
The Influence of Coumestrol on Sphingolipid Signaling Pathway and Insulin Resistance Development in Primary Rat Hepatocytes

 , , , , , and
, , , , , and
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Study Design
2.2. Experimental Procedures
2.3. Immunoblotting Analyses
2.4. Intracellular Sphingolipid Analyses
2.5. Data Analysis
3. Results
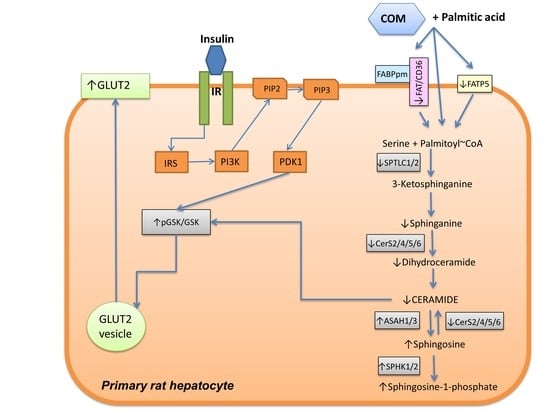
3.1. Effects of Coumestrol on Sphingolipid Concentrations in Primary Rat Hepatocytes
3.2. Effects of Coumestrol on the Expression of Insulin Signaling Pathway Proteins in Primary Rat Hepatocytes
3.3. Effects of Coumestrol on the Expression of Enzymes Involved in Ceramide De Novo Synthesis Pathway in Primary Rat Hepatocytes
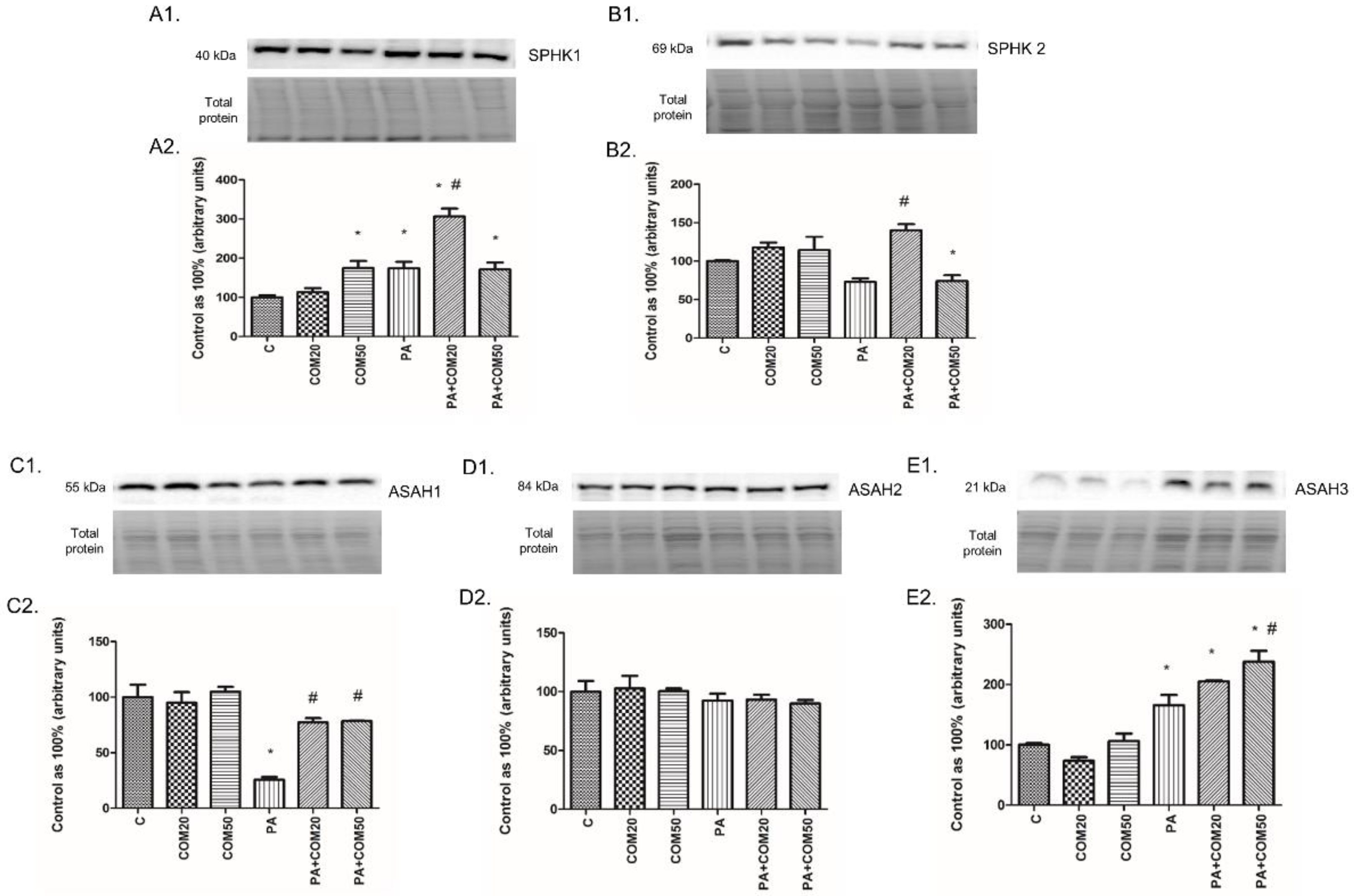
3.4. Effects of Coumestrol on the Expression of Enzymes That Share Ceramide De Novo Synthesis and Salvage Pathways in Primary Rat Hepatocytes
3.5. Effects of Coumestrol on the Expression of Enzymes Involved in the Sphingolipid Salvage Pathway in Primary Rat Hepatocytes
3.6. Effects of Coumestrol on the Expression of Proteins Involved in Fatty Acid Uptake and β-Oxidation in Primary Rat Hepatocytes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| Akt | protein kinase B |
| ANOVA | analysis of variance |
| ASAH1 | acid ceramidase |
| ASAH2 | neutral ceramidase |
| ASAH3 | alkaline ceramidase |
| BSA | bovine serum albumin |
| CER | ceramide |
| COM | coumestrol |
| FABPpm | fatty acid binding protein |
| FAT/CD36 | fatty acid translocase |
| FATP1 | fatty acid transport protein 1 |
| FATP2 | fatty acid transport protein 2 |
| FATP5 | fatty acid transport protein 5 |
| FBS | fetal bovine serum |
| FFA | free fatty acids |
| GSK-3β | glycogen synthase kinase 3β |
| HBBS | Hank’s Balanced Salt Solution |
| HPLC | high-performance liquid chromatography |
| HRP | horseradish peroxidase |
| IL-6 | interleukin 6 |
| CerS 4 | (dihydro)ceramide synthase 4 |
| CerS 6 | (dihydro)ceramide synthase 6 |
| PA | palmitic acid |
| pAkt | phosphorylated protein kinase B |
| PBS | phosphate-buffered saline |
| pGSK-3β | phosphorylated glycogen synthase kinase 3β |
| PI3K | phosphatidylinositol-3 kinase |
| S1P | sphingosine-1-phosphate |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| SFA | sphinganine |
| SFO | sphingosine |
| SPHK1 | sphingosine kinase |
| SPHK2 | sphingosine kinase 2 |
| SPTLC1 | serine palmitoyltransferase 1 |
| SPTLC2 | serine palmitoyltransferase 2 |
| T2DM | type II diabetes mellitus |
| TBS | Tris-buffered saline |
| TNF-α | tumor necrosis factor α |
| β-HADH | β-hydroxyacyl-CoA-dehydrogenase |
References
- Bhupathiraju, S.N.; Hu, F.B. Epidemiología de la Obesidad y la Diabetes y sus Complicaciones Cardiovasculares. Circ. Res. 2016, 118, 1723–1735. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.H.; Hu, F.B. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Perry, R.J.; Samuel, V.T.; Petersen, K.F.; Shulman, G.I. The role of hepatic lipids in hepatic insulin resistance and type 2 diabetes. Nature 2014, 510, 84–91. [Google Scholar] [CrossRef]
- Lara-Castro, C.; Garvey, W.T. Intracellular Lipid Accumulation in Liver and Muscle and the Insulin Resistance Syndrome. Endocrinol. Metab. Clin. N. Am. 2008, 37, 841–856. [Google Scholar] [CrossRef]
- Sokolowska, E.; Blachnio-Zabielska, A. The Role of Ceramides in Insulin Resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef]
- Konstantynowicz-Nowicka, K.; Harasim, E.; Baranowski, M.; Chabowski, A. New evidence for the role of ceramide in the development of hepatic insulin resistance. PLoS ONE 2015, 10, e0116858. [Google Scholar] [CrossRef]
- Kitatani, K.; Idkowiak-Baldys, J.; Hannun, Y.A. The sphingolipid salvage pathway in ceramide metabolism and signaling. Cell. Signal. 2008, 20, 1010–1018. [Google Scholar] [CrossRef]
- Bandet, C.L.; Tan-Chen, S.; Bourron, O.; Le Stunff, H.; Hajduch, E. Sphingolipid metabolism: New insight into ceramide-induced lipotoxicity in muscle cells. Int. J. Mol. Sci. 2019, 20, 479. [Google Scholar] [CrossRef]
- Charytoniuk, T.; Iłowska, N.; Berk, K.; Chabowski, A.; Konstantynowicz-, K. The effect of enterolactone on sphingolipid pathway and hepatic insulin resistance development in HepG2 cells. Life Sci. 2018, 217, 1–7. [Google Scholar] [CrossRef]
- Petersen, M.C.; Shulman, G.I. Roles of Diacylglycerols and Ceramides in Hepatic Insulin Resistance. Trends Pharmacol. Sci. 2017, 38, 649–665. [Google Scholar] [CrossRef]
- Montero, G.; Arriagada, F.; Günther, G.; Bollo, S.; Mura, F.; Berríos, E.; Morales, J. Phytoestrogen coumestrol: Antioxidant capacity and its loading in albumin nanoparticles. Int. J. Pharm. 2019, 562, 86–95. [Google Scholar] [CrossRef]
- Li, H.; Kang, J.H.; Han, J.M.; Cho, M.H.; Chung, Y.J.; Park, K.H.; Shin, D.H.; Park, H.Y.; Choi, M.S.; Jeong, T.S. Anti-Obesity Effects of Soy Leaf via Regulation of Adipogenic Transcription Factors and Fat Oxidation in Diet-Induced Obese Mice and 3T3-L1 Adipocytes. J. Med. Food 2015, 18, 899–908. [Google Scholar] [CrossRef]
- Hopert, A.C.; Bayer, A.; Frank, K.; Strunck, E.; Wünsche, W.; Vollmer, G. Characterization of estrogenicity of phytoestrogens in an endometrial-derived experimental model. Environ. Health Perspect. 1998, 106, 581–586. [Google Scholar] [CrossRef]
- Glisic, M.; Kastrati, N.; Gonzalez-Jaramillo, V.; Bramer, W.M.; Ahmadizar, F.; Chowdhury, R.; Danser, A.H.J.; Roks, A.J.M.; Voortman, T.; Franco, O.H.; et al. Associations between phytoestrogens, glucose homeostasis, and risk of diabetes in women: A systematic review and meta-analysis. Adv. Nutr. 2018, 9, 726–740. [Google Scholar] [CrossRef] [PubMed]
- Rietjens, I.M.C.M.; Louisse, J.; Beekmann, K. The potential health effects of dietary phytoestrogens. Br. J. Pharmacol. 2017, 174, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Sankar, P.; Zachariah, B.; Vickneshwaran, V.; Jacob, S.E.; Sridhar, M.G. Amelioration of oxidative stress and insulin resistance by soy isoflavones (from Glycine max) in ovariectomized Wistar rats fed with high fat diet: The molecular mechanisms. Exp. Gerontol. 2015, 63, 67–75. [Google Scholar] [CrossRef]
- Amanat, S.; Eftekhari, M.H.; Fararouei, M.; Bagheri Lankarani, K.; Massoumi, S.J. Genistein supplementation improves insulin resistance and inflammatory state in non-alcoholic fatty liver patients: A randomized, controlled trial. Clin. Nutr. 2018, 37, 1210–1215. [Google Scholar] [CrossRef]
- Oza, M.J.; Kulkarni, Y.A. Biochanin A improves insulin sensitivity and controls hyperglycemia in type 2 diabetes. Biomed. Pharmacother. 2018, 107, 1119–1127. [Google Scholar] [CrossRef]
- Seglen, P.O. Preparation of Isolated Rat Liver Cells. In Methods in Cell Biology; Norsk Hydro’s Institute for Cancer Research, Department of Tissue Culture, The Norwegian Radium Hospital: Oslo, Norway, 1976; Volume 13, pp. 29–83. [Google Scholar]
- Moya, M.; Benet, M.; Guzmán, C.; Tolosa, L.; García-Monzón, C.; Pareja, E.; Castell, J.V.; Jover, R. Foxa1 reduces lipid accumulation in human hepatocytes and is down-regulated in nonalcoholic fatty liver. PLoS ONE 2012, 7, e30014. [Google Scholar] [CrossRef]
- Chabowski, A.; Zendzian-Piotrowska, M.; Konstantynowicz, K.; Pankiewicz, W.; Mikłosz, A.; Łukaszuk, B.; Górski, J. Fatty acid transporters involved in the palmitate and oleate induced insulin resistance in primary rat hepatocytes. Acta Physiol. 2013, 207, 346–357. [Google Scholar] [CrossRef]
- Knapp, M.; Lisowska, A.; Zabielski, P.; Musiał, W.; Baranowski, M. Sustained decrease in plasma sphingosine-1-phosphate concentration and its accumulation in blood cells in acute myocardial infarction. Prostaglandins Other Lipid Mediat. 2013, 106, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [PubMed]
- Hermida, M.A.; Dinesh Kumar, J.; Leslie, N.R. GSK3 and its interactions with the PI3K/AKT/mTOR signalling network. Adv. Biol. Regul. 2017, 65, 5–15. [Google Scholar] [CrossRef]
- Lee, J.; Kim, M.S. The role of GSK3 in glucose homeostasis and the development of insulin resistance. Diabetes Res. Clin. Pract. 2007, 77, 49–57. [Google Scholar] [CrossRef]
- Bourbon, N.A.; Sandirasegarane, L.; Kester, M. Ceramide-induced inhibition of Akt is mediated through protein kinase Cζ. Implications for growth arrest. J. Biol. Chem. 2002, 277, 3286–3292. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ge, Z.; Ang, S.; Meng, R.; Bi, Y.; Zhu, D. Erythropoietin ameliorates PA-induced insulin resistance through the IRS/AKT/FOXO1 and GSK-3β signaling pathway, and inhibits the inflammatory response in HepG2 cells. Mol. Med. Rep. 2017, 16, 2295–2301. [Google Scholar] [CrossRef]
- Lin, C.F.; Chen, C.L.; Chiang, C.W.; Jan, M.S.; Huang, W.C.; Lin, Y.S. GSK-3β acts downstream of PP2A and the PI 3-kinase-Akt pathway, and upstream of caspase-2 in ceramide-induced mitochondrial apoptosis. J. Cell Sci. 2007, 120, 2935–2943. [Google Scholar] [CrossRef]
- Vinayavekhin, N.; Sueajai, J.; Chaihad, N.; Panrak, R.; Chokchaisiri, R.; Sangvanich, P.; Suksamrarn, A.; Piyachaturawat, P. Serum lipidomics analysis of ovariectomized rats under Curcuma comosa treatment. J. Ethnopharmacol. 2016, 192, 273–282. [Google Scholar] [CrossRef]
- Bonen, A.; Jain, S.S.; Snook, L.A.; Han, X.X.; Yoshida, Y.; Buddo, K.H.; Lally, J.S.; Pask, E.D.; Paglialunga, S.; Beaudoin, M.S.; et al. Extremely rapid increase in fatty acid transport and intramyocellular lipid accumulation but markedly delayed insulin resistance after high fat feeding in rats. Diabetologia 2015, 58, 2381–2391. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Chen, J.; Lay, A.; Don, A.; Vadas, M.; Xia, P. Loss of sphingosine kinase 1 predisposes to the onset of diabetes via promoting pancreatic β-cell death in diet-induced obese mice. FASEB J. 2013, 27, 4294–4304. [Google Scholar] [CrossRef]
- Fang, H.; Feng, Q.; Shi, Y.; Zhou, J.; Wang, Q.; Zhong, L. Hepatic insulin resistance induced by mitochondrial oxidative stress can be ameliorated by sphingosine 1-phosphate. Mol. Cell. Endocrinol. 2020, 501, 110660. [Google Scholar] [CrossRef]
- Obinata, H.; Hla, T. Sphingosine 1-phosphate and inflammation. Int. Immunol. 2019, 31, 617–625. [Google Scholar] [CrossRef]
- Cartier, A.; Hla, T. Sphingosine 1-phosphate: Lipid signaling in pathology and therapy. Science 2019, 366, 6463. [Google Scholar] [CrossRef]
- Kleuser, B. Divergent role of sphingosine 1-phosphate in liver health and disease. Int. J. Mol. Sci. 2018, 19, 722. [Google Scholar] [CrossRef] [PubMed]
- Raichur, S.; Wang, S.T.; Chan, P.W.; Li, Y.; Ching, J.; Chaurasia, B.; Dogra, S.; Öhman, M.K.; Takeda, K.; Sugii, S.; et al. CerS2 haploinsufficiency inhibits β-oxidation and confers susceptibility to diet-induced steatohepatitis and insulin resistance. Cell Metab. 2014, 20, 687–695. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zywno, H.; Bzdega, W.; Kolakowski, A.; Kurzyna, P.; Harasim-Symbor, E.; Sztolsztener, K.; Chabowski, A.; Konstantynowicz-Nowicka, K. The Influence of Coumestrol on Sphingolipid Signaling Pathway and Insulin Resistance Development in Primary Rat Hepatocytes. Biomolecules 2021, 11, 268. https://doi.org/10.3390/biom11020268
Zywno H, Bzdega W, Kolakowski A, Kurzyna P, Harasim-Symbor E, Sztolsztener K, Chabowski A, Konstantynowicz-Nowicka K. The Influence of Coumestrol on Sphingolipid Signaling Pathway and Insulin Resistance Development in Primary Rat Hepatocytes. Biomolecules. 2021; 11(2):268. https://doi.org/10.3390/biom11020268
Chicago/Turabian StyleZywno, Hubert, Wiktor Bzdega, Adrian Kolakowski, Piotr Kurzyna, Ewa Harasim-Symbor, Klaudia Sztolsztener, Adrian Chabowski, and Karolina Konstantynowicz-Nowicka. 2021. "The Influence of Coumestrol on Sphingolipid Signaling Pathway and Insulin Resistance Development in Primary Rat Hepatocytes" Biomolecules 11, no. 2: 268. https://doi.org/10.3390/biom11020268
APA StyleZywno, H., Bzdega, W., Kolakowski, A., Kurzyna, P., Harasim-Symbor, E., Sztolsztener, K., Chabowski, A., & Konstantynowicz-Nowicka, K. (2021). The Influence of Coumestrol on Sphingolipid Signaling Pathway and Insulin Resistance Development in Primary Rat Hepatocytes. Biomolecules, 11(2), 268. https://doi.org/10.3390/biom11020268