The Long-Acting D3 Partial Agonist MC-25-41 Attenuates Motivation for Cocaine in Sprague-Dawley Rats

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
2.1. Animals
2.2. Surgery
2.3. Drug Preparation
2.4. Locomotion
2.5. Progressive Ratio Schedule
2.6. Multiple Schedule of Sucrose and Cocaine Reinforcement
2.7. Within-Session Cocaine Self-Administration Dose-Response Function
2.8. Statistical Analysis
3. Results
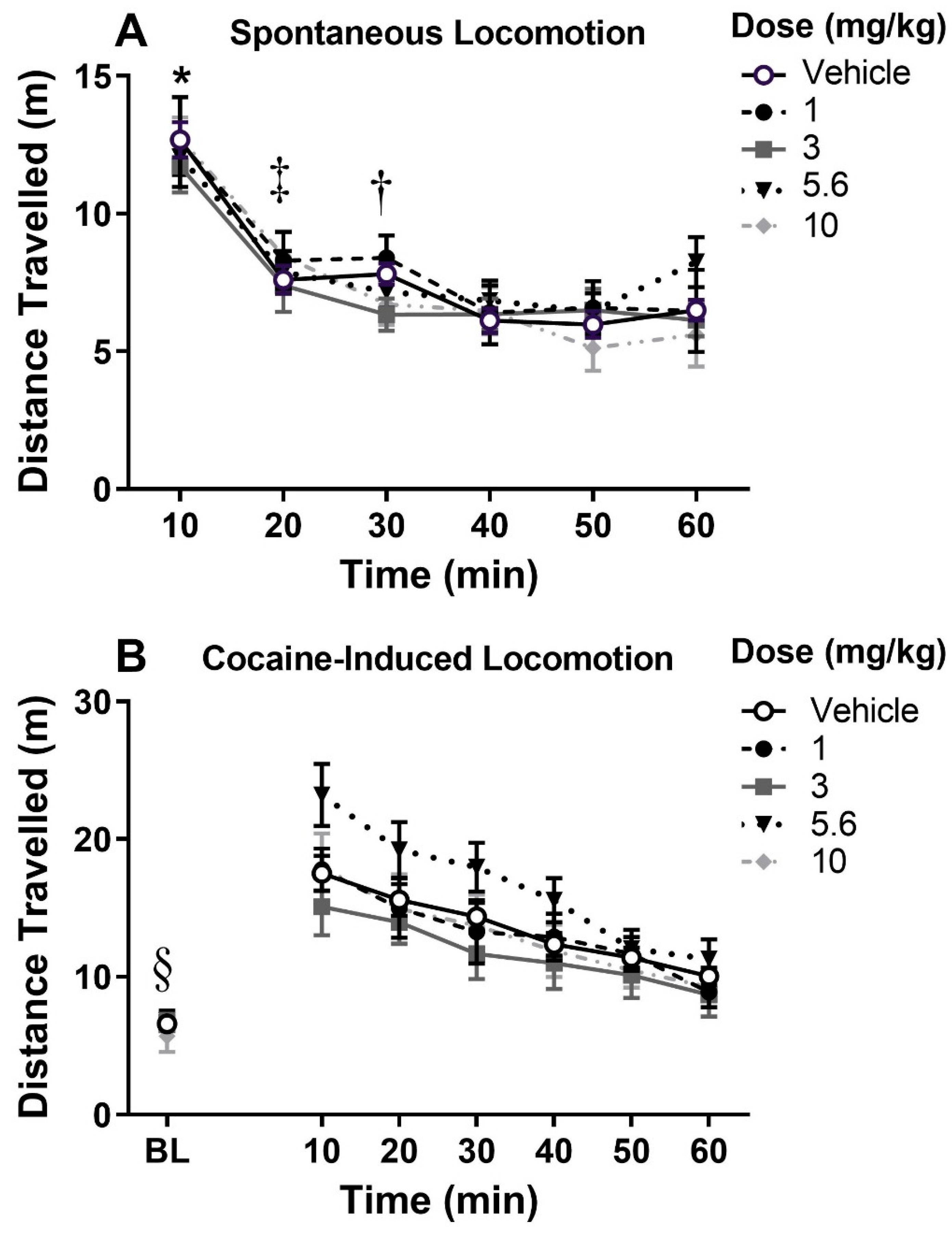
3.1. Spontaneous and Cocaine-Induced Locomotion
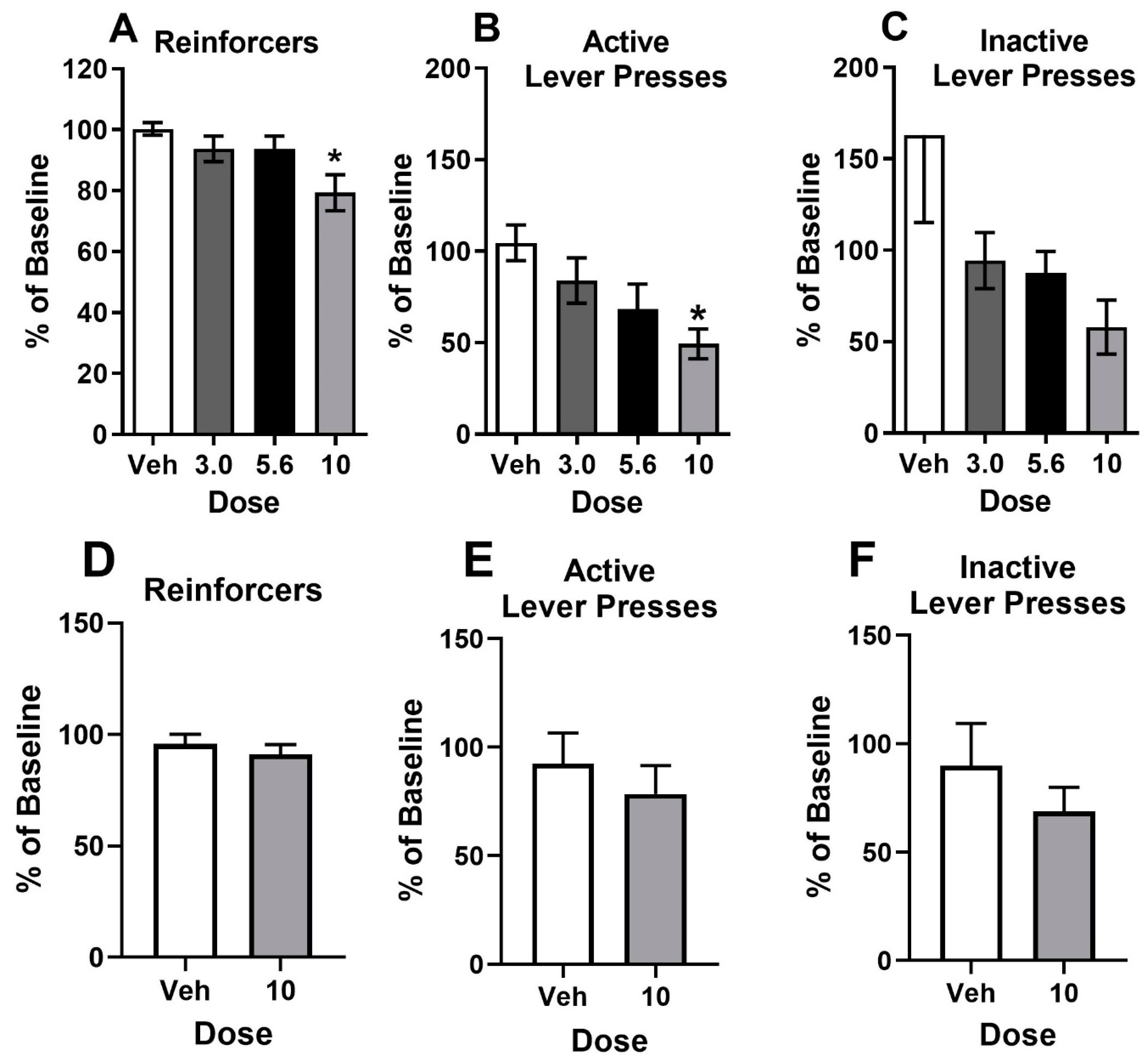
3.2. Progressive Ratio Schedule of Cocaine or Sucrose Reinforcement
3.3. Multiple Schedule of Sucrose and Cocaine Reinforcement
3.4. Behavioral Economics Analysis of Cocaine Self-Administration
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jones, C.M.; Baldwin, G.T.; Compton, W.M. Recent increases in cocaine-related overdose deaths and the role of opioids. Am. J. Public Health 2017, 107, 430–432. [Google Scholar] [CrossRef] [PubMed]
- Van Zee, A. The promotion and marketing of oxycontin: Commercial triumph, public health tragedy. Am. J. Public Health 2009, 99, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Mash, D.C.; Staley, J.K. D3 Dopamine and Kappa Opioid Receptor Alterations in Human Brain of Cocaine-overdose Victims. Ann. N. Y. Acad. Sci. 1999, 877, 507–522. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Volkow, N.D. The neural basis of addiction: A pathology of motivation and choice. Am. J. Psychiatry 2005, 162, 1403–1413. [Google Scholar] [CrossRef] [PubMed]
- Wise, R.A. Dopamine, learning and motivation. Nat. Rev. Neurosci. 2004, 5, 483–494. [Google Scholar] [CrossRef]
- Neisewander, J.L.; Cheung, T.H.C.; Pentkowski, N.S. Dopamine D3 and 5-HT1B receptor dysregulation as a result of psychostimulant intake and forced abstinence: Implications for medications development. Neuropharmacology 2014, 76, 301–319. [Google Scholar] [CrossRef]
- You, Z.B.; Bi, G.H.; Galaj, E.; Kumar, V.; Cao, J.; Gadiano, A.; Rais, R.; Slusher, B.S.; Gardner, E.L.; Xi, Z.X.; et al. Dopamine D3R antagonist VK4-116 attenuates oxycodone self-administration and reinstatement without compromising its antinociceptive effects. Neuropsychopharmacology 2019, 44, 1415–1424. [Google Scholar] [CrossRef]
- Rasmussen, K.; White, D.A.; Acri, J.B. NIDA’s medication development priorities in response to the Opioid Crisis: Ten most wanted. Neuropsychopharmacology 2019, 44, 657–659. [Google Scholar] [CrossRef]
- Sokoloff, P.; Le Foll, B. The dopamine D3 receptor, a quarter century later. Eur. J. Neurosci. 2017, 45, 2–19. [Google Scholar] [CrossRef]
- Pilla, M.; Perachon, S.; Sautel, F.; Garrido, F.; Mann, A.; Wermuth, C.G.; Schwartz, J.C.; Everittt, B.J.; Sokoloff, P. Selective inhibition of cocaine-seeking behaviour by a partial dopamine D3 receptor agonist. Nature 1999, 400, 371–375. [Google Scholar] [CrossRef]
- Powell, G.L.; Bonadonna, J.P.; Vannan, A.; Xu, K.; Mach, R.H.; Luedtke, R.R.; Neisewander, J.L. Dopamine D3 receptor partial agonist LS-3-134 attenuates cocaine-motivated behaviors. Pharmacol. Biochem. Behav. 2018, 175, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Di Ciano, P.; Underwood, R.J.; Hagan, J.J.; Everitt, B.J. Attenuation of cue-controlled cocaine-seeking by a selective D3 dopamine receptor antagonist SB-277011-A. Neuropsychopharmacology 2003, 28, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.X.; Newman, A.H.; Gilbert, J.G.; Pak, A.C.; Peng, X.Q.; Ashby, C.R.; Gitajn, L.; Gardner, E.L. The novel dopamine D3 receptor antagonist NGB 2904 inhibits cocaine’s rewarding effects and cocaine-induced reinstatement of drug-seeking behavior in rats. Neuropsychopharmacology 2006, 31, 1393–1405. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, M.; Barrett, A.C.; Butler, P.; Negus, S.S.; Caine, S.B. Effects of acute and chronic treatments with dopamine D2 and D3 receptor ligands on cocaine versus food choice in rats. J. Pharmacol. Exp. Ther. 2017, 362, 161–176. [Google Scholar] [CrossRef]
- Xi, Z.X.; Gilbert, J.G.; Pak, A.C.; Ashby, C.R.; Heidbreder, C.A.; Gardner, E.L. Selective dopamine D 3 receptor antagonism by SB-277011A attenuates cocaine reinforcement as assessed by progressive-ratio and variable-cost-variable-payoff fixed-ratio cocaine self-administration in rats. Eur. J. Neurosci. 2005, 21, 3427–3438. [Google Scholar] [CrossRef]
- Gilbert, J.G.; Newman, A.H.; Gardner, E.L.; Ashby, C.R.; Heidbreder, C.A.; Pak, A.C.; Peng, X.-Q.; Xi, Z.-X. Acute administration of SB-277011A, NGB 2904, or BP 897 inhibits cocaine cue-induced reinstatement of drug-seeking behavior in rats: Role of dopamine D3 receptors. Synapse 2005, 57, 17–28. [Google Scholar] [CrossRef]
- Bickel, W.K.; Johnson, M.W.; Koffarnus, M.N.; MacKillop, J.; Murphy, J.G. The Behavioral Economics of Substance Use Disorders: Reinforcement Pathologies and Their Repair. Annu. Rev. Clin. Psychol. 2014, 10, 641–677. [Google Scholar] [CrossRef]
- Bentzley, B.S.; Jhou, T.C.; Aston-Jones, G. Economic demand predicts addiction-like behavior and therapeutic efficacy of oxytocin in the rat. Proc. Natl. Acad. Sci. USA 2014, 111, 11822–11827. [Google Scholar] [CrossRef]
- Oleson, E.B.; Richardson, J.M.; Roberts, D.C.S. A novel IV cocaine self-administration procedure in rats: Differential effects of dopamine, serotonin, and GABA drug pre-treatments on cocaine consumption and maximal price paid. Psychopharmacology 2011, 214, 567–577. [Google Scholar] [CrossRef]
- Fragale, J.E.; Pantazis, C.B.; James, M.H.; Aston-Jones, G. The role of orexin-1 receptor signaling in demand for the opioid fentanyl. Neuropsychopharmacology 2019, 44, 1690–1697. [Google Scholar] [CrossRef]
- Porter-Stransky, K.A.; Bentzley, B.S.; Aston-Jones, G. Individual differences in orexin-I receptor modulation of motivation for the opioid remifentanil. Addict. Biol. 2017, 22, 303–317. [Google Scholar] [CrossRef] [PubMed]
- Cox, B.M.; Bentzley, B.S.; Regen-Tuero, H.; See, R.E.; Reichel, C.M.; Aston-Jones, G. Oxytocin Acts in Nucleus Accumbens to Attenuate Methamphetamine Seeking and Demand. Biol. Psychiatry 2017, 81, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.J.; Taylor, M.; Griffin, S.A.; Amani, A.; Hayatshahi, H.; Korzekwa, K.; Ye, M.; Mach, R.H.; Liu, J.; Luedtke, R.R.; et al. Design, synthesis, and evaluation of N-(4-(4-phenyl piperazin-1-yl)butyl)-4-(thiophen-3-yl)benzamides as selective dopamine D3 receptor ligands. Bioorganic Med. Chem. Lett. 2019, 29, 2690–2694. [Google Scholar] [CrossRef] [PubMed]
- Oleson, E.B.; Roberts, D.C. Behavioral economic assessment of price and cocaine consumption following self-administration histories that produce escalation of either final ratios or intake. Neuropsychopharmacology 2009, 34, 796–804. [Google Scholar] [CrossRef]
- Kufahl, P.R.; Zavala, A.R.; Singh, A.; Thiel, K.J.; Dickey, E.D.; Joyce, J.N.; Neisewander, J.L. c-Fos expression associated with reinstatement of cocaine-seeking behavior by response-contingent conditioned cues. Synapse 2009, 63, 823–835. [Google Scholar] [CrossRef]
- Pentkowski, N.S.; Duke, F.D.; Weber, S.M.; Pockros, L.A.; Teer, A.P.; Hamilton, E.C.; Thiel, K.J.; Neisewander, J.L. Stimulation of Medial Prefrontal Cortex Serotonin 2C (5-HT2C) Receptors Attenuates Cocaine-Seeking Behavior. Neuropsychopharmacology 2010, 35, 2037–2048. [Google Scholar] [CrossRef]
- Roberts, D.; Richardson, N. Self-administration of psychomotor stimulants using progressive ratio schedules of reinforcement. In Animal Models of Drug Addiction; Boulton, A., Baker, G., Wu, P., Eds.; Humana Press: Totowa, NJ, USA, 1992; pp. 233–269. [Google Scholar]
- Cheung, T.H.; Loriaux, A.L.; Weber, S.M.; Chandler, K.N.; Lenz, J.D.; Schaan, R.F.; Mach, R.H.; Luedtke, R.R.; Neisewander, J.L. Reduction of cocaine self-administration and D3 receptor-mediated behavior by two novel dopamine D3 receptor-selective partial agonists, OS-3-106 and WW-III-55. J. Pharmacol. Exp. Ther. 2013, 347, 410–423. [Google Scholar] [CrossRef]
- Bentzley, B.S.; Fender, K.M.; Aston-Jones, G. The behavioral economics of drug self-administration: A review and new analytical approach for within-session procedures. Psychopharmacology 2013, 226, 113–125. [Google Scholar] [CrossRef]
- Koffarnus, M.N.; Franck, C.T.; Stein, J.S.; Bickel, W.K. A modified exponential behavioral economic demand model to better describe consumption data. Exp. Clin. Psychopharmacol. 2015, 23, 504–512. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2018. Available online: https://www.R-project.org/ (accessed on 15 June 2020).
- Pinheiro, J.; Bates, D.; Debroy, S.; Sarkar, D. Linear and Nonlinear Mixed Effects. R Packag. Version 3.1-122 2015, 3, 1–96. [Google Scholar]
- Hursh, S.R.; Roma, P.G. Behavioral Economics and the Analysis of Consumption and Choice. Manag. Decis. Econ. 2016, 37, 224–238. [Google Scholar] [CrossRef]
- Bentzley, B.S.; Aston-Jones, G. Orexin-1 receptor signaling increases motivation for cocaine-associated cues. Eur. J. Neurosci. 2015, 41, 1149–1156. [Google Scholar] [CrossRef] [PubMed]
- Cheung, T.H.C.; Nolan, B.C.; Hammerslag, L.R.; Weber, S.M.; Durbin, J.P.; Peartree, N.A.; Mach, R.H.; Luedtke, R.R.; Neisewander, J.L. Phenylpiperazine derivatives with selectivity for dopamine D3 receptors modulate cocaine self-administration in rats. Neuropharmacology 2012, 63, 1346–1359. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kosten, T.R.; George, T.P.; Kosten, T.A. The potential of dopamine agonists in drug addiction. Expert Opin. Investig. Drugs 2002, 11, 491–499. [Google Scholar] [PubMed]
- Newman, A.H.; Blaylock, B.L.; Nader, M.A.; Bergman, J.; Sibley, D.R.; Skolnick, P. Medication discovery for addiction: Translating the dopamine D3 receptor hypothesis. Biochem. Pharmacol. 2012, 84, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Le Foll, B.; Schwartz, J.C.; Sokoloff, P. Dopamine D3 receptor agents as potential new medications for drug addiction. Eur. Psychiatry 2000, 15, 140–146. [Google Scholar] [CrossRef]
- Moreira, F.A.; Dalley, J.W. Dopamine receptor partial agonists and addiction. Eur. J. Pharmacol. 2015, 752, 112–115. [Google Scholar] [CrossRef] [PubMed]
- Childress, A.R.; O’Brien, C.P. Dopamine receptor partial agonists could address the duality of cocaine craving. Trends Pharmacol. Sci. 2000, 21, 6–9. [Google Scholar] [CrossRef]
- Higley, A.E.; Kiefer, S.W.; Li, X.; Gaál, J.; Xi, Z.X.; Gardner, E.L. Dopamine D3 receptor antagonist SB-277011A inhibits methamphetamine self-administration and methamphetamine-induced reinstatement of drug-seeking in rats. Eur. J. Pharmacol. 2011, 659, 187–192. [Google Scholar] [CrossRef]
- Orio, L.; Wee, S.; Newman, A.H.; Pulvirenti, L.; Koob, G.F. The dopamine D3 receptor partial agonist CJB090 and antagonist PG01037 decrease progressive ratio responding for methamphetamine in rats with extended-access. Addict. Biol. 2010, 15, 312–323. [Google Scholar] [CrossRef]
- Piazza, P.V.; Deroche-Gamonent, V.; Rouge-Pont, F.; Le Moal, M. Vertical shifts in self-administration dose-response functions predict a drug-vulnerable phenotype predisposed to addiction. J. Neurosci. 2000, 20, 4226–4232. [Google Scholar] [CrossRef] [PubMed]
- Kenakin, T. Pharmacological Analysis of Drug-Receptor Interaction; Raven: New York, NY, USA, 1993; ISBN 9780781700658. [Google Scholar]
- Ahmed, S.H.; Koob, G.F. Transition from Moderate to Excessive Drug Intake: Change in Hedonic Set Point. Science 1998, 282, 298–300. [Google Scholar] [CrossRef] [PubMed]
- Egli, M.; White, D.A.; Acri, J.B. Considerations in the Evaluation of Potential Efficacy of Medications for Alcohol and Drug Use Disorders: An. Editorial, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 126. [Google Scholar]
- Vorel, S.R.; Ashby, C.R., Jr.; Paul, M.; Liu, X.; Hayes, R.; Hagan, J.J.; Middlemiss, D.N.; Stemp, G.; Gardner, E.L. Dopamine D3 receptor antagonism inhibits cocaine-seeking and cocaine-enhanced brain reward in rats. J. Neurosci. 2002, 22, 9595–9603. [Google Scholar] [CrossRef]
- Achat-Mendes, C.; Platt, D.M.; Newman, A.H.; Spealman, R.D. The dopamine D3 receptor partial agonist CJB 090 inhibits the discriminative stimulus but not the reinforcing or priming effects of cocaine in squirrel monkeys. Psychopharmacology 2009, 206, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Gyertyan, I.; Kiss, B.; Gal, K.; Laszlovszky, I.; Horvath, A.; Gemesi, L.I.; Saghy, K.; Pasztor, G.; Zajer, M.; Kapas, M.; et al. Effects of RGH-237 [N-{4-[4-(3-aminocarbonyl-phenyl)-piperazin-1-yl]-butyl}-4-bromo-benzamide], an orally active, selective dopamine D(3) receptor partial agonist in animal models of cocaine abuse. J. Pharmacol. Exp. Ther. 2007, 320, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Gál, K.; Gyertyán, I. Dopamine D3 as well as D2 receptor ligands attenuate the cue-induced cocaine-seeking in a relapse model in rats. Drug Alcohol Depend. 2006, 81, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Yang, R.-F.; Wu, N.; Su, R.-B.; Li, J.; Peng, X.-Q.; Li, X.; Gaál, J.; Xi, Z.-X.; Gardner, E.L. YQA14: A novel dopamine D3 receptor antagonist that inhibits cocaine self-administration in rats and mice, but not in D3 receptor-knockout mice. Addict. Biol. 2012, 17, 259–273. [Google Scholar] [CrossRef]
- Burmeister, J.J.; Lungren, E.M.; Kirschner, K.F.; Neisewander, J.L. Differential roles of 5-HT receptor subtypes in cue and cocaine reinstatement of cocaine-seeking behavior in rats. Neuropsychopharmacology 2004, 29, 660–668. [Google Scholar] [CrossRef]
- Harvey-Lewis, C.; Li, Z.; Higgins, G.A.; Fletcher, P.J. The 5-HT2C receptor agonist lorcaserin reduces cocaine self-administration, reinstatement of cocaine-seeking and cocaine induced locomotor activity. Neuropharmacology 2016, 101, 237–245. [Google Scholar] [CrossRef]
- Bontempi, L.; Savoia, P.; Bono, F.; Fiorentini, C.; Missale, C. Dopamine D3 and acetylcholine nicotinic receptor heteromerization in midbrain dopamine neurons: Relevance for neuroplasticity. Eur. Neuropsychopharmacol. 2017, 27, 313–324. [Google Scholar] [CrossRef]
- Sabioni, P.; Di Ciano, P.; Le Foll, B. Effect of a D3 receptor antagonist on context-induced reinstatement of nicotine seeking. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 64, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Galaj, E.; Ewing, S.; Ranaldi, R. Dopamine D1 and D3 receptor polypharmacology as a potential treatment approach for substance use disorder. Neurosci. Biobehav. Rev. 2018, 89, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Heidbreder, C.A.; Newman, A.H. Current perspectives on selective dopamine D3 receptor antagonists as pharmacotherapeutics for addictions and related disorders. Ann. N. Y. Acad. Sci. 2010, 1187, 4–34. [Google Scholar] [CrossRef]
- Keck, T.M.; John, W.S.; Czoty, P.W.; Nader, M.A.; Newman, A.H. Identifying Medication Targets for Psychostimulant Addiction: Unraveling the Dopamine D3 Receptor Hypothesis. J. Med. Chem. 2015, 58, 5361–5380. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Xing, B.; Chu, Z.; Liu, F.; Lei, G.; Zhu, L.; Gao, Y.; Chen, T.; Dang, Y. Dopamine D3 Receptor Knockout Mice Exhibit Abnormal Nociception in a Sex-Different Manner. J. Neurosci. Res. 2017, 1445, 1438–1445. [Google Scholar] [CrossRef] [PubMed]
- Klinker, F.; Köhnemann, K.; Paulus, W.; Liebetanz, D. Dopamine D3 receptor status modulates sexual dimorphism in voluntary wheel running behavior in mice. Behav. Brain Res. 2017, 333, 235–241. [Google Scholar] [CrossRef]
- Jackson, L.R.; Robinson, T.E.; Becker, J.B. Sex differences and hormonal influences on acquisition of cocaine self-administration in rats. Neuropsychopharmacology 2006, 31, 129–138. [Google Scholar] [CrossRef]
- Martelle, S.E.; Nader, S.H.; Czoty, P.W.; John, W.S.; Duke, A.N.; Garg, P.K.; Garg, S.; Newman, A.H.; Nader, M.A. Further Characterization of Quinpirole-Elicited Yawning as a Model of Dopamine D3 Receptor Activation in Male and Female Monkeys. J. Pharmacol. Exp. Ther. 2014, 350, 205–211. [Google Scholar] [CrossRef]
- Becker, J.B. Gender differences in dopaminergic function in striatum and nucleus accumbens. Pharmacol. Biochem. Behav. 1999, 64, 803–812. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Powell, G.L.; Namba, M.D.; Vannan, A.; Bonadonna, J.P.; Carlson, A.; Mendoza, R.; Chen, P.-J.; Luetdke, R.R.; Blass, B.E.; Neisewander, J.L. The Long-Acting D3 Partial Agonist MC-25-41 Attenuates Motivation for Cocaine in Sprague-Dawley Rats. Biomolecules 2020, 10, 1076. https://doi.org/10.3390/biom10071076
Powell GL, Namba MD, Vannan A, Bonadonna JP, Carlson A, Mendoza R, Chen P-J, Luetdke RR, Blass BE, Neisewander JL. The Long-Acting D3 Partial Agonist MC-25-41 Attenuates Motivation for Cocaine in Sprague-Dawley Rats. Biomolecules. 2020; 10(7):1076. https://doi.org/10.3390/biom10071076
Chicago/Turabian StylePowell, Gregory L., Mark D. Namba, Annika Vannan, John Paul Bonadonna, Andrew Carlson, Rachel Mendoza, Peng-Jen Chen, Robert R. Luetdke, Benjamin E. Blass, and Janet L. Neisewander. 2020. "The Long-Acting D3 Partial Agonist MC-25-41 Attenuates Motivation for Cocaine in Sprague-Dawley Rats" Biomolecules 10, no. 7: 1076. https://doi.org/10.3390/biom10071076
APA StylePowell, G. L., Namba, M. D., Vannan, A., Bonadonna, J. P., Carlson, A., Mendoza, R., Chen, P.-J., Luetdke, R. R., Blass, B. E., & Neisewander, J. L. (2020). The Long-Acting D3 Partial Agonist MC-25-41 Attenuates Motivation for Cocaine in Sprague-Dawley Rats. Biomolecules, 10(7), 1076. https://doi.org/10.3390/biom10071076