G.J.; Lee, H.B.; Kim, K.M.; Jung, H.S.; Lee, S.W.; Jang, K.S.; Cho, K.Y.; Kim, J.C. Griseofulvin from Xylaria sp. strain F0010, an endophytic fungus of Abies holophylla and its antifungal activity against plant pathogenic fungi. J. Microbiol. Biotechnol. 2005, 15, 112–117. [Google Scholar]
- Brooks, W.C.; Paguigan, N.D.; Raja, H.A.; Moy, F.J.; Cech, N.B.; Pearce, C.J.; Oberlies, N.H. qNMR for profiling the production of fungal secondary metabolites. Magn. Reson. Chem. MRC 2017, 55, 670–676. [Google Scholar] [CrossRef]
- Knowles, S.L.; Raja, H.A.; Wright, A.J.; Lee, A.M.L.; Caesar, L.K.; Cech, N.B.; Mead, M.E.; Steenwyk, J.L.; Ries, L.N.A.; Goldman, G.H.; et al. Mapping the fungal battlefield: Using in situ chemistry and deletion mutants to monitor interspecific chemical interactions between fungi. Front. Microbiol. 2019, 10, 285. [Google Scholar] [CrossRef]
- Oberlies, N.H.; Knowles, S.L.; Amrine, C.S.M.; Kao, D.; Kertesz, V.; Raja, H.A. Droplet probe: Coupling chromatography to the in situ evaluation of the chemistry of nature. Nat. Prod. Rep. 2019, 36, 944–959. [Google Scholar] [CrossRef]
- Sudakin, D.L. Biopesticides. Toxicol. Rev. 2003, 22, 83–90. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Duke, S.O. Rationale for a natural products approach to herbicide discovery. Pest Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef]
- Mascarin, G.M.; Jaronski, S.T. The production and uses of Beauveria bassiana as a microbial insecticide. World J. Microbiol. Biotechnol. 2016, 32, 177. [Google Scholar] [CrossRef] [PubMed]
- Stone, L.B.L.; Bidochka, M.J. The multifunctional lifestyles of Metarhizium: Evolution and applications. Appl. Microbiol. Biotechnol. 2020, 104, 9935–9945. [Google Scholar] [CrossRef]
- Lozano-Tovar, M.D.; Garrido-Jurado, I.; Lafont, F.; Quesada-Moraga, E. Insecticidal activity of a destruxin-containing extract of Metarhizium brunneum against Ceratitis capitata (Diptera: Tephritidae). J. Econ. Entomol. 2015, 108, 462–472. [Google Scholar] [CrossRef]
- Oliveira, D.G.P.; Pauli, G.; Mascarin, G.M.; Delalibera, I. A protocol for determination of conidial viability of the fungal entomopathogens Beauveria bassiana and Metarhizium anisopliae from commercial products. J. Microbiol. Methods 2015, 119, 44–52. [Google Scholar] [CrossRef]
- Calcagno, C.; Novero, M.; Genre, A.; Bonfante, P.; Lanfranco, L. The exudate from an arbuscular mycorrhizal fungus induces nitric oxide accumulation in Medicago truncatula roots. Mycorrhiza 2012, 22, 259–269. [Google Scholar] [CrossRef]
- Vujanovic, V.; Germida, J.J. Endophytic Microbial Symbionts in Plant Prenatal Care 2019. U.S. Patent 10,212,912, 26 February 2019. [Google Scholar]
- Taevernier, L.; Wynendaele, E.; De Vreese, L.; Burvenich, C.; De Spiegeleer, B. The mycotoxin definition reconsidered towards fungal cyclic depsipeptides. J. Environ. Sci. Health Part C Environ. Carcinog. Ecotoxicol. Rev. 2016, 34, 114–135. [Google Scholar] [CrossRef] [PubMed]
- Pleadin, J.; Frece, J.; Markov, K. Mycotoxins in food and feed. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Elsevier: Amsterdam, The Netherlands, 2019; Volume 89, pp. 297–345. [Google Scholar]
- Richard, J.L. Some major mycotoxins and their mycotoxicoses–An overview. Int. J. Food Microbiol. 2007, 119, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Foroud, N.A.; Eudes, F. Trichothecenes in cereal grains. Int. J. Mol. Sci. 2009, 10, 147–173. [Google Scholar] [CrossRef]
- McCormick, S.P.; Stanley, A.M.; Stover, N.A.; Alexander, N.J. Trichothecenes: From simple to complex mycotoxins. Toxins 2011, 3, 802–814. [Google Scholar] [CrossRef]
- De Carvalho, M.P.; Weich, H.; Abraham, W.-R. Macrocyclic trichothecenes as antifungal and anticancer compounds. Curr. Med. Chem. 2016, 23, 23–35. [Google Scholar] [CrossRef]
- Zhu, M.; Cen, Y.; Ye, W.; Li, S.; Zhang, W. Recent advances on macrocyclic trichothecenes, their bioactivities and biosynthetic pathway. Toxins 2020, 12, 417. [Google Scholar] [CrossRef]
- Forner, A.; Llovet, J.M.; Bruix, J. Hepatocellular carcinoma. Lancet Lond. Engl. 2012, 379, 1245–1255. [Google Scholar] [CrossRef]
- Shi, M.; Wang, H.-N.; Xie, S.-T.; Luo, Y.; Sun, C.-Y.; Chen, X.-L.; Zhang, Y.-Z. Antimicrobial peptaibols, novel suppressors of tumor cells, targeted calcium-mediated apoptosis and autophagy in human hepatocellular carcinoma cells. Mol. Cancer 2010, 9, 26. [Google Scholar] [CrossRef]
- Shi, M.; Zhang, T.; Sun, L.; Luo, Y.; Liu, D.-H.; Xie, S.-T.; Song, X.-Y.; Wang, G.-F.; Chen, X.-L.; Zhou, B.-C.; et al. Calpain, Atg5 and Bak play important roles in the crosstalk between apoptosis and autophagy induced by influx of extracellular calcium. Apoptosis Int. J. Program. Cell Death 2013, 18, 435–451. [Google Scholar] [CrossRef]
- Chugh, J.K.; Wallace, B.A. Peptaibols: Models for ion channels. Biochem. Soc. Trans. 2001, 29, 565–570. [Google Scholar] [CrossRef]
- Marik, T.; Tyagi, C.; Balázs, D.; Urbán, P.; Szepesi, Á.; Bakacsy, L.; Endre, G.; Rakk, D.; Szekeres, A.; Andersson, M.A.; et al. Structural diversity and bioactivities of peptaibol compounds from the Longibrachiatum Clade of the filamentous fungal genus Trichoderma. Front. Microbiol. 2019, 10, 1434. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Janso, J.E.; Yang, H.Y.; Bernan, V.S.; Lin, S.L.; Yu, K. Culicinin D, an antitumor peptaibol produced by the fungus Culicinomyces clavisporus, strain LL-12I252. J. Nat. Prod. 2006, 69, 736–741. [Google Scholar] [CrossRef]
- Rogozhin, E.A.; Sadykova, V.S.; Baranova, A.A.; Vasilchenko, A.S.; Lushpa, V.A.; Mineev, K.S.; Georgieva, M.L.; Kul’ko, A.B.; Krasheninnikov, M.E.; Lyundup, A.V.; et al. A novel lipopeptaibol emericellipsin A with antimicrobial and antitumor activity produced by the extremophilic fungus Emericellopsis alkalina. Molecules 2018, 23, 2785. [Google Scholar] [CrossRef] [PubMed]
- Grigoletto, D.F.; Trivella, D.B.B.; Tempone, A.G.; Rodrigues, A.; Correia, A.M.L.; Lira, S.P. Antifungal compounds with anticancer potential from Trichoderma sp. P8BDA1F1, an endophytic fungus from Begonia venosa. Braz. J. Microbiol. Publ. Braz. Soc. Microbiol. 2020, 51, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Sica, V.P.; Rees, E.R.; Raja, H.A.; Rivera-Chávez, J.; Burdette, J.E.; Pearce, C.J.; Oberlies, N.H. In situ mass spectrometry monitoring of fungal cultures led to the identification of four peptaibols with a rare threonine residue. Phytochemistry 2017, 143, 45–53. [Google Scholar] [CrossRef]
- Guo, H.; Schmidt, A.; Stephan, P.; Raguž, L.; Braga, D.; Kaiser, M.; Dahse, H.-M.; Weigel, C.; Lackner, G.; Beemelmanns, C. Precursor-directed diversification of cyclic tetrapeptidic pseudoxylallemycins. Chembiochem. Eur. J. Chem. Biol. 2018, 19, 2307–2311. [Google Scholar] [CrossRef]
- Kobayashi, T.; Ikeno, S.; Hosokawa, N.; Uehara, Y.; Hori, M.; Tsuchiya, K. Destruxin E, a cyclodepsipeptide antibiotic, reduces cyclin D1 levels and inhibits anchorage-independent growth of v-Ki-ras-expressed pMAM-ras-REF cells. Biol. Pharm. Bull. 2004, 27, 587–590. [Google Scholar] [CrossRef][Green Version]
- Huynh, T.-T.; Rao, Y.K.; Lee, W.-H.; Chen, H.-A.; Le, T.D.-Q.; Tzeng, D.T.W.; Wang, L.-S.; Wu, A.T.H.; Lin, Y.-F.; Tzeng, Y.-M.; et al. Destruxin B inhibits hepatocellular carcinoma cell growth through modulation of the Wnt/β-catenin signaling pathway and epithelial-mesenchymal transition. Toxicol. In Vitro Int. J. Publ. Assoc. BIBRA 2014, 28, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.-T.; Rao, Y.K.; Ye, M.; Wu, W.-S.; Chang, T.-C.; Wang, L.-S.; Wu, C.-H.; Wu, A.T.H.; Tzeng, Y.-M. Preclinical evaluation of destruxin B as a novel Wnt signaling target suppressing proliferation and metastasis of colorectal cancer using non-invasive bioluminescence imaging. Toxicol. Appl. Pharmacol. 2012, 261, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-P.; Wang, C.-W.; Liao, W.-C.; Yang, C.-R.; Yeh, C.-T.; Tsai, C.-H.; Yang, C.-C.; Tzeng, Y.-M. In vitro and in vivo anticancer effects of destruxin B on human colorectal cancer. Anticancer Res. 2012, 32, 2735–2745. [Google Scholar]
- Wu, C.-C.; Chen, T.-H.; Liu, B.-L.; Wu, L.-C.; Chen, Y.-C.; Tzeng, Y.-M.; Hsu, S.-L. Destruxin B isolated from entomopathogenic fungus Metarhizium anisopliae induces apoptosis via a Bcl-2 family-dependent mitochondrial pathway in human nonsmall cell lung cancer cells. Evid. Based Complement. Altern. Med. ECAM 2013, 2013, 548929. [Google Scholar] [CrossRef]
- Huang Liu, R.; Chen, S.-P.; Lu, T.-M.; Tsai, W.-Y.; Tsai, C.-H.; Yang, C.-C.; Tzeng, Y.-M. Selective apoptotic cell death effects of oral cancer cells treated with destruxin B. BMC Complement. Altern. Med. 2014, 14, 207. [Google Scholar] [CrossRef] [PubMed]
- Jones, D. Ultrastructure and composition of the cell walls of Sclerotinia sclerotiorum. Trans. Br. Mycol. Soc. 1970, 54, 351-IN6. [Google Scholar] [CrossRef]
- Wong, J.H.; Xia, L.; Ng, T.B. A review of defensins of diverse origins. Curr. Protein Pept. Sci. 2007, 8, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Wolf, J.M.; Espadas, J.; Luque-Garcia, J.; Reynolds, T.; Casadevall, A. Lipid biosynthetic genes affect Candida albicans extracellular vesicle morphology, cargo, and immunostimulatory properties. Eukaryot. Cell 2015, 14, 745–754. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
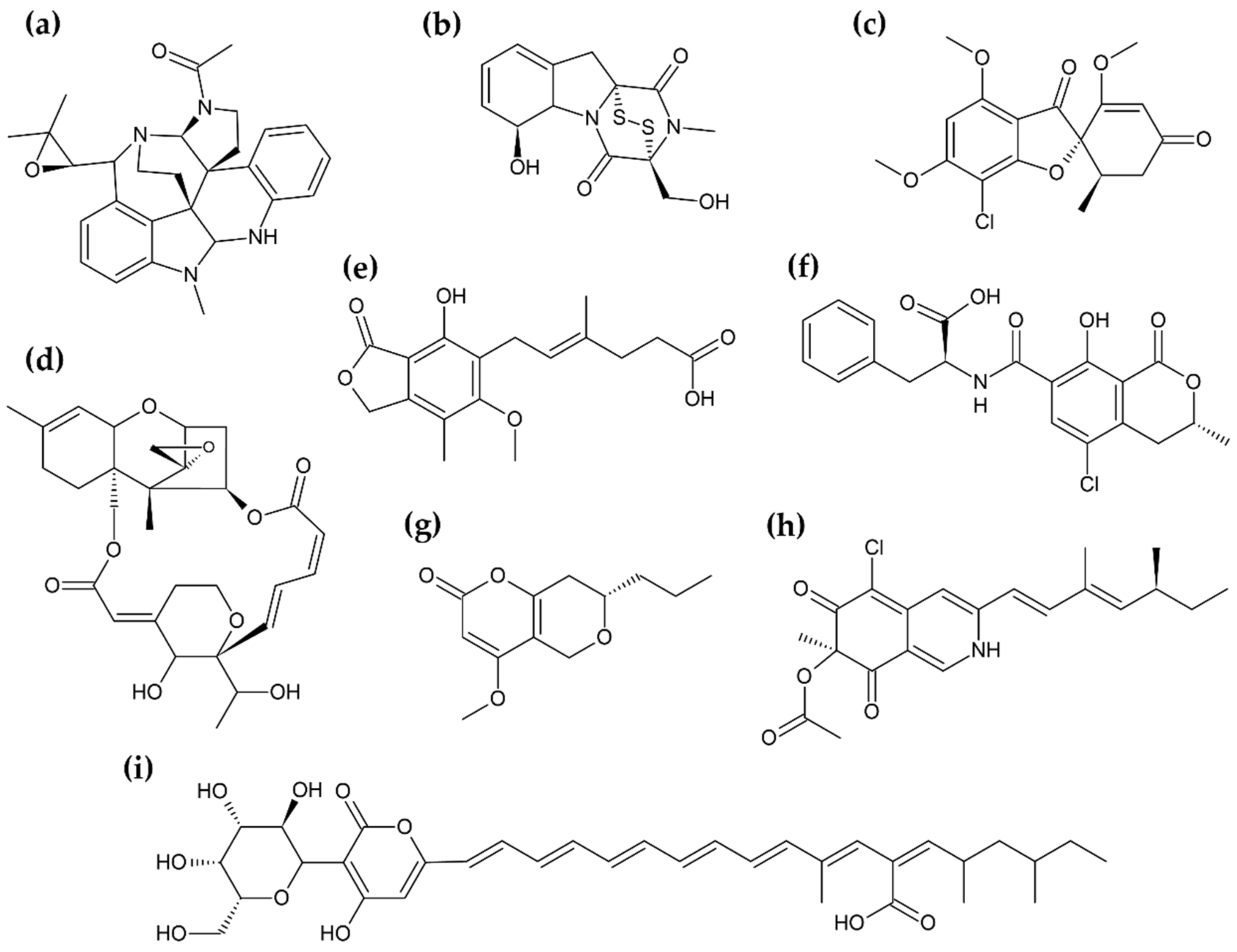{kind=link}
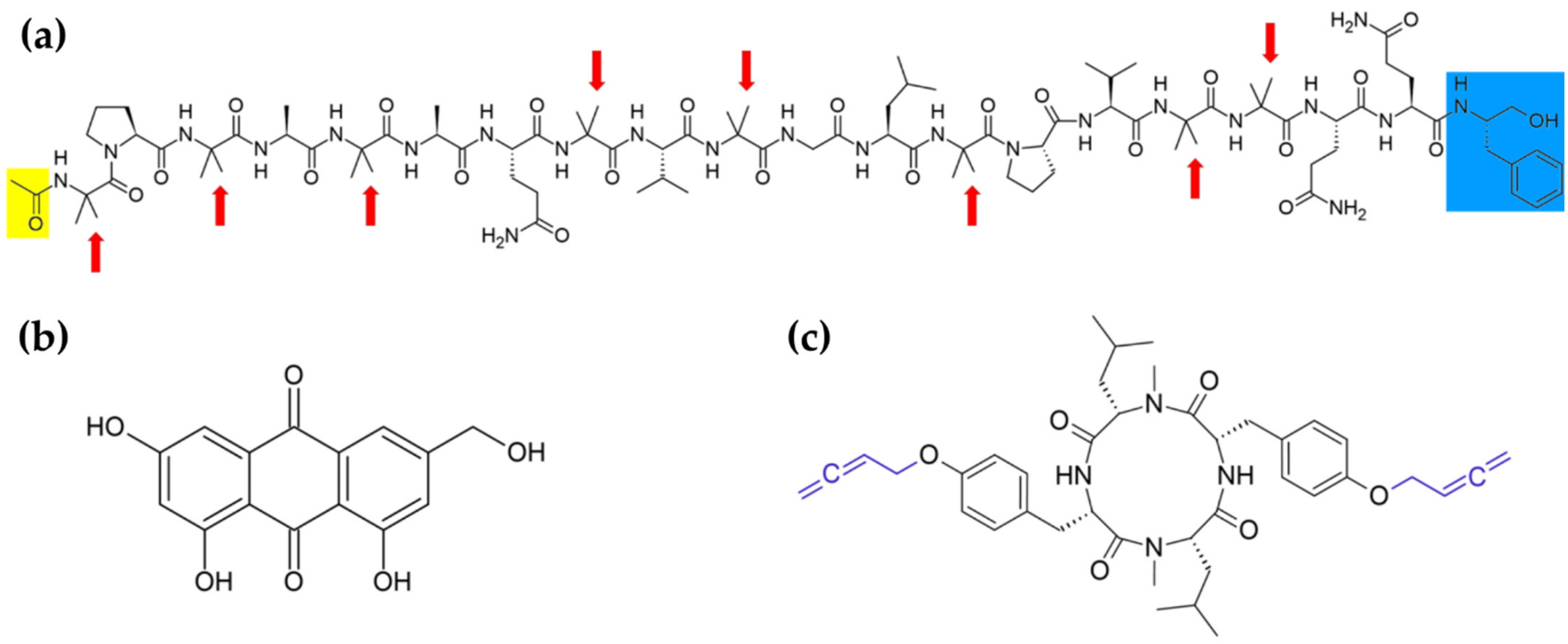{kind=link}
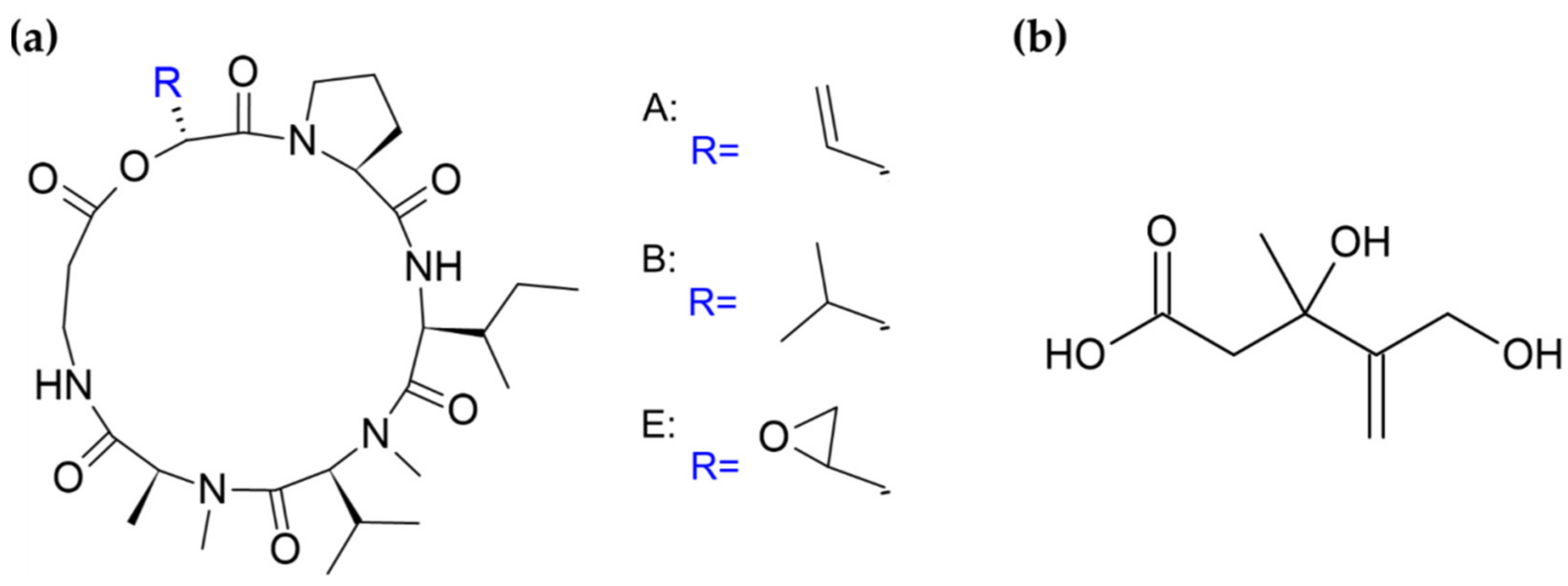{kind=link}
| Fungus Name | Growth Medium | Incubation T | Identified Compounds | Bioactivity | Reference |
|---|---|---|---|---|---|
| Sclerotinia sclerotiorum | PDA | 25 °C | proteins (amylases, cellulases, hemicellulases, polygalacturonases) | plant cell wall degrading (enzymatic) | [22] |
| Ustilaginoidea virens | PSA | 25 ± 1 °C | proteins (acetylxylan esterase, cellobiose dehydrogenase, endo-1,3(4)-beta-glucanase; superoxide dismutase) | plant cell wall degrading (enzymatic);oxidative stress generation | [21] |
| Chaetomium globosum, Penicillium expansum, Stachybotrys sp., Trichoderma atroviride, Trichoderma trixiae | MEA | 23 ± 1 °C | – | cytotoxic (mycotoxins) | [23] |
| Penicillium chrysogenum | specific | 25 °C | proteins (i.e., PAFC) | antifungal | [24] |
| Cercospora armoraciae | PDA | 25 °C ± 1 °C | proteins (putative indirect role—SMs biosynthesis enzymes) | antimicrobial, phytotoxic | [25] |
| Penicillium expansum | MEA | 23 ± 1 °C | chaetoglobosins, communesins | cytotoxic (mycotoxins) | [20] |
| Trichoderma guizhouense | specific (synthetic), coculture | 25 °C | hydrogen peroxide | oxidative stress generation (in another fungus) | [26] |
| Trichoderma atroviride, Trichoderma citrinoviride, Trichoderma paraviridescens, Trichoderma trixiae | MEA | 22 °C | peptaibols (trichorzianines, trilongins, trichostrigocins, trichostrigocin-like peptaibols) | cytotoxic (mycotoxins) | [19] |
| Sclerotinia ginseng | PDA | 20 ± 1 °C | proteins (cellulases, pectinases) | plant cell wall degrading (enzymatic) | [16] |
| Pseudoxylaria sp. | PDA, coculture | RT | pseudoxylallemycins (n = 4) | antibacterial, anticancer | [27] |
| Xylariacubensis | MEA; ISP-2 agar + amphotericin B | –; 30 °C | griseofulvin; griseofulvin, dechlorogriseofulvin | antifungal | [28,29] |
| Coniolariella sp. | agar | – | mevalocidin | phytotoxic (bioherbicide) | [30] |
| Clohesyomyces aquaticus | – | – | phomopsinone A | antifungal | [31] |
| Penicillium restrictum | PDA | 25 °C | polyhydroxyanthraquinones (n = 9) | antibacterial (QS inhibitor) | [18] |
| Stachybotryschartarum | MEA | 25 °C | macrocyclic trichothecenes (n = 6) | cytotoxic (mycotoxins) | [5] |
| Epicoccum nigrum | CYA | 25 °C | polyene compounds (epipyrones) | antifungal | [32,33] |
| Aspergillus niger, Aspergillus ochraceus | specific (coffee-, wheat-based) | 25 °C | ochratoxin A | cytotoxic (mycotoxin) | [34] |
| Rhizoctonia solani | PDA | 24 ± 1 °C | phenolic compounds | antifungal, phytotoxic | [17] |
| Sclerotinia sclerotiorum | PDA | 22 ± 1 °C | proteins (arabinofuranosidases; cerato-platanin, necrosis inducing protein) | plant cell wall degrading (enzymatic); phytotoxic | [35] |
| Metarhiziumanisopliae | specific (3 synthetic media) | 25 °C | destruxins (A, B, E) | insecticidal, phytotoxic, anticancer, etc. | [15] |
| Penicillium citreonigrum | specific + 5-azacytidine | 25 °C | sclerotioramine; sclerotiorin | antibacterial, antifungal; antibacterial | [36] |
| Penicillium nordicum, Penicillium verrucosum | CYA, MEA | 25 °C | ochratoxins (A, B) | cytotoxic (mycotoxins) | [11] |
| Sclerotium rolfsii | PDA | 25 ± 2 °C | phenolic compounds | antifungal | [37] |
| Penicillium brevicompactum | specific | 20 °C | bubble protein (defensin);mycophenolic acid | antifungal; antimicrobial | [38,39] |
| Aspergillus fumigatus | SDA + seawater;DCA + seawater | 27 °C | gliotoxin | cytotoxic (mycotoxin) | [40,41] |
| Fusarium culmorum, Penicillium claviforme, Sclerotinia sclerotiorum, Sclerotium rolfsii | PDA; specific (carrot-based, synthetic) | 20, 25 °C | proteins (cellulase, polygalacturonase) | plant cell wall degrading (enzymatic) | [10,42] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krain, A.; Siupka, P. Fungal Guttation, a Source of Bioactive Compounds, and Its Ecological Role—A Review. Biomolecules 2021, 11, 1270. https://doi.org/10.3390/biom11091270
Krain A, Siupka P. Fungal Guttation, a Source of Bioactive Compounds, and Its Ecological Role—A Review. Biomolecules. 2021; 11(9):1270. https://doi.org/10.3390/biom11091270
Chicago/Turabian StyleKrain, Adam, and Piotr Siupka. 2021. "Fungal Guttation, a Source of Bioactive Compounds, and Its Ecological Role—A Review" Biomolecules 11, no. 9: 1270. https://doi.org/10.3390/biom11091270
APA StyleKrain, A., & Siupka, P. (2021). Fungal Guttation, a Source of Bioactive Compounds, and Its Ecological Role—A Review. Biomolecules, 11(9), 1270. https://doi.org/10.3390/biom11091270