Chemical Composition and Immunomodulatory Activity of Hypericum perforatum Essential Oils †

 , ,
, , 
 and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Materials
2.3. Essential Oil Extraction
2.4. Gas Chromatography–Mass Spectrometry (GC-MS) Analysis
2.5. Isolation of Human Neutrophils
2.6. Ca2+ Mobilization Assay
2.7. Chemotaxis Assay
2.8. ROS Production Assay
2.9. Kinase Kd Determination
2.10. Human Neutrophil Elastase (HNE) Inhibition Assay
2.11. Cytotoxicity Assay
2.12. Molecular Modeling
3. Results and Discussion
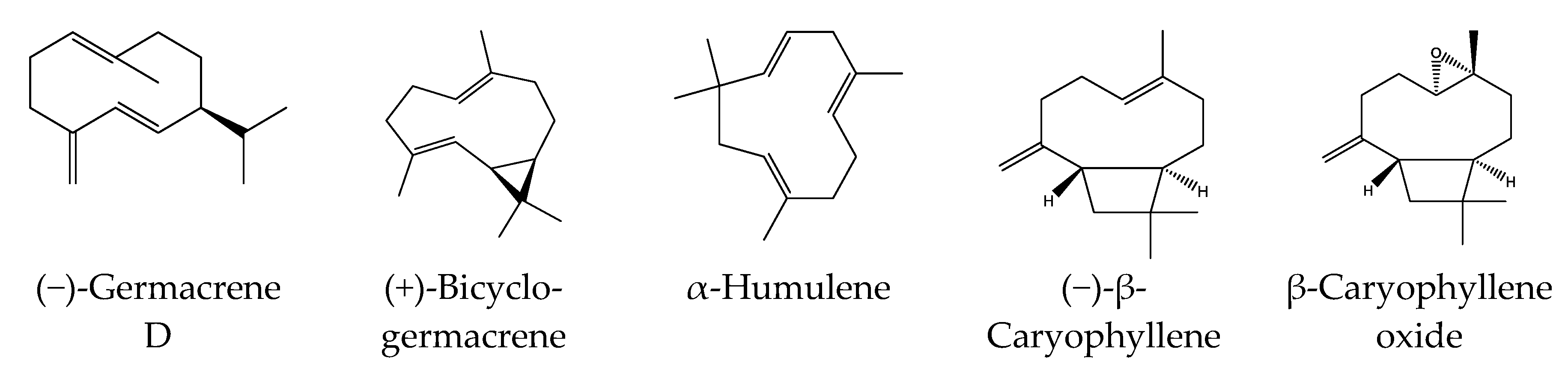
3.1. Essential Oil Composition
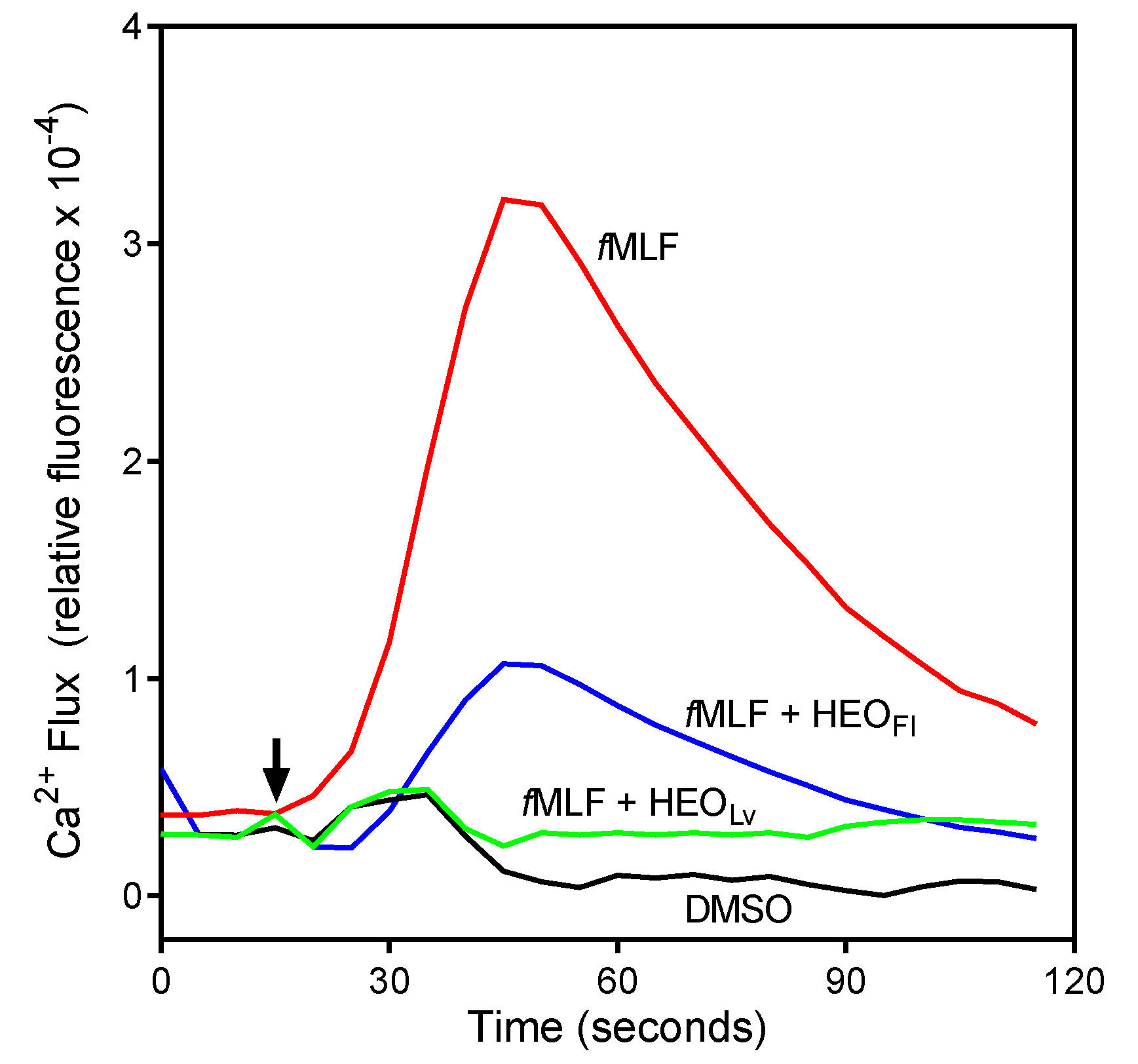
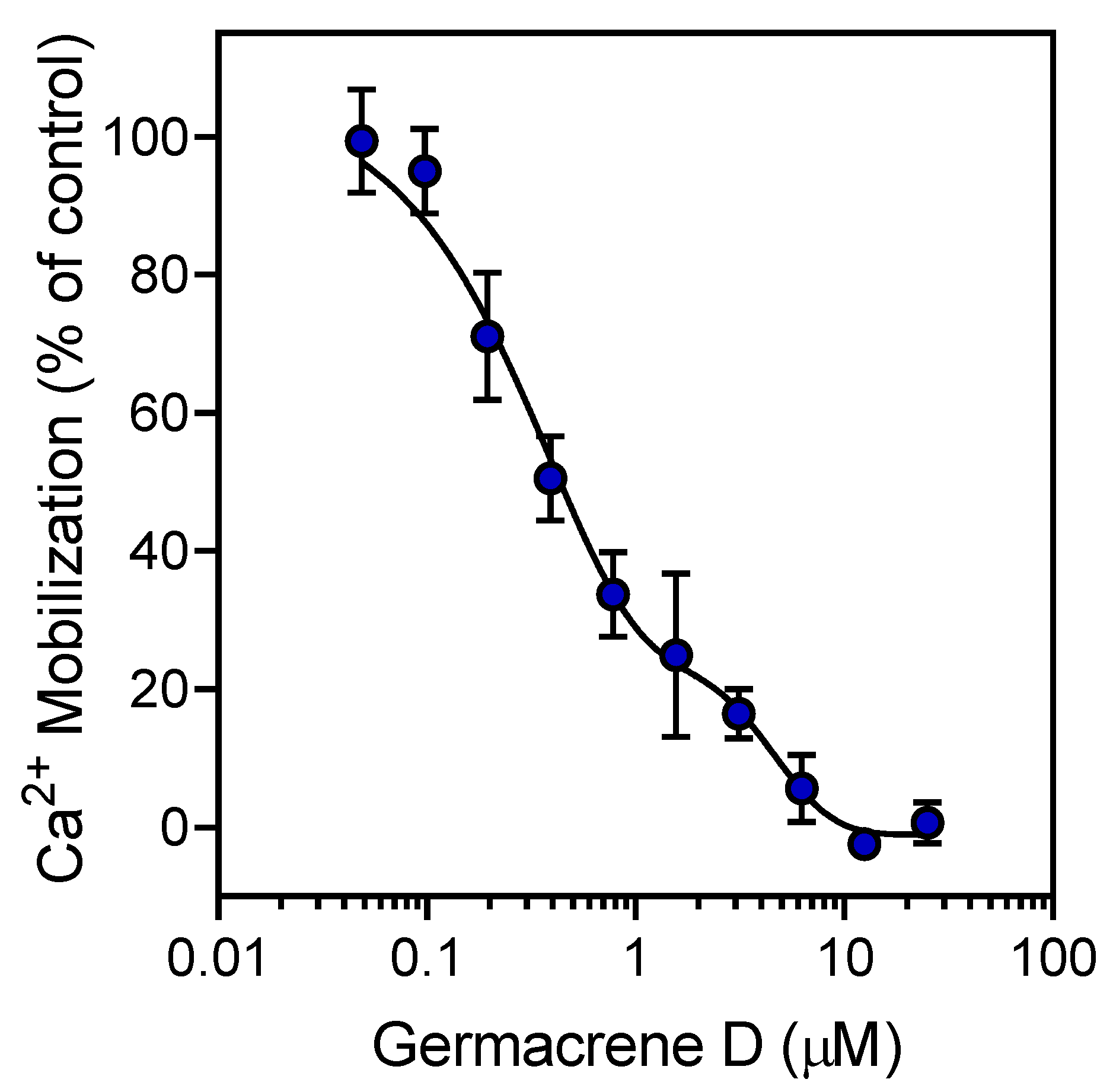
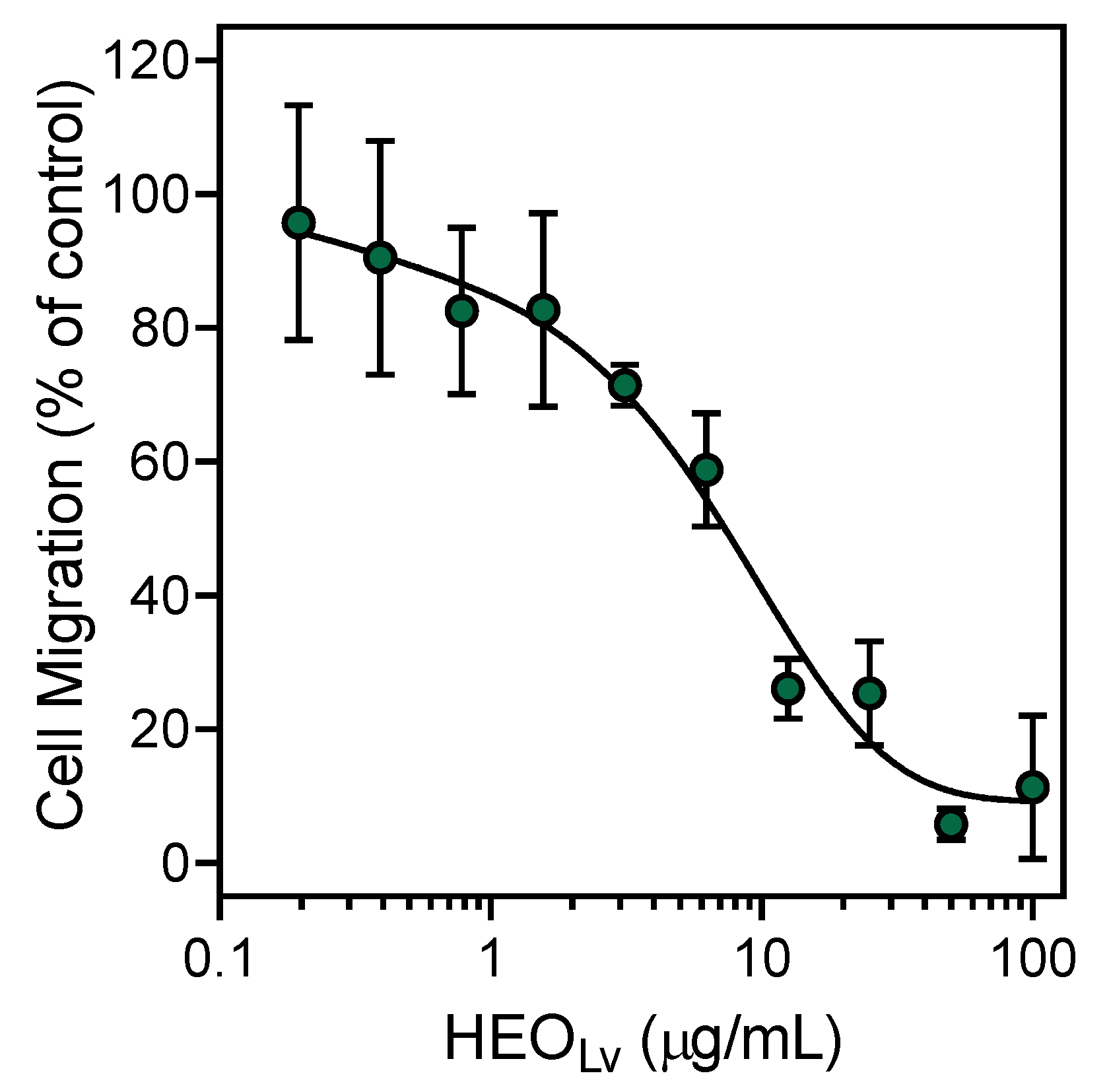
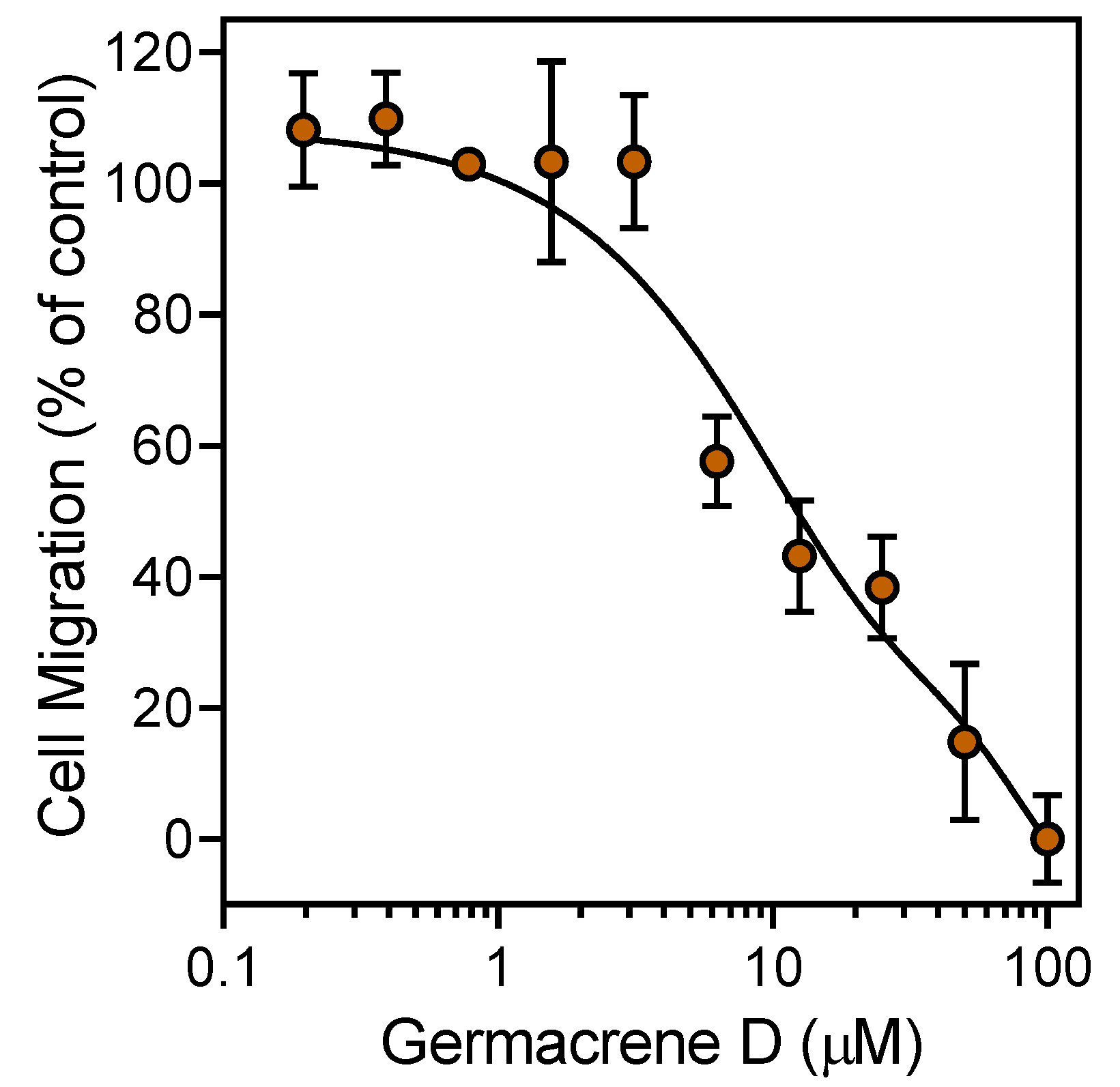
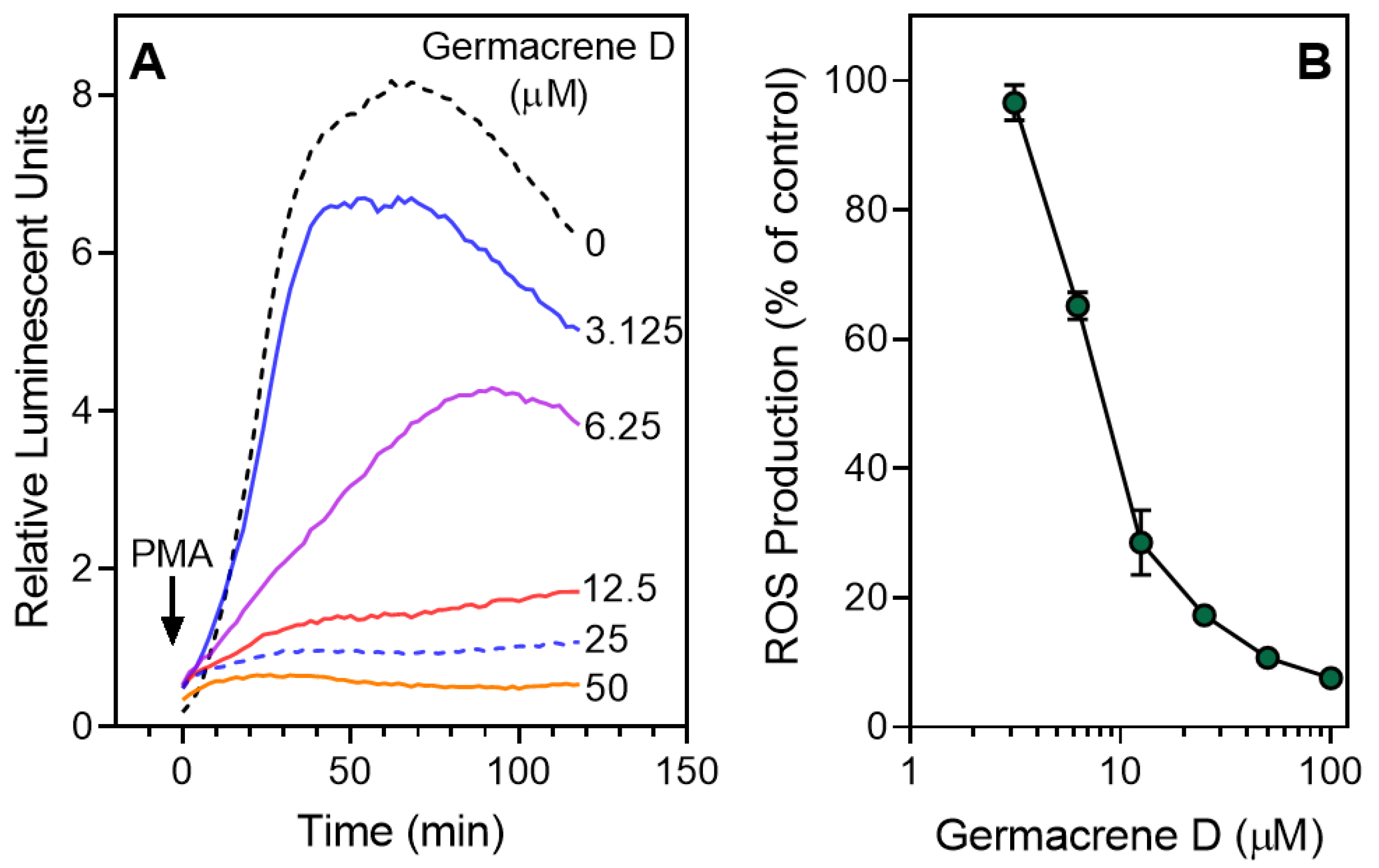
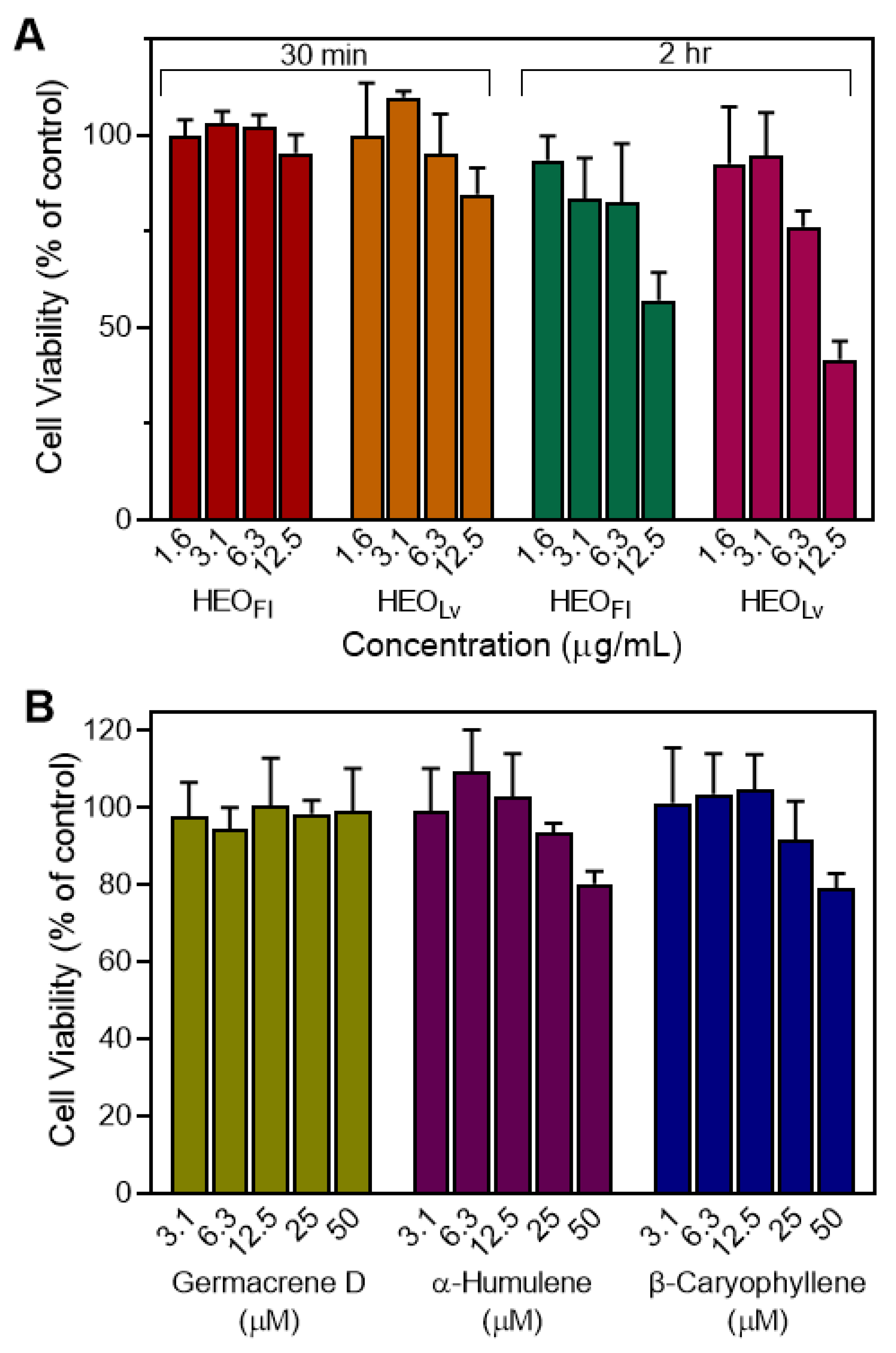
3.2. Effect of the Essential Oils and Their Components on Neutrophil Functional Responses
3.3. Identification of Potential Protein Targets for Selected Sesquiterpenes
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zhang, R.; Ji, Y.; Zhang, X.; Kennelly, E.J.; Long, C. Ethnopharmacology of Hypericum species in China: A comprehensive review on ethnobotany, phytochemistry and pharmacology. J. Ethnopharmacol. 2020, 254, 112686. [Google Scholar] [CrossRef]
- Velingkar, V.S.; Gupta, G.L.; Hegde, N.B. A current update on phytochemistry, pharmacology and herb-drug interactions of Hypericum perforatum. Phytochem. Rev. 2017, 16, 725–744. [Google Scholar] [CrossRef]
- Marrelli, M.; Statti, G.; Conforti, F.; Menichini, F. New potential pharmaceutical applications of Hypericum Species. Mini-Rev. Med. Chem. 2016, 16, 710–720. [Google Scholar] [CrossRef]
- Galeotti, N. Hypericum perforatum (St John’s wort) beyond depression: A therapeutic perspective for pain conditions. J. Ethnopharmacol. 2017, 200, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Dogan, S.; Gokalsin, B.; Senkardes, I.; Dogan, A.; Sesal, N.C. Anti-quorum sensing and anti-biofilm activities of Hypericum perforatum extracts against Pseudomonas aeruginosa. J. Ethnopharmacol. 2019, 235, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, U.; Kaya, H.; Turan, M.; Bir, F.; Sahin, B. Investigation the effect of Hypericum perforatum on corneal alkali burns. Cutan. Ocul. Toxicol. 2019, 38, 356–359. [Google Scholar] [CrossRef] [PubMed]
- Wise, K.; Selby-Pham, S.; Bennett, L.; Selby-Pham, J. Pharmacokinetic properties of phytochemicals in Hypericum perforatum influence efficacy of regulating oxidative stress. Phytomedicine 2019, 59, 152763. [Google Scholar] [CrossRef] [PubMed]
- Kandilarov, I.K.; Zlatanova, H.I.; Georgieva-Kotetarova, M.T.; Kostadinova, I.I.; Katsarova, M.N.; Dimitrova, S.Z.; Lukanov, L.K.; Sadakov, F. Antidepressant effect and recognition memory improvement of two novel plant extract combinations—Antistress I and anti-stress II on rats subjected to a model of mild chronic stress. Folia Med. 2018, 60, 110–116. [Google Scholar] [CrossRef]
- Crockett, S.L. Essential Oil and Volatile Components of the Genus Hypericum (Hypericaceae). Nat. Prod. Commun. 2010, 5, 1493–1506. [Google Scholar] [CrossRef]
- Hostettmann, K.; Wolfender, J.-L. St. John’s Wort and its active principles in depression and anxiety. In Phytochemistry; Müller, W.E., Ed.; Springer Science & Business Media: Basel, Switzerland, 2005; pp. 5–20. [Google Scholar]
- Belwal, T.; Devkota, H.P.; Singh, M.K.; Sharma, R.; Upadhayay, S.; Joshi, C.; Pande, V. St. John’s Wort (Hypericum perforatum). In Nonvitamin and Nonmineral Nutritional Supplements; Academic Press: London, UK, 2019; pp. 415–432. [Google Scholar]
- de Cassia da Silveira, E.S.R.; Andrade, L.N.; Dos Reis Barreto de Oliveira, R.; de Sousa, D.P. A review on anti-inflammatory activity of phenylpropanoids found in essential oils. Molecules 2014, 19, 1459–1480. [Google Scholar] [CrossRef]
- Perry, N.B.; Anderson, R.E.; Brennan, N.J.; Douglas, M.H.; Heaney, A.J.; McGimpsey, J.A.; Smallfield, B.M. Essential oils from dalmatian sage (Salvia officinalis L.): Variations among individuals, plant parts, seasons, and sites. J. Agric. Food Chem. 1999, 47, 2048–2054. [Google Scholar] [CrossRef] [PubMed]
- Marcetic, M.D.; Milenkovic, M.T.; Lakusic, D.V.; Lakusic, B.S. Chemical Composition and Antimicrobial Activity of the Essential Oil and Methanol Extract of Hypericum aegypticum subsp. webbii (Spach) N. Robson. Chem. Biodivers. 2016, 13, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Zorzetto, C.; Sanchez-Mateo, C.C.; Rabanal, R.M.; Lupidi, G.; Petrelli, D.; Vitali, L.A.; Bramucci, M.; Quassinti, L.; Caprioli, G.; Papa, F.; et al. Phytochemical analysis and in vitro biological activity of three Hypericum species from the Canary Islands (Hypericum reflexum, Hypericum canariense and Hypericum grandifolium). Fitoterapia 2015, 100, 95–109. [Google Scholar] [CrossRef] [PubMed]
- Akhbari, M.; Batooli, H.; Mozdianfard, M. Comparative study of composition and biological activities of SDE prepared essential oils from flowers and fruits of two Hypericum species from central Iran. Nat. Prod. Res. 2012, 26, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Toker, Z.; Kizil, G.; Ozen, H.C.; Kizil, M.; Ertekin, S. Compositions and antimicrobial activities of the essential oils of two Hypericum species from Turkey. Fitoterapia 2006, 77, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Maggi, F.; Cecchini, C.; Cresci, A.; Coman, M.M.; Tirillini, B.; Sagratini, G.; Papa, F.; Vittori, S. Chemical composition and antimicrobial activity of the essential oils from several Hypericum taxa (Guttiferae) growing in central Italy (Appennino Umbro-Marchigiano). Chem. Biodivers. 2010, 7, 447–466. [Google Scholar] [CrossRef]
- Beutler, B. Innate immunity: An overview. Mol. Immunol. 2004, 40, 845–859. [Google Scholar] [CrossRef]
- Witko-Sarsat, V.; Rieu, P.; Descamps-Latscha, B.; Lesavre, P.; Halbwachs-Mecarelli, L. Neutrophils: Molecules, functions and pathophysiological aspects. Lab. Investig. 2000, 80, 617–653. [Google Scholar] [CrossRef]
- Fletcher, S.; Steffy, K.; Averett, D. Masked oral prodrugs of toll-like receptor 7 agonists: A new approach for the treatment of infectious disease. Curr. Opin. Investig. Drugs 2006, 7, 702–708. [Google Scholar]
- Schepetkin, I.A.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. High-throughput screening for small-molecule activators of neutrophils: Identification of novel N-formyl peptide receptor agonists. Mol. Pharmacol. 2007, 71, 1061–1074. [Google Scholar] [CrossRef]
- Reshetnikov, V.; Hahn, J.; Maueroder, C.; Czegley, C.; Munoz, L.E.; Herrmann, M.; Hoffmann, M.H.; Mokhir, A. Chemical tools for targeted amplification of reactive oxygen species in neutrophils. Front. Immunol. 2018, 9, 1827. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Kushnarenko, S.V.; Ozek, G.; Kirpotina, L.N.; Sinharoy, P.; Utegenova, G.A.; Abidkulova, K.T.; Ozek, T.; Baser, K.H.; Kovrizhina, A.R.; et al. Modulation of human neutrophil responses by the essential oils from Ferula akitschkensis and their constituents. J. Agric. Food Chem. 2016, 64, 7156–7170. [Google Scholar] [CrossRef] [PubMed]
- Ozek, G.; Schepetkin, I.A.; Utegenova, G.A.; Kirpotina, L.N.; Andrei, S.R.; Ozek, T.; Baser, K.H.C.; Abidkulova, K.T.; Kushnarenko, S.V.; Khlebnikov, A.I.; et al. Chemical composition and phagocyte immunomodulatory activity of Ferula iliensis essential oils. J. Leukoc. Biol. 2017, 101, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Kushnarenko, S.V.; Ozek, G.; Kirpotina, L.N.; Utegenova, G.A.; Kotukhov, Y.A.; Danilova, A.N.; Ozek, T.; Baser, K.H.; Quinn, M.T. Inhibition of human neutrophil responses by the essential oil of Artemisia kotuchovii and its constituents. J. Agric. Food Chem. 2015, 63, 4999–5007. [Google Scholar] [CrossRef] [PubMed]
- Ozek, G.; Ishmuratova, M.; Tabanca, N.; Radwan, M.M.; Goger, F.; Ozek, T.; Wedge, D.E.; Becnel, J.J.; Cutler, S.J.; Can Baser, K.H. One-step multiple component isolation from the oil of Crinitaria tatarica (Less.) Sojak by preparative capillary gas chromatography with characterization by spectroscopic and spectrometric techniques and evaluation of biological activity. J. Sep. Sci. 2012, 35, 650–660. [Google Scholar] [CrossRef]
- Karaman, M.W.; Herrgard, S.; Treiber, D.K.; Gallant, P.; Atteridge, C.E.; Campbell, B.T.; Chan, K.W.; Ciceri, P.; Davis, M.I.; Edeen, P.T.; et al. A quantitative analysis of kinase inhibitor selectivity. Nat. Biotechnol. 2008, 26, 127–132. [Google Scholar] [CrossRef]
- Giovannoni, M.P.; Cantini, N.; Crocetti, L.; Guerrini, G.; Iacovone, A.; Schepetkin, I.A.; Vergelli, C.; Khlebnikov, A.I.; Quinn, M.T. Further modifications of 1H-pyrrolo[2,3-b]pyridine derivatives as inhibitors of human neutrophil elastase. Drug Dev. Res. 2019, 80, 617–628. [Google Scholar] [CrossRef]
- Liu, X.; Ouyang, S.; Yu, B.; Liu, Y.; Huang, K.; Gong, J.; Zheng, S.; Li, Z.; Li, H.; Jiang, H. PharmMapper server: A web server for potential drug target identification using pharmacophore mapping approach. Nucleic Acids Res. 2010, 38, W609–W614. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Rancic, A.; Sokovic, M.; Vukojevic, J.; Simic, A.; Marin, P.; Duletic-Lausevic, S.; Djokovic, D. Chemical composition and antimicrobial activities of essential oils of Myrrhis odorata (L.) Scop, Hypericum perforatum L. and Helichrysum arenarium (L.) Moench. J. Essent. Oil Res. 2005, 17, 341–345. [Google Scholar] [CrossRef]
- Ghasemi Pirbalouti, A.; Fatahi-Vanani, M.; Craker, L.; Shirmardi, H. Chemical composition and bioactivity of essential oils of Hypericum helianthemoides, Hypericum perforatum and Hypericum scabrum. Pharm. Biol. 2014, 52, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Đorđević, A.S. Chemical composition of Hypericum perforatum L. essential oil. Adv. Technol. 2015, 4, 64–68. [Google Scholar]
- Yüce, E. Analysis of the Essential Oils of two Hypericum species (H. lanuginosum var. lanuginosum Lam. and H. perforatum L.) from Turkey. Hacettepe J. Biol. Chem. 2016, 44, 29–34. [Google Scholar] [CrossRef]
- Parchin, R.A.; Ebadollahi, A. Biological activities of Hypericum perforatum L. essential oil against red flour beetle, Tribolium castaneum (Herbst)(Coleoptera: Tenebrionidae). J. Entomol. 2016, 13, 91–97. [Google Scholar]
- Morshedloo, M.R.; Nabizadeh, M.; Akramian, M.; Yazdani, D. Characterization of the volatile oil compositions from Hypericum perforatum L. shoot cultures in different basal media. Azarian J. Agric. 2017, 4, 7–11. [Google Scholar]
- Saleh, B. Volatile constituents of three Hypericum (Hypericaceae) species using GC-MS analysis. Int. J. Pharm. Life Sci. 2019, 10, 6396–6405. [Google Scholar]
- Zeliou, K.; Koui, E.M.; Papaioannou, C.; Koulakiotis, N.S.; Iatrou, G.; Tsarbopoulos, A.; Papasotiropoulos, V.; Lamari, F.N. Metabolomic fingerprinting and genetic discrimination of four Hypericum taxa from Greece. Phytochemistry 2020, 174, 112290. [Google Scholar] [CrossRef]
- Bergonzi, M.C.; Bilia, A.R.; Gallori, S.; Guerrini, D.; Vincieri, F.F. Variability in the content of the constituents of Hypericum perforatum L. and some commercial extracts. Drug Dev. Ind. Pharm. 2001, 27, 491–497. [Google Scholar] [CrossRef]
- Cirak, C.; Bertoli, A.; Pistelli, L.; Seyis, F. Essential oil composition and variability of Hypericum perforatum from wild populations of northern Turkey. Pharm. Biol. 2010, 48, 906–914. [Google Scholar] [CrossRef]
- Morshedloo, M.R.; Ebadi, A.; Maggi, F.; Fattahi, R.; Yazdani, D.; Jafari, M. Chemical characterization of the essential oil compositions from Iranian populations of Hypericum perforatum L. Ind. Crop. Prod. 2015, 76, 565–573. [Google Scholar] [CrossRef]
- Morshedloo, M.R.; Ebadi, A.; Fatahi Moghaddam, M.R.; Yazdani, D. Essential oil composition, total phenol compounds and antioxidant activity of Hypericum perforatum L. extract collected from North of Iran. J. Med. Plants 2012, 1, 218–226. [Google Scholar]
- Smelcerovic, A.; Spiteller, M.; Ligon, A.P.; Smelcerovic, Z.; Raabe, N. Essential oil composition of Hypericum L. species from Southeastern Serbia and their chemotaxonomy. Biochem. Syst. Ecol. 2007, 35, 99–113. [Google Scholar] [CrossRef]
- Radusiene, J.; Judzentiene, A.; Bernotiene, G. Essential oil composition and variability of Hypericum perforatum L. growing in Lithuania. Biochem. Syst. Ecol. 2005, 33, 113–124. [Google Scholar] [CrossRef]
- Mockute, D.; Bernotiene, G.; Judzentiene, A. The essential oils with dominant germacrene D of Hypericum perforatum L. growing wild in Lithuania. J. Essent. Oil Res. 2008, 20, 128–131. [Google Scholar] [CrossRef]
- Baser, K.H.C.; Ozek, T.; Nuriddinov, H.R.; Demirci, A.B. Essential oils of two Hypericum species from Uzbekistan. Chem. Nat. Compd. 2002, 38, 54–57. [Google Scholar] [CrossRef]
- Fraternale, D.; Flamini, G.; Ascrizzi, R. In Vitro anticollagenase and antielastase activities of essential oil of Helichrysum italicum subsp. italicum (Roth) G. Don. J. Med. Food 2019, 22, 1041–1046. [Google Scholar] [CrossRef]
- Laothaweerungsawat, N.; Sirithunyalug, J.; Chaiyana, W. Chemical compositions and anti-skin-ageing activities of Origanum vulgare L. essential oil from tropical and Mediterranean region. Molecules 2020, 25, 1101. [Google Scholar] [CrossRef]
- Fachini-Queiroz, F.C.; Kummer, R.; Estevao-Silva, C.F.; Carvalho, M.D.; Cunha, J.M.; Grespan, R.; Bersani-Amado, C.A.; Cuman, R.K. Effects of thymol and carvacrol, constituents of Thymus vulgaris L. essential oil, on the inflammatory response. Evid. Based Complementary Altern. Med. 2012, 2012, 657026. [Google Scholar]
- Danielli, L.J.; de Souza, T.J.T.; Maciel, A.J.; Ferrao, M.F.; Fuentefria, A.M.; Apel, M.A. Influence of monoterpenes in biological activities of Nectandra megapotamica (Spreng.) Mez essential oils. Biomolecules 2019, 9, 112. [Google Scholar] [CrossRef]
- Basile, A.; Senatore, F.; Gargano, R.; Sorbo, S.; Del Pezzo, M.; Lavitola, A.; Ritieni, A.; Bruno, M.; Spatuzzi, D.; Rigano, D.; et al. Antibacterial and antioxidant activities in Sideritis italica (Miller) Greuter et Burdet essential oils. J. Ethnopharmacol. 2006, 107, 240–248. [Google Scholar] [CrossRef]
- Cosentino, M.; Luini, A.; Bombelli, R.; Corasaniti, M.T.; Bagetta, G.; Marino, F. The essential oil of bergamot stimulates reactive oxygen species production in human polymorphonuclear leukocytes. Phytother. Res. 2014, 28, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Stranden, M.; Liblikas, I.; Konig, W.A.; Almaas, T.J.; Borg-Karlson, A.K.; Mustaparta, H. (-)-Germacrene D receptor neurones in three species of heliothine moths: Structure-activity relationships. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2003, 189, 563–577. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.W.; Huang, C.D.; Zheng, H.H.; Zhao, N.; Feng, X.L.; Ma, S.J.; Zhang, A.L.; Zhang, Q. Meroterpene-Like α-glucosidase inhibitors based on biomimetic reactions starting from β-caryophyllene. Molecules 2020, 25, 260. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J.Z.; Xie, X.Q.; Altmann, K.H.; Karsak, M.; Zimmer, A. β-caryophyllene is a dietary cannabinoid. Proc. Natl. Acad. Sci. USA 2008, 105, 9099–9104. [Google Scholar] [CrossRef] [PubMed]
- D’Ascola, A.; Irrera, N.; Ettari, R.; Bitto, A.; Pallio, G.; Mannino, F.; Atteritano, M.; Campo, G.M.; Minutoli, L.; Arcoraci, V.; et al. Exploiting curcumin synergy with natural products using quantitative analysis of dose-effect relationships in an experimental in vitro model of osteoarthritis. Front. Pharmacol. 2019, 10, 1347. [Google Scholar] [CrossRef]
- Tian, X.; Liu, H.; Xiang, F.; Xu, L.; Dong, Z. β-Caryophyllene protects against ischemic stroke by promoting polarization of microglia toward M2 phenotype via the TLR4 pathway. Life Sci. 2019, 237, 116915. [Google Scholar] [CrossRef]
- Alberti, T.B.; Barbosa, W.L.R.; Vieira, J.L.F.; Raposo, N.R.B.; Dutra, R.C. (-)-β-Caryophyllene, a CB2 receptor-selective phytocannabinoid, suppresses motor paralysis and neuroinflammation in a murine model of multiple sclerosis. Int. J. Mol. Sci. 2017, 18, 691. [Google Scholar] [CrossRef]
- Chang, H.J.; Kim, J.M.; Lee, J.C.; Kim, W.K.; Chun, H.S. Protective Effect of β-Caryophyllene, a Natural bicyclic sesquiterpene, against cerebral ischemic injury. J. Med. Food 2013, 16, 471–480. [Google Scholar] [CrossRef]
- Machado, K.D.C.; Islam, M.T.; Ali, E.S.; Rouf, R.; Uddin, S.J.; Dev, S.; Shilpi, J.A.; Shill, M.C.; Reza, H.M.; Das, A.K.; et al. A systematic review on the neuroprotective perspectives of β-caryophyllene. Phytother. Res. 2018, 32, 2376–2388. [Google Scholar] [CrossRef]
- Amiel, E.; Ofir, R.; Dudai, N.; Soloway, E.; Rabinsky, T.; Rachmilevitch, S. β-Caryophyllene, a compound isolated from the biblical balm of Gilead (Commiphora gileadensis), is a selective apoptosis inducer for tumor cell lines. Evid. Based Complementary Altern. Med. 2012, 2012, 872394. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, C.F.; Dai, X.L.; Ao, Y.; Li, Y.M. Inhibitory effect of trans-caryophyllene (TC) on leukocyte-endothelial attachment. Toxicol. Appl. Pharm. 2017, 329, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Andrade-Silva, M.; Correa, L.B.; Candea, A.L.P.; Cavalher-Machado, S.C.; Barbosa, H.S.; Rosas, E.C.; Henriques, M.G. The cannabinoid 2 receptor agonist β-caryophyllene modulates the inflammatory reaction induced by Mycobacterium bovis BCG by inhibiting neutrophil migration. Inflamm. Res. 2016, 65, 869–879. [Google Scholar] [CrossRef]
- Chung, H.M.; Wang, W.H.; Hwang, T.L.; Chen, J.J.; Fang, L.S.; Wen, Z.H.; Wang, Y.B.; Wu, Y.C.; Sung, P.J. Rumphellols A and B, new caryophyllene sesquiterpenoids from a Formosan gorgonian coral, Rumphella antipathies. Int. J. Mol. Sci. 2014, 15, 15679–15688. [Google Scholar] [CrossRef] [PubMed]
- Legault, J.; Dahl, W.; Debiton, E.; Pichette, A.; Madelmont, J.C. Antitumor activity of balsam fir oil: Production of reactive oxygen species induced by α-humulene as possible mechanism of action. Planta Med. 2003, 69, 402–407. [Google Scholar] [PubMed]
- Ouattara, Z.A.; Boti, J.B.; Ahibo, C.A.; Bekro, Y.A.; Casanova, J.; Tomi, F.; Bighelli, A. Composition and chemical variability of Ivoirian Polyalthia oliveri leaf oil. Chem. Biodivers. 2016, 13, 293–298. [Google Scholar] [CrossRef]
- Rogerio, A.P.; Andrade, E.L.; Leite, D.F.P.; Figueiredo, C.P.; Calixto, J.B. Preventive and therapeutic anti-inflammatory properties of the sesquiterpene α-humulene in experimental airways allergic inflammation. Br. J. Pharmacol. 2009, 158, 1074–1087. [Google Scholar] [CrossRef]
- Medeiros, R.; Passos, G.F.; Vitor, C.E.; Koepp, J.; Mazzuco, T.L.; Pianowski, L.F.; Campos, M.M.; Calixto, J.B. Effect of two active compounds obtained from the essential oil of Cordia verbenacea on the acute inflammatory responses elicited by LPS in the rat paw. Br. J. Pharmacol. 2007, 151, 618–627. [Google Scholar] [CrossRef]
- Oh, M.S.; Yang, J.Y.; Kim, M.G.; Lee, H.S. Acaricidal activities of β-caryophyllene oxide and structural analogues derived from Psidium cattleianum oil against house dust mites. Pest Manag. Sci. 2014, 70, 757–762. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Mysliveckova, Z.; Szotakova, B.; Spicakova, A.; Lnenickova, K.; Ambroz, M.; Kubicek, V.; Krasulova, K.; Anzenbacher, P.; Skalova, L. The inhibitory effects of β-caryophyllene, β-caryophyllene oxide and α-humulene on the activities of the main drug-metabolizing enzymes in rat and human liver in vitro. Chem. Biol. Interact. 2017, 278, 123–128. [Google Scholar] [CrossRef]
- Schipilliti, L.; Bonaccorsi, I.; Sciarrone, D.; Dugo, L.; Mondello, L.; Dugo, G. Determination of petitgrain oils landmark parameters by using gas chromatography-combustion-isotope ratio mass spectrometry and enantioselective multidimensional gas chromatography. Anal. Bioanal. Chem. 2013, 405, 679–690. [Google Scholar] [CrossRef]
- Saroglou, V.; Marin, P.D.; Rancic, A.; Veljic, M.; Skaltsa, H. Composition and antimicrobial activity of the essential oil of six Hypericum species from Serbia. Biochem. Syst. Ecol. 2007, 35, 146–152. [Google Scholar] [CrossRef]
- Stranden, M.; Borg-Karlson, A.K.; Mustaparta, H. Receptor neuron discrimination of the germacrene D enantiomers in the moth Helicoverpa armigera. Chem. Senses 2002, 27, 143–152. [Google Scholar] [CrossRef]
- Nishimura, K.; Shinoda, N.; Hirose, Y. A New Sesquiterpene, bicyclogermacrene. Tetrahedron Lett. 1969, 10, 3097–3100. [Google Scholar] [CrossRef]
- Konig, W.A.; Rieck, A.; Saritas, Y.; Hardt, I.H.; Kubeczka, K.H. Sesquiterpene hydrocarbons in the essential oil of Meum athamanticum. Phytochemistry 1996, 42, 461–464. [Google Scholar] [CrossRef]
- Dixit, N.; Kim, M.H.; Rossaint, J.; Yamayoshi, I.; Zarbock, A.; Simon, S.I. Leukocyte function antigen-1, kindlin-3, and calcium flux orchestrate neutrophil recruitment during inflammation. J. Immunol. 2012, 189, 5954–5964. [Google Scholar] [CrossRef]
- Qian, F.; Deng, J.; Wang, G.; Ye, R.D.; Christman, J.W. Pivotal Role of mitogen-activated protein kinase-activated protein kinase 2 in inflammatory pulmonary diseases. Curr. Protein Pept. Sci. 2016, 17, 332–342. [Google Scholar] [CrossRef]
- Sun, L.; Wu, Q.; Nie, Y.; Cheng, N.; Wang, R.; Wang, G.; Zhang, D.; He, H.; Ye, R.D.; Qian, F. A Role for MK2 in enhancing neutrophil-derived ROS production and aggravating liver ischemia/reperfusion injury. Front. Immunol. 2018, 9, 2610. [Google Scholar] [CrossRef]
- Bialopiotrowicz, E.; Gorniak, P.; Noyszewska-Kania, M.; Pula, B.; Makuch-Lasica, H.; Nowak, G.; Bluszcz, A.; Szydlowski, M.; Jablonska, E.; Piechna, K.; et al. Microenvironment-induced PIM kinases promote CXCR4-triggered mTOR pathway required for chronic lymphocytic leukaemia cell migration. J. Cell. Mol. Med. 2018, 22, 3548–3559. [Google Scholar] [CrossRef]
- Woolley, J.F.; Naughton, R.; Stanicka, J.; Gough, D.R.; Bhatt, L.; Dickinson, B.C.; Chang, C.J.; Cotter, T.G. H2O2 production downstream of FLT3 is mediated by p22phox in the endoplasmic reticulum and is required for STAT5 signalling. PLoS ONE 2012, 7, e34050. [Google Scholar] [CrossRef]
- Giner, B.; Lafuente, C.; Lapena, D.; Errazquin, D.; Lomba, L. QSAR study for predicting the ecotoxicity of NADES towards Aliivibrio fischeri. Exploring the use of mixing rules. Ecotoxicol. Environ. Saf. 2020, 191, 110004. [Google Scholar] [CrossRef]
- Kishk, S.M.; McLean, K.J.; Sood, S.; Smith, D.; Evans, J.W.D.; Helal, M.A.; Gomaa, M.S.; Salama, I.; Mostafa, S.M.; de Carvalho, L.P.S.; et al. Design and synthesis of imidazole and triazole pyrazoles as Mycobacterium tuberculosis CYP121A1 inhibitors. ChemistryOpen 2019, 8, 995–1011. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Major Compound (%) | Ref. |
|---|---|---|
| Greece | α-pinene (7.5); (E)-β-caryophyllene (10.3); germacrene D (5.5); β-selinene (14.7); α-selinene (14.6) | [39] |
| Iran | decane (27.7); menthol (8.9); methyl decanoate (4.6); β-elemene (4.6) | [37] |
| eudesma-4(15),7-dien-1β-ol (8.1–7.5); thymol (7.0–7.2); 1,4-trans-1,7-trans-acorenone (5.2–5.5) | [37] | |
| decane (59.6); dodecane (12.9); ethyl cyclohexane (6.8); 5-methyl nonane (4.7); 3-methyl nonane (4.3) | [36] | |
| 2,6-dimethyl-heptane (6.2–36.1); α-pinene (5.6–26.0); δ-cadinene (0.0–22.6); γ-cadinene (0.0–16.9) | [42] | |
| α-pinene (12.5); β-pinene (8.3); (E)-β-ocimene (4.4); 2-methyl decane (4.0); undecane (7.0); germacrene D (6.9); α-selinene (4.2) | [33] | |
| α-pinene (21.9); nonane (9.8); n-octane (9.1); dodecanol (6.8) | [43] | |
| Serbia | germacrene D (18.6); β-caryophyllene (11.2); 2-methyl octane (9.5); α-pinene (6.5); bicyclogermacrene (5.0); (E)-β-ocimene (4.6) | [34] |
| Syria | β-selinenol (18.1); elemol (12.8); β-elemene (10.7) | [38] |
| Turkey | β-selinene (19.4); bicyclogermacrene (15.3); tetradecene (8.2); α-amorphene (8.1) | [35] |
| No | RRI | Compound | Fl | Lv | No | RRI | Compound | Fl | Lv |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 965 | 3-methyl nonane | 0.6 | 48 | 1687 | α-humulene | 1.2 | ||
| 2 | 1032 | α-pinene | 2.2 | 4.9 | 49 | 1690 | cryptone | 0.9 | |
| 3 | 1035 | α-thujene | 1.4 | 50 | 1693 | β-acoradiene | 0.4 | ||
| 4 | 1048 | MBOL | 1.5 | 51 | 1700 | limonen-4-ol | 3.2 | ||
| 5 | 1065 | 2-methyl decane | 0.2 | 52 | 1704 | γ-muurolene | 1.4 | ||
| 6 | 1100 | undecane | 0.2 | 53 | 1706 | α-terpineol | 6.1 | ||
| 7 | 1118 | β-pinene | 0.9 | 4.6 | 54 | 1708 | ledene | 0.3 | |
| 8 | 1132 | sabinene | 5.6 | 55 | 1726 | germacrene D | 25.7 | ||
| 9 | 1174 | myrcene | 2.1 | 56 | 1740 | α-muurolene | 0.8 | ||
| 10 | 1176 | α-phellandrene | t | 57 | 1743 | α-cadinene | 0.3 | ||
| 11 | 1188 | α-terpinene | 0.5 | 58 | 1743 | eremophilene | 1.7 | ||
| 12 | 1203 | limonene | 0.5 | 59 | 1755 | bicyclogermacrene | 2.5 | ||
| 13 | 1218 | β-phellandrene | 1 | 60 | 1758 | (E,E)-α-farnesene | 0.7 | ||
| 14 | 1225 | (Z)-3-hexenal | 0.2 | 61 | 1773 | δ-cadinene | 2.5 | ||
| 15 | 1246 | (Z)-β-ocimene | 0.5 | 62 | 1776 | γ-cadinene | 1.0 | ||
| 16 | 1255 | γ-terpinene | 1.1 | 63 | 1799 | cubenene | 0.1 | ||
| 17 | 1266 | (E)-β-ocimene | 4.2 | 64 | 1804 | myrtenol | 2.7 | ||
| 18 | 1280 | p-cymene | 0.5 | 1.2 | 65 | 1815 | 2,6-dimethyl-3(E),5(Z),7-octatriene-2-ol | 0.1 | |
| 19 | 1290 | terpinolene | 0.3 | 66 | 1830 | 1.5 | |||
| 20 | 1466 | α-cubebene | 0.1 | 67 | 1853 | cis-calamenene | 0.2 | ||
| 21 | 1475 | acetic acid | 1.3 | 68 | 1857 | geraniol | 1.4 | t | |
| 22 | 1493 | α-ylangene | 0.1 | 69 | 1864 | p-cymen-8-ol | 2.9 | 0.1 | |
| 23 | 1495 | bicycloelemene | 0.1 | 70 | 1900 | epi-cubebol | t | ||
| 24 | 1497 | α-copaene | 0.2 | 71 | 1945 | 1,5-epoxy-salvial(4)14-ene | t | ||
| 25 | 1506 | decanal | t | 72 | 1953 | palustrol | 0.4 | ||
| 26 | 1525 | α-funebrene | t | 73 | 1973 | dodecanol | 1.2 | ||
| 27 | 1535 | β-bourbonene | 0.2 | 74 | 2006 | MDCB * | 9.8 | ||
| 28 | 1536 | italicene | t | 75 | 2008 | caryophyllene oxide | 1.2 | ||
| 29 | 1544 | α-gurjunene | 0.5 | 76 | 2050 | (E)-nerolidol | 0.8 | ||
| 30 | 1549 | β-cubebene | t | 77 | 2057 | ledol | 1.2 | ||
| 31 | 1553 | linalool | 1.3 | 0.2 | 78 | 2069 | germacrene D-4β-ol | t | |
| 32 | 1562 | octanol | 1.1 | 79 | 2080 | cubenol | 0.2 | ||
| 33 | 1571 | TMEOL | 0.9 | t | 80 | 2088 | 1-epi-cubenol | 0.2 | |
| 34 | 1586 | pinocarvone | 0.9 | 81 | 2093 | junenol | 0.3 | ||
| 35 | 1587 | β-funebrene | 1.1 | 82 | 2098 | globulol | 0.3 | ||
| 36 | 1589 | β-ylangene | 0.3 | 83 | 2099 | trans-ascaridol glycol * | 4.6 | ||
| 37 | 1602 | MHDO | t | 84 | 2104 | viridiflorol | 0.2 | ||
| 38 | 1608 | β-pinone | 1.3 | 85 | 2115 | HMCH | 3.4 | ||
| 39 | 1608 | β-copaene | 0.4 | 86 | 2144 | spathulenol | 1.6 | 1.9 | |
| 40 | 1611 | terpinen-4-ol | 7.4 | 2.6 | 87 | 2179 | tetradecanol | 0.7 | |
| 41 | 1612 | β-caryophyllene | t | 9.5 | 88 | 2184 | cis-p-menth-3-en-1,2-diol | 9.1 | |
| 42 | 1613 | β-cedrene | 0.4 | 89 | 2187 | T-cadinol | 0.6 | ||
| 43 | 1638 | CMEOL | 1.1 | 90 | 2209 | T-muurolol | 0.8 | ||
| 44 | 1661 | alloaromadendrene | 0.6 | 91 | 2219 | torreyol | 0.2 | ||
| 45 | 1668 | (Z)-β-farnesene | 0.6 | 92 | 2255 | α-cadinol | 1.5 | 1.9 | |
| 46 | 1670 | trans-pinocarveol | 0.6 | 93 | 2260 | alismol | 0.1 | ||
| 47 | 1683 | trans-verbenol | 1.6 | 94 | 2329 | trans-sobrerol * | 1.7 |
| Total (%) | 71.3 | 97.2 |
|---|---|---|
| Monoterpene hydrocarbons (%) | 3.6 | 27.9 |
| Oxygenated monoterpenes (%) | 49.2 | 3.0 |
| Sesquiterpene hydrocarbons (%) | 0 | 52.9 |
| Oxygenated sesquiterpenes (%) | 3.1 | 10.3 |
| Miscellaneous compounds (%) | 15.4 | 3.1 |
| Essential Oil or Pure Compound | Composition (%) | Ca2+ Flux a | Chemotaxis b | ROS Production c | |
|---|---|---|---|---|---|
| IC50 (μg/mL) | |||||
| HEOFl | 11.3 ± 1.8 | 5.7 ± 1.8 | 9.5 ± 0.9 | ||
| HEOLv | 2.3 ± 0.4 | 5.2 ± 1.1 | 1.2 ± 0.4 | ||
| HEOFl | HEOLv | IC50 (μM) | |||
| Caryophyllene oxide | 0 | 1.2 | N.A. d | N.A. d | N.A. d |
| p-Cymen-8-ol | 2.9 | 0.1 | N.A. d | N.A. d | N.A. d |
| p-Cymene | 0.5 | 1.2 | N.A. d | N.A. d | N.A. d |
| Myrcene | 0 | 2.1 | N.A. d | N.A. d | N.A. d |
| α-Terpinene | 0 | 0.5 | N.A. d | N.A. d | N.A. d |
| Limonene | 0 | 0.5 | N.A. d | N.A. d | N.A. d |
| Myrtenol | 2.7 | 0 | N.A. d | N.A. | N.A. |
| (E/Z)-β-Ocimene | 0 | 4.7 | N.A. d | N.A. d | N.A. d |
| α-Pinene | 2.2 | 4.9 | N.A. d | N.A. d | N.A. d |
| (1S)-(-)-β-Pinene | 0.9 | 4.6 | N.A. d | 22.7 ± 2.6 d | N.A. d |
| (±)-Sabinene | 0 | 5.6 | N.A. d | 37.4 ± 4.3 d | N.A. d |
| (−)-Terpinen-4-ol | 7.4 | 2.6 | N.A. d | N.A. | N.A. |
| γ-Terpinene | 0 | 1.1 | N.A. | 32.5 ± 4.6 d | N.A. |
| α-Terpineol | 6.1 | 0 | N.A. d | N.A. | N.A. |
| Terpinolene | 0 | 0.3 | N.A. d | N.A. d | N.A. d |
| (-)-Linalool | 1.3 | 0.2 | N.A. d | N.A. d | N.A. d |
| MHDO | < 0.1 | 0 | 8.2 ± 2.5 d | 3.6 ± 0.5 d | 2.8 ± 0.4 d |
| Germacrene D | 0 | 25.7 | 0.51 ± 0.08 | 5.4 ± 2.3 | 9.9 ± 1.9 |
| Geraniol | 1.4 | < 0.1 | N.A. | N.A. | 50.1 ± 3.2 |
| α-Humulene | 0 | 1.2 | 0.31 ± 0.06 | 12.0 ± 3.4 | 2.2 ± 0.8 |
| β-Caryophyllene | < 0.1 | 9.5 | 0.33 ± 0.02 | 17.6 ± 5.7 | 2.6 ± 0.9 |
| Rank | PDB ID | Target Name | Fit Score | Rank | PDB ID | Target Name | Fit Score |
|---|---|---|---|---|---|---|---|
| (−)-β-Caryophyllene | α-Humulene | ||||||
| 1 | 1XDD | Integrin α-L | 0.998 | 1 | 2P3G | MAPKAPK2 | 0.989 |
| 2 | 1REU | BMP-2 | 0.987 | 2 | 3BMP | BMP-2 | 0.987 |
| 3 | 2P3G | MAPKAPK2 | 0.981 | 3 | 1J96 | AKR1C2 | 0.981 |
| 4 | 1J96 | AKR1C2 | 0.981 | 4 | 1SHJ | Caspase-7 | 0.961 |
| 5 | 1L6L | ApoA-II | 0.957 | 5 | 2O65 | PIM1 | 0.925 |
| 6 | 1P49 | Steroid sulfatase | 0.954 | 6 | 1P49 | Steroid sulfatase | 0.908 |
| 7 | 1E7E | Serum albumin | 0.919 | 7 | 2PIN | THRB | 0.885 |
| 8 | 2PG2 | KIF11 | 0.919 | 8 | 1E7A | Serum albumin | 0.880 |
| (−)-Germacrene D | Bicyclogermacrene | ||||||
| 1 | 1E7A | Serum albumin | 0.999 | 1 | 1REU | BMP-2 | 0.998 |
| 2 | 1SHJ | Caspase-7 | 0.990 | 2 | 2O65 | PIM1 | 0.993 |
| 3 | 1P49 | Steroid sulfatase | 0.977 | 3 | 1E7E | Serum albumin | 0.991 |
| 4 | 2O65 | PIM1 | 0.969 | 4 | 1J96 | AKR1C2 | 0.990 |
| 5 | 2PG2 | KIF11 | 0.958 | 5 | 2P3G | MAPKAPK2 | 0.988 |
| 6 | 1L6L | ApoA-II | 0.951 | 6 | 2PG2 | KIF11 | 0.920 |
| 7 | 2P3G | MAPKAPK2 | 0.948 | 7 | 1L6L | ApoA-II | 0.913 |
| 8 | 1J96 | AKR1C2 | 0.924 | 8 | 1SHJ | Caspase-7 | 0.913 |
| Property | Germacrene D | Bicyclo- germacrene | α-Humulene | β-Caryophyllene | β-Caryophyllene Oxide |
|---|---|---|---|---|---|
| Formula | C15H24 | C15H24 | C15H24 | C15H24 | C15H24O |
| M.W. | 204.35 | 204.35 | 204.35 | 204.35 | 220.35 |
| Heavy atoms | 15 | 15 | 15 | 15 | 16 |
| Fraction Csp3 | 0.60 | 0.73 | 0.60 | 0.73 | 0.87 |
| Rotatable bonds | 1 | 0 | 0 | 0 | 0 |
| H-bond acceptors | 0 | 0 | 0 | 0 | 1 |
| H-bond donors | 0 | 0 | 0 | 0 | 0 |
| MR | 70.68 | 68.78 | 70.42 | 68.78 | 68.27 |
| tPSA | 0.00 | 0.00 | 0.00 | 0.00 | 12.53 |
| Log P | 4.30 | 4.15 | 4.26 | 4.24 | 3.68 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schepetkin, I.A.; Özek, G.; Özek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Chemical Composition and Immunomodulatory Activity of Hypericum perforatum Essential Oils. Biomolecules 2020, 10, 916. https://doi.org/10.3390/biom10060916
Schepetkin IA, Özek G, Özek T, Kirpotina LN, Khlebnikov AI, Quinn MT. Chemical Composition and Immunomodulatory Activity of Hypericum perforatum Essential Oils. Biomolecules. 2020; 10(6):916. https://doi.org/10.3390/biom10060916
Chicago/Turabian StyleSchepetkin, Igor A., Gulmira Özek, Temel Özek, Liliya N. Kirpotina, Andrei I. Khlebnikov, and Mark T. Quinn. 2020. "Chemical Composition and Immunomodulatory Activity of Hypericum perforatum Essential Oils" Biomolecules 10, no. 6: 916. https://doi.org/10.3390/biom10060916
APA StyleSchepetkin, I. A., Özek, G., Özek, T., Kirpotina, L. N., Khlebnikov, A. I., & Quinn, M. T. (2020). Chemical Composition and Immunomodulatory Activity of Hypericum perforatum Essential Oils. Biomolecules, 10(6), 916. https://doi.org/10.3390/biom10060916