
Associations of Circulating Irisin with FNDC5 Expression in Fat and Muscle in Type 1 and Type 2 Diabetic Mice




Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.1.1. Streptozotocin-Induced Type 1 Diabetic Mice
2.1.2. High-Fat Diet-Induced Obese Mice
2.1.3. db/m and db/db Mice
2.2. Blood Glucose, Lipid, and Hormonal Analysis
2.3. Organ Collection
2.4. Gene Expression Analysis by Quantitative PCR
2.5. Statistical Analysis
3. Results
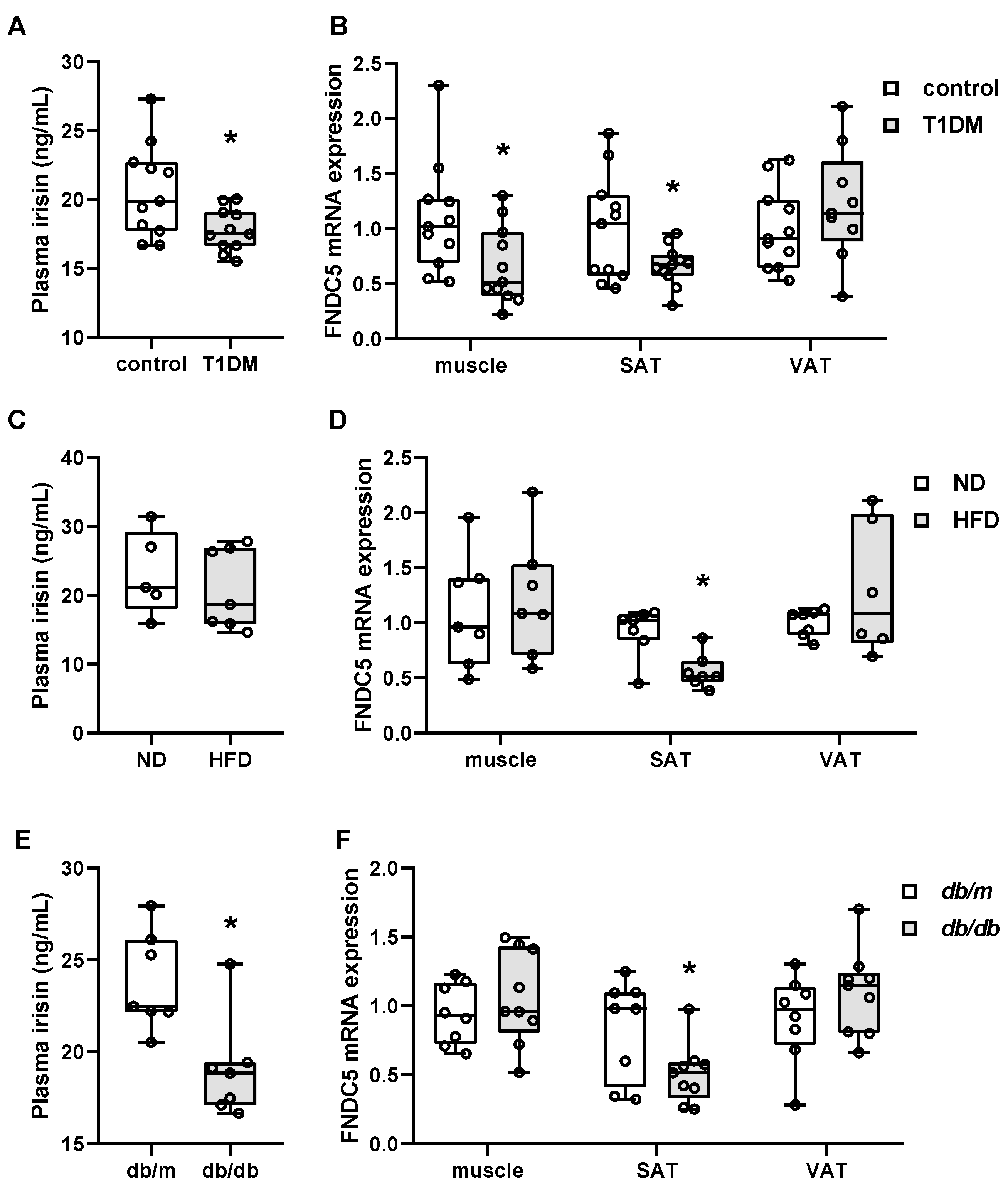
3.1. FNDC5 Expression and Circulating Irisin Levels in Streptozotocin-Induced Type 1 Diabetic Mice
3.2. FNDC5 Expression and Circulating Irisin Levels in High-Fat Diet-Induced Obese Mice
3.3. FNDC5 Expression and Circulating Irisin Levels in db/m and db/db Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bostroem, P.; Wu, J.; Jedrychowski, M.P.; Korde, A.; Ye, L.; Lo, J.C.; Rasbach, K.A.; Bostroem, E.A.; Choi, J.H.; Long, J.Z.; et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nat. Cell Biol. 2012, 481, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Lira, V.A.; Benton, C.R.; Yan, Z.; Bonen, A. PGC-1α regulation by exercise training and its influences on muscle function and insulin sensitivity. Am. J. Physiol. Metab. 2010, 299, E145–E161. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, B.K.; Febbraio, M.A. Muscles, exercise and obesity: Skeletal muscle as a secretory organ. Nat. Rev. Endocrinol. 2012, 8, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, A.; Heeren, J. Adipose tissue browning and metabolic health. Nat. Rev. Endocrinol. 2014, 10, 24–36. [Google Scholar] [CrossRef]
- Panati, K.; Suneetha, Y.; Narala, V.R. Irisin/FNDC5—An updated review. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 689–697. [Google Scholar] [PubMed]
- Xin, C.; Liu, J.; Zhang, J.; Zhu, D.; Wang, H.; Xiong, L.; Lee, Y.; Ye, J.; Lian, K.; Xu, C.; et al. Irisin improves fatty acid oxidation and glucose utilization in type 2 diabetes by regulating the AMPK signaling pathway. Int. J. Obes. 2016, 40, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.-Q.; Chen, D.; Sun, H.-J.; Ding, L.; Wang, J.-J.; Chen, Q.; Li, Y.-H.; Zhou, Y.-B.; Han, Y.; Zhang, F.; et al. FNDC5 overexpression and irisin ameliorate glucose/lipid metabolic derangements and enhance lipolysis in obesity. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2015, 1852, 1867–1875. [Google Scholar] [CrossRef]
- Ma, E.B.; Sahar, N.E.; Jeong, M.; Huh, J.Y. Irisin Exerts Inhibitory Effect on Adipogenesis Through Regulation of Wnt Signaling. Front. Physiol. 2019, 10, 1085. [Google Scholar] [CrossRef]
- Liu, T.-Y.; Shi, C.-X.; Gao, R.; Sun, H.-J.; Xiong, X.-Q.; Ding, L.; Chen, Q.; Li, Y.-H.; Wang, J.-J.; Kang, Y.-M.; et al. Irisin inhibits hepatic gluconeogenesis and increases glycogen synthesis via the PI3K/Akt pathway in type 2 diabetic mice and hepatocytes. Clin. Sci. 2015, 129, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.-Y.; Xiong, X.-Q.; Ren, X.-S.; Zhao, M.-X.; Shi, C.-X.; Wang, J.-J.; Zhou, Y.-B.; Zhang, F.; Han, Y.; Gao, X.-Y.; et al. FNDC5 Alleviates Hepatosteatosis by Restoring AMPK/mTOR-Mediated Autophagy, Fatty Acid Oxidation, and Lipogenesis in Mice. Diabetes 2016, 65, 3262–3275. [Google Scholar] [CrossRef]
- Duan, H.; Ma, B.; Ma, X.; Wang, H.; Ni, Z.; Wang, B.; Li, X.; Jiang, P.; Umar, M.; Li, M. Anti-diabetic activity of recombinant irisin in STZ-induced insulin-deficient diabetic mice. Int. J. Biol. Macromol. 2016, 84, 457–463. [Google Scholar] [CrossRef]
- Huh, J.Y.; Panagiotou, G.; Mougios, V.; Brinkoetter, M.; Vamvini, M.T.; Schneider, B.E.; Mantzoros, C.S. FNDC5 and irisin in humans: I. Predictors of circulating concentrations in serum and plasma and II. mRNA expression and circulating concentrations in response to weight loss and exercise. Metabolism 2012, 61, 1725–1738. [Google Scholar] [CrossRef] [PubMed]
- Stengel, A.; Hofmann, T.; Goebel-Stengel, M.; Elbelt, U.; Kobelt, P.; Klapp, B.F. Circulating levels of irisin in patients with anorexia nervosa and different stages of obesity—Correlation with body mass index. Peptides 2013, 39, 125–130. [Google Scholar] [CrossRef]
- Crujeiras, A.B.; Zulet, M.A.; Lopez-Legarrea, P.; de la Iglesia, R.; Pardo, M.; Carreira, M.C.; Martínez, J.A.; Casanueva, F.F. Association between circulating irisin levels and the promotion of insulin resistance during the weight maintenance period after a dietary weight-lowering program in obese patients. Metabolism 2014, 63, 520–531. [Google Scholar] [CrossRef]
- Park, K.H.; Zaichenko, L.; Brinkoetter, M.; Thakkar, B.; Sahin-Efe, A.; Joung, K.E.; Tsoukas, M.A.; Geladari, E.V.; Huh, J.Y.; Dincer, F.; et al. Circulating Irisin in Relation to Insulin Resistance and the Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2013, 98, 4899–4907. [Google Scholar] [CrossRef] [PubMed]
- Pardo, M.; Crujeiras, A.B.; Amil, M.; Aguera, Z.; Jiménez-Murcia, S.; Baños, R.; Botella, C.; De La Torre, R.; Estivill, X.; Fagundo, A.B.; et al. Association of Irisin with Fat Mass, Resting Energy Expenditure, and Daily Activity in Conditions of Extreme Body Mass Index. Int. J. Endocrinol. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Moreno-Navarrete, J.M.; Ortega, F.; Serrano, M.; Guerra, E.; Pardo, G.; Tinahones, F.; Ricart, W.; Fernández-Real, J.M. Irisin Is Expressed and Produced by Human Muscle and Adipose Tissue in Association With Obesity and Insulin Resistance. J. Clin. Endocrinol. Metab. 2013, 98, E769–E778. [Google Scholar] [CrossRef]
- Cai, L.; Tan, M.; Tan, W.; Zeng, X.; Wan, N.; Wong, S.H.-S.; O’Reilly, J.; Sun, F.; Yang, J.; Chen, Y. Associations of Circulating Irisin Concentrations With Cardiometabolic Risk Factors Among Children Vary by Physical Activity or Sedentary Time Levels. Front. Endocrinol. 2019, 10, 549. [Google Scholar] [CrossRef] [PubMed]
- Perakakis, N.; Triantafyllou, G.A.; Fernández-Real, J.M.; Huh, J.Y.; Park, K.H.; Seufert, J.; Mantzoros, C.S. Physiology and role of irisin in glucose homeostasis. Nat. Rev. Endocrinol. 2017, 13, 324–337. [Google Scholar] [CrossRef]
- Qiu, S.; Cai, X.; Yin, H.; Zügel, M.; Sun, Z.; Steinacker, J.M.; Schumann, U. Association between circulating irisin and insulin resistance in non-diabetic adults: A meta-analysis. Metabolism 2016, 65, 825–834. [Google Scholar] [CrossRef] [PubMed]
- Sesti, G.; Andreozzi, F.; Fiorentino, T.V.; Mannino, G.C.; Sciacqua, A.; Marini, M.A.; Perticone, F. High circulating irisin levels are associated with insulin resistance and vascular atherosclerosis in a cohort of nondiabetic adult subjects. Acta Diabetol. 2014, 51, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.-H.; Zhang, X.-W.; Chen, W.-K.; Huang, Q.-X.; Chen, Q.-Q. Relationship between serum irisin levels and urinary albumin excretion in patients with type 2 diabetes. J. Diabetes Complicat. 2015, 29, 384–389. [Google Scholar] [CrossRef]
- Liu, J.-J.; Wong, M.D.; Toy, W.C.; Tan, C.S.; Liu, S.; Ng, X.W.; Tavintharan, S.; Sum, C.F.; Lim, S.C. Lower circulating irisin is associated with type 2 diabetes mellitus. J. Diabetes Complicat. 2013, 27, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Ding, Z.; Lv, G.; Li, J.; Zhou, P.; Zhang, J. Lower irisin level in patients with type 2 diabetes mellitus: A case-control study and meta-analysis. J. Diabetes 2015, 8, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Amanat, S.; Sinaei, E.; Panji, M.; MohammadporHodki, R.; Bagheri-Hosseinabadi, Z.; Asadimehr, H.; Fararouei, M.; Dianatinasab, A. A Randomized Controlled Trial on the Effects of 12 Weeks of Aerobic, Resistance, and Combined Exercises Training on the Serum Levels of Nesfatin-1, Irisin-1 and HOMA-IR. Front. Physiol. 2020, 11, 562895. [Google Scholar] [CrossRef] [PubMed]
- Dianatinasab, A.; Koroni, R.; Bahramian, M.; Bagheri-Hosseinabadi, Z.; Vaismoradi, M.; Fararouei, M.; Amanat, S. The effects of aerobic, resistance, and combined exercises on the plasma irisin levels, HOMA-IR, and lipid profiles in women with metabolic syndrome: A randomized controlled trial. J. Exerc. Sci. Fit. 2020, 18, 168–176. [Google Scholar] [CrossRef]
- Hassan, I.I.; Hassan, A.B.; Rajab, H.A.; Saadi, F.S.; Abdulah, D.M.; Majeed, A.A.A.; Khaleel, B.B.; Taher, S.M.; Ahmed, I.H. Association of irisin and oxidative stress with biochemical parameters in patients with metabolic syndrome. Horm. Mol. Biol. Clin. Investig. 2019, 39. [Google Scholar] [CrossRef]
- Ebert, T.; Gebhardt, C.; Scholz, M.; Wohland, T.; Schleinitz, D.; Fasshauer, M.; Blüher, M.; Stumvoll, M.; Kovács, P.; Tönjes, A. Relationship Between 12 Adipocytokines and Distinct Components of the Metabolic Syndrome. J. Clin. Endocrinol. Metab. 2018, 103, 1015–1023. [Google Scholar] [CrossRef]
- Du, X.-L.; Jiang, W.-X.; Lv, Z.-T. Lower Circulating Irisin Level in Patients with Diabetes Mellitus: A Systematic Review and Meta-Analysis. Horm. Metab. Res. 2016, 48, 644–652. [Google Scholar] [CrossRef]
- Roca-Rivada, A.; Castelao, C.; Senin, L.L.; Landrove, M.O.; Baltar, J.; Crujeiras, A.B.; Seoane, L.M.; Casanueva, F.F.; Pardo, M. FNDC5/Irisin Is Not Only a Myokine but Also an Adipokine. PLoS ONE 2013, 8, e60563. [Google Scholar] [CrossRef]
- Kazeminasab, F.; Marandi, S.M.; Ghaedi, K.; Safaeinejad, Z.; Esfarjani, F.; Nasr-Esfahani, M.H. A comparative study on the effects of high-fat diet and endurance training on the PGC-1α-FNDC5/irisin pathway in obese and nonobese male C57BL/6 mice. Appl. Physiol. Nutr. Metab. 2018, 43, 651–662. [Google Scholar] [CrossRef]
- Andrade, J.M.O.; Barcala-Jorge, A.S.; Batista-Jorge, G.C.; Paraíso, A.F.; De Freitas, K.M.; Lelis, D.D.F.; Guimarães, A.L.S.; De Paula, A.M.B.; Santos, S.H.S. Effect of resveratrol on expression of genes involved thermogenesis in mice and humans. Biomed. Pharmacother. 2019, 112, 108634. [Google Scholar] [CrossRef]
- Guilford, B.L.; Parson, J.C.; Grote, C.W.; Vick, S.N.; Ryals, J.M.; Wright, D.E. Increased FNDC5 is associated with insulin resistance in high fat-fed mice. Physiol. Rep. 2017, 5, e13319. [Google Scholar] [CrossRef] [PubMed]
- Kurdiova, T.; Balaz, M.; Vician, M.; Maderova, D.; Vlcek, M.; Valkovic, L.; Srbecky, M.; Imrich, R.; Kyselovicova, O.; Belan, V.; et al. Effects of obesity, diabetes and exercise onFndc5gene expression and irisin release in human skeletal muscle and adipose tissue:in vivoandin vitrostudies. J. Physiol. 2014, 592, 1091–1107. [Google Scholar] [CrossRef]
- Pellegrinelli, V.; Carobbio, S.; Vidal-Puig, A. Adipose tissue plasticity: How fat depots respond differently to pathophysiological cues. Diabetologia 2016, 59, 1075–1088. [Google Scholar] [CrossRef]
- Hwang, I.; Lee, J.; Huh, J.Y.; Park, J.; Lee, H.B.; Ho, Y.-S.; Ha, H. Catalase Deficiency Accelerates Diabetic Renal Injury through Peroxisomal Dysfunction. Diabetes 2012, 61, 728–738. [Google Scholar] [CrossRef]
- Piao, L.; Dorotea, D.; Jiang, S.; Koh, E.H.; Oh, G.T.; Ha, H. Impaired Peroxisomal Fitness in Obese Mice, a Vicious Cycle Exacerbating Adipocyte Dysfunction via Oxidative Stress. Antioxid. Redox Signal. 2019, 31, 1339–1351. [Google Scholar] [CrossRef] [PubMed]
- ZhuGe, D.-L.; Javaid, H.M.A.; Sahar, N.E.; Zhao, Y.-Z.; Huh, J.Y. Fibroblast growth factor 2 exacerbates inflammation in adipocytes through NLRP3 inflammasome activation. Arch. Pharmacal Res. 2020, 43, 1311–1324. [Google Scholar] [CrossRef] [PubMed]
- Irving, B.A.; Robinson, M.M.; Nair, K.S. Age effect on myocellular remodeling: Response to exercise and nutrition in humans. Ageing Res. Rev. 2012, 11, 374–389. [Google Scholar] [CrossRef] [PubMed]
- Pang, M.; Yang, J.; Rao, J.; Wang, H.; Zhang, J.; Wang, S.; Chen, X.; Dong, X. Time-Dependent Changes in Increased Levels of Plasma Irisin and Muscle PGC-1α and FNDC5 after Exercise in Mice. Tohoku J. Exp. Med. 2018, 244, 93–103. [Google Scholar] [CrossRef]
- Abedpoor, N.; Taghian, F.; Ghaedi, K.; Niktab, I.; Safaeinejad, Z.; Rabiee, F.; Tanhaei, S.; Nasr-Esfahani, M.H. PPARγ/Pgc-1α-Fndc5 pathway up-regulation in gastrocnemius and heart muscle of exercised, branched chain amino acid diet fed mice. Nutr. Metab. 2018, 15, 59. [Google Scholar] [CrossRef]
- Wajchenberg, B.L. Subcutaneous and Visceral Adipose Tissue. Endocr. Rev. 2000, 21, 697–738. [Google Scholar] [CrossRef]
- Després, J.-P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nat. Cell Biol. 2006, 444, 881–887. [Google Scholar] [CrossRef]
- Esser, N.; L’Homme, L.; De Roover, A.; Kohnen, L.; Scheen, A.J.; Moutschen, M.; Piette, J.; Legrand-Poels, S.; Paquot, N. Obesity phenotype is related to NLRP3 inflammasome activity and immunological profile of visceral adipose tissue. Diabetologia 2013, 56, 2487–2497. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.S.; Massaro, J.M.; Hoffmann, U.; Pou, K.M.; Maurovich-Horvat, P.; Liu, C.-Y.; Vasan, R.S.; Murabito, J.M.; Meigs, J.B.; Cupples, L.A.; et al. Abdominal Visceral and Subcutaneous Adipose Tissue Compartments: Association with metabolic risk factors in the Framingham Heart Study. Circulation 2007, 116, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Boström, P.; Sparks, L.M.; Ye, L.; Choi, J.H.; Giang, A.-H.; Khandekar, M.; Virtanen, K.A.; Nuutila, P.; Schaart, G.; et al. Beige Adipocytes Are a Distinct Type of Thermogenic Fat Cell in Mouse and Human. Cell 2012, 150, 366–376. [Google Scholar] [CrossRef]
- Varela-Rodríguez, B.M.; Pena-Bello, L.; Juiz-Valiña, P.; Vidal-Bretal, B.; Cordido, F.; Sangiao-Alvarellos, S. FNDC5 expression and circulating irisin levels are modified by diet and hormonal conditions in hypothalamus, adipose tissue and muscle. Sci. Rep. 2016, 6, 29898. [Google Scholar] [CrossRef]
- Gutierrez-Repiso, C.; Garcia-Serrano, S.; Rodriguez-Pacheco, F.; García-Escobar, E.; Haro-Mora, J.J.; Garcia-Arnes, J.; Valdés, S.; Gonzalo, M.; Soriguer, F.; Moreno-Ruiz, F.J.; et al. FNDC5 could be regulated by leptin in adipose tissue. Eur. J. Clin. Investig. 2014, 44, 918–925. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Control | Type 1 Diabetic | |
| Body weight (g) | 26.4 ± 0.3 | 22.3 ± 0.5 * |
| Kidney weight (g) | 0.17 ± 0.01 | 0.21 ± 0.10 * |
| Glucose (mM) | 11.9 ± 0.8 | 28.6 ± 0.7 * |
| HbA1c (%) | 4.2 ± 0.1 | 8.2 ± 0.4 * |
| Free fatty acid (mM) | 0.99 ± 0.12 | 1.37 ± 0.12 * |
| Triglyceride (mM) | 0.24 ± 0.01 | 0.49 ± 0.06 * |
| Total cholesterol (mM) | 2.49 ± 0.05 | 2.55 ± 0.15 |
| LDL/VLDL-C (mM) | 0.48 ± 0.02 | 0.64 ± 0.05 * |
| HDL-C (mM) | 4.06 ± 0.12 | 3.88 ± 0.16 |
| Urine volume (mL/day) | 1.0 ± 0.2 | 18.7 ± 2.2 * |
| Normal Diet | High-Fat Diet | |
| Body weight (g) | 25.6 ± 0.5 | 33.7 ± 1.7 * |
| Glucose (mM) | 9.2 ± 1.6 | 13.2 ± 1.0 * |
| Insulin (ng/mL) | 0.44 ± 0.03 | 0.63 ± 0.1 * |
| HbA1c (%) | 3.9 ± 0.1 | 4.2 ± 0.1 * |
| HOMA-IR | 5.79 ± 1.15 | 10.64 ± 1.64 * |
| GTT AUC | 23,949 ± 720 | 38,944 ± 1048 * |
| Free fatty acid (mM) | 1.42 ± 0.17 | 0.56 ± 0.06 * |
| Triglyceride (mM) | 0.49 ± 0.06 | 0.74 ± 0.10 |
| Total cholesterol (mM) | 1.94 ± 0.31 | 3.01 ± 0.07 * |
| LDL-C (mM) | 0.51 ± 0.09 | 0.61 ± 0.04 |
| HDL-C (mM) | 1.76 ± 0.32 | 2.97 ± 0.07 * |
| Plasma LPO (μM) | 3.4 ± 0.9 | 10.0 ± 0.7 * |
| db/m | db/db | |
| Body weight (g) | 27.7 ± 1.1 | 41.1 ± 2.5 * |
| Subcutaneous fat weight (g) | 0.23 ± 0.03 | 1.32 ± 0.13 * |
| Epididymal fat weight (g) | 0.29 ± 0.04 | 1.10 ± 0.12 * |
| Brown fat weight (g) | 0.19 ± 0.03 | 0.73 ± 0.08 * |
| Kidney weight (g) | 0.28 ± 0.03 | 0.34 ± 0.01 |
| Glucose (mM) | 11.7 ± 1.0 | 34.0 ± 2.3 * |
| HbA1c (%) | 4.0 ± 0.1 | 9.8 ± 0.2 * |
| GTT AUC (mg dL−1 min) | 28113 ± 1281 | 111747 ± 3919 * |
| Urine volume (mL/day) | 0.57 ± 0.19 | 1.96 ± 0.33 * |
| Plasma Irisin | FNDC5 mRNA (Muscle) | FNDC5 mRNA (SAT) | FNDC5 mRNA (VAT) | ||
|---|---|---|---|---|---|
| Plasma irisin | r | 1 | |||
| p-value | |||||
| FNDC5 mRNA (muscle) | r | 0.202 | 1 | ||
| p-value | 0.381 | ||||
| FNDC5 mRNA (SAT) | r | 0.749 * | 0.061 | 1 | |
| p-value | 0.000 | 0.787 | |||
| FNDC5 mRNA (VAT) | r | 0.022 | −0.205 | −0.337 | 1 |
| p-value | 0.928 | 0.401 | 0.146 | ||
| Body weight | r | 0.542 * | 0.527 * | 0.504 * | −0.042 |
| p-value | 0.009 | 0.012 | 0.014 | 0.859 | |
| Blood glucose | r | −0.489 * | −0.381 | −0.467 * | 0.176 |
| p-value | 0.021 | 0.080 | 0.025 | 0.458 | |
| HbA1c | r | −0.450 * | −0.447 * | −0.383 | 0.298 |
| p-value | 0.036 | 0.037 | 0.071 | 0.202 | |
| Free fatty acid | r | −0.449 | −0.170 | −0.425 | 0.079 |
| p-value | 0.054 | 0.487 | 0.062 | 0.755 | |
| Triglyceride | r | −0.359 | −0.547 * | −0.308 | 0.010 |
| p-value | 0.110 | 0.010 | 0.163 | 0.968 | |
| Total cholesterol | r | −0.100 | −0.351 | −0.076 | 0.003 |
| p-value | 0.667 | 0.119 | 0.738 | 0.989 | |
| LDL/VLDL-C | r | −0.322 | −0.494 * | −0.317 | 0.114 |
| p-value | 0.144 | 0.019 | 0.141 | 0.632 | |
| HDL-C | r | 0.364 | −0.065 | 0.183 | −0.161 |
| p-value | 0.096 | 0.779 | 0.414 | 0.509 | |
| Urine volume | r | −0.446 * | −0.510 * | −0.425 * | 0.136 |
| p-value | 0.038 | 0.015 | 0.043 | 0.567 | |
| Plasma Irisin | FNDC5 mRNA (Muscle) | FNDC5 mRNA (SAT) | FNDC5 mRNA (VAT) | ||
|---|---|---|---|---|---|
| Plasma irisin | r | 1 | |||
| p-value | |||||
| FNDC5 mRNA (muscle) | r | 0.271 | 1 | ||
| p-value | 0.394 | ||||
| FNDC5 mRNA (SAT) | r | 0.153 | 0.110 | 1 | |
| p-value | 0.636 | 0.721 | |||
| FNDC5 mRNA (VAT) | r | 0.195 | 0.245 | −0.534 | 1 |
| p-value | 0.565 | 0.420 | 0.074 | ||
| Body weight | r | −0.461 | 0.026 | −0.779* | 0.283 |
| p-value | 0.132 | 0.929 | 0.002 | 0.348 | |
| Fasting glucose | r | 0.067 | 0.386 | −0.450 | 0.427 |
| p-value | 0.836 | 0.173 | 0.123 | 0.146 | |
| Fasting insulin | r | −0.368 | 0.047 | −0.599 | 0.300 |
| p-value | 0.265 | 0.884 | 0.051 | 0.370 | |
| HbA1c | r | −0.303 | 0.245 | −0.444 | 0.266 |
| p-value | 0.339 | 0.398 | 0.129 | 0.380 | |
| HOMA-IR | r | −0.172 | 0.198 | −0.600 | 0.505 |
| p-value | 0.614 | 0.536 | 0.051 | 0.113 | |
| GTT AUC | r | −0.116 | 0.110 | −0.871 * | 0.364 |
| p-value | 0.720 | 0.708 | 0.000 | 0.222 | |
| Free fatty acid | r | −0.029 | −0.069 | 0.655 * | −0.438 |
| p-value | 0.929 | 0.815 | 0.015 | 0.135 | |
| Triglyceride | r | −0.447 | 0.096 | −0.430 | −0.128 |
| p-value | 0.145 | 0.744 | 0.143 | 0.676 | |
| Total cholesterol | r | −0.061 | 0.456 | −0.577 * | 0.213 |
| p-value | 0.850 | 0.101 | 0.039 | 0.484 | |
| LDL-C | r | 0.108 | 0.128 | −0.115 | 0.301 |
| p-value | 0.738 | 0.663 | 0.707 | 0.317 | |
| HDL-C | r | −0.163 | 0.455 | −0.602 * | 0.185 |
| p-value | 0.613 | 0.102 | 0.030 | 0.546 | |
| LPO | r | −0.120 | 0.475 | −0.655 * | 0.349 |
| p-value | 0.710 | 0.086 | 0.015 | 0.242 | |
| Plasma irisin | FNDC5 mRNA (Muscle) | FNDC5 mRNA (SAT) | FNDC5 mRNA (VAT) | ||
|---|---|---|---|---|---|
| Plasma irisin | r | 1 | |||
| p-value | |||||
| FNDC5 mRNA (muscle) | r | −0.282 | 1 | ||
| p-value | 0.351 | ||||
| FNDC5 mRNA (SAT) | r | 0.547 | −0.150 | 1 | |
| p-value | 0.160 | 0.593 | |||
| FNDC5 mRNA (VAT) | r | −0.171 | 0.121 | −0.538 * | 1 |
| p-value | 0.637 | 0.643 | 0.047 | ||
| Body weight | r | −0.688 * | 0.067 | −0.542 * | 0.079 |
| p-value | 0.009 | 0.778 | 0.037 | 0.763 | |
| SAT weight | r | −0.611 * | 0.037 | −0.619 * | 0.063 |
| p-value | 0.027 | 0.877 | 0.014 | 0.809 | |
| VAT weight | r | −0.848 * | 0.150 | −0.537 * | 0.048 |
| p-value | 0.000 | 0.529 | 0.039 | 0.854 | |
| Brown fat weight | r | −0.708 * | 0.149 | −0.587 * | 0.305 |
| p-value | 0.007 | 0.531 | 0.022 | 0.234 | |
| Kidney weight | r | −0.444 | 0.347 | −0.451 | −0.226 |
| p-value | 0.129 | 0.133 | 0.092 | 0.384 | |
| Blood glucose | r | −0.593 * | 0.371 | −0.751 * | 0.050 |
| p-value | 0.033 | 0.118 | 0.002 | 0.854 | |
| HbA1c | r | −0.602 * | 0.093 | −0.727 * | 0.087 |
| p-value | 0.030 | 0.706 | 0.003 | 0.748 | |
| GTT AUC | r | −0.649 * | 0.294 | −0.730 * | 0.070 |
| p-value | 0.016 | 0.209 | 0.002 | 0.791 | |
| Urine volume | r | −0.259 | 0.345 | −0.569* | −0.022 |
| p-value | 0.392 | 0.137 | 0.027 | 0.933 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Piao, L.; Ma, E.B.; Ha, H.; Huh, J.Y. Associations of Circulating Irisin with FNDC5 Expression in Fat and Muscle in Type 1 and Type 2 Diabetic Mice. Biomolecules 2021, 11, 322. https://doi.org/10.3390/biom11020322
Jiang S, Piao L, Ma EB, Ha H, Huh JY. Associations of Circulating Irisin with FNDC5 Expression in Fat and Muscle in Type 1 and Type 2 Diabetic Mice. Biomolecules. 2021; 11(2):322. https://doi.org/10.3390/biom11020322
Chicago/Turabian StyleJiang, Songling, Lingjuan Piao, Eun Bi Ma, Hunjoo Ha, and Joo Young Huh. 2021. "Associations of Circulating Irisin with FNDC5 Expression in Fat and Muscle in Type 1 and Type 2 Diabetic Mice" Biomolecules 11, no. 2: 322. https://doi.org/10.3390/biom11020322
APA StyleJiang, S., Piao, L., Ma, E. B., Ha, H., & Huh, J. Y. (2021). Associations of Circulating Irisin with FNDC5 Expression in Fat and Muscle in Type 1 and Type 2 Diabetic Mice. Biomolecules, 11(2), 322. https://doi.org/10.3390/biom11020322