Immunomodulatory Activities of Selected Essential Oils


Abstract

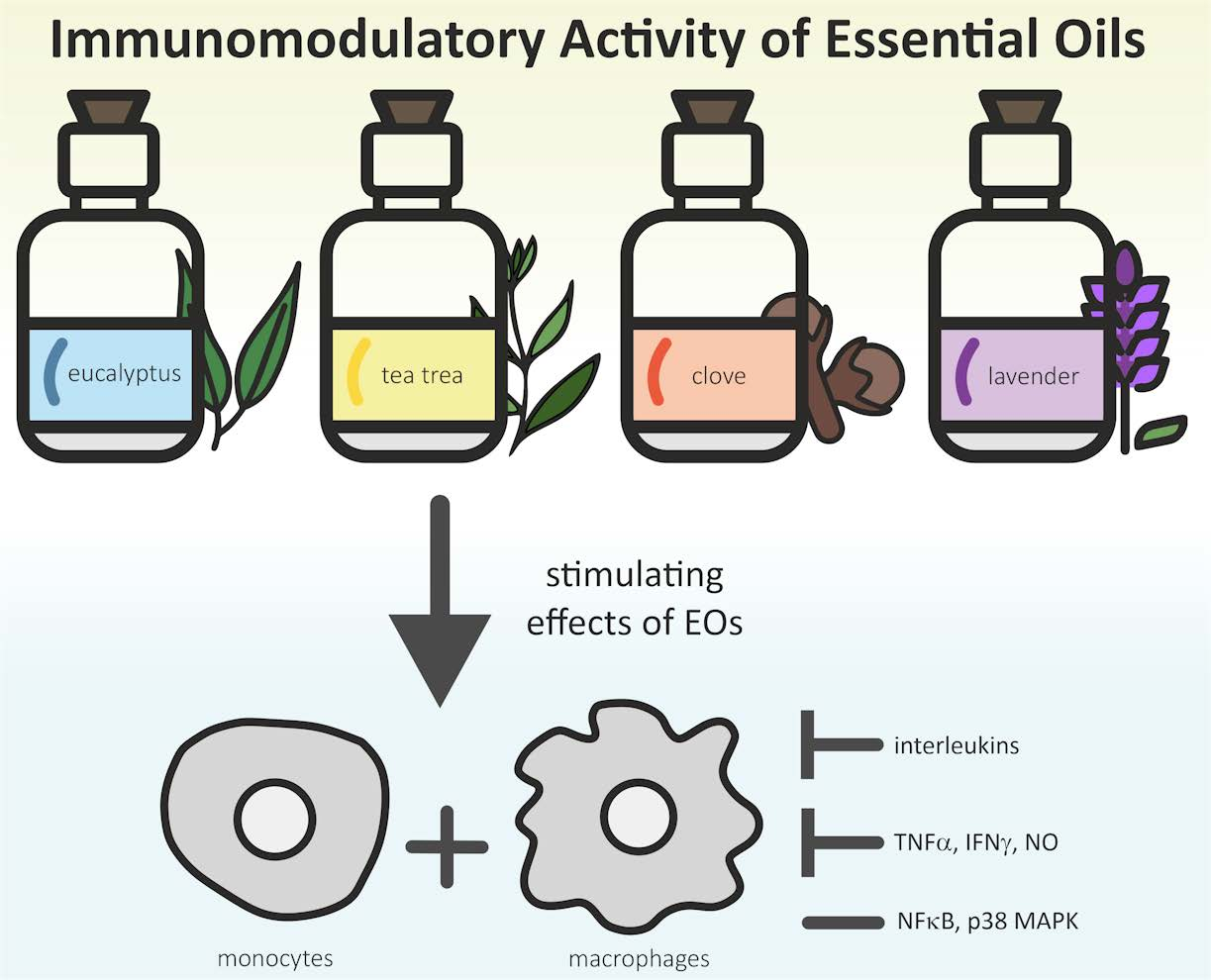
1. Introduction—The Immune System and Herbal Medicine
2. Methods
3. Eucalyptus Essential Oil
4. Clove Essential Oil
5. Tea Tree Essential Oil
6. Lavender Essential Oil
7. Conclusions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Explanation |
|---|---|
| APC | antigen-presenting cells |
| CEO | clove essential oil |
| COX-2 | cyclooxygenase |
| DTH | delayed-type hypersensitivity |
| EE | eugenol emulsion |
| EEO | eucalyptus essential oil |
| EO | essential oil |
| HO-1 | heme oxygenase-1 |
| IFNγ | interferon-γ |
| IL | interleukin |
| iNOS | inducible nitric oxide synthase |
| IκB | inhibitor of κB kinase |
| LEO | lavender essential oil |
| LPS | lipopolysaccharide |
| MAC | Melaleuca alternifolia concentrate |
| MDM | monocyte-derived macrophages |
| MUC5B | gel-forming mucin |
| NFκB | nuclear factor kappa-light-chain-enhancer of activated B-cells |
| NLR | nod-like receptor |
| NO | nitric oxide |
| Nrf2 | NF-E2-related factor 2 |
| p38 MAPK | p38-mitogen-activated protein kinase |
| PPAR | peroxisome proliferator-activated receptor |
| SRBC | sheep red blood cells |
| TNFα | tumor necrosis factor α |
| TREM-1 | triggering receptor expressed on myeloid cells |
| TTO | tea tree oil |
| VL | visceral leishmaniasis |
| WBC | total white blood cell count |
References
- Hotta, M.; Nakata, R.; Katsukawa, M.; Hori, K.; Takahashi, S.; Inoue, H. Carvacrol, a component of thyme oil, activates PPARα and γ and suppresses COX-2 expression. J. Lipid Res. 2009, 51, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.H.; Brand, C.; Carson, C.F.; Riley, T.V.; Prager, R.H.; Finlay-Jones, J.J. Terpinen-4-ol, the main component of the essential oil of Melaleuca alternifolia (tea tree oil), suppresses inflammatory mediator production by activated human monocytes. Inflamm. Res. 2000, 49, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, M.N.M.; Aquino, S.G.; Rossa Junior, C.; Spolidorio, D.M.P. Terpinen-4-ol and alpha-terpineol (tea tree oil components) inhibit the production of IL-1β, IL-6 and IL-10 on human macrophages. Inflamm. Res. 2014, 63, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 49. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, Z.; Huang, Z.; Zhao, M.; Li, P.; Zhou, W.; Zhang, K.; Zheng, X.; Lin, L.; Tang, J.; et al. Variation in Essential Oil and Bioactive Compounds of Curcuma kwangsiensis Collected from Natural Habitats. Chem. Biodivers. 2017, 14. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef]
- Fujimura, Y.; Kawano, C.; Maeda-Murayama, A.; Nakamura, A.; Koike-Miki, A.; Yukihira, D.; Hayakawa, E.; Ishii, T.; Tachibana, H.; Wariishi, H.; et al. A Chemometrics-driven Strategy for the Bioactivity Evaluation of Complex Multicomponent Systems and the Effective Selection of Bioactivity-predictive Chemical Combinations. Sci. Rep. 2017, 7, 2257. [Google Scholar] [CrossRef]
- Hammoda, H.M.; Harraz, F.M.; Farag, M.A.; El-Aswad, A.F.; El-Hawiet, A.; Eid, A.M. Volatiles profiling and bioactivities of Cupressus spp. leaf and cone essential oils as analyzed via chemometrics tools. J. Essent. Oil Res. 2019, 31, 53–62. [Google Scholar] [CrossRef]
- Sonnenberg, G.F.; Hepworth, M.R. Functional interactions between innate lymphoid cells and adaptive immunity. Nat. Rev. Immunol. 2019, 19, 599–613. [Google Scholar] [CrossRef]
- Redgrove, K.A.; McLaughlin, E.A. The Role of the Immune Response in Chlamydia trachomatis Infection of the Male Genital Tract: A Double-Edged Sword. Front. Immunol. 2014, 5, 534. [Google Scholar] [CrossRef] [PubMed]
- Ashley, N.T.; Weil, Z.M.; Nelson, R.J. Inflammation: Mechanisms, Costs, and Natural Variation. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 385–406. [Google Scholar] [CrossRef]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M.; et al. A guiding map for inflammation. Nat. Immunol. 2017, 18, 826–831. [Google Scholar] [CrossRef] [PubMed]
- Gea-Banacloche, J.C. Immunomodulation. In Principles of Molecular Medicine; Runge, M.S., Patterson, C., Eds.; Humana Press: Totowa, NJ, USA, 2006; pp. 893–904. [Google Scholar]
- Huang, C.-F.; Lin, S.-S.; Liao, P.-H.; Young, S.-C.; Yang, C.-C. The Immunopharmaceutical Effects and Mechanisms of Herb Medicine. Cell. Mol. Immunol. 2008, 5, 23–31. [Google Scholar] [CrossRef]
- Dong, L.; Liu, J.; Zhong, Z.; Wang, S.; Wang, H.; Huo, Y.; Wei, Z.; Yu, L. Dietary tea tree oil supplementation improves the intestinal mucosal immunity of weanling piglets. Anim. Feed Sci. Technol. 2019, 255, 114209. [Google Scholar] [CrossRef]
- Kang, S.; Min, H. Ginseng, the ‘Immunity Boost’: The Effects of Panax ginseng on Immune System. J. Ginseng Res. 2012, 36, 354–368. [Google Scholar] [CrossRef]
- Sandner, G.; Mueller, A.S.; Zhou, X.; Stadlbauer, V.; Schwarzinger, B.; Schwarzinger, C.; Wenzel, U.; Maenner, K.; van der Klis, J.D.; Hirtenlehner, S.; et al. Ginseng Extract Ameliorates the Negative Physiological Effects of Heat Stress by Supporting Heat Shock Response and Improving Intestinal Barrier Integrity: Evidence from Studies with Heat-Stressed Caco-2 Cells, C. elegans and Growing Broilers. Molecules 2020, 25, 835. [Google Scholar] [CrossRef]
- Reyes, A.W.B.; Hop, H.T.; Arayan, L.T.; Huy, T.X.N.; Park, S.J.; Kim, K.D.; Min, W.; Lee, H.J.; Rhee, M.H.; Kwak, Y.-S.; et al. The host immune enhancing agent Korean red ginseng oil successfully attenuates Brucella abortus infection in a murine model. J. Ethnopharmacol. 2017, 198, 5–14. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Quispe, C.; Llaique, H.; Villalobos, M.; Smeriglio, A.; Trombetta, D.; Ezzat, S.M.; Salem, M.A.; Zayed, A.; et al. Insights into Eucalyptus genus chemical constituents, biological activities and health-promoting effects. Trends Food Sci. Technol. 2019, 91, 609–624. [Google Scholar] [CrossRef]
- Barbosa, L.C.; Filomeno, C.A.; Teixeira, R.R. Chemical Variability and Biological Activities of Eucalyptus spp. Essential Oils. Molecules 2016, 21, 1671. [Google Scholar] [CrossRef]
- Yin, C.; Liu, B.; Wang, P.; Li, X.; Li, Y.; Zheng, X.; Tai, Y.; Wang, C.; Liu, B. Eucalyptol alleviates inflammation and pain responses in a mouse model of gout arthritis. Br. J. Pharmacol. 2020, 177, 2042–2057. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.; Sharma, A.; Bachheti, R.K.; Pandey, D.P. A Comparative Study of the Chemical Composition of the Essential oil from Eucalyptus globulus Growing in Dehradun (India) and Around the World. Orient. J. Chem. 2016, 32, 331–340. [Google Scholar] [CrossRef]
- Sitaraman, R.; Rao, G. A Pediatric Case of Accidental Eucalyptus Oil Poisoning from New Delhi, India: Emergency Measures, Historical Context, and Implications for Practice. Cureus 2019, 11, e5734. [Google Scholar] [CrossRef]
- Ittyachen, A.M.; George, G.R.; Radhakrishnan, M.; Joy, Y. Eucalyptus oil poisoning: Two case reports. J. Med. Case Rep. 2019, 13, 326. [Google Scholar] [CrossRef] [PubMed]
- Flaman, Z.; Pellechia-Clarke, S.; Bailey, B.; McGuigan, M. Unintentional exposure of young children to camphor and eucalyptus oils. Paediatr. Child Health 2001, 6, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Sebei, K.; Sakouhi, F.; Herchi, W.; Khouja, M.L.; Boukhchina, S. Chemical composition and antibacterial activities of seven Eucalyptus species essential oils leaves. Biol. Res. 2015, 48, 7. [Google Scholar] [CrossRef] [PubMed]
- Nurdjannah, N.; Bermawie, N. Cloves. In Handbook of Herbs and Spices; Elsevier: Amsterdam, The Netherlands, 2012; pp. 197–215. [Google Scholar]
- Jirovetz, L.; Buchbauer, G.; Stoilova, I.; Stoyanova, A.; Krastanov, A.; Schmidt, E. Chemical composition and antioxidant properties of clove leaf essential oil. J. Agric. Food Chem. 2006, 54, 6303–6307. [Google Scholar] [CrossRef]
- Carson, C.F.; Hammer, K.A.; Riley, T.V. Melaleuca alternifolia (Tea Tree) oil: A review of antimicrobial and other medicinal properties. Clin. Microbiol. Rev. 2006, 19, 50–62. [Google Scholar] [CrossRef]
- De Groot, A.C.; Schmidt, E. Tea tree oil: Contact allergy and chemical composition. Contact Derm. 2016, 75, 129–143. [Google Scholar] [CrossRef]
- Białoń, M.; Krzyśko-Łupicka, T.; Nowakowska-Bogdan, E.; Wieczorek, P.P. Chemical Composition of Two Different Lavender Essential Oils and Their Effect on Facial Skin Microbiota. Molecules 2019, 24, 3270. [Google Scholar] [CrossRef]
- Kıvrak, Ş. Essential oil composition and antioxidant activities of eight cultivars of Lavender and Lavandin from western Anatolia. Ind. Crops Prod. 2018, 117, 88–96. [Google Scholar] [CrossRef]
- Serafino, A.; Sinibaldi Vallebona, P.; Andreola, F.; Zonfrillo, M.; Mercuri, L.; Federici, M.; Rasi, G.; Garaci, E.; Pierimarchi, P. Stimulatory effect of Eucalyptus essential oil on innate cell-mediated immune response. BMC Immunol. 2008, 9, 17. [Google Scholar] [CrossRef] [PubMed]
- Jordan, A.R.; Racine, R.R.; Hennig, M.J.P.; Lokeshwar, V.B. The Role of CD44 in Disease Pathophysiology and Targeted Treatment. Front. Immunol. 2015, 6, 182. [Google Scholar] [CrossRef] [PubMed]
- Yadav, N.; Chandra, H. Suppression of inflammatory and infection responses in lung macrophages by eucalyptus oil and its constituent 1,8-cineole: Role of pattern recognition receptors TREM-1 and NLRP3, the MAP kinase regulator MKP-1, and NFκB. PLoS ONE 2017, 12, e0188232. [Google Scholar] [CrossRef] [PubMed]
- Juergens, U.R.; Engelen, T.; Racké, K.; Stöber, M.; Gillissen, A.; Vetter, H. Inhibitory activity of 1,8-cineol (eucalyptol) on cytokine production in cultured human lymphocytes and monocytes. Pulm. Pharmacol. Ther. 2004, 17, 281–287. [Google Scholar] [CrossRef]
- Broz, P.; Dixit, V.M. Inflammasomes: Mechanism of assembly, regulation and signalling. Nat. Rev. Immunol. 2016, 16, 407–420. [Google Scholar] [CrossRef]
- Ding, S.; Xu, S.; Ma, Y.; Liu, G.; Jang, H.; Fang, J. Modulatory Mechanisms of the NLRP3 Inflammasomes in Diabetes. Biomolecules 2019, 9, 850. [Google Scholar] [CrossRef]
- Liu, L.; Dong, Y.; Ye, M.; Jin, S.; Yang, J.; Joosse, M.E.; Sun, Y.; Zhang, J.; Lazarev, M.; Brant, S.R.; et al. The Pathogenic Role of NLRP3 Inflammasome Activation in Inflammatory Bowel Diseases of Both Mice and Humans. J. Crohns. Colitis 2017, 11, 737–750. [Google Scholar] [CrossRef]
- Paramel Varghese, G.; Folkersen, L.; Strawbridge, R.J.; Halvorsen, B.; Yndestad, A.; Ranheim, T.; Krohg-Sørensen, K.; Skjelland, M.; Espevik, T.; Aukrust, P.; et al. NLRP3 Inflammasome Expression and Activation in Human Atherosclerosis. J. Am. Heart Assoc. 2016, 5, e003031. [Google Scholar] [CrossRef]
- Roe, K.; Gibot, S.; Verma, S. Triggering receptor expressed on myeloid cells-1 (TREM-1): A new player in antiviral immunity? Front. Microbiol. 2014, 5, 627. [Google Scholar] [CrossRef]
- Goldstein, D.M.; Kuglstatter, A.; Lou, Y.; Soth, M.J. Selective p38α Inhibitors Clinically Evaluated for the Treatment of Chronic Inflammatory Disorders. J. Med. Chem. 2010, 53, 2345–2353. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-y.; Tang, F.-d.; Mao, G.-g.; Bian, R.-l. Effect of alpha-pinene on nuclear translocation of NF-kappa B in THP-1 cells. Acta Pharmacol. Sin. 2004, 25, 480–484. [Google Scholar] [PubMed]
- Zhou, J.-y.; Wang, X.-F.; Tang, F.-d.; Zhou, J.-Y.; Lu, G.-H.; Wang, Y.; Bian, R.-l. Inhibitory effect of 1,8-cineol (eucalyptol) on Egr-1 expression in lipopolysaccharide-stimulated THP-1 cells. Acta Pharmacol. Sin. 2007, 28, 908–912. [Google Scholar] [CrossRef]
- Rouzer, C.A.; Marnett, L.J. Cyclooxygenases: Structural and functional insights. J. Lipid Res. 2009, 50, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Batiha, G.E.; Alkazmi, L.M.; Wasef, L.G.; Beshbishy, A.M.; Nadwa, E.H.; Rashwan, E.K. Syzygium aromaticum L. (Myrtaceae): Traditional Uses, Bioactive Chemical Constituents, Pharmacological and Toxicological Activities. Biomolecules 2020, 10, 202. [Google Scholar] [CrossRef]
- Mateen, S.; Rehman, M.T.; Shahzad, S.; Naeem, S.S.; Faizy, A.F.; Khan, A.Q.; Khan, M.S.; Husain, F.M.; Moin, S. Anti-oxidant and anti-inflammatory effects of cinnamaldehyde and eugenol on mononuclear cells of rheumatoid arthritis patients. Eur. J. Pharmacol. 2019, 852, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Pavesi, C.; Banks, L.; Hudaib, T. Antifungal and antibacterial activities of eugenol and non-polar extract of Syzygium aromaticum L. J. Pharm. Sci. Res. 2018, 10, 337–339. [Google Scholar]
- Ghofran, O.; Safari, T.; Shahraki, M.R. Effects of Eugenol on Pain Response to the Formalin Test and Plasma Antioxidant Activity in High Fructose Drinking Water in Male Rats. Int. J. Prev. Med. 2019, 10, 151. [Google Scholar]
- Gaylor, R.; Michel, J.; Thierry, D.; Panja, R.; Fanja, F.; Pascal, D. Bud, leaf and stem essential oil composition of Syzygium aromaticum from Madagascar, Indonesia and Zanzibar. Int. J. Basic Appl. Sci. 2014, 3, 224–233. [Google Scholar] [CrossRef][Green Version]
- Prashar, A.; Locke, I.C.; Evans, C.S. Cytotoxicity of clove (Syzygium aromaticum) oil and its major components to human skin cells. Cell Prolif. 2006, 39, 241–248. [Google Scholar] [CrossRef]
- Janes, S.E.; Price, C.S.; Thomas, D. Essential oil poisoning: N-acetylcysteine for eugenol-induced hepatic failure and analysis of a national database. Eur. J. Pediatr. 2005, 164, 520–522. [Google Scholar] [CrossRef] [PubMed]
- Carrasco, F.R.; Schmidt, G.; Romero, A.L.; Sartoretto, J.L.; Caparroz-Assef, S.M.; Bersani-Amado, C.A.; Cuman, R.K.N. Immunomodulatory activity of Zingiber officinale Roscoe, Salvia officinalis L. and Syzygium aromaticum L. essential oils: Evidence for humor- and cell-mediated responses. J. Pharm. Pharmacol. 2009, 61, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Halder, S.; Mehta, A.K.; Mediratta, P.K.; Sharma, K.K. Essential oil of clove (Eugenia caryophyllata) augments the humoral immune response but decreases cell mediated immunity. Phytother. Res. 2011, 25, 1254–1256. [Google Scholar] [CrossRef] [PubMed]
- Islamuddin, M.; Chouhan, G.; Want, M.Y.; Ozbak, H.A.; Hemeg, H.A.; Afrin, F. Immunotherapeutic Potential of Eugenol Emulsion in Experimental Visceral Leishmaniasis. PLoS Negl. Trop. Dis. 2016, 10, e0005011. [Google Scholar] [CrossRef]
- Parker, D. CD80/CD86 signaling contributes to the proinflammatory response of Staphylococcus aureus in the airway. Cytokine 2018, 107, 130–136. [Google Scholar] [CrossRef]
- Dibazar, S.P.; Fateh, S.; Daneshmandi, S. Immunomodulatory effects of clove (Syzygium aromaticum) constituents on macrophages: In vitro evaluations of aqueous and ethanolic components. J. Immunotoxicol. 2015, 12, 124–131. [Google Scholar] [CrossRef]
- Choi, C.Y.; Park, K.-R.; Lee, J.-H.; Jeon, Y.J.; Liu, K.-H.; Oh, S.; Kim, D.-E.; Yea, S.S. Isoeugenol suppression of inducible nitric oxide synthase expression is mediated by down-regulation of NF-kappaB, ERK1/2, and p38 kinase. Eur. J. Pharmacol. 2007, 576, 151–159. [Google Scholar] [CrossRef]
- Magalhães, C.B.; Casquilho, N.V.; Machado, M.N.; Riva, D.R.; Travassos, L.H.; Leal-Cardoso, J.H.; Fortunato, R.S.; Faffe, D.S.; Zin, W.A. The anti-inflammatory and anti-oxidative actions of eugenol improve lipopolysaccharide-induced lung injury. Respir. Physiol. Neurobiol. 2019, 259, 30–36. [Google Scholar] [CrossRef]
- Persoons, J.H.; Schornagel, K.; Tilders, F.F.; De Vente, J.; Berkenbosch, F.; Kraal, G. Alveolar macrophages autoregulate IL-1 and IL-6 production by endogenous nitric oxide. Am. J. Respir. Cell Mol. Biol. 1996, 14, 272–278. [Google Scholar] [CrossRef]
- Bachiega, T.F.; de Sousa, J.P.; Bastos, J.K.; Sforcin, J.M. Clove and eugenol in noncytotoxic concentrations exert immunomodulatory/anti-inflammatory action on cytokine production by murine macrophages. J. Pharm. Pharmacol. 2012, 64, 610–616. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Salehi, B.; Varoni, E.M.; Sharopov, F.; Yousaf, Z.; Ayatollahi, S.A.; Kobarfard, F.; Sharifi-Rad, M.; Afdjei, M.H.; Sharifi-Rad, M.; et al. Plants of the Melaleuca Genus as Antimicrobial Agents: From Farm to Pharmacy. Phytother. Res. 2017, 31, 1475–1494. [Google Scholar] [CrossRef] [PubMed]
- Hammer, K.A.; Carson, C.F.; Riley, T.V.; Nielsen, J.B. A review of the toxicity of Melaleuca alternifolia (tea tree) oil. Food Chem. Toxicol. 2006, 44, 616–625. [Google Scholar] [CrossRef]
- Low, P.; Clark, A.M.; Chou, T.-C.; Chang, T.-C.; Reynolds, M.; Ralph, S.J. Immunomodulatory activity of Melaleuca alternifolia concentrate (MAC): Inhibition of LPS-induced NF-κB activation and cytokine production in myeloid cell lines. Int. Immunopharmacol. 2015, 26, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Karin, M. NF-κB, inflammation, immunity and cancer: Coming of age. Nat. Rev. Immunol. 2018, 18, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-Y.; Chen, P.-Y.; Lin, J.-C.; Kirkby, N.S.; Ou, C.-H.; Chang, T.-C. Melaleuca alternifolia Induces Heme Oxygenase-1 Expression in Murine RAW264.7 Cells through Activation of the Nrf2-ARE Pathway. Am. J. Chin. Med. 2017, 45, 1631–1648. [Google Scholar] [CrossRef] [PubMed]
- Budhiraja, S.S.; Cullum, M.E.; Sioutis, S.S.; Evangelista, L.; Habanova, S.T. Biological activity of Melaleuca alternifola (Tea Tree) oil component, terpinen-4-ol, in human myelocytic cell line HL-60. J. Manip. Physiol. Ther. 1999, 22, 447–453. [Google Scholar] [CrossRef]
- Salehi, B.; Mnayer, D.; Özçelik, B.; Altin, G.; Kasapoğlu, K.N.; Daskaya-Dikmen, C.; Sharifi-Rad, M.; Selamoglu, Z.; Acharya, K.; Sen, S.; et al. Plants of the Genus Lavandula: From Farm to Pharmacy. Nat. Prod. Commun. 2018, 13, 1385–1402. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential Oils as Antimicrobial Agents-Myth or Real Alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef]
- Prashar, A.; Locke, I.C.; Evans, C.S. Cytotoxicity of lavender oil and its major components to human skin cells. Cell Prolif. 2004, 37, 221–229. [Google Scholar] [CrossRef]
- Di Sotto, A.; Mazzanti, G.; Carbone, F.; Hrelia, P.; Maffei, F. Genotoxicity of lavender oil, linalyl acetate, and linalool on human lymphocytes in vitro. Environ. Mol. Mutagen. 2011, 52, 69–71. [Google Scholar] [CrossRef]
- Giovannini, D.; Gismondi, A.; Basso, A.; Canuti, L.; Braglia, R.; Canini, A.; Mariani, F.; Cappelli, G. Lavandula angustifolia Mill. Essential Oil Exerts Antibacterial and Anti-Inflammatory Effect in Macrophage Mediated Immune Response to Staphylococcus aureus. Immunol. Investig. 2016, 45, 11–28. [Google Scholar] [CrossRef]
- Chen, P.J.; Chou, C.C.; Yang, L.; Tsai, Y.L.; Chang, Y.C.; Liaw, J.J. Effects of Aromatherapy Massage on Pregnant Women’s Stress and Immune Function: A Longitudinal, Prospective, Randomized Controlled Trial. J. Altern. Complement. Med. 2017, 23, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Kuriyama, H.; Watanabe, S.; Nakaya, T.; Shigemori, I.; Kita, M.; Yoshida, N.; Masaki, D.; Tadai, T.; Ozasa, K.; Fukui, K.; et al. Immunological and Psychological Benefits of Aromatherapy Massage. Evid. Based Complement. Alternat. Med. 2005, 2, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Sasannejad, P.; Saeedi, M.; Shoeibi, A.; Gorji, A.; Abbasi, M.; Foroughipour, M. Lavender essential oil in the treatment of migraine headache: A placebo-controlled clinical trial. Eur. Neurol. 2012, 67, 288–291. [Google Scholar] [CrossRef] [PubMed]
- Ueno-Iio, T.; Shibakura, M.; Yokota, K.; Aoe, M.; Hyoda, T.; Shinohata, R.; Kanehiro, A.; Tanimoto, M.; Kataoka, M. Lavender essential oil inhalation suppresses allergic airway inflammation and mucous cell hyperplasia in a murine model of asthma. Life Sci. 2014, 108, 109–115. [Google Scholar] [CrossRef]
- Lambrecht, B.N.; Hammad, H.; Fahy, J.V. The Cytokines of Asthma. Immunity 2019, 50, 975–991. [Google Scholar] [CrossRef]
- Wang, W.; Shao, S.; Wang, S. The role for human nasal epithelial nuclear factor kappa B activation in histamine-induced mucin 5 subtype B overproduction. Int. Forum Allergy Rhinol. 2016, 6, 264–270. [Google Scholar] [CrossRef]
- Lee, K.Y. M1 and M2 polarization of macrophages: A mini-review. Med. Biol. Sci. Eng. 2019, 2, 1–5. [Google Scholar] [CrossRef]
- Horky, P.; Skalickova, S.; Smerkova, K.; Skladanka, J. Essential Oils as a Feed Additives: Pharmacokinetics and Potential Toxicity in Monogastric Animals. Animals 2019, 9, 352. [Google Scholar] [CrossRef]
- Tammar, S.; Salem, N.; Bettaieb Rebey, I.; Sriti, J.; Hammami, M.; Khammassi, S.; Marzouk, B.; Ksouri, R.; Msaada, K. Regional effect on essential oil composition and antimicrobial activity of Thymus capitatus L. J. Essent. Oil Res. 2019, 31, 129–137. [Google Scholar] [CrossRef]
- Peterfalvi, A.; Miko, E.; Nagy, T.; Reger, B.; Simon, D.; Miseta, A.; Czéh, B.; Szereday, L. Much More Than a Pleasant Scent: A Review on Essential Oils Supporting the Immune System. Molecules 2019, 24, 4530. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Mehdizadeh, L.; Moghaddam, M. Essential Oils: Biological Activity and Therapeutic Potential. In Therapeutic, Probiotic, and Unconventional Foods; Elsevier: Amsterdam, The Netherlands, 2018; pp. 167–179. [Google Scholar]


| Eucalyptus [23,27] | Clove [28,29] | Tea Tree [30,31] | Lavender [32,33] | ||||
|---|---|---|---|---|---|---|---|
| Compound | Content | Compound | Content | Compound | Content | Compound | Content |
| eucalyptol | 50–80% | eugenol | 70–76% | terpinen-4-ol | 30–48% | linalool | 28–45% |
| α-pinene | 2–26% | β-caryophyllene | 10–17% | γ-terpinene | 10–28% | camphor | 3–12% |
| α-terpinyl acetate | 2–5% | α-humulene | 2% | α-terpinene | 5–13% | eucalyptol | 2–10% |
| α-terpineol | 2–3% | eugenyl acetate | 1–12% | α-terpineol | 1–8% | terpinen-4-ol | 2–7% |
| trans-pinocarveol | 1–8% | α-cubebene | 1–2% | p-cymene | 1–8% | β-caryophyllene | 1–6% |
| globulol | 1–6% | α-copaene | 1–2% | α-pinene | 1–6% | borneol | 1–10% |
| limonene | 1–4% | nerolidol | 0–1% | limonene | 1–3% | limonene | 0–3% |
| pinocarvone | 1–4% | farnesol | <1% | sabinene | 0–4% | α-pinene | 0–2% |
| γ-terpinene | 1–2% | methyl chavicol | <1% | δ-cadinene | 0–3% | β-pinene | 0–2% |
| p-cymene | 1–7% | caryophyllene oxide | <1% | eucalyptol | 0–15% | myrcene | 0–2% |
| Essential Oil | Effects on Immune Functions | Concentrations |
|---|---|---|
| Eucalyptus | Increased the phagocytic activities of macrophages and peripheral blood monocytes and enhanced bacterial clearance [34,36]; restored the number of circulating granulocytes and their phagocytic ability in immunosuppressed models [34]; inhibited the production of IL-1α, IL-1β, IL-4, IL-6, TNFα, and NO [34,36]; attenuated the activation of p38 MAPK, NFκB, and TREM-1 [36]; suppressed COX-2 promoter activity by 25% [1] | EEO 0.008 and 0.016% [v/v] (in vitro), EEO 12 mg/kg/day for 15 days (in vivo) [34]; EEO and 1,8-cineole 0.02% [v/v] (in vitro) [36]; EEO 0.01% (in vitro) [1] |
| Clove | Many contradictory results; stimulated cell-mediated immunity in immunocompetent mice and restored WBC count and humoral immunity in immunosuppressed mice [54]; inhibited cell-mediated responses and improved humoral immune responses in immunocompetent rats [55]; suppressed NO and TNFα production by macrophages [58]; stimulated [58] and inhibited [62] IL-6 production; enhanced cell-mediated and humoral immune responses in experimental VL [56]; suppressed COX-2 promoter activity by 40% [1] | CEO (<98% eugenol) 100, 200, 400 mg/kg/day for 7 days (in vivo) [54]; CEO (87.34% eugenol) 0.1 mL/kg/day (in vivo) [55]; ethanolic CEO extract (74% eugenol), aqueous CEO extract (43% eugenol) 0.001–1000 µg/mL (in vitro) [58]; clove extract 100 µg/well, eugenol extract 50 and 100 µg/well (in vitro) [62]; eugenol emulsion 25, 50, and 75 mg/kg/day for 10 days (in vivo) [56]; CEO 0.01% (in vitro) [1] |
| Tea tree | Stimulated the differentiation of immature myelocytes into active phagocytizing monocytes and increased CD11b receptor expression [68]; suppressed the production of TNFα, IL-1β, IL-8, IL-10, and prostaglandin E2 by blood peripheral monocytes [2]; MAC reduced the production of NO and proinflammatory cytokines, inhibited NFκB activation and induced HO-1 expression [65,67] | TTO and terpinen-4-ol 20-90 µmol/L (in vitro) [68]; water soluble components of TTO at 0.125% (42% terpinen-4-ol, 3% α-terpineol and 2% 1,8-cineole) (in vitro) [2]; MAC (60–64% terpinen-4-ol, 8–14% p-cymene) 0.004–0.016% [v/v] (in vitro) [65]; MAC (60% terpinen-4-ol) 0.01–0.5% (in vitro) [67] |
| Lavender | Increased the phagocytic activity of macrophages and reduced intracellular bacterial replication and the production of IL-1α, IL-1β, and IL-6 [73]; attenuated IL-5 and IL-13 secretion and inhibited eosinophilic infiltration and mucus production in mouse asthma models [77]; aromatherapy massage increased IgA levels [74] and the number of CD8+ and CD16+ cells [75] | LEO (39% linalool, 11.97% camphor, 10.54% eucalyptol) dilution of 1:50,000 for 106 cells (in vitro) [73]; LEO (31.78% linalyl acetate, 25.56% linalool) 20 µL on 10 × 10 filter paper (in vivo) [77]; LEO 2% (aromatherapy clinical trial) [74]; essential oil blend of lavender (36.31% linalool, 34.05% linalyl acetate), cypress (61.85% β-pinene, 15.2% 3-carene), and sweet marjoram (21.26% terpinen-4-ol, 13.46% γ-terpinene) (aromatherapy clinical trial) [75] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandner, G.; Heckmann, M.; Weghuber, J. Immunomodulatory Activities of Selected Essential Oils. Biomolecules 2020, 10, 1139. https://doi.org/10.3390/biom10081139
Sandner G, Heckmann M, Weghuber J. Immunomodulatory Activities of Selected Essential Oils. Biomolecules. 2020; 10(8):1139. https://doi.org/10.3390/biom10081139
Chicago/Turabian StyleSandner, Georg, Mara Heckmann, and Julian Weghuber. 2020. "Immunomodulatory Activities of Selected Essential Oils" Biomolecules 10, no. 8: 1139. https://doi.org/10.3390/biom10081139
APA StyleSandner, G., Heckmann, M., & Weghuber, J. (2020). Immunomodulatory Activities of Selected Essential Oils. Biomolecules, 10(8), 1139. https://doi.org/10.3390/biom10081139