
Metabolic Profiles, Bioactive Compounds, and Antioxidant Capacity in Lentinula edodes Cultivated on Log versus Sawdust Substrates

Abstract
1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. Mushroom Material
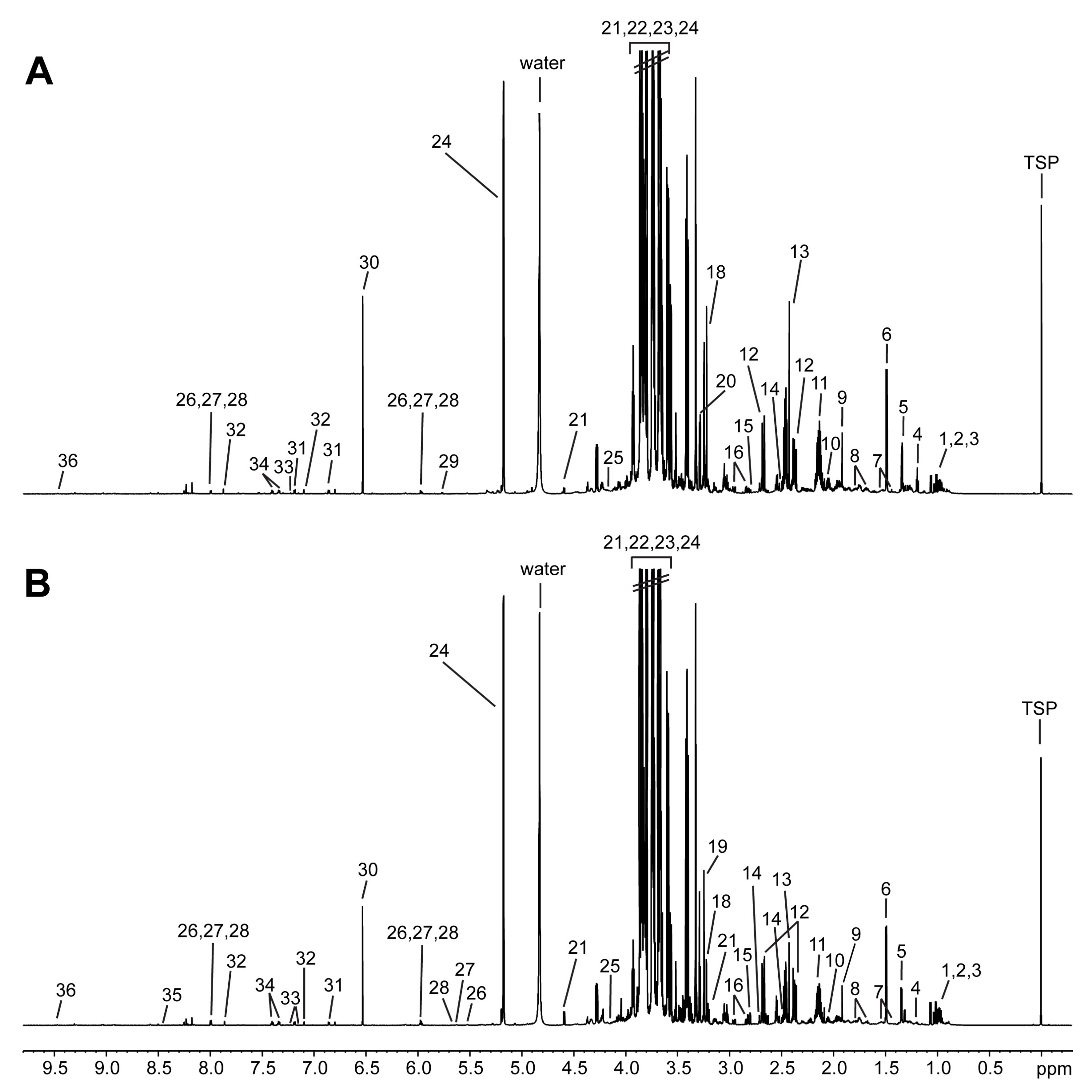
2.3. Metabolite Profiling Using 1H NMR Analysis
2.4. UPLC-QTOF-MS Analysis
2.5. Preparation of the Methanolic Extract
2.6. Determination of Bioactive Compounds
2.7. Determination of Antioxidant Capacity
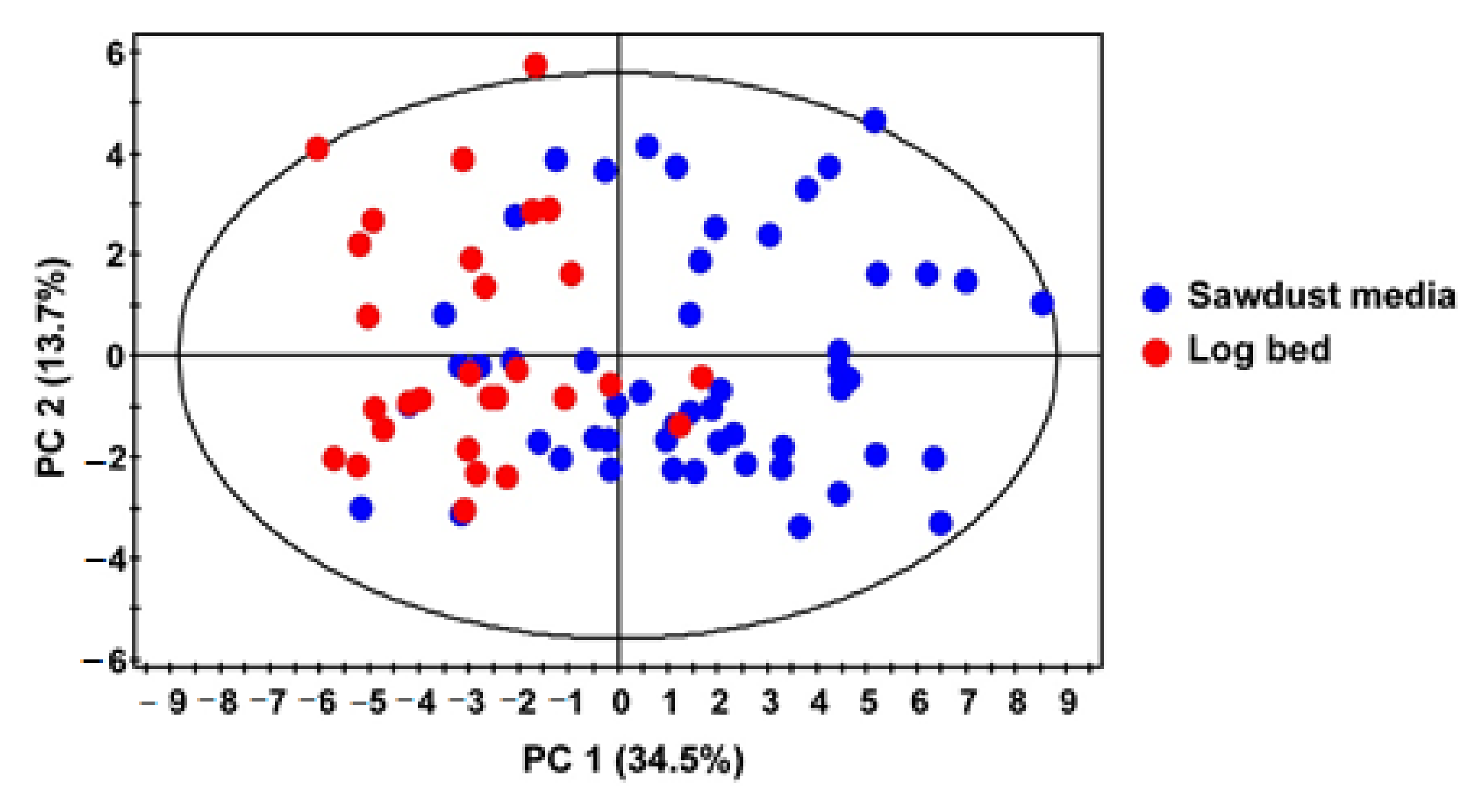
2.8. Statistical Analysis
3. Results and Discussion
3.1. Metabolite Profiling Using 1H NMR
3.2. Metabolite Profiling Using UPLC-QTOF-MS
3.3. Extraction Yield and Analysis of Bioactive Components
3.4. Determination of Antioxidant Capacity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Miles, P.G.; Chang, S.-T. Mushrooms: Cultivation, Nutritional Value, Medicinal Effect, and Environmental Impact; CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Chen, L.; Gong, Y.; Cai, Y.; Liu, W.; Zhou, Y.; Xiao, Y.; Xu, Z.; Liu, Y.; Lei, X.; Wang, G. Genome sequence of the edible cultivated mushroom Lentinula edodes (Shiitake) reveals insights into lignocellulose degradation. PLoS ONE 2016, 11, e0160336. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhao, Y.; Huang, J.; Zeng, H.; Zheng, B. Effects of different drying methods on the product quality and volatile compounds of whole shiitake mushrooms. Food Chem. 2016, 197, 714–722. [Google Scholar] [CrossRef]
- Boa, E.R. Wild Edible Fungi: A Global Overview of Their Use and Importance to People; FAO: Rome, Italy, 2004; p. 163. [Google Scholar]
- Mattila, P.; Salo-Väänänen, P.; Könkö, K.; Aro, H.; Jalava, T. Basic composition and amino acid contents of mushrooms cultivated in Finland. J. Agric. Food Chem. 2002, 50, 6419–6422. [Google Scholar] [CrossRef]
- Fukushima, M.; Ohashi, T.; Fujiwara, Y.; Sonoyama, K.; Nakano, M. Cholesterol-lowering effects of maitake (Grifola frondosa) fiber, shiitake (Lentinus edodes) fiber, and enokitake (Flammulina velutipes) fiber in rats. Exp. Biol. Med. 2001, 226, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Morales, D.; Gil-Ramirez, A.; Smiderle, F.R.; Piris, A.J.; Ruiz-Rodriguez, A.; Soler-Rivas, C. Vitamin D-enriched extracts obtained from shiitake mushrooms (Lentinula edodes) by supercritical fluid extraction and UV-irradiation. Innov. Food Sci. Emerg. Technol. 2017, 41, 330–336. [Google Scholar] [CrossRef]
- Xue, Z.; Chen, Y.; Jia, Y.; Wang, Y.; Lu, Y.; Chen, H.; Zhang, M. Structure, thermal and rheological properties of different soluble dietary fiber fractions from mushroom Lentinula edodes (Berk.) Pegler residues. Food Hydrocoll. 2019, 95, 10–18. [Google Scholar] [CrossRef]
- Carneiro, A.A.; Ferreira, I.C.; Dueñas, M.; Barros, L.; Da Silva, R.; Gomes, E.; Santos-Buelga, C. Chemical composition and antioxidant activity of dried powder formulations of Agaricus blazei and Lentinus edodes. Food Chem. 2013, 138, 2168–2173. [Google Scholar] [CrossRef] [PubMed]
- Soković, M.; Ćirić, A.; Glamočlija, J.; Stojković, D. The bioactive properties of mushrooms. Wild Plants Mushrooms Nuts Funct. Food Prop. Appl. 2017, 4, 83–122. [Google Scholar]
- Zhuang, H.; Chen, Z.; Feng, T.; Yang, Y.; Zhang, J.; Liu, G.; Li, Z.; Ye, R. Characterization of Lentinus edodes β-glucan influencing the in vitro starch digestibility of wheat starch gel. Food Chem. 2017, 224, 294–301. [Google Scholar] [CrossRef]
- Pire, D.; Wright, J.; Albertó, E. Cultivation of shiitake using sawdust from widely available local woods in Argentina. Micol. Apl. Int. 2001, 13, 87–91. [Google Scholar]
- Tabata, T.; Tomioka, K.; Iwasaka, Y.; Shinohara, H.; Ogura, T. Comparison of chemical compositions of shiitake (Lentinus edodes (Berk.) Sing) cultivated on logs and sawdust substrate. Food Sci. Technol. Res. 2006, 12, 252–255. [Google Scholar] [CrossRef][Green Version]
- Barros, L.; Calhelha, R.C.; Vaz, J.A.; Ferreira, I.C.; Baptista, P.; Estevinho, L.M. Antimicrobial activity and bioactive compounds of Portuguese wild edible mushrooms methanolic extracts. Eur. Food Res. Technol. 2007, 225, 151–156. [Google Scholar] [CrossRef]
- Woldegiorgis, A.Z.; Abate, D.; Haki, G.D.; Ziegler, G.R. Antioxidant property of edible mushrooms collected from Ethiopia. Food Chem. 2014, 157, 30–36. [Google Scholar] [CrossRef]
- Huang, R.; Wu, W.; Shen, S.; Fan, J.; Chang, Y.; Chen, S.; Ye, X. Evaluation of colorimetric methods for quantification of citrus flavonoids to avoid misuse. Anal. Methods 2018, 10, 2575–2587. [Google Scholar] [CrossRef]
- Erbiai, E.H.; da Silva, L.P.; Saidi, R.; Lamrani, Z.; Esteves da Silva, J.C.; Maouni, A. Chemical Composition, Bioactive Compounds, and Antioxidant Activity of Two Wild Edible Mushrooms Armillaria mellea and Macrolepiota procera from Two Countries (Morocco and Portugal). Biomolecules 2021, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-N.; Sung, T.-J.; Chou, C.-H.; Liu, K.-L.; Hsieh, L.-P.; Hsieh, C.-W. Characterization and antioxidant activities of yellow strain Flammulina velutipes (Jinhua mushroom) polysaccharides and their effects on ROS content in L929 cell. Antioxidants 2019, 8, 298. [Google Scholar] [CrossRef] [PubMed]
- Worley, B.; Halouska, S.; Powers, R. Utilities for quantifying separation in PCA/PLS-DA scores plots. Anal. Biochem. 2013, 433, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Dong, M.; Qin, L.; Xue, J.; Du, M.; Lin, S.-Y.; Xu, X.-B.; Zhu, B.-W. Simultaneous quantification of free amino acids and 5′-nucleotides in shiitake mushrooms by stable isotope labeling-LC-MS/MS analysis. Food Chem. 2018, 268, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Manninen, H.; Rotola-Pukkila, M.; Aisala, H.; Hopia, A.; Laaksonen, T. Free amino acids and 5′-nucleotides in Finnish forest mushrooms. Food Chem. 2018, 247, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Ming, T.; Li, J.; Huo, P.; Wei, Y.; Chen, X. Analysis of free amino acids in Russula griseocarnosa harvested at different stages of maturity using iTRAQ®-LC-MS/MS. Food Anal. Methods 2014, 7, 1816–1823. [Google Scholar] [CrossRef]
- Gao, S.; Huang, Z.; Feng, X.; Bian, Y.; Huang, W.; Liu, Y. Bioconversion of rice straw agro-residues by Lentinula edodes and evaluation of non-volatile taste compounds in mushrooms. Sci. Rep. 2020, 10, 1814. [Google Scholar] [CrossRef]
- Li, W.; Chen, W.; Yang, Y.; Zhang, J.; Feng, J.; Yu, H.; Zhou, S.; Li, X.; Liu, Y. Effects of culture substrates on taste component content and taste quality of Lentinula edodes. Int. J. Food Sci. Technol. 2017, 52, 981–991. [Google Scholar] [CrossRef]
- Bar-Peled, M.; O’Neill, M.A. Plant nucleotide sugar formation, interconversion, and salvage by sugar recycling. Annu. Rev. Plant Biol. 2011, 62, 127–155. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.; Duthie, G. Plant secondary metabolites and gut health: The case for phenolic acids. Proc. Nutr. Soc. 2011, 70, 389–396. [Google Scholar] [CrossRef]
- Pontiki, E.; Hadjipavlou-Litina, D.; Litinas, K.; Geromichalos, G. Novel cinnamic acid derivatives as antioxidant and anticancer agents: Design, synthesis and modeling studies. Molecules 2014, 19, 9655–9674. [Google Scholar] [CrossRef]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef] [PubMed]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant properties of ferulic acid and its possible application. Ski. Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial properties of green tea catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-H.; Lin, H.-C.; Mau, J.-L. Antioxidant properties of several commercial mushrooms. Food Chem. 2002, 77, 229–235. [Google Scholar] [CrossRef]
- Cheung, L.; Cheung, P.C. Mushroom extracts with antioxidant activity against lipid peroxidation. Food Chem. 2005, 89, 403–409. [Google Scholar] [CrossRef]
- Cheung, L.; Cheung, P.C.; Ooi, V.E. Antioxidant activity and total phenolics of edible mushroom extracts. Food Chem. 2003, 81, 249–255. [Google Scholar] [CrossRef]
- Velioglu, Y.; Mazza, G.; Gao, L.; Oomah, B. Antioxidant activity and total phenolics in selected fruits, vegetables, and grain products. J. Agric. Food Chem. 1998, 46, 4113–4117. [Google Scholar] [CrossRef]
- Alkin, M.; Söğüt, E.; Seydim, A.C. Determination of bioactive properties of different edible mushrooms from Turkey. J. Food Meas. Charact. 2021, 15, 3608–3617. [Google Scholar] [CrossRef]
- Kaur, S.; Mondal, P. Study of total phenolic and flavonoid content, antioxidant activity and antimicrobial properties of medicinal plants. J. Microbiol. Exp. 2014, 1, 00005. [Google Scholar] [CrossRef]
- Krüzselyi, D.; Móricz, Á.M.; Vetter, J. Comparison of different morphological mushroom parts based on the antioxidant activity. LWT 2020, 127, 109436. [Google Scholar] [CrossRef]
- Morales, D.; Piris, A.J.; Ruiz-Rodriguez, A.; Prodanov, M.; Soler-Rivas, C. Extraction of bioactive compounds against cardiovascular diseases from Lentinula edodes using a sequential extraction method. Biotechnol. Prog. 2018, 34, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Reis, F.S.; Martins, A.; Barros, L.; Ferreira, I.C. Antioxidant properties and phenolic profile of the most widely appreciated cultivated mushrooms: A comparative study between in vivo and in vitro samples. Food Chem. Toxicol. 2012, 50, 1201–1207. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| No. | Metabolite | Log Bed (μM, n = 29) | Sawdust Media (μM, n = 52) | x-Fold Change |
|---|---|---|---|---|
| Log Bed/Sawdust Media | ||||
| 1 | Acetate | 397.88 ± 693.76 | 675.71 ± 1225.33 | 0.59 |
| 2 | Formate | 6.44 ± 2.25 | 6.52 ± 2.03 | 0.99 |
| 3 | Citrate | 527.57 ± 211.06 | 392.08 ± 103.01 | 1.35 ** |
| 4 | Fumarate | 1024.32 ± 252.79 | 1112.18 ± 225.77 | 0.92 |
| 5 | Malate | 4782.12 ± 893.68 | 5052.55 ± 886.63 | 0.95 |
| 6 | Succinate | 365.91 ± 181.96 | 300.12 ± 162.8 | 1.22 |
| 7 | Alanine | 1208.7 ± 487.68 | 1174.01 ± 467.17 | 1.03 |
| 8 | Arginine | 442.31 ± 226.7 | 641.1 ± 267.12 | 0.69 ** |
| 9 | Asparagine | 499.69 ± 176.29 | 588.61 ± 187.99 | 0.85 * |
| 10 | Aspartate | 618.73 ± 434.3 | 707.73 ± 407.19 | 0.87 |
| 11 | Glutamate | 1005.86 ± 331.19 | 1267.12 ± 440.44 | 0.79 ** |
| 12 | Glutamine | 5447.25 ± 1941.88 | 6693.75 ± 2364.08 | 0.81 * |
| 13 | Histidine | 162.39 ± 56.05 | 197.05 ± 68.67 | 0.82 * |
| 14 | Isoleucine | 132.63 ± 51.58 | 225.35 ± 102.73 | 0.59 *** |
| 15 | Leucine | 189.6 ± 68.7 | 322.53 ± 139.72 | 0.59 *** |
| 16 | Lysine | 453.17 ± 146.7 | 729.61 ± 217.9 | 0.62 *** |
| 17 | Ornithine | 635.24 ± 280.67 | 1054.49 ± 411.71 | 0.60 *** |
| 18 | Phenylalanine | 88.08 ± 25.01 | 134.59 ± 43.22 | 0.65 *** |
| 19 | Threonine | 488.2 ± 132.49 | 655.21 ± 176.73 | 0.75 *** |
| 20 | Tryptophan | 35.15 ± 15.18 | 51.49 ± 18.8 | 0.68 *** |
| 21 | Tyrosine | 88.52 ± 30.03 | 173.57 ± 86.12 | 0.51 *** |
| 22 | Valine | 257.18 ± 89.27 | 442.15 ± 179.74 | 0.58 *** |
| 23 | Arabinitol | 11,832.42 ± 5501.65 | 11,989.78 ± 5200.2 | 0.99 |
| 24 | Mannitol | 18,828.69 ± 3851.1 | 26,495.17 ± 6170.15 | 0.71 *** |
| 25 | Trehalose | 10,132.53 ± 2783.68 | 8688.03 ± 3165.51 | 1.17 |
| 26 | Glucose | 463.2 ± 753.62 | 370.91 ± 757.63 | 1.25 |
| 27 | myo-Inositol | 208.54 ± 59.41 | 258.13 ± 149.75 | 0.81 |
| 28 | UDP-N-Acetylglucosamine | 63.28 ± 11.53 | 112.86 ± 23.95 | 0.56 *** |
| 29 | UDP-galactose | 19.8 ± 3.5 | 28.25 ± 5.5 | 0.70 *** |
| 30 | UDP-glucose | 63.16 ± 11.03 | 97.21 ± 16.49 | 0.65 *** |
| 31 | Ethanol | 186.21 ± 270.16 | 91.77 ± 148.89 | 2.03 * |
| 32 | NAD+ | 35.23 ± 5.38 | 46.46 ± 8.98 | 0.76 *** |
| 33 | Uracil | 16.65 ± 15.27 | 11.78 ± 12.73 | 1.41 |
| 34 | Choline | 504.11 ± 231.99 | 231.63 ± 138.76 | 2.18 *** |
| 35 | sn-Glycero-3-phosphocholine | 383.91 ± 125.94 | 816.89 ± 310.72 | 0.47 *** |
| 36 | Trimethylamine N-oxide | 183.94 ± 37.73 | 173.6 ± 60.96 | 1.06 |
| No. | Proposed Identity | Rt (min) | Ionization Mode | Precursor Ions (m/z) | Difference (ppm) | Molecular Formula | x-Fold Change | |
|---|---|---|---|---|---|---|---|---|
| Theoretical | Observed | Log Bed/Sawdust Media | ||||||
| 1 | 3,4-Dimethoxybenzoic acid | 3.69 | − | 181.0501 | 181.0502 | 0.64 | C9H10O4 | 1.24 *** |
| 2 | Apigenin | 6.8 | − | 269.045 | 269.0463 | 4.83 | C15H10O5 | 0.74 * |
| 3 | Benzoic acid | 0.75 | − | 121.029 | 121.0284 | 4.59 | C7H6O2 | 3.17 *** |
| 4 | Caffeic acid | 3.86 | − | 179.0344 | 179.0344 | 0.2 | C9H8O4 | 0.33 |
| 5 | Catechin | 3.27 | − | 289.0712 | 289.0718 | 2.02 | C15H14O6 | 7.2 ** |
| 6 | Cinnamic acid | 5.72 | − | 147.0446 | 147.0446 | 0.03 | C9H8O2 | 1.31 *** |
| 7 | Coumaric acid | 4.35 | + | 165.0552 | 165.0538 | 8.3 | C9H8O3 | 0.15 *** |
| 8 | Ferulic acid | 4.59 | − | 193.0501 | 193.0517 | 8.37 | C10H10O4 | 0.40 * |
| 9 | Gallic acid | 2.49 | − | 169.0137 | 169.012 | 10.06 | C7H6O5 | 1.06 *** |
| 10 | Glycitein | 5.58 | − | 283.0607 | 283.0608 | 0.53 | C16H12O5 | 0.09 *** |
| 11 | L-ascorbic acid | 0.81 | − | 175.0243 | 175.0225 | 10.08 | C6H8O6 | 0.23*** |
| 12 | Niacinamide | 2 | + | 123.0558 | 123.0546 | 10.06 | C6H6N2O | 0.14 * |
| 13 | Riboflavin | 3.85 | +,− | 377.1461 | 377.1457 | 1.09 | C17H20N4O6 | 4.28 *** |
| 14 | Salicylic acid | 4.5 | − | 137.0239 | 137.0228 | 7.81 | C7H6O3 | 2.92 |
| 15 | Shikimic acid | 0.8 | − | 173.045 | 173.0433 | 9.82 | C7H10O5 | 1.10 *** |
| 16 | Syringic acid | 2.57 | − | 197.045 | 197.0449 | 0.51 | C9H10O5 | 1.34 *** |
| 17 | Vanillic acid | 3.44 | − | 167.0344 | 167.0349 | 2.78 | C8H8O4 | 0.34 ** |
| Bioactive Compounds | L. edodes | ||
|---|---|---|---|
| Log Bed (n = 29) | Sawdust Media (n = 52) | p Value | |
| Extraction yield (%) | 24.94 ± 4.48 | 25.08 ± 3.36 | 0.420 ns |
| Total phenolic content (mg GAE/g dry extract) | 5.06 ± 0.62 | 4.86 ± 0.68 | 0.017 * |
| Total flavonoid content (mg RE/g dry extract) | 1.35 ± 0.43 | 1.20 ± 0.53 | 0.000 *** |
| Total tannin content (mg CE/g dry extract) | 0.62 ± 0.22 | 0.62 ± 0.26 | 0.583 ns |
| Assays | L. edodes | ||
|---|---|---|---|
| Log Bed (n = 29) | Sawdust Media (n = 52) | p Value | |
| DPPH radical-scavenging activity | 0.88 ± 0.17 | 0.94 ± 0.17 | 0.012 * |
| ABTS radical-scavenging activity | 1.90 ± 0.42 | 2.38 ± 0.61 | 0.000 *** |
| Reducing power activity | 2.33 ± 0.58 | 2.92 ± 0.53 | 0.000 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, M.; Choi, J.Y.; Kim, M.-S. Metabolic Profiles, Bioactive Compounds, and Antioxidant Capacity in Lentinula edodes Cultivated on Log versus Sawdust Substrates. Biomolecules 2021, 11, 1654. https://doi.org/10.3390/biom11111654
Nam M, Choi JY, Kim M-S. Metabolic Profiles, Bioactive Compounds, and Antioxidant Capacity in Lentinula edodes Cultivated on Log versus Sawdust Substrates. Biomolecules. 2021; 11(11):1654. https://doi.org/10.3390/biom11111654
Chicago/Turabian StyleNam, Miso, Ji Yeon Choi, and Min-Sun Kim. 2021. "Metabolic Profiles, Bioactive Compounds, and Antioxidant Capacity in Lentinula edodes Cultivated on Log versus Sawdust Substrates" Biomolecules 11, no. 11: 1654. https://doi.org/10.3390/biom11111654
APA StyleNam, M., Choi, J. Y., & Kim, M.-S. (2021). Metabolic Profiles, Bioactive Compounds, and Antioxidant Capacity in Lentinula edodes Cultivated on Log versus Sawdust Substrates. Biomolecules, 11(11), 1654. https://doi.org/10.3390/biom11111654