
Depside and Depsidone Synthesis in Lichenized Fungi Comes into Focus through a Genome-Wide Comparison of the Olivetoric Acid and Physodic Acid Chemotypes of Pseudevernia furfuracea


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Chemotypes
2.2. DNA Extraction and Genome Sequencing
2.3. Genome Assembly and Annotation
2.4. Identification and Annotations of Biosynthetic Gene Clusters
2.5. Identification of Homologous BGCs
2.6. Phylogenetic Analyses
2.7. Candidate Cluster for Olivetoric Acid and Physodic Acid Biosynthesis
2.8. Identification of HexA and HexB
2.9. Metatranscriptome Analyses and Quantification of PKS, CytP450 and HexA and HexB Transcripts
3. Results
3.1. Genomes of the P. furfuracea Chemotypes
3.2. Predicted BGCs
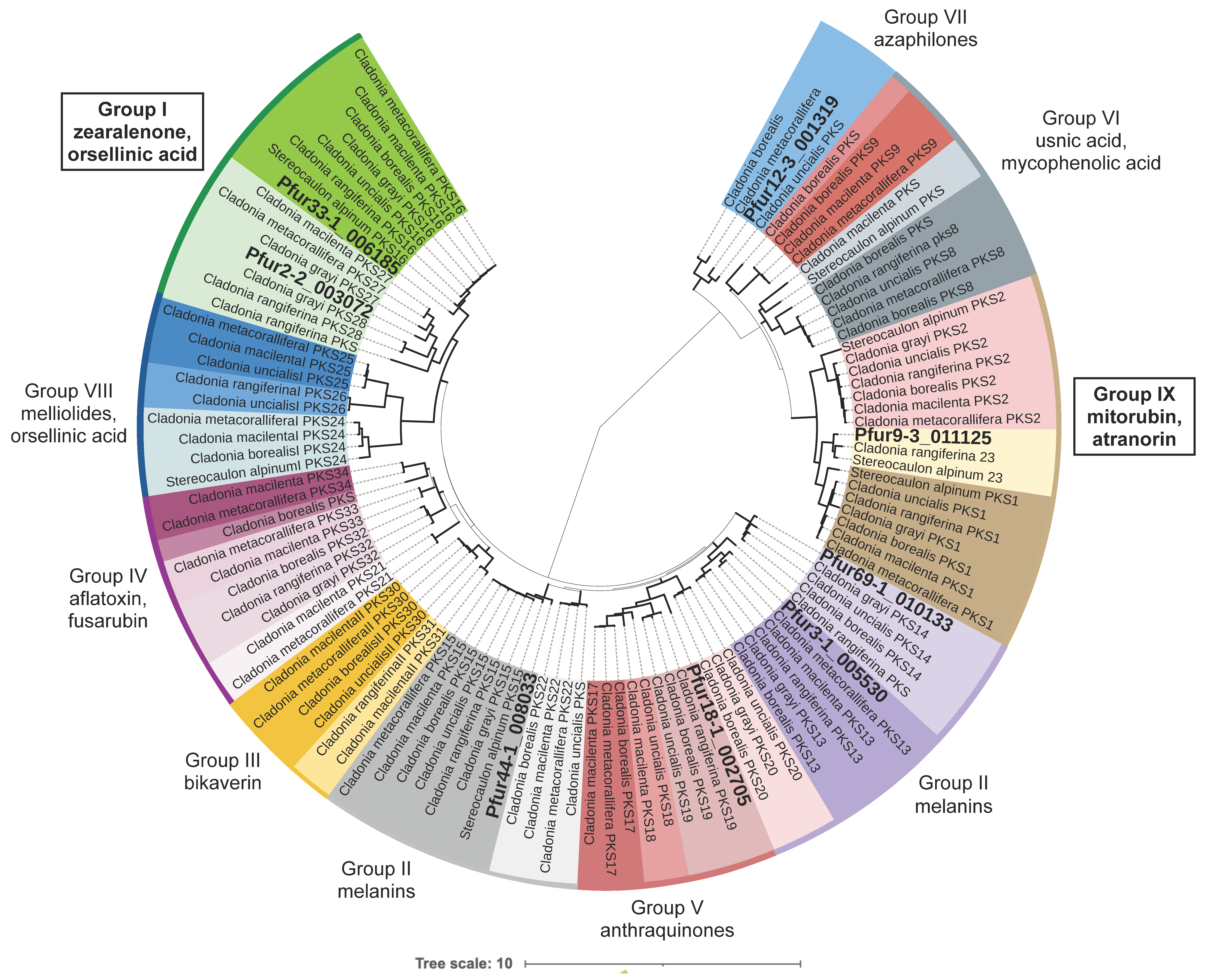
3.3. Phylogenetic Analyses
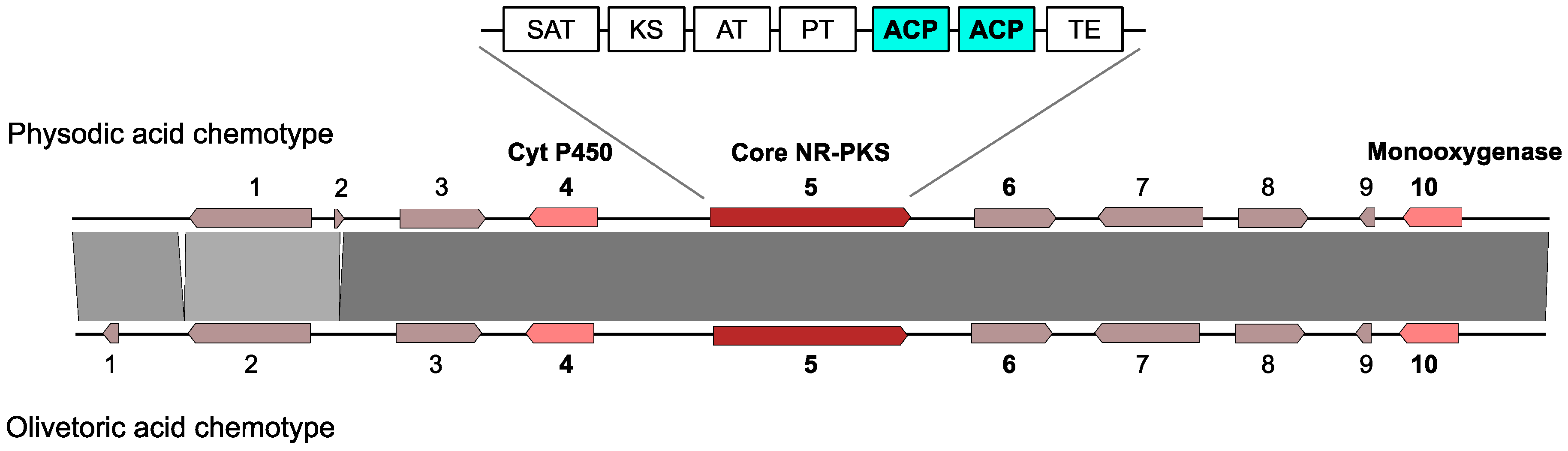
3.4. Selection of the Candidate Cluster for Olivetoric Acid and Physodic Acid
3.5. A Putative Atranorin Cluster Is Present in P. furfuracea
3.6. The Two Genes for a Metabolite FAS Are Present in P. furfuracea
3.7. Transcription of the Olivetoric Acid and Physodic Acid Cluster and of HexA and HexB
4. Discussion
4.1. True Intraspecific Variation Underlies Differences in BGCs between Chemotypes
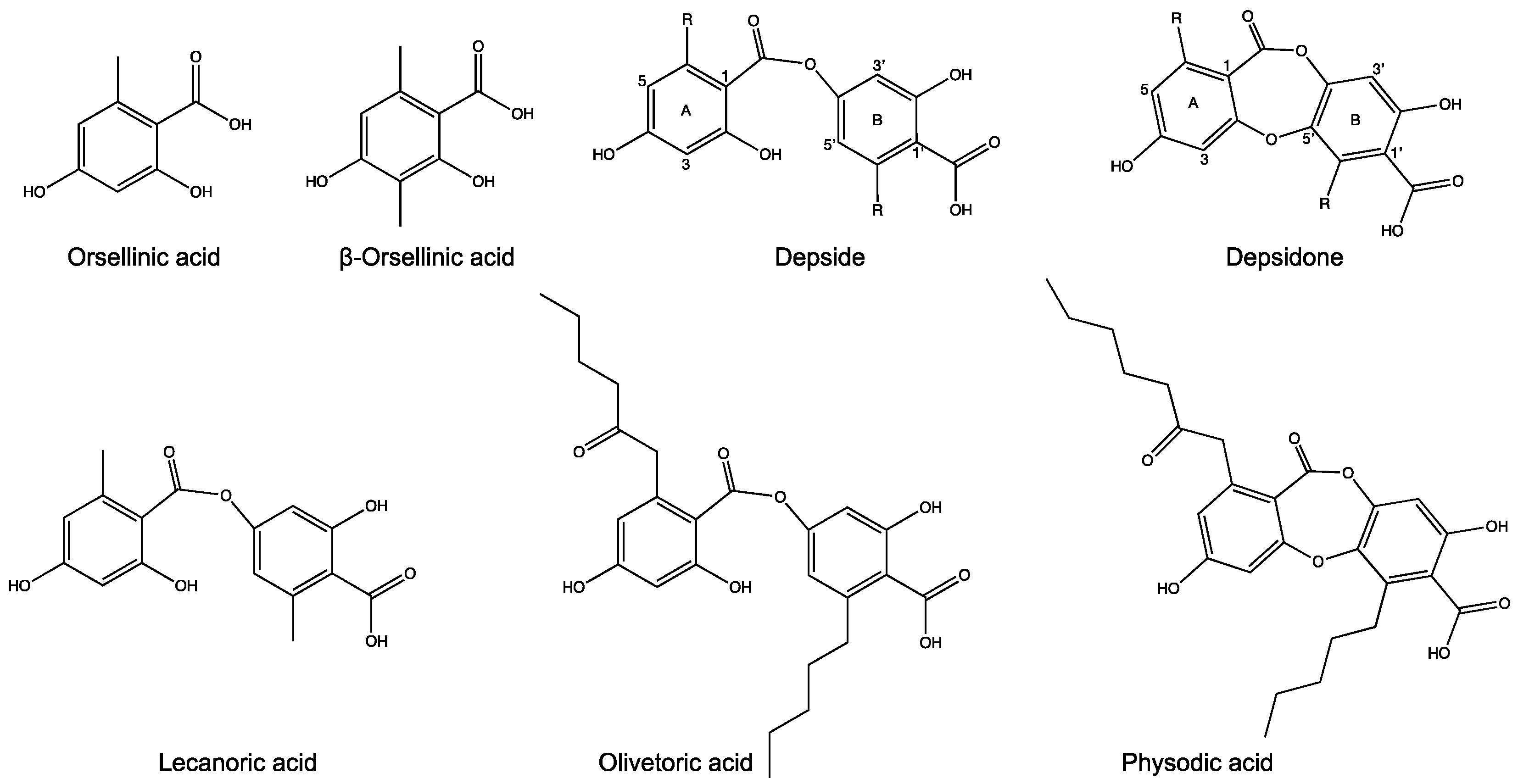
4.2. The Same Candidate BGC Is Linked to Depside and Depsidone Biosynthesis
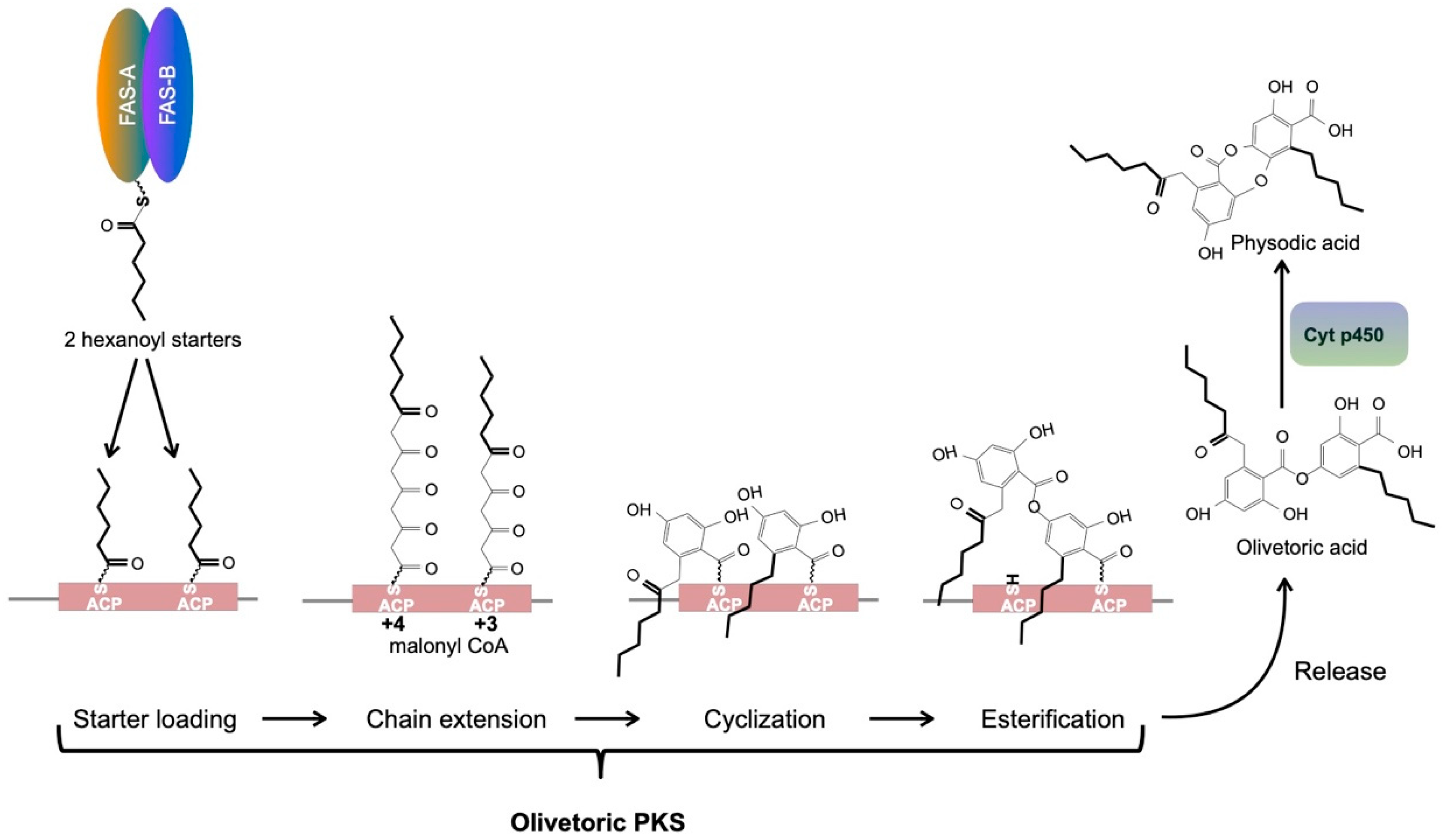
4.3. A Metabolite Fatty Acid Synthase Is the Likely Provider of the Hexanoyl Starter for Olivetoric Acid Synthesis
4.4. An Updated Scheme of Orcinol Depside and Depsidone Synthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shukla, V.; Joshi, G.P.; Rawat, M.S.M. Lichens as a potential natural source of bioactive compounds: A review. Phytochem. Rev. 2010, 9, 303–314. [Google Scholar] [CrossRef]
- Shrestha, G.; St. Clair, L.L. Lichens: A promising source of antibiotic and anticancer drugs. Phytochem. Rev. 2013, 12, 229–244. [Google Scholar] [CrossRef]
- Ingelfinger, R.; Henke, M.; Roser, L.; Ulshöfer, T.; Calchera, A.; Singh, G.; Parnham, M.J.; Geisslinger, G.; Fürst, R.; Schmitt, I.; et al. Unraveling the pharmacological potential of lichen extracts in the context of cancer and inflammation with a broad screening approach. Front. Pharmacol. 2020, 11, 1322. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, T.R. A theory of biogenesis of lichen depsides and depsidones. Proc. Indian Acad. Sci. 1944, 20A, 1–14. [Google Scholar] [CrossRef]
- Elix, J.; Jenie, U.; Parker, J. A novel synthesis of the lichen depsidones divaronic acid and stenosporonic acid, and the biosynthetic implications. Aust. J. Chem. 1987, 40, 1451–1464. [Google Scholar] [CrossRef]
- Calchera, A.; Dal Grande, F.; Bode, H.B.; Schmitt, I. Biosynthetic gene content of the “perfume lichens” Evernia prunastri and Pseudevernia furfuracea. Molecules 2019, 24, 203. [Google Scholar] [CrossRef] [Green Version]
- Meiser, A.; Otte, J.; Schmitt, I.; Grande, F.D. Sequencing genomes from mixed DNA samples - Evaluating the metagenome skimming approach in lichenized fungi. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Kroken, S.; Glass, N.L.; Taylor, J.W.; Yoder, O.C.; Turgeon, B.G. Phylogenomic analysis of type I polyketide synthase genes in pathogenic and saprobic ascomycetes. Proc. Natl. Acad. Sci. USA 2003, 100, 15670–15675. [Google Scholar] [CrossRef] [Green Version]
- Cox, R.J.; Simpson, T.J. Fungal type I polyketide synthases. Methods Enzymol. 2009, 459, 49–78. [Google Scholar]
- Armaleo, D.; Sun, X.; Culberson, C. Insights from the first putative biosynthetic gene cluster for a lichen depside and depsidone. Mycologia 2011, 103, 741–754. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.; Liu, R.; Woo, S.; Kang, K.B.; Park, H.; Yu, Y.H.; Ha, H.-H.; Oh, S.-Y.; Yang, J.H.; Kim, H.; et al. Linking a gene cluster to atranorin, a major cortical substance of lichens, through genetic dereplication and heterologous expression. MBio 2021, e0111121. [Google Scholar]
- Feng, C.; Wei, Q.; Hu, C.; Zou, Y. Biosynthesis of diphenyl ethers in fungi. Org. Lett. 2019, 21, 3114–3118. [Google Scholar] [CrossRef]
- Hitchman, T.S.; Schmidt, E.W.; Trail, F.; Rarick, M.D.; Linz, J.E.; Townsend, C.A. Hexanoate synthase, a specialized type I fatty acid synthase in aflatoxin B1 biosynthesis. Bioorg. Chem. 2001, 29, 293–307. [Google Scholar] [CrossRef]
- Smith, S.; Tsai, S.C. The type I fatty acid and polyketide synthases: A tale of two megasynthases. Nat. Prod. Rep. 2007, 24, 1041–1072. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, C.M.H.; Townsend, C.A. Initial characterization of a type I fatty acid synthase and polyketide synthase multienzyme complex NorS in the biosynthesis of aflatoxin B1. Chem. Biol. 2002, 9, 981–988. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.W.; Adams, T.H.; Keller, N.P. Aspergillus has distinct fatty acid synthases for primary and secondary metabolism. Proc. Natl. Acad. Sci. USA 1996, 93, 14873–14877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Culberson, C.F.; Culberson, W.L. Chemosyndromic variation in lichens. Syst. Bot. 1976, 1, 325. [Google Scholar] [CrossRef]
- Kealey, J.T.; Craig, J.P.; Barr, P.J. Identification of a lichen depside polyketide synthase gene by heterologous expression in Saccharomyces cerevisiae. Metab. Eng. Commun. 2021, e00172. [Google Scholar] [CrossRef] [PubMed]
- Halvorsen, R.; Bendiksen, E. The chemical variation of Pseudevernia furfuracea in Norway. Nord. J. Bot. 1982, 2, 371–380. [Google Scholar] [CrossRef]
- Culberson, W.L.; Culberson, C.F.; Johnson, A. Pseudevernia furfuracea-olivetorina relationships: Chemistry and ecology. Mycologia 1977, 69, 604. [Google Scholar] [CrossRef]
- Kosanić, M.; Manojlović, N.; Janković, S.; Stanojković, T.; Ranković, B. Evernia prunastri and Pseudoevernia furfuraceae lichens and their major metabolites as antioxidant, antimicrobial and anticancer agents. Food Chem. Toxicol. 2013, 53, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Feige, G.B.; Lumbsch, H.T.; Huneck, S.; Elix, J.A. Identification of lichen substances by a standardized high-performance liquid chromatographic method. J. Chromatogr. A 1993, 646, 417–427. [Google Scholar] [CrossRef]
- Benatti, M.N.; Gernert, M.; Schmitt, I. Parmotrema hydrium, a new species of Parmeliaceae in southeastern Brazil. Acta Bot. Brasilica 2013, 27, 810–814. [Google Scholar] [CrossRef] [Green Version]
- Cubero, O.F.; Crespo, A. Isolation of nucleic acids from lichens. In Protocols in Lichenology; Kranner, I., Beckett, R., Varma, A., Eds.; Springer: Berlin, Germany, 2002; pp. 381–391. [Google Scholar]
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef]
- Boetzer, M.; Pirovano, W. SSPACE-LongRead: Scaffolding bacterial draft genomes using long read sequence information. BMC Bioinform. 2014, 15, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huson, D.H.; Beier, S.; Flade, I.; Górska, A.; El-Hadidi, M.; Mitra, S.; Ruscheweyh, H.-J.; Tappu, R. MEGAN Community Edition - Interactive exploration and analysis of large-scale microbiome sequencing data. PLoS Comput. Biol. 2016, 12, e1004957. [Google Scholar] [CrossRef] [Green Version]
- Bradnam, K.R.; Fass, J.N.; Alexandrov, A.; Baranay, P.; Bechner, M.; Birol, I.; Boisvert, S.; Chapman, J.A.; Chapuis, G.; Chikhi, R.; et al. Assemblathon 2: Evaluating de novo methods of genome assembly in three vertebrate species. Gigascience 2013, 2, 10. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.; Stajich, J. Funannotate v1.7.4. Zenodo 2019. [Google Scholar]
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, W339–W346. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Borodovsky, M.; Lomsadze, A. Eukaryotic gene prediction using GeneMark.hmm-E and GeneMark-ES. Curr. Protoc. Bioinform. 2011, 35, 4.6.1–4.6.10. [Google Scholar] [CrossRef] [Green Version]
- Quevillon, E.; Silventoinen, V.; Pillai, S.; Harte, N.; Mulder, N.; Apweiler, R.; Lopez, R. InterProScan: Protein domains identifier. Nucleic Acids Res. 2005, 33, W116. [Google Scholar] [CrossRef] [Green Version]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. EggNOG 5.0: A hierarchical, functionally and phylogenetically annotated orthology resource based on 5090 organisms and 2502 viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huerta-Cepas, J.; Forslund, K.; Coelho, L.P.; Szklarczyk, D.; Jensen, L.J.; Von Mering, C.; Bork, P. Fast genome-wide functional annotation through orthology assignment by eggNOG-mapper. Mol. Biol. Evol. 2017, 34, 2115–2122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sønderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar] [CrossRef]
- Käll, L.; Krogh, A.; Sonnhammer, E.L.L. Advantages of combined transmembrane topology and signal peptide prediction-the Phobius web server. Nucleic Acids Res. 2007, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kautsar, S.A.; Blin, K.; Shaw, S.; Navarro-Muñoz, J.C.; Terlouw, B.R.; Van Der Hooft, J.J.J.; Van Santen, J.A.; Tracanna, V.; Suarez Duran, H.G.; Pascal Andreu, V.; et al. MIBiG 2.0: A repository for biosynthetic gene clusters of known function. Nucleic Acids Res. 2020, 48, D454–D458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullivan, M.J.; Petty, N.K.; Beatson, S.A. Easyfig: A genome comparison visualizer. Bioinformatics 2011, 27, 1009–1010. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Le, S.; Li, Y.; Hu, F. SeqKit: A cross-platform and ultrafast toolkit for FASTA/Q file manipulation. PLoS ONE 2016, 11, e0163962. [Google Scholar] [CrossRef]
- Rice, P.; Longden, L.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Kjærbølling, I.; Vesth, T.C.; Frisvad, J.C.; Nybo, J.L.; Theobald, S.; Kuo, A.; Bowyer, P.; Matsuda, Y.; Mondo, S.; Lyhne, E.K.; et al. Linking secondary metabolites to gene clusters through genome sequencing of six diverse Aspergillus species. Proc. Natl. Acad. Sci. USA 2018, 115, E753–E761. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lünne, F.; Niehaus, E.M.; Lipinski, S.; Kunigkeit, J.; Kalinina, S.A.; Humpf, H.U. Identification of the polyketide synthase PKS7 responsible for the production of lecanoric acid and ethyl lecanorate in Claviceps purpurea. Fungal Genet. Biol. 2020, 145, 103481. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, D.; Divakar, P.K.; Grewe, F.; Crespo, A.; Dal Grande, F.; Lumbsch, H.T. Genome-wide analysis of biosynthetic gene cluster reveals correlated gene loss with absence of usnic acid in lichen-forming fungi. Genome Biol. Evol. 2020, 12, 1858–1868. [Google Scholar] [CrossRef]
- Rubio-Piña, J.A.; Zapata-Pérez, O. Isolation of total RNA from tissues rich in polyphenols and polysaccharides of mangrove plants. Electron. J. Biotechnol. 2011, 14. [Google Scholar]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Pertea, G.; Trapnell, C.; Pimentel, H.; Kelley, R.; Salzberg, S.L. TopHat2: Accurate alignment of transcriptomes in the presence of insertions, deletions and gene fusions. Genome Biol. 2013, 14, R36. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Dillies, M.A.; Rau, A.; Aubert, J.; Hennequet-Antier, C.; Jeanmougin, M.; Servant, N.; Keime, C.; Marot, N.S.; Castel, D.; Estelle, J.; et al. A comprehensive evaluation of normalization methods for Illumina high-throughput RNA sequencing data analysis. Brief. Bioinform. 2013, 14, 671–683. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef] [Green Version]
- Oshlack, A.; Wakefield, M.J. Transcript length bias in RNA-seq data confounds systems biology. Biol. Direct 2009, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Drott, M.T.; Bastos, R.W.; Rokas, A.; Ries, L.N.A.; Gabaldón, T.; Goldman, G.H.; Keller, N.P.; Greco, C. Diversity of secondary metabolism in Aspergillus nidulans clinical isolates. mSphere 2020, 5, e00156-20. [Google Scholar] [CrossRef] [Green Version]
- Moore, B.D.; Andrew, R.L.; Külheim, C.; Foley, W.J. Explaining intraspecific diversity in plant secondary metabolites in an ecological context. New Phytol. 2014, 201, 733–750. [Google Scholar] [CrossRef]
- Zhu, B.; Ibrahim, M.; Cui, Z.; Xie, G.; Jin, G.; Kube, M.; Li, B.; Zhou, X. Multi-omics analysis of niche specificity provides new insights into ecological adaptation in bacteria. ISME J. 2016, 10, 2072–2075. [Google Scholar] [CrossRef] [Green Version]
- Thynne, E.; Mead, O.L.; Chooi, Y.-H.; McDonald, M.C.; Solomon, P.S. Acquisition and loss of secondary metabolites shaped the evolutionary path of three emerging phytopathogens of wheat. Genome Biol. Evol. 2019, 11, 890–905. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Calchera, A.; Schulz, M.; Drechsler, M.; Bode, H.B.; Schmitt, I.; Dal Grande, F. Climate-specific biosynthetic gene clusters in populations of a lichen-forming fungus. Environ. Microbiol. 2021, 23, 4260–4275. [Google Scholar] [CrossRef]
- Wisecaver, J.H.; Slot, J.C.; Rokas, A. The Evolution of Fungal Metabolic Pathways. PLoS Genet. 2014, 10, e1004816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lind, A.L.; Wisecaver, J.H.; Lameiras, C.; Wiemann, P.; Palmer, J.M.; Keller, N.P.; Rodrigues, F.; Goldman, G.H.; Rokas, A. Drivers of genetic diversity in secondary metabolic gene clusters within a fungal species. PLoS Biol. 2017, 15, e2003583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rokas, A.; Wisecaver, J.H.; Lind, A.L. The birth, evolution and death of metabolic gene clusters in fungi. Nat. Rev. Microbiol. 2018, 16, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Vicente, C.M.; Thibessard, A.; Lorenzi, J.N.; Benhadj, M.; Hôtel, L.; Gacemi-Kirane, D.; Lespinet, O.; Leblond, P.; Aigle, B. Comparative genomics among closely related Streptomyces strains revealed specialized metabolite biosynthetic gene cluster diversity. Antibiotics 2018, 7, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macheleidt, J.; Mattern, D.J.; Fischer, J.; Netzker, T.; Weber, J.; Schroeckh, V.; Valiante, V.; Brakhage, A.A. Regulation and role of fungal secondary metabolites. Annu. Rev. Genet. 2016, 50, 371–392. [Google Scholar] [CrossRef]
- Keller, N.P. Fungal secondary metabolism: Regulation, function and drug discovery. Nat. Rev. Microbiol. 2019, 17, 167–180. [Google Scholar] [CrossRef]
- Fox, E.M.; Howlett, B.J. Secondary metabolism: Regulation and role in fungal biology. Curr. Opin. Microbiol. 2008, 11, 481–487. [Google Scholar] [CrossRef]
- Patra, B.; Schluttenhofer, C.; Wu, Y.; Pattanaik, S.; Yuan, L. Transcriptional regulation of secondary metabolite biosynthesis in plants. Biochim. Biophys. Acta Gene Regul. Mech. 2013, 1829, 1236–1247. [Google Scholar] [CrossRef]
- Collemare, J.; Seidl, M.F. Chromatin-dependent regulation of secondary metabolite biosynthesis in fungi: Is the picture complete? FEMS Microbiol. Rev. 2019, 43, 591–607. [Google Scholar] [CrossRef] [Green Version]
- Yin, W.; Keller, N.P. Transcriptional regulatory elements in fungal secondary metabolism. J. Microbiol. 2011, 49, 329–339. [Google Scholar] [CrossRef] [Green Version]
- Wasil, Z.; Pahirulzaman, K.A.K.; Butts, C.; Simpson, T.J.; Lazarus, C.M.; Cox, R.J. One pathway, many compounds: Heterologous expression of a fungal biosynthetic pathway reveals its intrinsic potential for diversity. Chem. Sci. 2013, 4, 3845–3856. [Google Scholar] [CrossRef] [Green Version]
- Jenni, S.; Leibundgut, M.; Maier, T.; Ban, N. Architecture of a fungal fatty acid synthase at 5 Å resolution. Science (80-.) 2006, 311, 1263–1267. [Google Scholar] [CrossRef]
- Watanabe, C.M.H.; Wilson, D.; Linz, J.E.; Townsend, C.A. Demonstration of the catalytic roles and evidence for the physical association of type I fatty acid synthases and a polyketide synthase in the biosynthesis of aflatoxin B1. Chem. Biol. 1996, 3, 463–469. [Google Scholar] [CrossRef] [Green Version]
- Gluck-Thaler, E.; Haridas, S.; Binder, M.; Grigoriev, I.V.; Crous, P.W.; Spatafora, J.W.; Bushley, K.; Slot, J.C. The architecture of metabolism maximizes biosynthetic diversity in the largest class of fungi. Mol. Biol. Evol. 2020, 37, 2838–2856. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physodic Acid Chemotype | Olivetoric Acid Chemotype | ||
|---|---|---|---|
| Genome accession nos. | JAIUPS000000000 | JAIUPT000000000 | |
| Sequencing Report | Subreads bases (Gb) | 8.182 | 8.122 |
| Average subreads length | 9563 | 9325 | |
| N50 raw reads | 12855 | 12479 | |
| Assembly Stats | # scaffolds | 104 | 53 |
| CDS | 11199 | 10480 | |
| size (Mb) | 34.2 | 37.3 | |
| N50 | 632 kb | 1.6 Mb | |
| % completeness | 96 | 92 | |
| homologous clusters | 51 | ||
| chemotype specific clusters | 5 | 1 | |
| BGCs Summary | R-PKSs | 19 | 14 |
| NR-PKSs | 4 | 4 | |
| hybrid | 10 | 8 | |
| T3-PKS | 1 | 1 | |
| NRPS | 4 | 4 | |
| NRPS-like | 10 | 12 | |
| Terpenes | 6 | 6 | |
| indole | 2 | 2 | |
| Clusters Detected in Only One Chemotype | |||||
| Cluster Number | Name of the Cluster | Detected in | Core Gene | Normalized Read Count in Physodic Acid Chemotype | Normalized Read Count in Olivetoric Acid Chemotype |
| 52 | Region 7.2 | physodic acid chemotype | R-PKS | 24.66 | 0 |
| 53 | Region 25.1 | physodic acid chemotype | R-PKS | 11.50 | 0 |
| 54 | Region 60.2 | physodic acid chemotype | hybrid | 24.44 | 0 |
| 55 | Region 65.1 | physodic acid chemotype | R-PKS | 18.30 | 0 |
| 56 | Region 27.1 | physodic acid chemotype | terpene | 59.99 | 0 |
| 57 | Region 10.1 | olivetoric acid chemotype | terpene | 0 | 133.94 |
| Clusters present in both chemotypes | |||||
| 1 | Region 12.3 | both chemotypes | NR-PKS | 18.19 | 47.66 |
| 2 | Region 18.1 | both chemotypes | NR-PKS | 16.43 | 56.19 |
| 3 | Region 44.1 | both chemotypes | NR-PKS | 29.05 | 43.41 |
| 4 | Region 33.1 | both chemotypes | NR-PKS | 37.08 | 42.14 |
| 9 | Region 10.1 | both chemotypes | R-PKS | 20.14 | 45.88 |
| 11 | Region 16.1 | both chemotypes | R-PKS | 25.53 | 48.84 |
| 12 | Region 2.4 | both chemotypes | R-PKS | 33.33 | 42.85 |
| 15 | Region 9.2 | both chemotypes | R-PKS | 35.00 | 39.86 |
| 6 | Region 9.3 | both chemotypes | R-PKS | 23.87 | 41.47 |
| 5 | Region 2.2 | both chemotypes | R-PKS | 26.24 | 53.15 |
| 44 | Region 38.1 | both chemotypes | terpene | 40.34 | 107.15 |
| 45 | Region 33.2 | both chemotypes | terpene | 83.56 | 119.52 |
| 46 | Region 2.3 | both chemotypes | terpene | 37.49 | 114.33 |
| 47 | Region 12.2 | both chemotypes | terpene | 50.63 | 149.93 |
| 48 | Region 60.1 | both chemotypes | terpene | 51.24 | 85.36 |
| Cluster Number | PKS | Compound Defining the Group (Kim et al. [11]) | PKS Category (Kim et al. [11]) | PKS Domains | Total Genes in the Cluster | Identified Genes in Cluster | Most Similar Known PKS (MIBiG) | Coverage and Similarity to MIBiG Cluster |
| 1 | Pfur12-3_001319 | VII (azaphilones, monascorubrin) | not known | SAT-KS-AT-ACP cMT-TD | 12 | Regulatory gene, NRPKS | monascorubrin | 95%, 58% |
| 2 | Pfur18-1_002705 | V (anthraquinones) | PKS20 | SAT-KS-AT-PT-ACP | 16 | metallo-beta-lactamase family protein, NRPKS, halogenase | RES-1214-2 | 61%, 100% |
| 3 | Pfur44-1_008033 | II (melanins) | PKS15 | SAT-KS-AT-PT-ACP ACP-TE | 11 | PKS, dehydrogenase/reductase (KR) | naphthalene | 99.5%, 53% |
| 4 | Pfur33-1_006185 | I (zearalenone, orsellinic acid) | PKS16 | SAT-KS-AT-ACP ACP-TE | 10 | CytP450, PKS, monooxygenase | grayanic acid | 99%, 73% |
| 5 | Pfur2-2_003072 | I (zearalenone, orsellinic acid) | PKS27 | SAT-KS-AT-PT-ACP-TE | 13 | O-methyltransferase, CytP450, crotonyl-CoA reductase/alcohol dehydrogenase, red-PKS, NRPKS, GATase_7 | grayanic acid | 100%, 38% |
| 6 | Pfur9-3_011125 | IX (mitorubin, atranorin) | PKS23 | SAT-KS-AT-PT-ACP cMT | 11 | alkyl hydroperoxide reductase/Thiol specific, PKS, CytP450, drug resistance transporter | ascochlorin cluster (Acremonium egyptiacum) | 100%, 38% |
| 7 | Pfur3-1_005530 | II (melanins) | PKS13 | SAT-KS-AT-PT-ACP-ACP-TE | 13 | serine/threonine protein kinase, Drug resistance transporter, monooxygenase FAD-binding, NRPKS, short-chain dehydrogenase/reductase SDR, O-methyltransferase, transcription regulator | 100%, 48% | |
| 8 | Pfur69-1_010133 | II (melanins) | PKS14 | SAT-KS-AT-PT-ACP-ACP-TE | 9 | Drug resistance transporter, O-methyltransferase, halogenase, monooxygenase FAD-binding, adh_short (DH-KR), NRPKS, monooxygenase FAD-binding | 6-hydroxymellein (Cladonia uncialis) | 99%, 78% |
| Physodic Acid Chemotype | Olivetoric Acid Chemotype | |||||
|---|---|---|---|---|---|---|
| Gene | Gene Identity | Gene Length | Number of Raw Reads Aligned | Reads (Normalized by CPM) | # Raw Reads Aligned | Reads (Normalized by CPM) |
| cluster 4 gene1 | unidentified | 3822 | 38 | 0 | no hits | - |
| cluster 4 gene2 | unidentified | 306 | no hits | - | 1 | 0 |
| cluster 4 gene3 | unidentified | 2685 | no hits | - | 19 | 0 |
| cluster 4 gene4 | CytP450 | 2139 | 8677 | 117.29 | 1222 | 17.4 |
| cluster 4 gene5 | NR-PKS | 6294 | 111226 | 511 | 1265 | 6.33 |
| cluster 4 gene6 | unidentified | 2550 | 13690 | 155.25 | 6304 | 75.38 |
| cluster 4 gene7 | unidentified | 3284 | 1035 | 9.11 | 118 | 1.125 |
| cluster 4 gene8 | unidentified | 2191 | 936 | 12.35 | 205 | 1.98 |
| cluster 4 gene9 | unidentified | 492 | no hits | - | 12 | 0.73 |
| cluster 4 gene10 | monooxygenase | 1851 | 46 | 0.71 | 19 | 0.307 |
| FAS | FAS-A | 5619 | 3143 | 16.17 | 944 | 5.12 |
| FAS | FAS-B | 6285 | 1971 | 9.06 | 911 | 4.42 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, G.; Armaleo, D.; Dal Grande, F.; Schmitt, I. Depside and Depsidone Synthesis in Lichenized Fungi Comes into Focus through a Genome-Wide Comparison of the Olivetoric Acid and Physodic Acid Chemotypes of Pseudevernia furfuracea. Biomolecules 2021, 11, 1445. https://doi.org/10.3390/biom11101445
Singh G, Armaleo D, Dal Grande F, Schmitt I. Depside and Depsidone Synthesis in Lichenized Fungi Comes into Focus through a Genome-Wide Comparison of the Olivetoric Acid and Physodic Acid Chemotypes of Pseudevernia furfuracea. Biomolecules. 2021; 11(10):1445. https://doi.org/10.3390/biom11101445
Chicago/Turabian StyleSingh, Garima, Daniele Armaleo, Francesco Dal Grande, and Imke Schmitt. 2021. "Depside and Depsidone Synthesis in Lichenized Fungi Comes into Focus through a Genome-Wide Comparison of the Olivetoric Acid and Physodic Acid Chemotypes of Pseudevernia furfuracea" Biomolecules 11, no. 10: 1445. https://doi.org/10.3390/biom11101445
APA StyleSingh, G., Armaleo, D., Dal Grande, F., & Schmitt, I. (2021). Depside and Depsidone Synthesis in Lichenized Fungi Comes into Focus through a Genome-Wide Comparison of the Olivetoric Acid and Physodic Acid Chemotypes of Pseudevernia furfuracea. Biomolecules, 11(10), 1445. https://doi.org/10.3390/biom11101445