Nematicidal Activity of Holigarna caustica (Dennst.) Oken Fruit Is Due to Linoleic Acid


 and
and 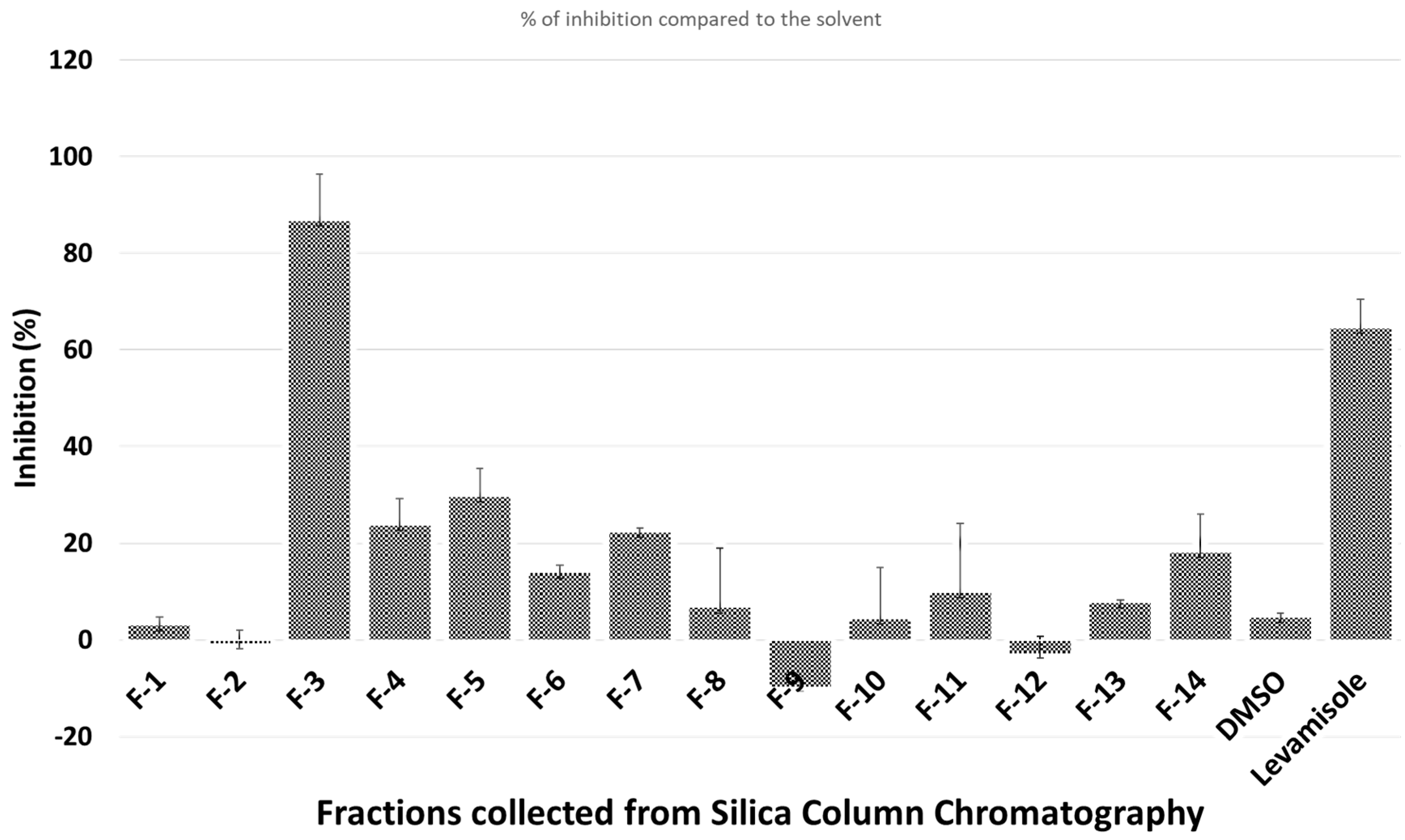{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Plant Collection and Extraction
2.3. Bioassay-Guided Fractionation and Isolation
2.4. HPLC-DAD Analysis
2.5. Mass Spectrometry (MS)
2.6. NMR Spectroscopy
2.7. Nematicidal Activity
2.8. Cytotoxicity Test
2.9. Statistical Analysis
3. Results and Discussion
3.1. Fruit Extracts of H. caustica Prevent the Movement of C. elegans
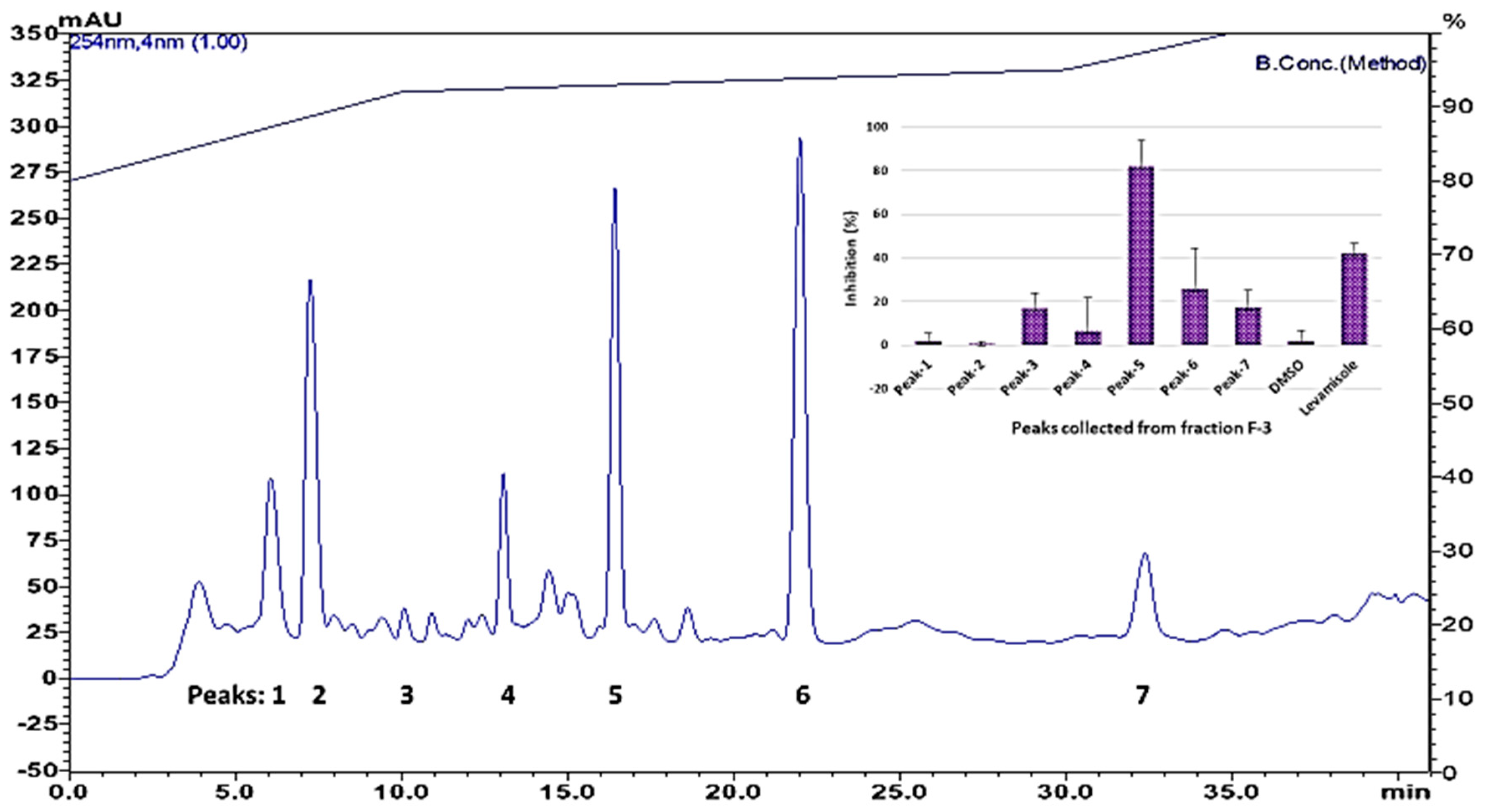
3.2. Bioassay-Guided Purification of Nematicidal Compounds
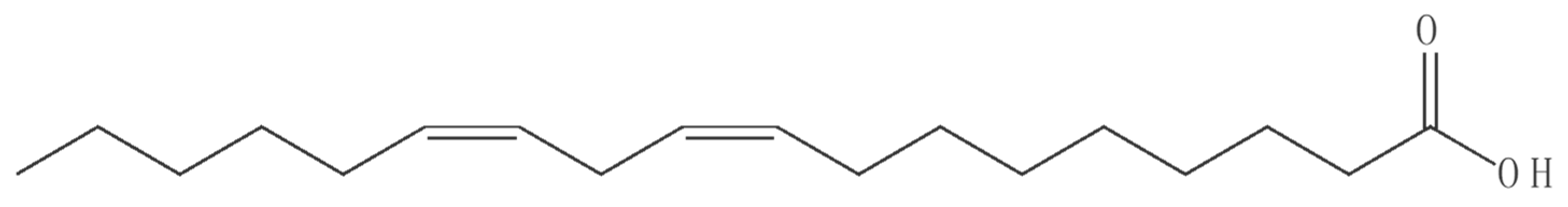
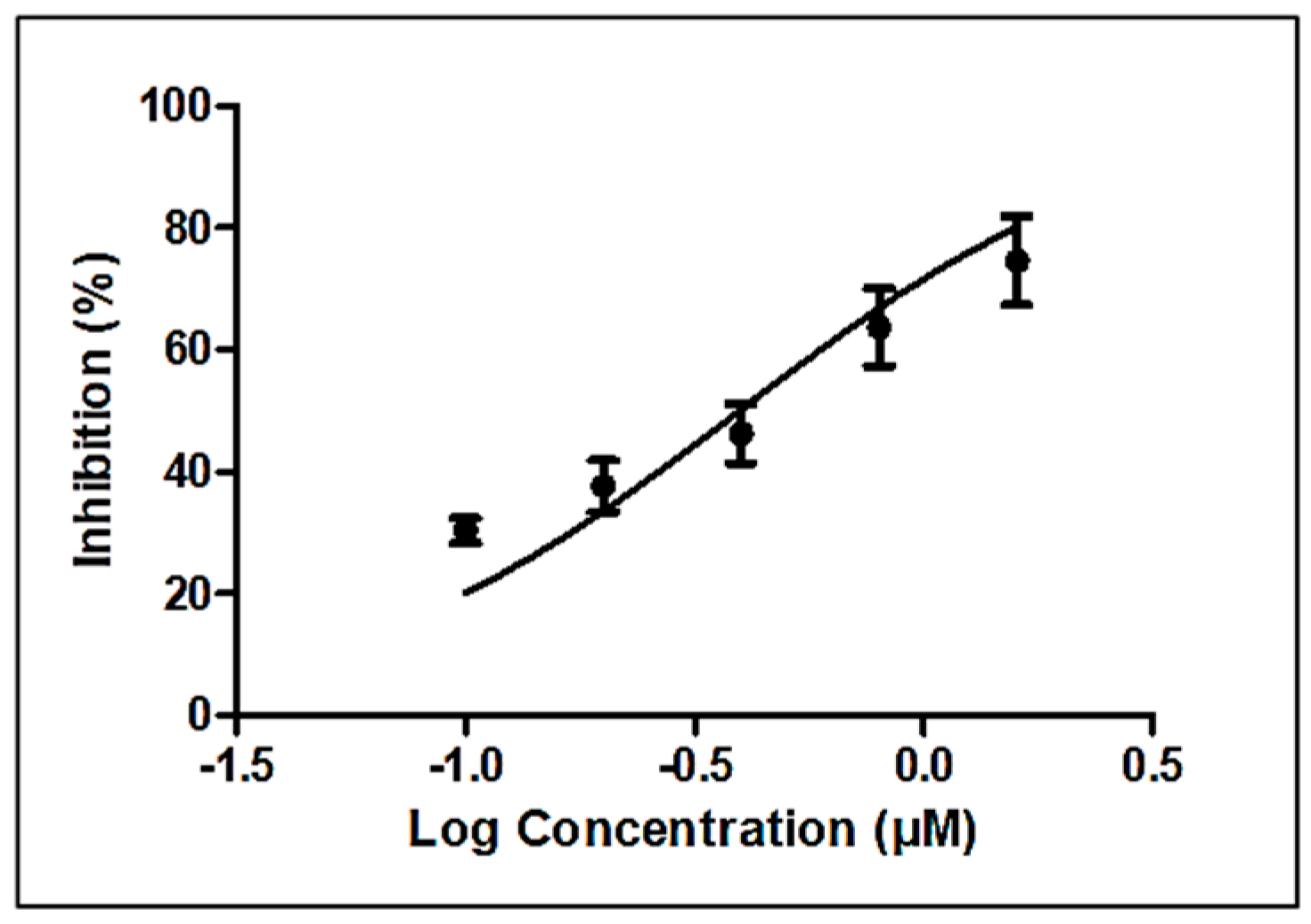
3.3. Nematicidal Activity of Linoleic Acid
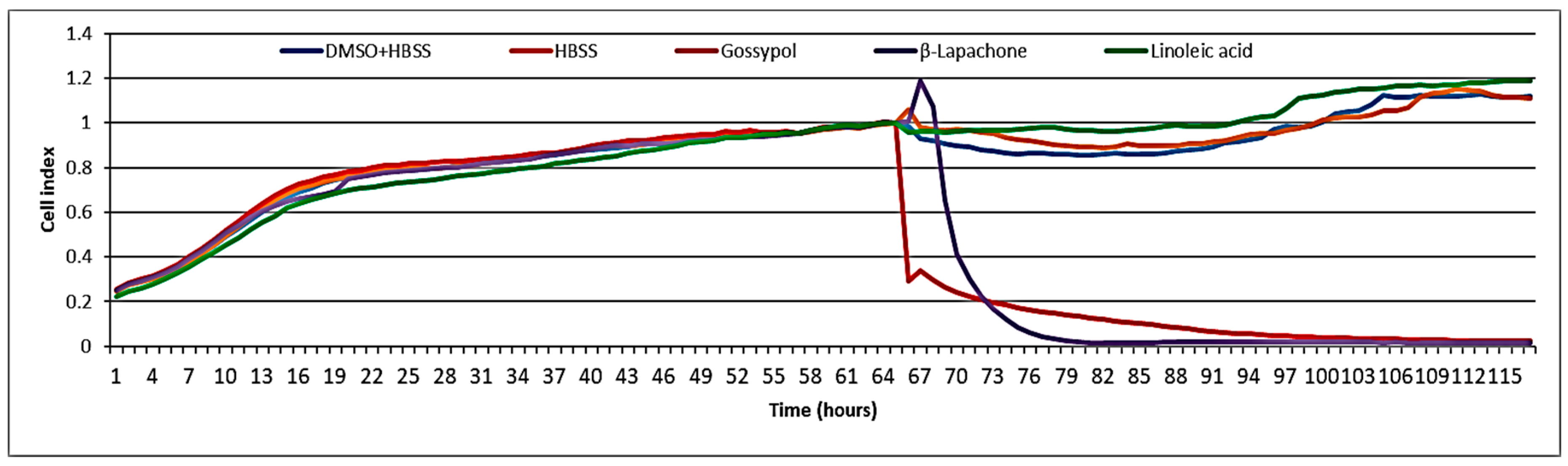
3.4. Cytotoxicity of Linoleic Acid
3.5. Suitability of H. caustica Fruit or Linoleic Acid as Anthelmintic
3.6. Mechanism of Action of Linoleic Acid
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Chemical Compounds Studied in This Article
Abbreviations
| ATCC | American Type Culture Collection, Manassas, Virginia, USA |
| BJ5ta | Normal fibroblast immortalized with hTERT |
| CRF | Chirang Reserve Forest |
| DMSO | Dimethyl sulfoxide |
| ESI | Electrospray ionization |
| FBS | Fetal bovine serum |
| HPLC | High performance liquid chromatography |
| IC50 | 50% Inhibitory concentration |
| MS | Mass spectrometry |
| NMR | Nuclear magnetic resonance |
| NTDs | Neglected tropical diseases |
| TFA | Trifluoro acetic acid |
| WHO | World Health Organization |
References
- Hotez, P.J.; Molyneux, D.H.; Fenwick, A.; Kumaresan, J.; Sachs, S.E.; Sachs, J.D.; Savioli, L. Control of Neglected Tropical Diseases. N. Engl. J. Med. 2007, 357, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Charlier, J.; van der Voort, M.; Kenyon, F.; Skuce, P.; Vercruysse, J. Chasing helminths and their economic impact on farmed ruminants. Trends Parasitol. 2014, 30, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, J.L. Global food security: The impact of veterinary parasites and parasitologists. Vet. Parasitol. 2013, 195, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Panda, S.K.; Luyten, W. Plant-Based Natural Products for the Discovery and Development of Novel Anthelmintics against Nematodes. Biomolecules 2020, 10, 426. [Google Scholar] [CrossRef]
- Hu, Y.; Xiao, S.-H.; Aroian, R.V. The new anthelmintic Tribendimidine is an l-type (levamisole and pyrantel) nicotinic acetylcholine receptor agonist. PLoS Negl. Trop. Dis. 2009, 3, e499. [Google Scholar] [CrossRef]
- Rath, S.K.; Mohapatra, N.; Dubey, D.; Panda, S.K.; Thatoi, H.N.; Dutta, S.K. Antimicrobial activity of Diospyros melanoxylon bark from Similipal Biosphere Reserve, Orissa, India. Afr. J. Biotechnol. 2009, 8, 1924–1928. [Google Scholar]
- Panda, S.K.; Luyten, W. Antiparasitic activity in Asteraceae with special attention to ethnobotanical use by the tribes of Odisha, India. Parasite 2018, 25, 10. [Google Scholar] [CrossRef]
- Idris, O.A.; Wintola, O.A.; Afolayan, A.J. Helminthiases; prevalence, transmission, host-parasite interactions, resistance to common synthetic drugs and treatment. Heliyon 2019, 5, e01161. [Google Scholar] [CrossRef]
- Panda, S.K. Ethno-medicinal uses and screening of plants for antibacterial activity from Similipal Biosphere Reserve, Odisha, India. J. Ethnopharmacol. 2014, 151, 158–175. [Google Scholar] [CrossRef]
- Adnan, M.; Nazim Uddin Chy, M.; Mostafa Kamal, A.T.M.; Barlow, J.W.; Faruque, M.O.; Yang, X.; Uddin, S.B. Evaluation of anti-nociceptive and anti-inflammatory activities of the methanol extract of Holigarna caustica (Dennst.) Oken leaves. J. Ethnopharmacol. 2019, 236, 401–411. [Google Scholar] [CrossRef]
- Panda, S.K.; Das, R.; Lavigne, R.; Luyten, W. Indian medicinal plant extracts to control multidrug-resistant S. aureus, including in biofilms. S. Afr. J. Bot. 2020, 128, 283–291. [Google Scholar] [CrossRef]
- Panda, S.K.; Das, R.; Leyssen, P.; Neyts, J.; Luyten, W. Assessing medicinal plants traditionally used in the Chirang Reserve Forest, Northeast India for antimicrobial activity. J. Ethnopharmacol. 2018, 225, 220–233. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Mohanta, Y.K.; Padhi, L.; Park, Y.-H.; Mohanta, T.K.; Bae, H. Large scale screening of ethnomedicinal plants for identification of potential antibacterial compounds. Molecules 2016, 21, 293. [Google Scholar] [CrossRef] [PubMed]
- Panda, S.K.; Padhi, L.; Leyssen, P.; Liu, M.; Neyts, J.; Luyten, W. Antimicrobial, anthelmintic, and antiviral activity of plants traditionally used for treating infectious disease in the Similipal Biosphere Reserve, Odisha, India. Front. Pharmacol. 2017, 8, 658. [Google Scholar] [CrossRef]
- Kerkoub, N.; Panda, S.K.; Yang, M.-R.; Lu, J.-G.; Jiang, Z.-H.; Nasri, H.; Luyten, W. Bioassay-Guided Isolation of Anti-Candida Biofilm Compounds From Methanol Extracts of the Aerial Parts of Salvia officinalis (Annaba, Algeria). Front. Pharmacol. 2018, 9, 1418. [Google Scholar] [CrossRef]
- Ma, Y.; Cong, W.; Huang, H.; Sun, L.; Mai, A.H.; Boonen, K.; Maryam, W.; De Borggraeve, W.; Luo, G.; Liu, Q.; et al. Identification of fukinolic acid from Cimicifuga heracleifolia and its derivatives as novel antiviral compounds against enterovirus A71 infection. Int. J. Antimicrob. Agents 2019, 53, 128–136. [Google Scholar] [CrossRef]
- Patiny, L.; Borel, A. ChemCalc: A Building Block for Tomorrow’s Chemical Infrastructure. J. Chem. Inf. Modeling 2013, 53, 1223–1228. [Google Scholar] [CrossRef]
- Bouhenna, M.M.; Mameri, N.; Pérez, M.V.; Talhi, O.; Bachari, K.; Silva, A.M.S.; Luyten, W. Anticancer Activity Study of Chromone and Coumarin Hybrids using Electrical Impedance Spectroscopy. Anti-Cancer Agents Med. Chem. 2018, 18, 854–864. [Google Scholar] [CrossRef]
- Panda, S.K.; Mohanta, Y.K.; Padhi, L.; Luyten, W. Antimicrobial activity of select edible plants from Odisha, India against food-borne pathogens. LWT 2019, 113, 108246. [Google Scholar] [CrossRef]
- Harun-Ur-Rashid, M.; Islam, S.; Kashem, S.B. Floristic diversity (Magnoliids and Eudicots) of Baraiyadhala National Park, Chittagong, Bangladesh. Bangladesh J. Plant Taxon. 2018, 25, 273–288. [Google Scholar] [CrossRef]
- Jadhav, V.; Kalase, V.; Patil, P. GC-MS analysis of bioactive compounds in methanolic extract of Holigarna grahamii (wight) Kurz. Int. J. Herb. Med. 2014, 2, 35–39. [Google Scholar]
- Risi, G.; Aguilera, E.; Ladós, E.; Suárez, G.; Carrera, I.; Álvarez, G.; Salinas, G. Caenorhabditis elegans infrared-based motility assay identified new hits for nematicide drug development. Vet. Sci. 2019, 6, 29. [Google Scholar] [CrossRef] [PubMed]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A Transparent Window into Biology: A Primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, L.C.A.; Barcelos, F.F.; Demuner, A.J.; Santos, M.A. Investigation-Research: Chemical constituents from Mucuna aterrima with activity against Meloidogyne incognita and Heterodera glycines. Nematropica 1999, 29, 81–88. [Google Scholar]
- Ayers, S.; Zink, D.L.; Mohn, K.; Powell, J.S.; Brown, C.M.; Bills, G.; Grund, A.; Thompson, D.; Singh, S.B. Anthelmintic constituents of Clonostachys candelabrum. J. Antibiot. 2010, 63, 119–122. [Google Scholar] [CrossRef]
- Stadler, M.; Anke, H.; Sterner, O. Linoleic acid—The nematicidal principle of several nematophagous fungi and its production in trap-forming submerged cultures. Arch. Microbiol. 1993, 160, 401–405. [Google Scholar] [CrossRef]
- Stadler, M.; Mayer, A.; Anke, H.; Sterner, O. Fatty Acids and Other Compounds with Nematicidal Activity from Cultures of Basidiomycetes. Planta Med. 1994, 60, 128–132. [Google Scholar] [CrossRef]
- Kumarasingha, R.; Karpe, A.V.; Preston, S.; Yeo, T.-C.; Lim, D.S.L.; Tu, C.-L.; Luu, J.; Simpson, K.J.; Shaw, J.M.; Gasser, R.B.; et al. Metabolic profiling and in vitro assessment of anthelmintic fractions of Picria fel-terrae Lour. Int. J. Parasitol. Drugs Drug Resist. 2016, 6, 171–178. [Google Scholar] [CrossRef]
- Faizi, S.; Fayyaz, S.; Bano, S.; Yawar Iqbal, E.; Siddiqi, H.; Naz, A. Isolation of Nematicidal Compounds from Tagetes patula L. Yellow Flowers: Structure–Activity Relationship Studies against Cyst Nematode Heterodera zeae Infective Stage Larvae. J. Agric. Food Chem. 2011, 59, 9080–9093. [Google Scholar] [CrossRef]
- Melariri, P.; Campbell, W.; Etusim, P.; Smith, P. In Vitro and in Vivo Antimalarial Activity of Linolenic and Linoleic Acids and their Methyl Esters. Adv. Stud. Biol. 2012, 4, 333–349. [Google Scholar]
- Shultz, T.D.; Chew, B.P.; Seaman, W.R. Differential stimulatory and inhibitory responses of human MCF-7 breast cancer cells to linoleic acid and conjugated linoleic acid in culture. Anticancer Res. 1992, 12, 2143–2145. [Google Scholar] [PubMed]
- Cunningham, D.; Harrison, L.; Shultz, T. Proliferative responses of normal human mammary and MCF-7 breast cancer cells to linoleic acid, conjugated linoleic acid and eicosanoid synthesis inhibitors in culture. Anticancer Res. 1997, 17, 197–203. [Google Scholar]
- Zock, P.L.; Katan, M.B. Linoleic acid intake and cancer risk: A review and meta-analysis. Am. J. Clin. Nutr. 1998, 68, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Jandacek, R.J.; Jandacek, J.R. Linoleic Acid: A Nutritional Quandary. Healthcare 2017, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Asselin, C.; Ducharme, A.; Ntimbane, T.; Ruiz, M.; Fortier, A.; Guertin, M.-C.; Lavoie, J.; Diaz, A.; Levy, É.; Tardif, J.-C.; et al. Circulating levels of linoleic acid and HDL-cholesterol are major determinants of 4-hydroxynonenal protein adducts in patients with heart failure. Redox Biol. 2014, 2, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Abdelmagid, S.A.; Clarke, S.E.; Nielsen, D.E.; Badawi, A.; El-Sohemy, A.; Mutch, D.M.; Ma, D.W.L. Comprehensive Profiling of Plasma Fatty Acid Concentrations in Young Healthy Canadian Adults. PLoS ONE 2015, 10, e0116195. [Google Scholar] [CrossRef]
- Watanabe, Y.; Fang, X.; Minemoto, Y.; Adachi, S.; Matsuno, R. Suppressive effect of saturated acyl L-ascorbate on the oxidation of linoleic acid encapsulated with maltodextrin or gum arabic by spray-drying. J. Agric. Food Chem. 2002, 50, 3984–3987. [Google Scholar] [CrossRef]
- Fang, X.; Watanabe, Y.; Adachi, S.; Matsumura, Y.; Mori, T.; Maeda, H.; Nakamura, A.; Matsuno, R. Microencapsulation of linoleic acid with low- and high-molecular-weight components of soluble soybean polysaccharide and its oxidation process. Biosci. Biotechnol. Biochem. 2003, 67, 1864–1869. [Google Scholar] [CrossRef][Green Version]
- Adnan, M.; Chy, M.N.U.; Kamal, A.T.M.M.; Chowdhury, K.A.A.; Rahman, M.A.; Ali Reza, A.S.M.; Moniruzzaman, M.; Rony, S.R.; Nasrin, M.S.; Azad, M.O.K.; et al. Intervention in neuropsychiatric disorders by suppressing inflammatory and oxidative stress signal and exploration of in silico studies for potential lead compounds from Holigarna caustica (Dennst.) oken leaves. Biomolecules 2020, 10, 561. [Google Scholar] [CrossRef]
- Davis, E.L.; Meyers, D.M.; Dullum, C.J.; Feitelson, J.S. Nematicidal Activity of Fatty Acid Esters on Soybean Cyst and Root-knot Nematodes. J. Nematol. 1997, 29, 677–684. [Google Scholar]
- Liu, M.; Kipanga, P.; Mai, A.H.; Dhondt, I.; Braeckman, B.P.; De Borggraeve, W.; Luyten, W. Bioassay-guided isolation of three anthelmintic compounds from Warburgia ugandensis Sprague subspecies ugandensis, and the mechanism of action of polygodial. Int. J. Parasitol. 2018, 48, 833–844. [Google Scholar] [CrossRef] [PubMed]
- Aoshima, H. Potentiation and inhibition of ionotropic neurotransmitter receptors expressed in Xenopus oocyte by linoleic acid and its hydroperoxide. J. Neurochem. 1996, 66, 1300–1305. [Google Scholar] [CrossRef] [PubMed]
- Kotze, A.C.; Hunt, P.W.; Skuce, P.; von Samson-Himmelstjerna, G.; Martin, R.J.; Sager, H.; Krücken, J.; Hodgkinson, J.; Lespine, A.; Jex, A.R.; et al. Recent advances in candidate-gene and whole-genome approaches to the discovery of anthelmintic resistance markers and the description of drug/receptor interactions. Int. J. Parasitol. Drugs Drug Resist. 2014, 4, 164–184. [Google Scholar] [CrossRef] [PubMed]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panda, S.K.; Das, R.; Mai, A.H.; De Borggraeve, W.M.; Luyten, W. Nematicidal Activity of Holigarna caustica (Dennst.) Oken Fruit Is Due to Linoleic Acid. Biomolecules 2020, 10, 1043. https://doi.org/10.3390/biom10071043
Panda SK, Das R, Mai AH, De Borggraeve WM, Luyten W. Nematicidal Activity of Holigarna caustica (Dennst.) Oken Fruit Is Due to Linoleic Acid. Biomolecules. 2020; 10(7):1043. https://doi.org/10.3390/biom10071043
Chicago/Turabian StylePanda, Sujogya Kumar, Raju Das, Anh Hung Mai, Wim M. De Borggraeve, and Walter Luyten. 2020. "Nematicidal Activity of Holigarna caustica (Dennst.) Oken Fruit Is Due to Linoleic Acid" Biomolecules 10, no. 7: 1043. https://doi.org/10.3390/biom10071043
APA StylePanda, S. K., Das, R., Mai, A. H., De Borggraeve, W. M., & Luyten, W. (2020). Nematicidal Activity of Holigarna caustica (Dennst.) Oken Fruit Is Due to Linoleic Acid. Biomolecules, 10(7), 1043. https://doi.org/10.3390/biom10071043