
Pregnane X Receptor (PXR) Polymorphisms and Cancer Treatment




Abstract
1. Introduction
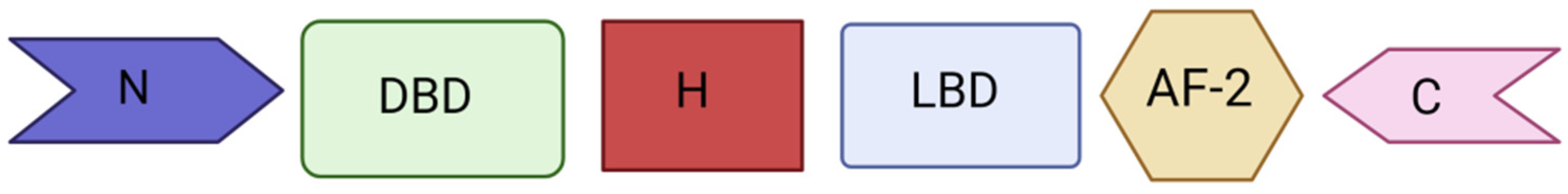
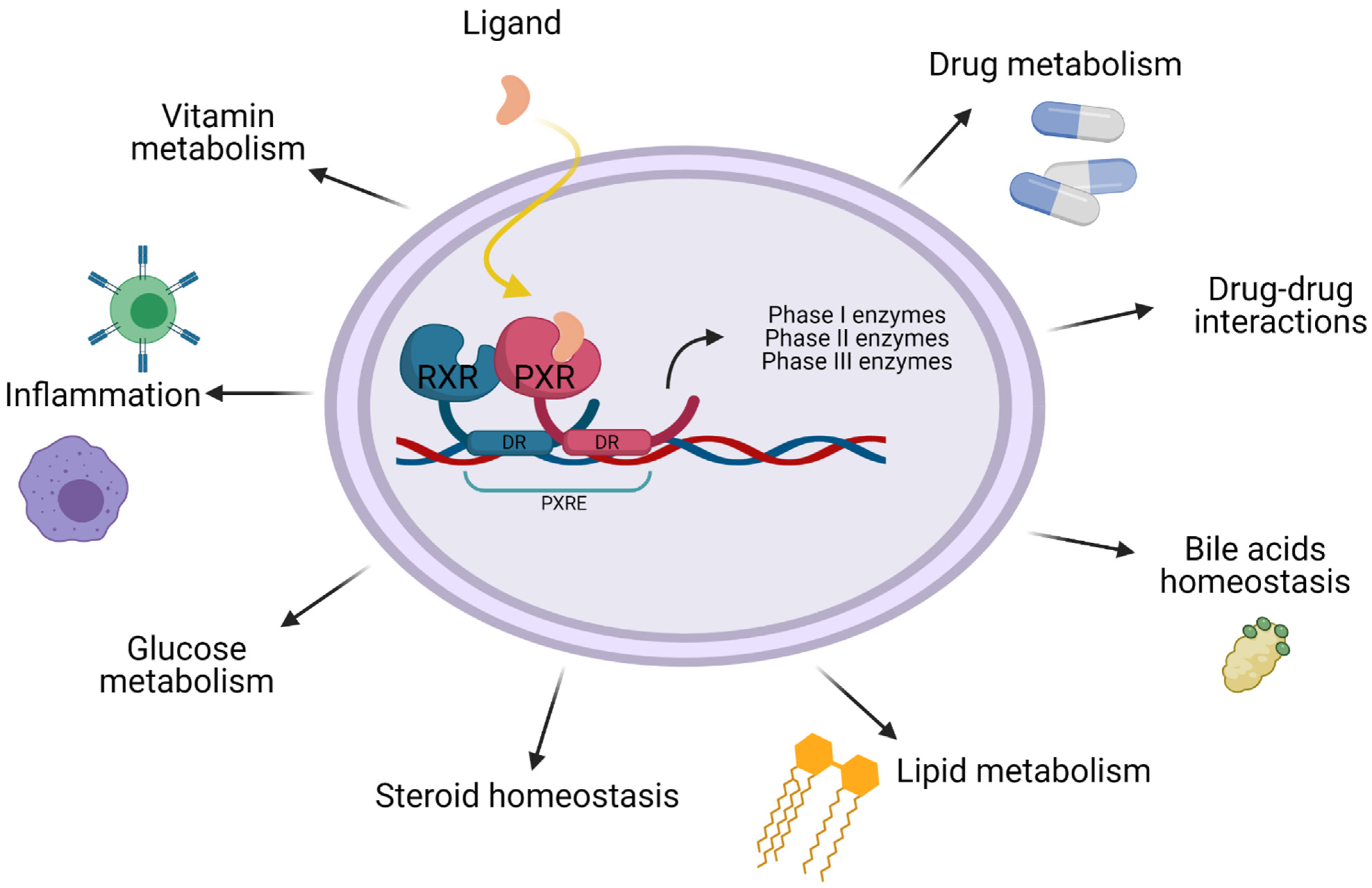
2. PXR Function
3. Regulation
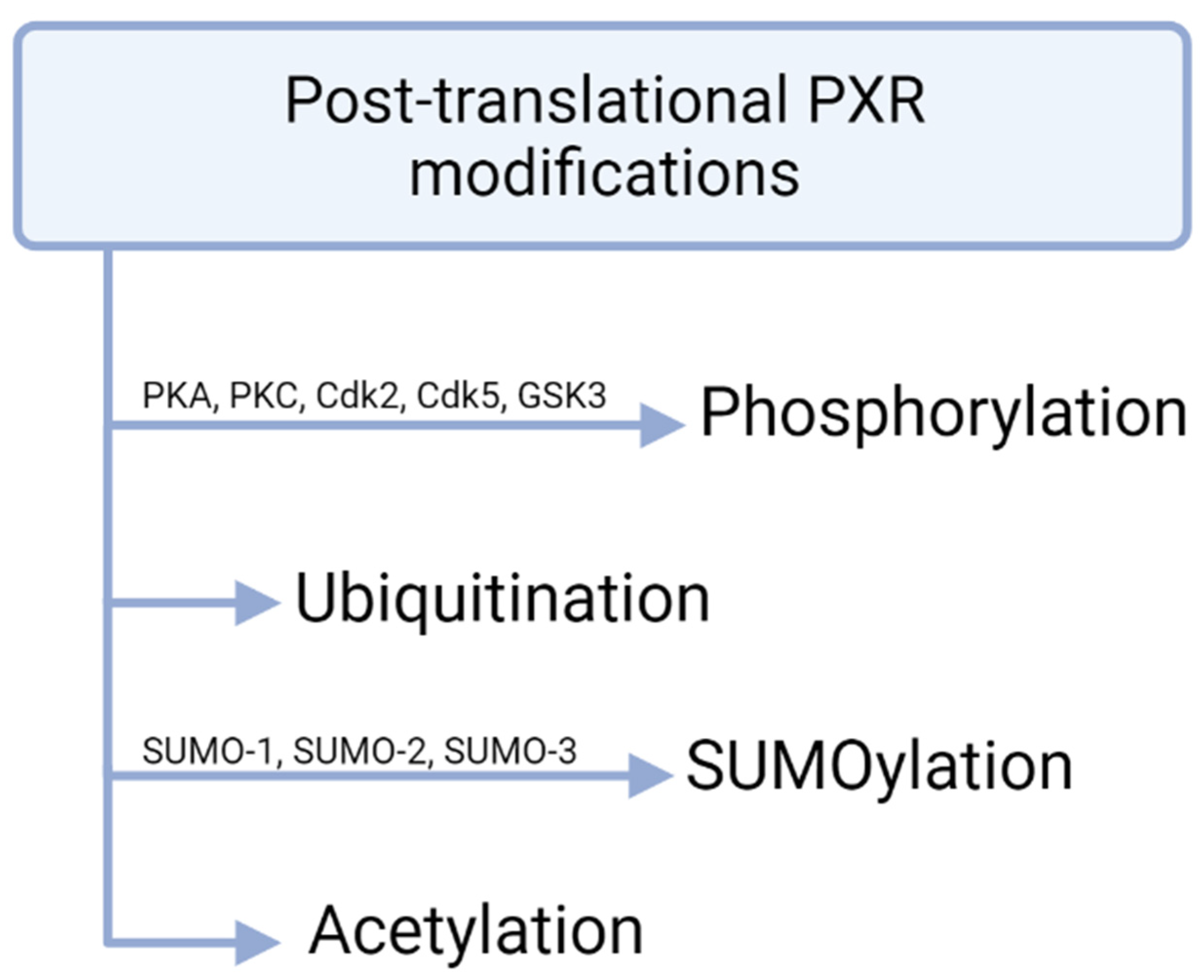
4. Post-Translational and Post-Transcriptional PXR Modifications
5. Agonists and Antagonists
6. PXR and Cancer
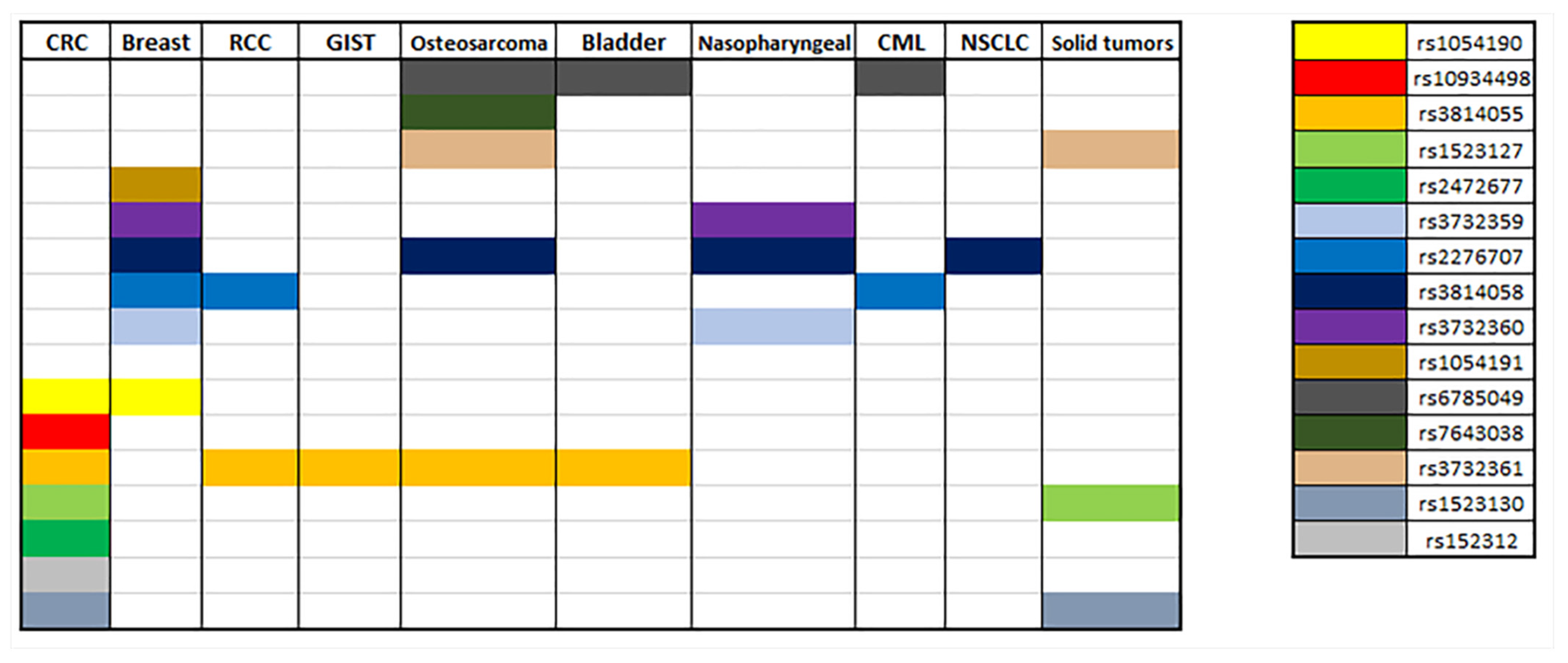
7. PXR Polymorphisms and Cancer Pharmacogenetics
7.1. Gastrointestinal Cancer
7.2. Breast Cancer
7.3. Renal Cell Carcinoma
7.4. Others
8. Discussion
9. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoshinari, K. Role of Nuclear Receptors PXR and CAR in Xenobiotic-Induced Hepatocyte Proliferation and Chemical Carcinogenesis. Biol. Pharm. Bull. 2019, 42, 1243–1252. [Google Scholar] [CrossRef]
- Kliewer, S.A.; Goodwin, B.; Willson, T.M. The Nuclear Pregnane X Receptor: A Key Regulator of Xenobiotic Metabolism. Endocr. Rev. 2002, 23, 687–702. [Google Scholar] [CrossRef] [PubMed]
- Kotta-Loizou, I.; Patsouris, E.; Theocharis, S. Pregnane X Receptor Polymorphisms Associated with Human Diseases. Expert Opin. Ther. Targets 2013, 17, 1167–1177. [Google Scholar] [CrossRef]
- Koutsounas, I.; Patsouris, E.; Theocharis, S. Pregnane X Receptor and Human Malignancy. Histol. Histopathol. 2013, 28, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Koutsounas, I.; Theocharis, S.; Patsouris, E.; Giaginis, C. Pregnane X Receptor (PXR) at the Crossroads of Human Metabolism and Disease. Curr. Drug Metab. 2013, 14, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Koutsounas, I.; Giaginis, C.; Alexandrou, P.; Zizi-Serbetzoglou, A.; Patsouris, E.; Kouraklis, G.; Theocharis, S. Pregnane X Receptor Expression in Human Pancreatic Adenocarcinoma. Pancreas 2015, 44, 1134–1140. [Google Scholar] [CrossRef] [PubMed]
- Sandanaraj, E.; Lal, S.; Selvarajan, V.; Ooi, L.L.; Zee, W.W.; Nan, S.W.; Ang, P.C.S.; Lee, E.J.D.; Chowbay, B. PXR Pharmacogenetics: Association of Haplotypes with Hepatic CYP3A4 and ABCB1 Messenger RNA Expression and Doxorubicin Clearance in Asian Breast Cancer Patients. Clin. Cancer Res. 2008, 14, 7116–7126. [Google Scholar] [CrossRef]
- Zhang, J.; Kuehl, P.; Green, E.D.; Touchman, J.W.; Watkins, P.B.; Daly, A.; Hall, S.D.; Maurel, P.; Relling, M.; Brimer, C.; et al. The Human Pregnane X Receptor: Genomic Structure and Identification and Functional Characterization of Natural Allelic Variants. Pharmacogenetics 2001, 11, 555–572. [Google Scholar] [CrossRef]
- Pavek, P.; Dvorak, Z. Xenobiotic-Induced Transcriptional Regulation of Xenobiotic Metabolizing Enzymes of the Cytochrome P450 Superfamily in Human Extrahepatic Tissues. Curr. Drug Metab. 2008, 9, 129–143. [Google Scholar] [CrossRef]
- Nishimura, M.; Naito, S.; Yokoi, T. Tissue-Specific MRNA Expression Profiles of Human Nuclear Receptor Subfamilies. Drug Metab. Pharmacokinet. 2004, 19, 135–149. [Google Scholar] [CrossRef] [PubMed]
- Pondugula, S.R.; Mani, S. Pregnane Xenobiotic Receptor in Cancer Pathogenesis and Therapeutic Response. Cancer Lett. 2013, 328, 1–9. [Google Scholar] [CrossRef]
- Zhang, B.; Xie, W.; Krasowski, M.D. PXR: A xenobiotic receptor of diverse function implicated in pharmacogenetics. Pharmacogenomics 2008, 9, 1695–1709. [Google Scholar] [CrossRef] [PubMed]
- Hogle, B.C.; Guan, X.; Folan, M.M.; Xie, W. PXR as a Mediator of Herb–Drug Interaction. J. Food Drug Anal. 2018, 26, S26–S31. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, W.; Hu, L.; Lv, J.; Wang, H.; Zhou, H.; Fan, L. Role of Pregnane X Receptor in Chemotherapeutic Treatment. Cancer Chemother. Pharmacol. 2014, 74, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Watkins, R.E.; Davis-searles, P.R.; Lambert, M.H.; Redinbo, M.R.; Hill, C. Coactivator Binding Promotes the Specific Interaction Between Ligand and the Pregnane X Receptor. J. Mol. Biol. 2003, 2836, 815–828. [Google Scholar] [CrossRef]
- Revathidevi, S.; Sudesh, R.; Vaishnavi, V.; Kaliyanasundaram, M.; MaryHelen, K.G.; Sukanya, G.; Munirajan, A.K. Screening for the 3’ UTR Polymorphism of the PXR Gene in South Indian Breast Cancer Patients and Its Potential Role in Pharmacogenomics. Asian Pac. J. Cancer Prev. 2016, 17, 3969–3975. [Google Scholar]
- Smutny, T.; Mani, S.; Pavek, P. Post-Translational and Post-Transcriptional Modifications of Pregnane X Receptor (PXR) in Regulation of the Cytochrome P450 Superfamily. Curr. Drug Metab. 2013, 14, 1059–1069. [Google Scholar] [CrossRef]
- Pondugula, S.R.; Pavek, P.; Mani, S. Pregnane X Receptor and Cancer: Context-Specificity Is Key. Nucl. Recept. Res. 2016, 3, 101198. [Google Scholar] [CrossRef]
- Qiao, E.; Ji, M.; Wu, J.; Ma, R.; Zhang, X.; He, Y.; Zha, Q.; Song, X.; Zhu, L.W.; Tang, J. Expression of the PXR Gene in Various Types of Cancer and Drug Resistance (Review). Oncol. Lett. 2013, 5, 1093–1100. [Google Scholar] [CrossRef] [PubMed]
- Mbatchi, L.C.; Brouillet, J.P.; Evrard, A. Genetic Variations of the Xenoreceptors NR1I2 and NR1I3 and Their Effect on Drug Disposition and Response Variability. Pharmacogenomics 2018, 19, 61–77. [Google Scholar] [CrossRef]
- Swart, M.; Dandara, C. Genetic Variation in the 3′-UTR of CYP1A2, CYP2B6, CYP2D6, CYP3A4, NR1I2, and UGT2B7: Potential Effects on Regulation by MicroRNA and Pharmacogenomics Relevance. Front. Genet. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dussault, I.; Lin, M.; Hollister, K.; Fan, M.; Termini, J.; Sherman, M.A.; Forman, B.M. A Structural Model of the Constitutive Androstane Receptor Defines Novel Interactions That Mediate Ligand-Independent Activity. Mol. Cell. Biol. 2002, 22, 5270–5280. [Google Scholar] [CrossRef] [PubMed]
- Chai, X.; Zeng, S.; Xie, W. Nuclear Receptors PXR and CAR: Implications for Drug Metabolism Regulation, Pharmacogenomics and Beyond. Expert Opin. Drug Metab. Toxicol. 2013, 9, 253–266. [Google Scholar] [CrossRef]
- Mukherjee, S.; Mani, S. Orphan Nuclear Receptors as Targets for Drug Development. Pharm. Res. 2010, 27, 1439–1468. [Google Scholar] [CrossRef]
- Bourguet, W.; Germain, P.; Gronemeyer, H. Nuclear Receptor Ligand-Binding Domains: Three-Dimensional Structures, Molecular Interactions and Pharmacological Implications. Trends Pharmacol. Sci. 2000, 21, 381–388. [Google Scholar] [CrossRef]
- Lazar, M.A. Nuclear Receptor Corepressors. Nucl. Recept. Signal. 2003, 1, nrs.01001. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeld, M.G.; Lunyak, V.V.; Glass, C.K. Sensors and Signals: A Coactivator/Corepressor/Epigenetic Code for Integrating Signal-Dependent Programs of Transcriptional Response. Genes Dev. 2006, 20, 1405–1428. [Google Scholar] [CrossRef]
- Orans, J.; Teotico, D.G.; Redinbo, M.R. The Nuclear Xenobiotic Receptor Pregnane X Receptor: Recent Insights and New Challenges. Mol. Endocrinol. 2005, 19, 2891–2900. [Google Scholar] [CrossRef]
- Cui, J.Y.; Gunewardena, S.S.; Rockwell, C.E.; Klaassen, C.D. ChIPing the Cistrome of PXR in Mouse Liver. Nucleic Acids Res. 2010, 38, 7943–7963. [Google Scholar] [CrossRef]
- Wahli, W. A Gut Feeling of the PXR, PPAR and NF-ΚB Connection. J. Intern. Med. 2008, 263, 613–619. [Google Scholar] [CrossRef]
- Staudinger, J.L.; Xu, C.; Biswas, A.; Mani, S. Post-Translational Modification of Pregnane x Receptor. Pharmacol. Res. 2011, 64, 4–10. [Google Scholar] [CrossRef]
- Cui, W.; Sun, M.; Zhang, S.; Shen, X.; Gal-, N.; Williams, T.D.; Staudinger, J.L. A SUMO-Acetyl Switch in PXR Biology. BBA Gene Regul. Mech. 2016, 1859, 1170–1182. [Google Scholar] [CrossRef]
- Vachirayonstien, T.; Yan, B. MicroRNA-30c-1-3p Is a Silencer of the Pregnane X Receptor by Targeting the 3′-Untranslated Region and Alters the Expression of Its Target Gene Cytochrome P450 3A4. Biochim. Biophys. Acta Gene Regul. Mech. 2016, 1859, 1238–1244. [Google Scholar] [CrossRef][Green Version]
- Takagi, S.; Nakajima, M.; Mohri, T.; Yokoi, T. Post-Transcriptional Regulation of Human Pregnane X Receptor by Micro-RNA Affects the Expression Of Cytochrome P450 3A4. J. Biol. Chem. 2008, 283, 9674–9680. [Google Scholar] [CrossRef] [PubMed]
- Lamba, V.; Ghodke, Y.; Guan, W.; Tracy, T.S. MicroRNA-34a Is Associated with Expression of Key Hepatic Transcription Factors and Cytochromes P450. Biochem. Biophys. Res. Commun. 2014, 445, 404–411. [Google Scholar] [CrossRef]
- Li, J.; Zhao, J.; Wang, H.; Li, X.; Liu, A.; Qin, Q.; Li, B. MicroRNA-140-3p Enhances the Sensitivity of Hepatocellular Carcinoma Cells to Sorafenib by Targeting Pregnenolone X Receptor. OncoTargets Ther. 2018, 5885–5894. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Turkistani, A.A.; Chang, W.; Hu, C.; Xu, Z.; Chang, T.K.H. Negative Regulation of Human Pregnane X Receptor by MicroRNA-18a-5p: Evidence for Suppression of MicroRNA-18a-5p Expression by Rifampin and Rilpivirine. Mol. Pharmacol. 2017, 48–56. [Google Scholar] [CrossRef]
- Reuter, T.; Herold-Mende, C.; Dyckhoff, G.; Rigalli, J.P.; Weiss, J. Functional Role of MiR-148a in Oropharyngeal Cancer: Influence on Pregnane X Receptor and P-Glycoprotein Expression. J. Recept. Signal Transduct. 2019, 39, 451–459. [Google Scholar] [CrossRef]
- Ramamoorthy, A.; Li, L.; Gaedigk, A.; Bradford, L.D.A.; Benson, E.A.; Flockhart, D.A.; Skaar, T.C. In Silico and in Vitro Identification of MicroRNAs That Regulate Hepatic Nuclear Factor 4α Expression. Drug Metab. Dispos. 2012, 40, 726–733. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, Y.; Chen, S.; Nie, D.; Chen, Y.; Tang, Y.; Chen, S.; Nie, D. Regulation of Drug Resistance by Human Pregnane X Receptor in Breast Cancer Regulation of Drug Resistance by Human Pregnane X Receptor in Breast Cancer. Cancer Biol. Ther. 2016, 4047, 1265–1272. [Google Scholar] [CrossRef]
- Zhou, H.; Tomlinson, B. Theranostics Meets Traditional Chinese Medicine: Rational Prediction of Drug—Herb Interactions. Expert Rev. Mol. Diagn. 2012, 12, 815–830. [Google Scholar]
- Smutny, T.; Pavek, P. Resveratrol as an Inhibitor of Pregnane X Receptor ( PXR ): Another Lesson in PXR Antagonism. J. Pharmacol. Sci. 2014, 178, 177–178. [Google Scholar] [CrossRef]
- Mani, S.; Dou, W.; Redinbo, M.R. PXR Antagonists and Implication in Drug Metabolism. Drug Metab. Rev. 2013, 45, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Synold, T.W.; Dussault, I.; Forman, B.M. The Orphan Nuclear Receptor SXR Coordinately Regulates Drug Metabolism and Efflux. Nat. Med. 2001, 7, 584–590. [Google Scholar] [CrossRef]
- Chai, S.C.; Wright, W.C.; Chen, T. Strategies for Developing Pregnane X Receptor Antagonists: Implications from Metabolism to Cancer. Med. Res. Rev. 2020, 40, 1061–1083. [Google Scholar] [CrossRef]
- Shen, Y.; Shi, Z.; Fan, J.T.; Yan, B. Dechlorination and Demethylation of Ochratoxin A Enhance Blocking Activity of PXR Activation, Suppress PXR Expression and Reduce Cytotoxicity. Toxicol. Lett. 2020, 332, 171–180. [Google Scholar] [CrossRef]
- Xing, Y.; Yan, J.; Niu, Y. PXR: A Center of Transcriptional Regulation in Cancer. Acta Pharm. Sin. B 2020, 10, 197–206. [Google Scholar] [CrossRef]
- Toporova, L.; Grimaldi, M.; Boulahtouf, A.; Balaguer, P. Assessing the Selectivity of FXR, LXRs, CAR, and RORγ Pharmaceutical Ligands with Reporter Cell Lines. Front. Pharmacol. 2020, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Burk, O.; Kronenberger, T.; Keminer, O.; Lee, S.M.L.; Schiergens, T.S.; Schwab, M.; Windshügel, B. Nelfinavir and Its Active Metabolite M8 Are Partial Agonists and Competitive Antagonists of the Human Pregnane X Receptor. Mol. Pharmacol. 2021, 99, 184–196. [Google Scholar] [CrossRef]
- Lin, W.; Wang, Y.M.; Chai, S.C.; Lv, L.; Zheng, J.; Wu, J.; Zhang, Q.; Wang, Y.D.; Griffin, P.R.; Chen, T. SPA70 Is a Potent Antagonist of Human Pregnane X Receptor. Nat. Commun. 2017, 8, 741. [Google Scholar] [CrossRef]
- Huber, A.D.; Wright, W.C.; Lin, W.; Majumder, K.; Low, J.A.; Wu, J.; Buchman, C.D.; Pintel, D.J.; Chen, T. Mutation of a Single Amino Acid of Pregnane X Receptor Switches an Antagonist to Agonist by Altering AF-2 Helix Positioning. Cell. Mol. Life Sci. 2021, 78, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lin, W.; Wright, W.C.; Chai, S.C.; Wu, J.; Chen, T. Building a Chemical Toolbox for Human Pregnane X Receptor Research: Discovery of Agonists, Inverse Agonists, and Antagonists Among Analogs Based on the Unique Chemical Scaffold of SPA70. J. Med. Chem. 2021, 64, 1733–1761. [Google Scholar] [CrossRef] [PubMed]
- Creamer, B.A.; Sloan, S.N.B.; Dennis, J.F.; Rogers, R.; Spencer, S.; McCuen, A.; Persaud, P.; Staudinger, J.L. Associations between Pregnane X Receptor and Breast Cancer Growth and Progression. Cells 2020, 9, 2295. [Google Scholar] [CrossRef] [PubMed]
- Van de Winkel, A.; Menke, V.; Capello, A.; Moons, L.M.G.; Pot, R.G.J.; van Dekken, H.; Siersema, P.D.; Kusters, J.G.; van der Laan, L.J.W.; Kuipers, E.J. Expression, Localization and Polymorphisms of the Nuclear Receptor PXR in Barrett’s Esophagus and Esophageal Adenocarcinoma. BMC Gastroenterol. 2011, 11, 108. [Google Scholar] [CrossRef]
- Zhang, L.; Qiu, F.; Lu, X.; Li, Y.; Fang, W.; Zhang, L.; Zhou, Y.; Yang, L.; Lu, J. A Functional Polymorphism in the 3’-UTR of PXR Interacts with Smoking to Increase Lung Cancer Risk in Southern and Eastern Chinese Smoker. Int. J. Mol. Sci. 2014, 15, 17457–17468. [Google Scholar] [CrossRef] [PubMed]
- Reuter, T.; Warta, R.; Theile, D.; Meid, A.D.; Rigalli, J.P.; Mogler, C.; Herpel, E.; Grabe, N.; Lahrmann, B.; Plinkert, P.K.; et al. Role of NR1I2 (Pregnane X Receptor) Polymorphisms in Head and Neck Squamous Cell Carcinoma. Naunyn. Schmiedebergs. Arch. Pharmacol. 2015, 388, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Ren, L.; Wang, Y.; Bi, X.; Li, X.; Wen, M.; Zhang, Q.; Yang, Y.; Jia, Y.; Li, Y.; et al. FBI-1 Enhanced the Resistance of Triple-Negative Breast Cancer Cells to Chemotherapeutic Agents via the MiR-30c/PXR Axis. Cell Death Dis. 2020, 11, 851. [Google Scholar] [CrossRef] [PubMed]
- Creusot, N.; Gassiot, M.; Alaterre, E.; Chiavarina, B.; Grimaldi, M.; Boulahtouf, A.; Toporova, L.; Gerbal-Chaloin, S.; Daujat-Chavanieu, M.; Matheux, A.; et al. The Anti-Cancer Drug Dabrafenib Is a Potent Activator of the Human Pregnane X Receptor. Cells 2020, 9, 1641. [Google Scholar] [CrossRef]
- De Mattia, E.; Polesel, J.; Roncato, R.; Labriet, A.; Bignucolo, A.; Dreussi, E.; Romanato, L.; Guardascione, M.; Buonadonna, A.; D’Andrea, M.; et al. Germline Polymorphisms in the Nuclear Receptors PXR and VDR as Novel Prognostic Markers in Metastatic Colorectal Cancer Patients Treated with FOLFIRI. Front. Oncol. 2019, 9, 1–11. [Google Scholar] [CrossRef]
- Mbatchi, L.C.; Robert, J.; Ychou, M.; Boyer, J.C.; Del Rio, M.; Gassiot, M.; Thomas, F.; Tubiana, N.; Evrard, A. Effect of Single Nucleotide Polymorphisms in the Xenobiotic-Sensing Receptors NR1I2 and NR1I3 on the Pharmacokinetics and Toxicity of Irinotecan in Colorectal Cancer Patients. Clin. Pharmacokinet. 2016, 55, 1145–1157. [Google Scholar] [CrossRef]
- Liu, J.; Chen, Z.; Chen, H.; Hou, Y.; Lu, W.; He, J.; Tong, H.; Zhou, Y.; Cai, W. Genetic Polymorphisms Contribute to the Individual Variations of Imatinib Mesylate Plasma Levels and Adverse Reactions in Chinese GIST Patients. Int. J. Mol. Sci. 2017, 18, 603. [Google Scholar] [CrossRef] [PubMed]
- Qian, Y.; Sun, L.N.; Liu, Y.J.; Zhang, Q.; Xu, J.H.; Ma, Z.Q.; Zhang, X.H.; Xu, H.; Wang, Y.Q. Genetic Polymorphisms and Adverse Events on Unbound Imatinib and Its Active Metabolite Concentration in Patients with Gastrointestinal Stromal Tumors. Front. Pharmacol. 2019, 10, 1–9. [Google Scholar] [CrossRef]
- Pamuła-Piłat, J.; Tęcza, K.; Kalinowska-Herok, M.; Grzybowska, E. Genetic 3′UTR Variations and Clinical Factors Significantly Contribute to Survival Prediction and Clinical Response in Breast Cancer Patients. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef]
- Van Der Veldt, A.A.M.; Eechoute, K.; Gelderblom, H.; Gietema, J.; Guchelaar, H.J.; Van Erp, N.P.; Van Den Eertwegh, A.J.M.; Haanen, J.B.; Mathijssen, R.H.J.; Wessels, J.A.M. Genetic Polymorphisms Associated with a Prolonged Progression-Free Survival in Patients with Metastatic Renal Cell Cancer Treated with Sunitinib. Clin. Cancer Res. 2011, 17, 620–629. [Google Scholar] [CrossRef]
- Beuselinck, B.; Karadimou, A.; Lambrechts, D.; Claes, B.; Wolter, P.; Couchy, G.; Berkers, J.; Paridaens, R.; Schöffski, P.; Méjean, A.; et al. Single-Nucleotide Polymorphisms Associated with Outcome in Metastatic Renal Cell Carcinoma Treated with Sunitinib. Br. J. Cancer 2013, 108, 887–900. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, C.F.; Bing, N.X.; Ball, H.A.; Rajagopalan, D.; Sternberg, C.N.; Hutson, T.E.; De Souza, P.; Xue, Z.G.; McCann, L.; King, K.S.; et al. Pazopanib Efficacy in Renal Cell Carcinoma: Evidence for Predictive Genetic Markers in Angiogenesis-Related and Exposure-Related Genes. J. Clin. Oncol. 2011, 29, 2557–2564. [Google Scholar] [CrossRef]
- Xu, C.; Ball, H.A.; Bing, N.; Sternberg, C.N.; Xue, Z.; McCann, L.; King, K.; Spraggs, C.F.; Mooser, V.E.; Pandite, L.N. Association of Genetic Markers in Angiogenesis- or Exposure-Related Genes with Overall Survival in Pazopanib (P) Treated Patients (Pts) with Advanced Renal Cell Carcinoma. J. Clin. Oncol. 2011, 29, 303. [Google Scholar] [CrossRef]
- Mbatchi, L.C.; Gassiot, M.; Pourquier, P.; Goberna, A.; Mahammedi, H.; Mourey, L.; Joly, F.; Lumbroso, S.; Evrard, A.; Houede, N. Association of NR1I2, CYP3A5 and ABCB1 Genetic Polymorphisms with Variability of Temsirolimus Pharmacokinetics and Toxicity in Patients with Metastatic Bladder Cancer. Cancer Chemother. Pharmacol. 2017, 80, 653–659. [Google Scholar] [CrossRef]
- Chew, S.C.; Lim, J.; Singh, O.; Chen, X.; Tan, E.H.; Lee, E.J.; Chowbay, B. Pharmacogenetic Effects of Regulatory Nuclear Receptors (PXR, CAR, RXRα and HNF4α) on Docetaxel Disposition in Chinese Nasopharyngeal Cancer Patients. Eur. J. Clin. Pharmacol. 2014, 70, 155–166. [Google Scholar] [CrossRef]
- Zhou, Y.; Yang, P.; Liu, Y.; Wang, L. Effect of Polymorphisms of NF-ΚB and PXR on Platinum-Based Chemotherapy for Non-Small Cell Lung Cancer. Zhong Nan Da Xue Xue Bao. Yi Xue Ban 2016, 41, 233–237. [Google Scholar] [CrossRef]
- Abumiya, M.; Mita, A.; Takahashi, S.; Yoshioka, T.; Kameoka, Y.; Takahashi, N.; Miura, M. Effects of Polymorphisms in NR1I2, CYP3A4, and ABC Transporters on the Steady-State Plasma Trough Concentrations of Bosutinib in Japanese Patient with Chronic Myeloid Leukemia. Med. Oncol. 2018, 35, 1–7. [Google Scholar] [CrossRef]
- Ren, W.; Zhou, C.; Liu, Y.; Su, K.; Jia, L.; Chen, L.; Li, M.; Ma, J.; Zhou, W.; Zhang, S.; et al. Genetic Associations of Docetaxel-Based Chemotherapy-Induced Myelosuppression in Chinese Han Population. J. Clin. Pharm. Ther. 2020, 45, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Mbatchi, L.C.; Schmitt, A.; Thomas, F.; Cazaubon, Y.; Robert, J.; Lumbroso, S.; Brouillet, J.P.; Pourquier, P.; Chatelut, E.; Boyer, J.C.; et al. Polymorphisms in SLCO1B3 and NR1I2 as Genetic Determinants of Hematotoxicity of Carboplatin and Paclitaxel Combination. Pharmacogenomics 2015, 16, 1439–1450. [Google Scholar] [CrossRef]
- Hegyi, M.; Arany, A.; Semsei, A.F.; Csordas, K.; Eipel, O.; Gezsi, A.; Kutszegi, N.; Csoka, M.; Muller, J.; Erdelyi, D.J.; et al. Pharmacogenetic Analysis of High-Dose Methotrexate Treatment in Children with Osteosarcoma. Oncotarget 2017, 8, 9388–9398. [Google Scholar] [CrossRef]
- Yang, M.; Pan, H.; Chen, H.; Liu, W.; Lu, L.; He, X.; Yi, H.; Tang, S. Association between NR1I2 Polymorphisms and Susceptibility to Anti-Tuberculosis Drug-Induced Hepatotoxicity in an Eastern Chinese Han Population: A Case-Control Study. Infect. Genet. Evol. 2020, 83, 104349. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xiang, X.; Huang, W.W.; Sandford, A.J.; Wu, S.Q.; Zhang, M.M.; Wang, M.G.; Chen, G.; He, J.Q. Association of PXR and CAR Polymorphisms and Antituberculosis Drug-Induced Hepatotoxicity. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Zhu, H.; Luo, X.; Ge, W. PXR Haplotype Clusters Will Affect the Pharmacokinetics of Ciclosporin in Chinese Renal Transplant Recipients. J. Pharm. Pharmacol. 2020, 72, 271–278. [Google Scholar] [CrossRef]
- Chen, Y.B.; Zhou, Z.Y.; Li, G.M.; Xiao, C.X.; Yu, W.B.; Zhong, S.l.; Cai, Y.F.; Jin, J.; Huang, M. Influences of an NR1I2 Polymorphism on Heterogeneous Antiplatelet Reactivity Responses to Clopidogrel and Clinical Outcomes in Acute Ischemic Stroke Patients. Acta Pharmacol. Sin. 2019, 40, 762–768. [Google Scholar] [CrossRef] [PubMed]
- D’Avolio, A.; Carcieri, C.; Cusato, J.; Simiele, M.; Calcagno, A.; Allegra, S.; Sciandra, M.; Trentini, L.; Di Perri, G.; Bonora, S. Intracellular Accumulation of Atazanavir/Ritonavir According to Plasma Concentrations and OATP1B1, ABCB1 and PXR Genetic Polymorphisms. J. Antimicrob. Chemother. 2014, 69, 3061–3066. [Google Scholar] [CrossRef] [PubMed]
- Schipani, A.; Siccardi, M.; D’Avolio, A.; Baietto, L.; Simiele, M.; Bonora, S.; Novoa, S.R.; Cuenca, L.; Soriano, V.; Chierakul, N.; et al. Population Pharmacokinetic Modeling of the Association between 63396C→T Pregnane X Receptor Polymorphism and Unboosted Atazanavir Clearance. Antimicrob. Agents Chemother. 2010, 54, 5242–5250. [Google Scholar] [CrossRef]
- Noetzli, M.; Guidi, M.; Ebbing, K.; Eyer, S.; Wilhelm, L.; Michon, A.; Thomazic, V.; Alnawaqil, A.M.; Maurer, S.; Zumbach, S.; et al. Population Pharmacokinetic Study of Memantine: Effects of Clinical and Genetic Factors. Clin. Pharmacokinet. 2013, 52, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Zeng, G.; Wang, L.; Shi, L.; Li, H.; Zhu, M.; Luo, J.; Zhang, Z. Variability of Voriconazole Concentrations in Patients with Hematopoietic Stem Cell Transplantation and Hematological Malignancies: Influence of Loading Dose, Procalcitonin, and Pregnane X Receptor Polymorphisms. Eur. J. Clin. Pharmacol. 2020, 76, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.; Lockhart, A.C.; Kim, R.B.; Rothenberg, M.L. Cancer Pharmacogenomics: Powerful Tools in Cancer Chemotherapy and Drug Development. Oncologist 2005, 10, 104–111. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP (rs) | Localization | Pathology | Therapy | Patients | Association | Ref. |
|---|---|---|---|---|---|---|
| rs1054190 (C > T) | 3′-UTR | mCRC | FOLFIRI | 247 Italian (discovery cohort) 90 Canadian (Replication cohort) | Worse OS and PFS (T allele) | [59] |
| Breast Cancer | Doxorubicin | 96 South Indian | Change in doxorubicin pharmacokinetics via miRNA altering | [16] | ||
| rs10934498 (G > A, G > C, G > T) | Intron 1 | mCRC | FOLFIRI/ FOLFIRINOX | 109 French | Decreased AUC of SN-38 Decreased biliary index Decreased risk of grade 3–4 hematotoxicity (A allele) | [60] |
| rs3814055 (C > T) | 5′-UTR | mCRC | FOLFIRI/ FOLFIRINOX | 109 French | Increased risk of grade 3–4 hematotoxicity (T allele) | [60] |
| GISTs | Imatinib | 68 Asian 62 patients | Decreased imatinib plasma concentrations and lower edema incidence (T allele) | [61,62] | ||
| RCC | Sunitinib | 136 patients | Shorter PFS, OS (T allele) | [64,65] | ||
| RCC | Pazopanib | 397 patients | Reduced RR, PFS, OS (T allele) | [66,67] | ||
| Bladder cancer | Temsirolimus | 54 patients | Decreased frequency of adverse events (T allele) High frequency of severe toxicity (CC genotype) Extended temsirolimus T1/2 (TT genotype) | [68] | ||
| Osteosarcoma | MTX | 59 patients | Increased first T1/2 of MTX (T allele) | [74] | ||
| rs1523127 (C > A) | 5′-UTR | mCRC | FOLFIRI/ FOLFIRINOX | 109 French | Increased risk of grade 3–4 hematotoxicity (C allele) | [60] |
| rs2472677 (C > G, C > T) | Intron 2 | mCRC | FOLFIRI/ FOLFIRINOX | 109 French | Increased risk of all type of grade 3–4 toxicity (G allele) | [60] |
| rs3732359 (G > A) | 3′-UTR | Breast cancer | FAC | 305 Caucasian | Increased OS (G allele) | [63] |
| Nasopharyngeal cancer | Docetaxel | 50 Asian | Decrease in nadir hemoglobin from baseline (G allele) | [69] | ||
| Solid tumors | Docetaxel | 110 Asian | Docetaxel-induced myelosuppresion grade ≥3 (G allele) | [72] | ||
| rs2276707 (C > T, C > G) | Intron 7 | Breast cancer | Doxorubicin/ Cyclophosphamide | 62 Asian | Haplotype cluster (rs2276707 and rs3814058) associated with reduced doxorubicin clearance | [7] |
| RCC | Sunitinib | 136 patients | Shorter PFS, OS (T allele) | [64,65] | ||
| CML | Bosutinib | 30 Asian | Increased bosutinib clearance (TT genotype) | [71] | ||
| rs3814058 (T > C) | 3′-UTR | Breast cancer | Doxorubicin/ Cyclophosphamide | 62 Asian | Haplotype cluster (rs2276707 and rs3814058) associated with reduced doxorubicin clearance | [7] |
| Nasopharyngeal cancer | Docetaxel | 50 Asian | Decrease in nadir hemoglobin from baseline (C allele) | [69] | ||
| NSCLC | Platinum-based | 262 Asian | High risk of hematological toxicity (C allele) | [70] | ||
| Osteosarcoma | MTX | 59 patients | Reduced risk of hepatotoxicity/bone marrow toxicity (C allele) | [74] | ||
| rs3732360 (C > T, C > G) | 3′-UTR | Breast cancer | Doxorubicin | 96 South Indian | Change in doxorubicin pharmacokinetics via miRNA altering | [16] |
| Nasopharyngeal cancer | Docetaxel | 50 Asian | Decrease in nadir hemoglobin from baseline (T allele) | [69] | ||
| rs1054191 (G > A) | 3′-UTR | Breast cancer | Doxorubicin | 96 South Indian | Change in doxorubicin pharmacokinetics via miRNA altering | [16] |
| rs6785049 (G > A, G > T) | Intron 6 | Bladder cancer | Temsirolimus | 54 patients | Decreased frequency of adverse events (G allele) Increased exposure to active entities (GG genotype) High frequency of severe toxicity (AA genotype) | [68] |
| CML | Bosutinib | 30 Asian | Increased bosutinib clearance (GG genotype) | [71] | ||
| Osteosarcoma | MTX | 59 patients | Increased 48-h MTX concentration (G allele) Reduced risk of hepatotoxicity/bone marrow toxicity (G allele) | [74] | ||
| rs7643038 (A > G) | 5′-UTR | Osteosarcoma | MTX | 59 patients | Increased first T1/2 of MTX (G allele) | [74] |
| rs3732361 (A > G, A > C) | 3′-UTR | Osteosarcoma | MTX | 59 patients | Increased 48-h MTX concentration (G allele) Reduced risk of hepatotoxicity/bone marrow toxicity (G allele) | [74] |
| rs1523130 (T > A, T > C, T > G) | 5′-UTR | mCRC | Irinotecan | 109 Caucasian | Reduced APC and NPC metabolism (T allele) | [60] |
| Solid tumors | Carboplatin/Paclitaxel | 201 patients | Reduced sensitivity to thrombocytopenia (AA genotype) ATG haplotype (rs1523130, rs3814055, rs1523127) linked to reduced sensitivity to thrombocytopenia | [73] | ||
| rs152312 (C > T) | 5′-UTR | mCRC | Irinotecan | 109 Caucasian | Reduced NPC metabolism (C allele) | [60] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Skandalaki, A.; Sarantis, P.; Theocharis, S. Pregnane X Receptor (PXR) Polymorphisms and Cancer Treatment. Biomolecules 2021, 11, 1142. https://doi.org/10.3390/biom11081142
Skandalaki A, Sarantis P, Theocharis S. Pregnane X Receptor (PXR) Polymorphisms and Cancer Treatment. Biomolecules. 2021; 11(8):1142. https://doi.org/10.3390/biom11081142
Chicago/Turabian StyleSkandalaki, Aikaterini, Panagiotis Sarantis, and Stamatios Theocharis. 2021. "Pregnane X Receptor (PXR) Polymorphisms and Cancer Treatment" Biomolecules 11, no. 8: 1142. https://doi.org/10.3390/biom11081142
APA StyleSkandalaki, A., Sarantis, P., & Theocharis, S. (2021). Pregnane X Receptor (PXR) Polymorphisms and Cancer Treatment. Biomolecules, 11(8), 1142. https://doi.org/10.3390/biom11081142