
Plasma Membrane Ca2+–ATPase in Rat and Human Odontoblasts Mediates Dentin Mineralization


 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Odontoblast Cell Culture
2.3. Acute Isolation of Rat Odontoblasts from Dental Pulp Slices
2.4. RT-PCR
2.5. Fluorescence Measurement of [Ca2+]i
2.6. Mineralization Assay
2.7. Solutions and Reagents
2.8. Immunostaining
2.9. Statistics and Offline Analysis
3. Results
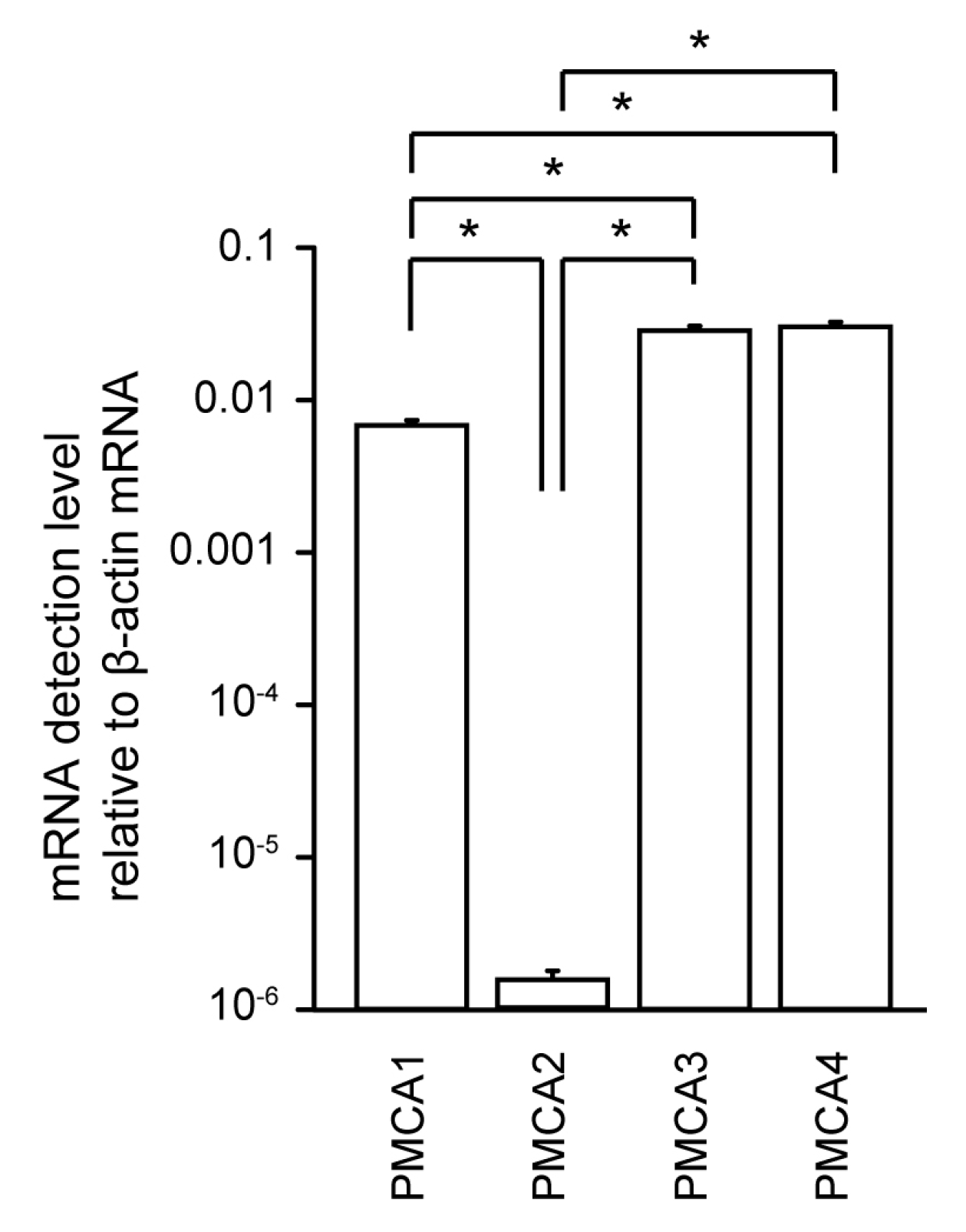
3.1. Measurement of PMCA1–4 mRNA Levels in Human Odontoblasts
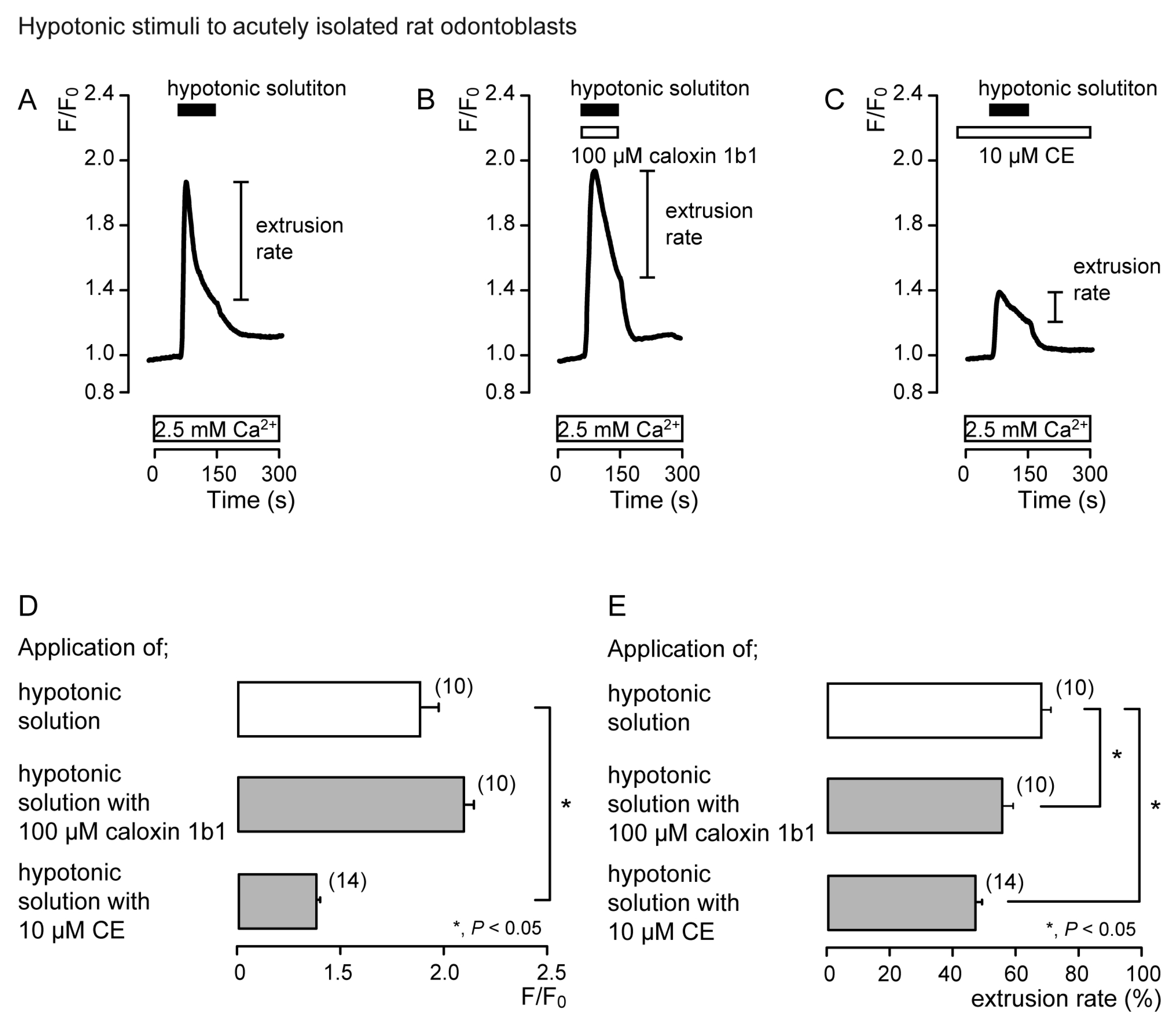
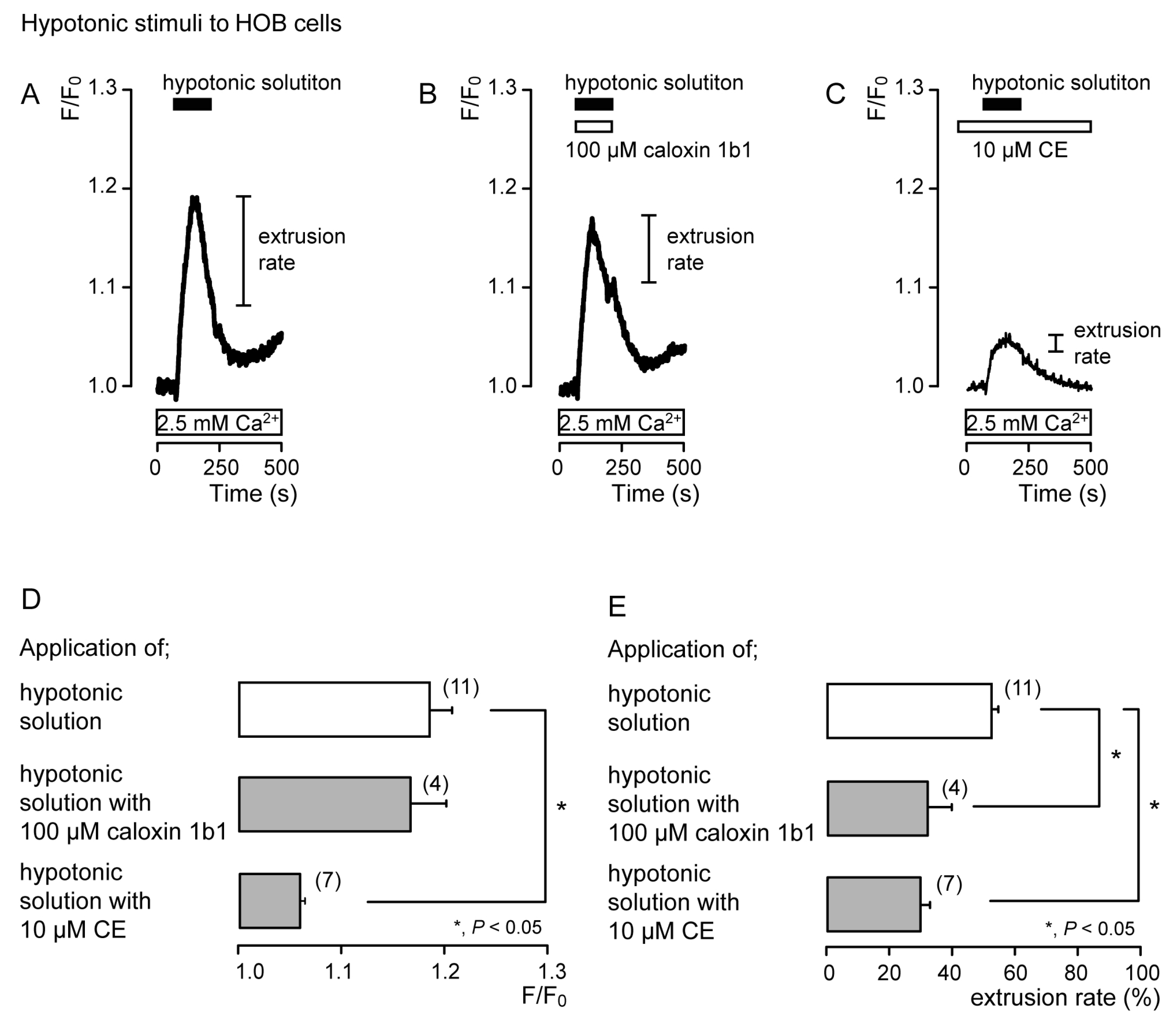
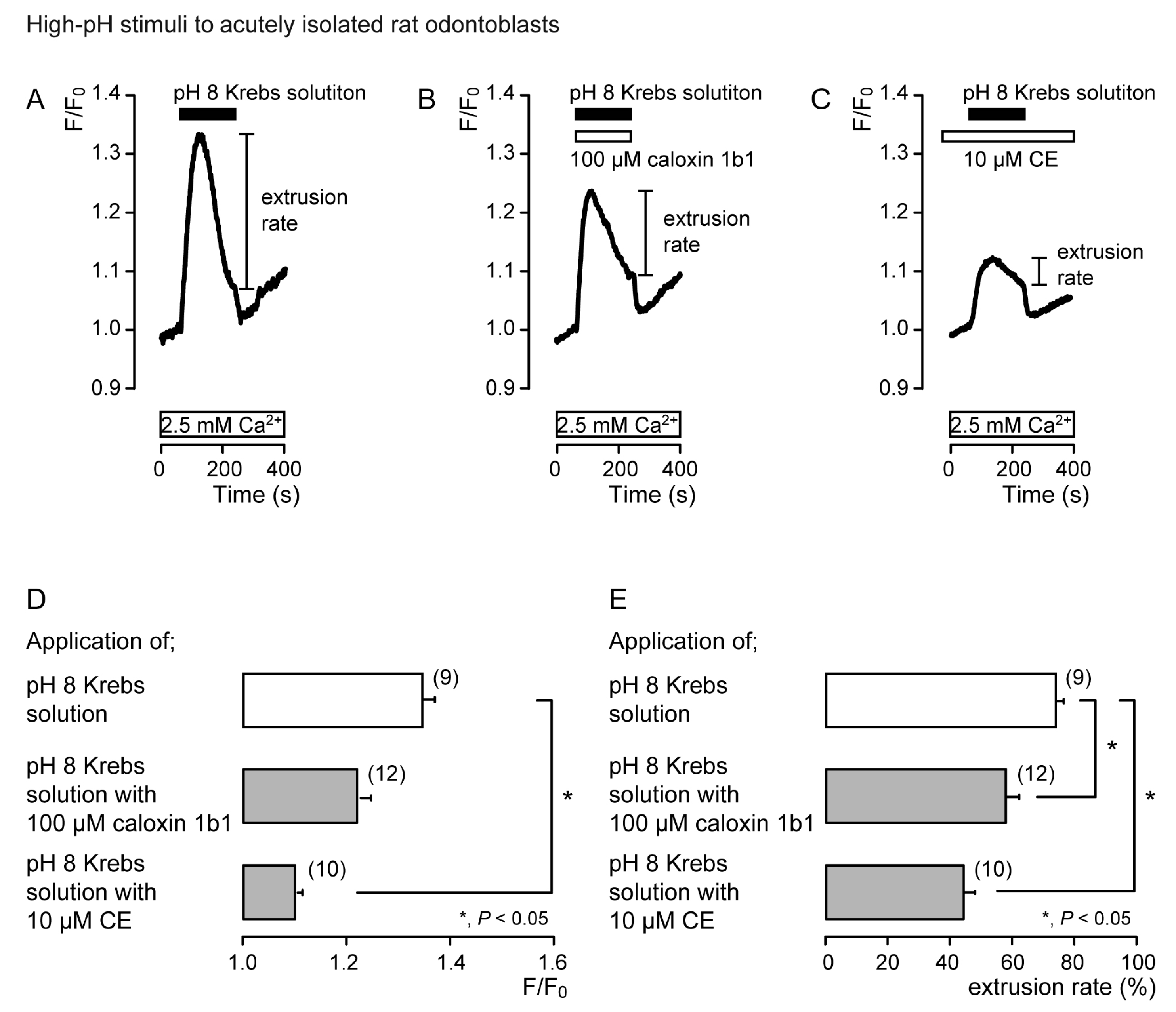
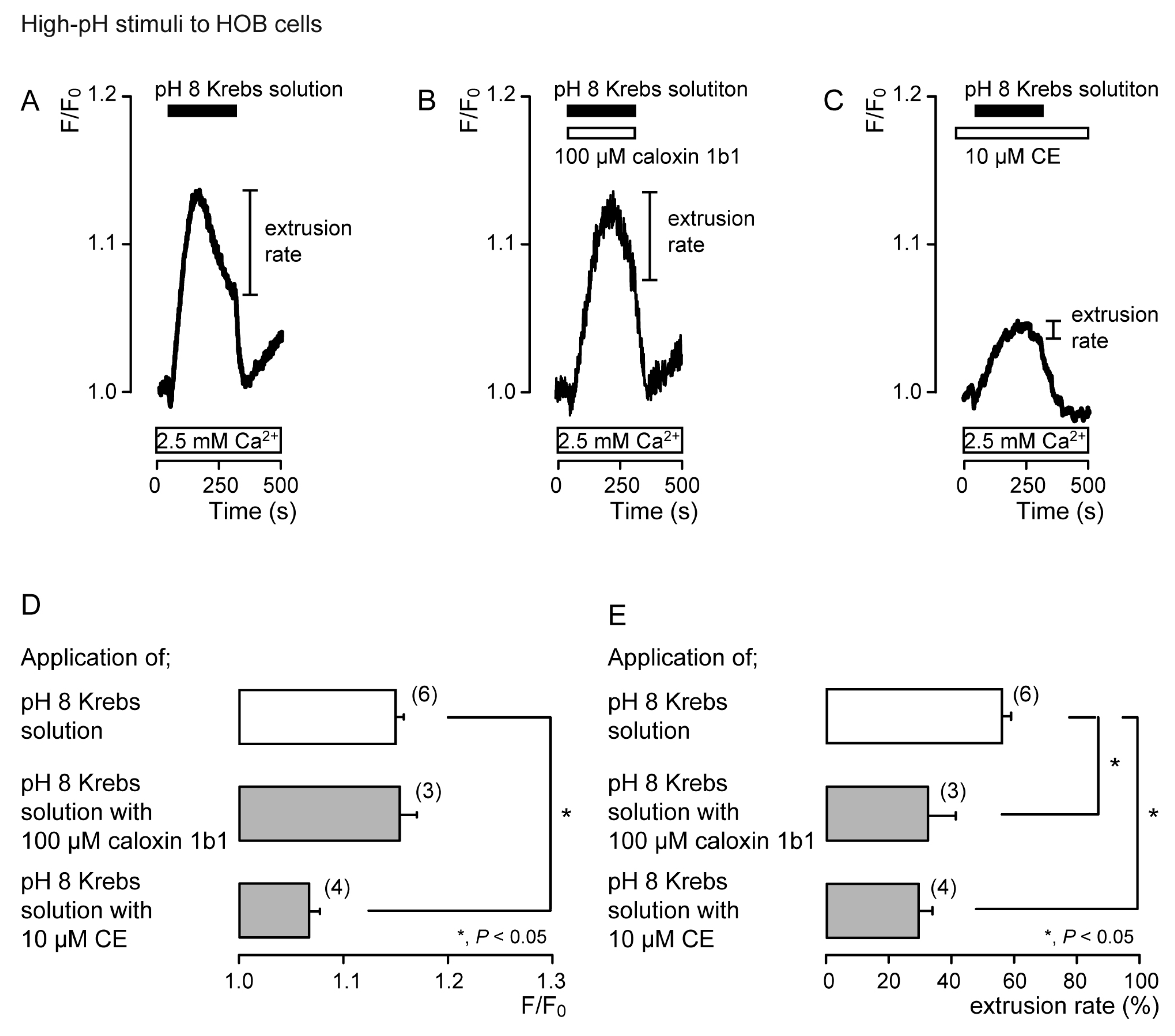
3.2. Extrusion of Ca2+ by PMCA Following Hypotonic or High-pH Stimulation in Acutely Isolated Rat Odontoblasts and HOB Cells
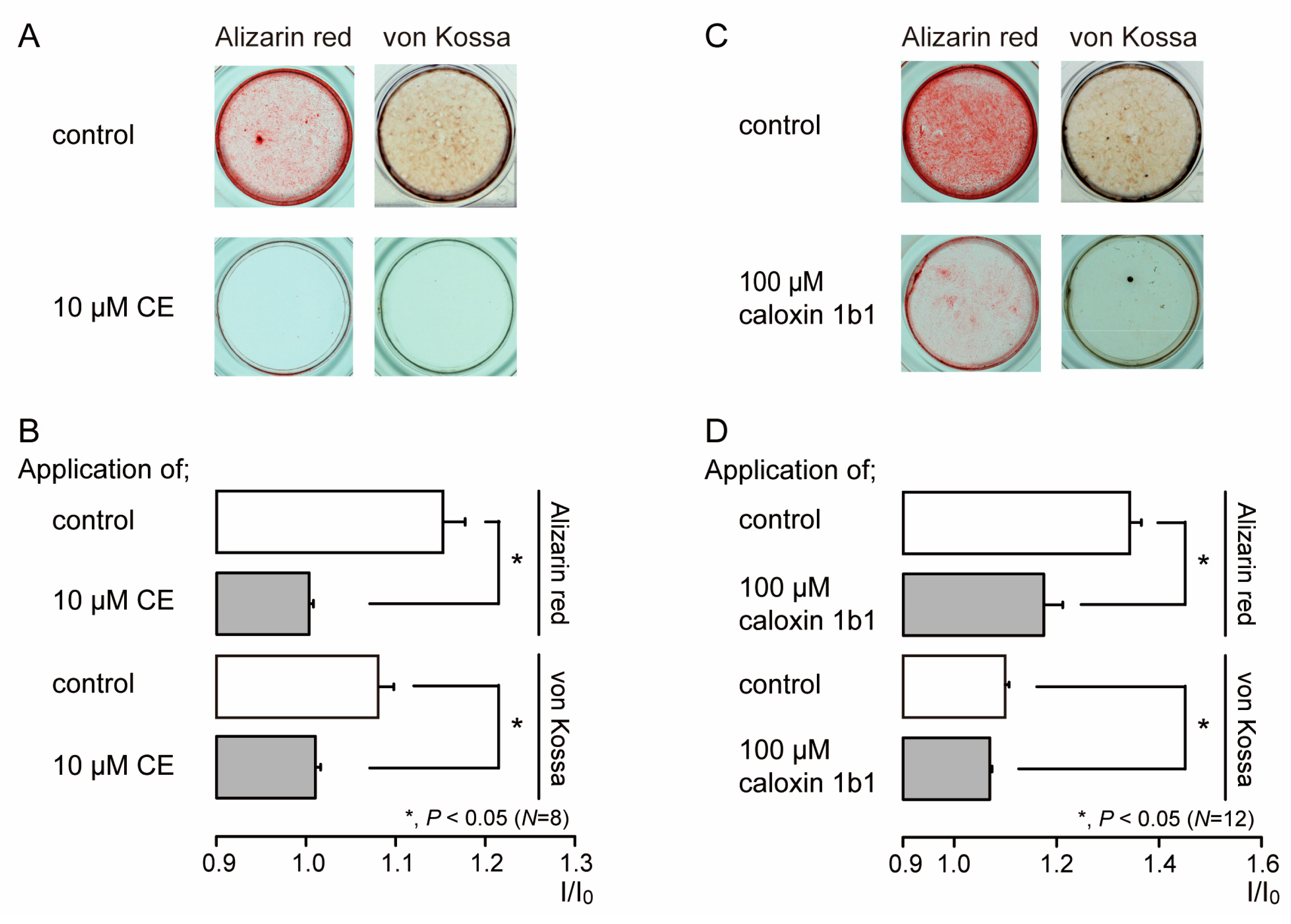
3.3. PMCA Mediates Mineralization by Odontoblasts
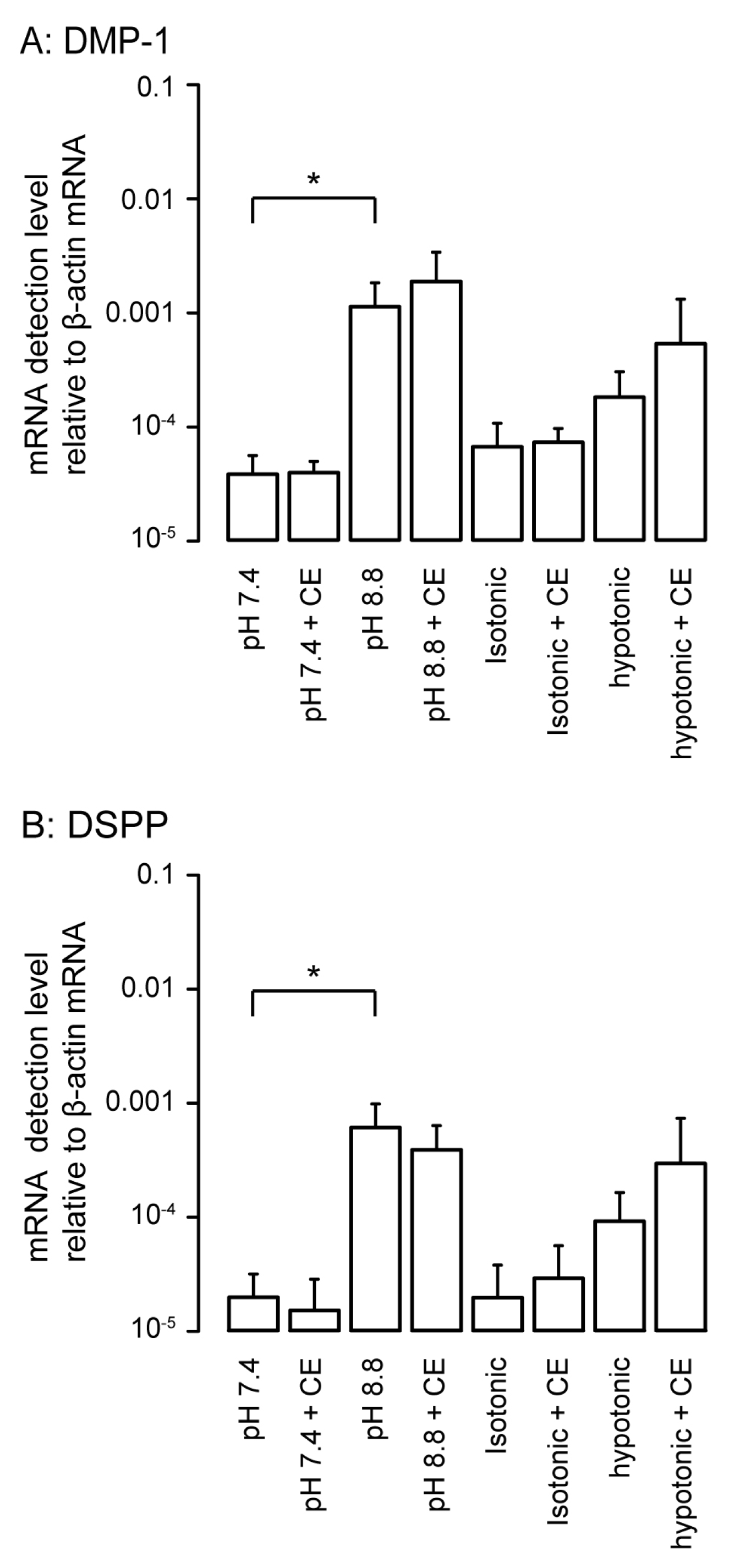
3.4. Effects of PMCA Inhibitor on Detected mRNA Levels of Non-Collagenous Extracellular Matrix Proteins as Odontoblast Markers
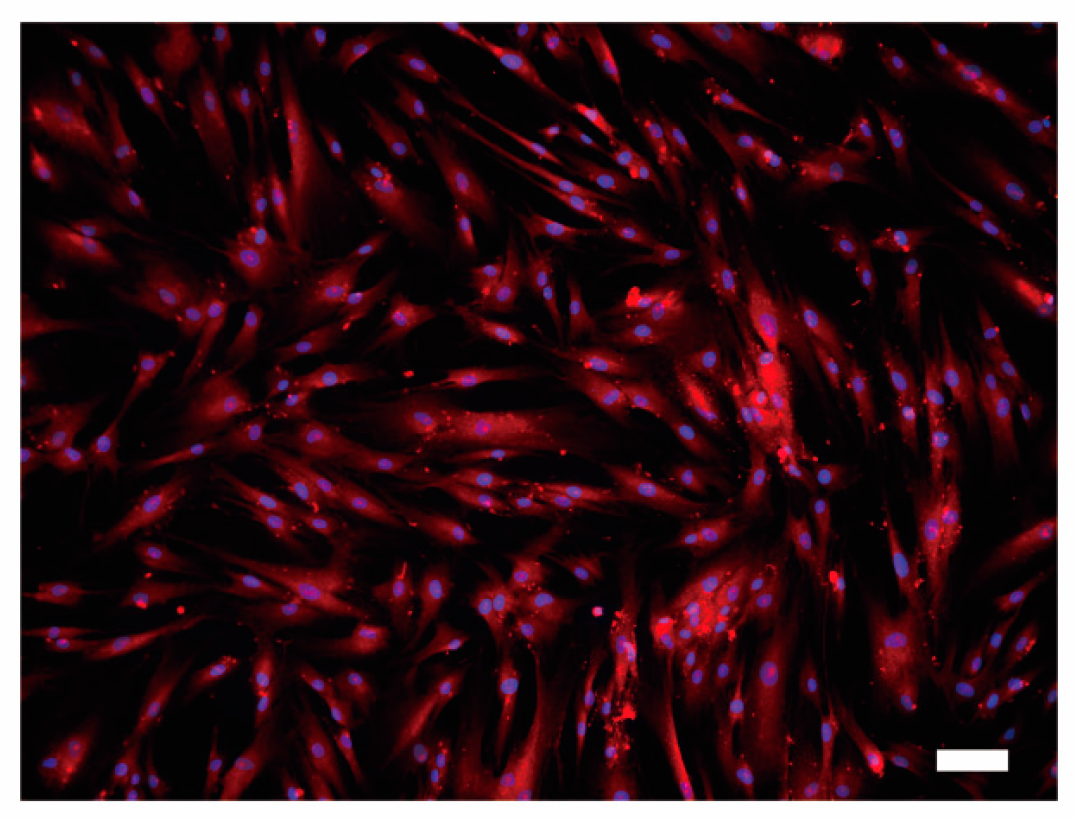
3.5. Immunofluoresence Analysis of PMCA1 in HOB Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shibukawa, Y.; Sato, M.; Kimura, M.; Sobhan, U.; Shimada, M.; Nishiyama, A.; Kawaguchi, A.; Soya, M.; Kuroda, H.; Katakura, A.; et al. Odontoblasts as Sensory Receptors: Transient Receptor Potential Channels, Pannexin-1, and Ionotropic ATP Receptors Mediate Intercellular Odontoblast-Neuron Signal Transduction. Pflüg. Arch. Eur. J. Physiol. 2015, 467, 843–863. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, M.; Sobhan, U.; Muramatsu, T.; Sato, M.; Ichikawa, H.; Sahara, Y.; Tazaki, M.; Shibukawa, Y. TRPV1-Mediated Calcium Signal Couples with Cannabinoid Receptors and Sodium–Calcium Exchangers in Rat Odontoblasts. Cell Calcium 2012, 52, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Sobhan, U.; Tsumura, M.; Kuroda, H.; Soya, M.; Masamura, A.; Nishiyama, A.; Katakura, A.; Ichinohe, T.; Tazaki, M.; et al. Hypotonic-Induced Stretching of Plasma Membrane Activates Transient Receptor Potential Vanilloid Channels and Sodium-Calcium Exchangers in Mouse Odontoblasts. J. Endod. 2013, 39, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Sato, M.; Furuya, T.; Kimura, M.; Kojima, Y.; Tazaki, M.; Sato, T.; Shibukawa, Y. Intercellular Odontoblast Communication via ATP Mediated by Pannexin-1 Channel and Phospholipase C-Coupled Receptor Activation. Front. Physiol. 2015, 6, 326. [Google Scholar] [CrossRef]
- Sato, M.; Ogura, K.; Kimura, M.; Nishi, K.; Ando, M.; Tazaki, M.; Shibukawa, Y. Activation of Mechanosensitive Transient Receptor Potential/Piezo Channels in Odontoblasts Generates Action Potentials in Cocultured Isolectin B4-Negative Medium-Sized Trigeminal Ganglion Neurons. J. Endod. 2018, 44, 984–991. [Google Scholar] [CrossRef]
- Kimura, M.; Sase, T.; Higashikawa, A.; Sato, M.; Sato, T.; Tazaki, M.; Shibukawa, Y. High pH-Sensitive TRPA1 Activation in Odontoblasts Regulates Mineralization. J. Dent. Res. 2016, 95, 1057–1064. [Google Scholar] [CrossRef]
- Kimura, M.; Nishi, K.; Higashikawa, A.; Ohyama, S.; Sakurai, K.; Tazaki, M.; Shibukawa, Y. High pH-Sensitive Store-Operated Ca2+ Entry Mediated by Ca2+ Release-Activated Ca2+ Channels in Rat Odontoblasts. Front. Physiol. 2018, 9, 443. [Google Scholar] [CrossRef]
- Nishiyama, A.; Sato, M.; Kimura, M.; Katakura, A.; Tazaki, M.; Shibukawa, Y. Intercellular Signal Communication among Odontoblasts and Trigeminal Ganglion Neurons via Glutamate. Cell Calcium 2016, 60, 341–355. [Google Scholar] [CrossRef]
- Tsumura, M.; Okumura, R.; Tatsuyama, S.; Ichikawa, H.; Muramatsu, T.; Matsuda, T.; Baba, A.; Suzuki, K.; Kajiya, H.; Sahara, Y.; et al. Ca2+ Extrusion via Na+-Ca2+ Exchangers in Rat Odontoblasts. J. Endod. 2010, 36, 668–674. [Google Scholar] [CrossRef]
- Pande, J.; Mallhi, K.K.; Grover, A.K. Role of Third Extracellular Domain of Plasma Membrane Ca2+–Mg2+-ATPase Based on the Novel Inhibitor Caloxin 3A1. Cell Calcium 2005, 37, 245–250. [Google Scholar] [CrossRef]
- Chen, Y.; Cao, J.; Zhong, J.; Chen, X.; Cheng, M.; Yang, J.; Gao, Y. Plasma Membrane Ca2+-ATPase Regulates Ca2+ Signaling and the Proliferation of Airway Smooth Muscle Cells. Eur. J. Pharmacol. 2014, 740, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Brini, M.; Carafoli, E. The Plasma Membrane Ca2+ ATPase and the Plasma Membrane Sodium Calcium Exchanger Cooperate in the Regulation of Cell Calcium. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–15. [Google Scholar] [CrossRef]
- Bruce, J.I.E. Metabolic Regulation of the PMCA: Role in Cell Death and Survival. Cell Calcium 2018, 69, 28–36. [Google Scholar] [CrossRef]
- Giacomello, M.; De Mario, A.; Scarlatti, C.; Primerano, S.; Carafoli, E. Plasma Membrane Calcium ATPases and Related Disorders. Int. J. Biochem. Cell Biol. 2013, 45, 753–762. [Google Scholar] [CrossRef]
- Szewczyk, M.M.; Pande, J.; Grover, A.K. Caloxins: A Novel Class of Selective Plasma Membrane Ca2+ Pump Inhibitors Obtained Using Biotechnology. Pflüg. Arch. 2008, 456, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Sato, I.; Shimada, K.; Ezure, H.; Sato, T.; Lance, V.A. Distribution of Calcium-ATPase in Developing Teeth of Embryonic American Alligators (Alligator Mississippiensis). J. Morphol. 1993, 218, 249–256. [Google Scholar] [CrossRef]
- Borke, J.L.; Zaki, A.E.; Eisenmann, D.R.; Ashrafi, S.H.; Ashrafi, S.S.; Penniston, J.T. Expression of Plasma Membrane Ca++ Pump Epitopes Parallels the Progression of Enamel and Dentin Mineralization in Rat Incisor. J. Histochem. Cytochem. 1993, 41, 175–181. [Google Scholar] [CrossRef]
- Lundgren, T.; Linde, A. Na+/Ca2+ Antiports in Membranes of Rat Incisor Odontoblasts. J. Oral Pathol. 1988, 17, 560–563. [Google Scholar] [CrossRef]
- Linde, A.; Lundgren, T. From Serum to the Mineral Phase. The Role of the Odontoblast in Calcium Transport and Mineral Formation. Int. J. Dev. Biol. 1995, 39, 213–222. [Google Scholar]
- Kitagawa, M.; Ueda, H.; Iizuka, S.; Sakamoto, K.; Oka, H.; Kudo, Y.; Ogawa, I.; Miyauchi, M.; Tahara, H.; Takata, T. Immortalization and Characterization of Human Dental Pulp Cells with Odontoblastic Differentiation. Arch. Oral Biol. 2007, 52, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, H.; Kim, H.J.; Shuprisha, A.; Shikano, T.; Tsumura, M.; Shibukawa, Y.; Tazaki, M. Voltage-Dependent Sodium Channels and Calcium-Activated Potassium Channels in Human Odontoblasts In Vitro. J. Endod. 2012, 38, 1355–1362. [Google Scholar] [CrossRef]
- Shibukawa, Y.; Suzuki, T. Ca2+ Signaling Mediated by IP3-Dependent Ca2+ Releasing and Store-Operated Ca2+ Channels in Rat Odontoblasts. J. Bone Miner. Res. 2003, 18, 30–38. [Google Scholar] [CrossRef]
- Son, A.R.; Yang, Y.M.; Hong, J.H.; Lee, S.I.; Shibukawa, Y.; Shin, D.M. Odontoblast TRP Channels and Thermo/Mechanical Transmission. J. Dent. Res. 2009, 88, 1014–1019. [Google Scholar] [CrossRef] [PubMed]
- Tokuda, M.; Tatsuyama, S.; Fujisawa, M.; Morimoto-Yamashita, Y.; Kawakami, Y.; Shibukawa, Y.; Torii, M. Dentin and Pulp Sense Cold Stimulus. Med. Hypotheses 2015, 84, 442–444. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative C(T) Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Tsien, R.Y.; Rink, T.J.; Poenie, M. Measurement of Cytosolic Free Ca2+ in Individual Small Cells Using Fluorescence Microscopy with Dual Excitation Wavelengths. Cell Calcium 1985, 6, 145–157. [Google Scholar] [CrossRef]
- Kojima, Y.; Kimura, M.; Higashikawa, A.; Kono, K.; Ando, M.; Tazaki, M.; Shibukawa, Y. Potassium Currents Activated by Depolarization in Odontoblasts. Front. Physiol. 2017, 8, 1078. [Google Scholar] [CrossRef] [PubMed]
- Tsumura, M.; Sobhan, U.; Sato, M.; Shimada, M.; Nishiyama, A.; Kawaguchi, A.; Soya, M.; Kuroda, H.; Tazaki, M.; Shibukawa, Y. Functional Expression of TRPM8 and TRPA1 Channels in Rat Odontoblasts. PLoS ONE 2013, 8, e82233. [Google Scholar] [CrossRef] [PubMed]
- Pande, J.; Mallhi, K.K.; Sawh, A.; Szewczyk, M.M.; Simpson, F.; Grover, A.K. Aortic Smooth Muscle and Endothelial Plasma Membrane Ca2+ Pump Isoforms Are Inhibited Differently by the Extracellular Inhibitor Caloxin 1b1. Am. J. Physiol. Cell Physiol. 2006, 290, C1341–C1349. [Google Scholar] [CrossRef]
- Groten, C.J.; Rebane, J.T.; Hodgson, H.M.; Chauhan, A.K.; Blohm, G.; Magoski, N.S. Ca2+ Removal by the Plasma Membrane Ca2+-ATPase Influences the Contribution of Mitochondria to Activity-Dependent Ca2+ Dynamics in Aplysia Neuroendocrine Cells. J. Neurophysiol. 2016, 115, 2615–2634. [Google Scholar] [CrossRef]
- Pande, J.; Szewczyk, M.M.; Kuszczak, I.; Grover, S.; Escher, E.; Grover, A.K. Functional Effects of Caloxin 1c2, a Novel Engineered Selective Inhibitor of Plasma Membrane Ca2+-Pump Isoform 4, on Coronary Artery. J. Cell. Mol. Med. 2008, 12, 1049–1060. [Google Scholar] [CrossRef]
- Mohamed, T.M.A.; Zakeri, S.A.; Baudoin, F.; Wolf, M.; Oceandy, D.; Cartwright, E.J.; Gul, S.; Neyses, L. Optimisation and Validation of a High Throughput Screening Compatible Assay to Identify Inhibitors of the Plasma Membrane Calcium ATPase Pump—A Novel Therapeutic Target for Contraception and Malaria. J. Pharm. Pharm. Sci. 2013, 16, 217–230. [Google Scholar] [CrossRef]
- Ryan, Z.C.; Craig, T.A.; Filoteo, A.G.; Westendorf, J.J.; Cartwright, E.J.; Neyses, L.; Strehler, E.E.; Kumar, R. Deletion of the Intestinal Plasma Membrane Calcium Pump, Isoform 1, Atp2b1, in Mice Is Associated with Decreased Bone Mineral Density and Impaired Responsiveness to 1, 25-Dihydroxyvitamin D3. Biochem. Biophys. Res. Commun. 2015, 467, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Abramowitz, J.; Suki, W.N. Ca-ATPase and Bone Cell Mineralization. Miner. Electrolyte Metab. 1996, 22, 336–344. [Google Scholar]
- Go, W.; Korzh, V. Plasma Membrane Ca2+ ATPase Atp2b1a Regulates Bone Mineralization in Zebrafish. Bone 2013, 54, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Domi, T.; Di Leva, F.; Fedrizzi, L.; Rimessi, A.; Brini, M. Functional Specificity of PMCA Isoforms? Ann. N. Y. Acad. Sci. 2007, 1099, 237–246. [Google Scholar] [CrossRef]
- Lundgren, T. Calcium Transport in Dentinogenesis. An Experimental Study in the Rat Incisor Odontoblast. Swed. Dent. J. Suppl. 1992, 82, 1–91. [Google Scholar] [PubMed]
- Shibukawa, Y.; Suzuki, T. Measurements of Cytosolic Free Ca2+ Concentrations in Odontoblasts. Bull. Tokyo Dent. Coll. 1997, 38, 177–185. [Google Scholar]
- Torabinejad, M.; Hong, C.U.; McDonald, F.; Pitt Ford, T.R. Physical and Chemical Properties of a New Root-End Filling Material. J. Endod. 1995, 21, 349–353. [Google Scholar] [CrossRef]
- Mohammadi, Z.; Dummer, P.M.H. Properties and Applications of Calcium Hydroxide in Endodontics and Dental Traumatology. Int. Endod. J. 2011, 44, 697–730. [Google Scholar] [CrossRef] [PubMed]
- Makani, S.; Chesler, M. Rapid Rise of Extracellular PH Evoked by Neural Activity Is Generated by the Plasma Membrane Calcium ATPase. J. Neurophysiol. 2010, 103, 667–676. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | 5′-Sequence-3′ | GenBank Number |
|---|---|---|
| β-actin | Forward TGGCACCCAGCACAATGAA Reverse CTAAGTCATAGTCCGCCTAGAAGCA | NM_001101.5 |
| PMCA1 | Forward ACCATATGCTAGAATGCCCACCTC Reverse CTGGTGAAATCTGGGCCCTAAC | NM_001001323.2 |
| PMCA2 | Forward AGAGCTTCCGCATGTACAGCAA Reverse CAAGCCATGGGCTCAATCAC | NM_001001331.4 |
| PMCA3 | Forward CGTAACGTCTATGACAGCATCTCCA Reverse TCCATGATCAAGTTCACCCACAA | NM_001001344.2 |
| PMCA4 | Forward TGGCATGGTTAAATCTGAATGG Reverse CTGCTTCAATTGTAAGGCAAAGG | NM_001001396.2 |
| DMP-1 | Forward TCCAGTCTCACAGCAGCTCA Reverse TCTCCGTGGAGTTGCTATCTTC | NM_004407.4 |
| DSPP | Forward TGATAGCAGTGACAGCACATCTGAC Reverse GTTGTTACCGTTACCAGACTTGCTC | NM_014208.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimura, M.; Mochizuki, H.; Satou, R.; Iwasaki, M.; Kokubu, E.; Kono, K.; Nomura, S.; Sakurai, T.; Kuroda, H.; Shibukawa, Y. Plasma Membrane Ca2+–ATPase in Rat and Human Odontoblasts Mediates Dentin Mineralization. Biomolecules 2021, 11, 1010. https://doi.org/10.3390/biom11071010
Kimura M, Mochizuki H, Satou R, Iwasaki M, Kokubu E, Kono K, Nomura S, Sakurai T, Kuroda H, Shibukawa Y. Plasma Membrane Ca2+–ATPase in Rat and Human Odontoblasts Mediates Dentin Mineralization. Biomolecules. 2021; 11(7):1010. https://doi.org/10.3390/biom11071010
Chicago/Turabian StyleKimura, Maki, Hiroyuki Mochizuki, Ryouichi Satou, Miyu Iwasaki, Eitoyo Kokubu, Kyosuke Kono, Sachie Nomura, Takeshi Sakurai, Hidetaka Kuroda, and Yoshiyuki Shibukawa. 2021. "Plasma Membrane Ca2+–ATPase in Rat and Human Odontoblasts Mediates Dentin Mineralization" Biomolecules 11, no. 7: 1010. https://doi.org/10.3390/biom11071010
APA StyleKimura, M., Mochizuki, H., Satou, R., Iwasaki, M., Kokubu, E., Kono, K., Nomura, S., Sakurai, T., Kuroda, H., & Shibukawa, Y. (2021). Plasma Membrane Ca2+–ATPase in Rat and Human Odontoblasts Mediates Dentin Mineralization. Biomolecules, 11(7), 1010. https://doi.org/10.3390/biom11071010